
Abstract
A thermally selected strain of rainbow trout has been established by selective breeding since 1966 in Miyazaki, Japan. In the present study, we compared the critical thermal maxima (CTMs), the temperatures at which organisms reach a predefined sublethal endpoint and lose their equilibrium, between a thermally selected and two normal (Donaldson) strains of rainbow trout. The CTM of one normal strain from Nikko (Nikko strain) acclimated to 20 °C (29.7 °C) was significantly lower than those of the thermally selected strain (30.0 °C) and the other Donaldson strain from Aomori (29.9 °C) (P < 0.05). The F1 generations, F1T and F1N, were produced by crossing thermally selected strain females with Nikko strain males and Nikko strain females with thermally selected strain males, respectively. No significant difference was observed in the CTM between F1T [30.1 ± 0.15 °C (n = 30)] and F1N [30.1 ± 0.16 °C (n = 30)] (P > 0.05) for fish acclimated to 20 °C, suggesting that the F1 offspring inherited the thermal tolerance trait from one thermally selected strain parent irrespective of whether it was the male or female. F2 offspring of F1T or F1N also showed the thermal tolerance trait. The coefficients of variation for CTM were also compared among all the datasets obtained in the present study and their values for F1 hybrids were lower than those of the parental generation of the Nikko strain (P < 0.05). In contrast, the coefficients of variation of F2s were the same as those of their parental generation. Furthermore, the thermally selected strain and Nikko strain as a reference family provide a F2 generation for segregating phenotypes, which is required for in-depth genetic analysis of the thermally selected rainbow trout strain.
Similar content being viewed by others


Introduction
Salmonids are globally important, cold water aquaculture species. The selective breeding of salmonids has been performed to improve the traits associated with growth and disease resistance (Henryon et al. 2005; Kause et al. 2007; Leeds et al. 2010; Sae-Lim et al. 2012; Janhunen et al. 2012). Undesirably high water temperatures have been recorded during summer months in cold water salmonid-inhabiting streams due to global warming and/or human activity-induced habitat alteration (Schindler 2001). Moreover, salmonids often experience a range of water temperatures that approach or exceed their lethal limits for at least a couple of days within a year (Lund et al. 2002). Rainbow trout Oncorhynchus mykiss is an important salmonid, but has limits for aquaculture in terms of temperature when cultured in temperate regions. High-temperature-resistant strains are desired for game fishing (Currie et al. 1998), and Miyazaki Prefectural Fisheries Research Institute Kobayashi Branch, Miyazaki Prefecture, Japan, has been breeding a rainbow trout strain endowed with thermal tolerance since 1966 in order to expand aquaculture areas for this species, which could be farmed at relatively high temperatures. Our previous studies demonstrated that the upper 50% lethal temperature (LT50) and hatching rate for embryos at high temperatures of an established, thermally selected strain were significantly higher than those of the normal strains (Ineno et al. 2005). Furthermore, death temperature (DT) and LT50 of fry of the selected strain acclimated to 20 °C were again significantly higher than those of the normal strains. Biochemical experiments also demonstrated that while the cytosolic proteins were released into the circulation system of the normal strain after exposure to high temperatures, the thermally selected strain showed no change after the same high temperature treatment (Itoi et al. 2001). Furthermore, we examined the differences in the gene expression pattern of embryos between the thermally selected and normal strains following exposure to a temperature of 22 °C. One of the genes upregulated in the thermally selected strain encoded cytochrome c oxidase subunit II in mitochondria (Ikeguchi et al. 2006). The transcripts accumulated in the embryos of the thermally selected strain as maternal ribonucleic acid in the unfertilized eggs. In addition, a comprehensive transcriptome analysis was carried out to analyze the gene expression of tissues of juveniles after hatching, in the thermally selected and normal strains, before and after high temperature exposure at 26 °C by using a next-generation sequencer. As a result, the levels of transcripts encoding heat shock proteins (Hsp) in the thermally selected strain were found to be markedly higher than those of the normal strain and the levels of HSP70b mRNA were 2000 times more than those of the normal strain regardless of exposure to high temperatures (Tang et al. 2012, 2016). However, Hsp70 expression at the protein level after exposure to the high temperature of 28 °C in F2 fry generated by crossbreeding the thermally selected and normal strains was only apparent in high temperature-tolerant individuals, and not in less-tolerant counterparts (Ojima et al. 2012).
In the present study, thermal tolerance was compared between the two Donaldson strains. Then pedigrees of the thermally selected were determined by crossbreeding with a low thermal tolerant strain, and the offspring examined for their thermal tolerance trait by using the critical thermal maximum (CTM) that indicates the temperature at which animals reach a predicted sublethal endpoint and lose their equilibrium.
Materials and methods
Materials
The parental strains included a thermally selected strain of rainbow trout, which was generated by breeding in high temperature treatments since 1966 in Miyazaki Prefectural Fisheries Research Institute Kobayashi Branch, Miyazaki Prefecture, Japan. The parental strains also included two pedigrees of a normal (Donaldson) strain, which have been bred at the Freshwater Fisheries Research Division of the National Research Institute of Aquaculture, Nikko Branch, Fisheries Research Agency, Tochigi Prefecture, Japan (Nikko strain), and the Freshwater Fisheries Research Institute, Aomori Prefectural Industrial Technology Research Center, Aomori Prefecture (Aomori strain). The Nikko strain was preserved as pure lines and bred in cold water for a long time. In contrast, the Aomori strain was obtained by selective breeding during aquaculture in the coastal region around the Tsugaru Channel in the northern region of Honshu Island. These two normal strains were compared for thermal tolerance by the following method, then a strain with a much lower thermal tolerance, Nikko, was selected as the parental normal strain to cross with the thermally selected strain.
Construction of F1 and F2 generations
Hybrid F1 generations, F1T and F1N, were produced by crossing thermally selected strain females with Nikko strain males and Nikko strain females with thermally selected strain males, respectively. These were produced from 50 diallel lots.
F1T was produced as follows: milt from ten thermally selected strain males were collected at Kobayashi Branch and transported to the Nikko Branch by using an ice-cold airtight container on the following day. Subsequently, 25 diallel lots were produced by crossing five thermally selected strain lots with active sperm with five Nikko strain lots. Twenty-five diallel lots of fertilized eggs thus obtained were incubated until the eyed egg stage in hatching tanks of the Nikko Branch at 10 °C, and subsequently transported to the Kobayashi Branch for breeding. Each F1 family was reared in a separate tank at 17 °C.
The opposite cross, F1N, was carried out using ten Nikko strain males and five thermally selected strain females following the above procedures. Twenty-five diallel lots of fertilized eggs were incubated at 13 °C until the eyed egg stage in hatching tanks of the Kobayashi Branch, and each diallel lot was bred individually at 17 °C at the branch. Then fish were fed daily with a commercial diet.
Two F2 pedigrees, F2T and F2N, were produced by a full-sib cross for F1T and F1N, respectively, and bred individually at the Kobayashi Branch at 17 °C.
Determination of thermal tolerance
Fifty juveniles each of the thermally selected Nikko and Aomori strains were transported to the Marine Ecology Research Institute Central Laboratory (MERI), Chiba Prefecture, Japan. The fish were fed daily with commercial diet at 2% of total body weight in 200-L Artemia hatching tanks. Subsequently CTM was determined in three 38-L circulation-type cylindrical tanks in which water temperature was regulated according to the temperature equipment installed outside at MERI. Fish were acclimated to 15 or 20 °C for more than 2 weeks. Then two individuals of the thermally selected strain and three individuals of the Nikko or Aomori strains were introduced into one tank and subjected to thermal experiments by raising the water temperature at a rate of 5 °C/h. The temperature at which fish lost their equilibrium was defined as the CTM. Usually the determination of the CTM took about 180 and 120 min when fish were acclimated to 15 and 20 °C, respectively (data not shown). At first, juveniles were determined for their CTMs after acclimation at 15 °C. This experiment was repeated five or six times using different individuals (Table 1) after allowing all fish to acclimate to 15 °C. Second, the remaining juveniles that had not been used in the first experiments after acclimation at 15 °C were acclimated to 20 °C for more than 2 weeks by raising the water temperature at a rate of 1 °C/day, and subjected to the same CTM experiments as the fish acclimated to 15 °C. This experiment was performed four or five time using different individuals acclimated to 20 °C.
Twenty juveniles from two groups of F1T and ten juveniles from four groups of F1N were transported to MERI and acclimated to 20 °C for more than 2 weeks under the same conditions as those described above. CTM was determined to compare the potential of tolerance to high temperatures between F1T and F1N by the same method as described above.
Sixty juveniles each from the two F2 pedigrees were transported to MERI, acclimated to 20 °C for more than 2 weeks, and CTM determined to compare the potential high-temperature tolerance between F2T and F2N by the same method as that described above.
Statistical analysis
Data were expressed as mean ± SD. Statistical significance was determined using the one-sided Student’s t-test. Differences were considered to be significant at P < 0.05. The coefficient of variation is a useful statistic data for comparing the degree of variation from one data series to another; it represents the ratio of the SD to the mean, and was calculated as follows: 100 × SD/mean (%).
Results
CTM of the thermally selected, Nikko and Aomori strains
The three rainbow trout strains, the thermally selected, Nikko and Aomori strains, were acclimated to either 15 or 20 °C; there were 18–28 individuals of each strain. Fish acclimated to 15 °C showed very similar values of CTM at 29.1–29.2 °C irrespective of the strain, with no significant difference (P > 0.05) (Table 1).
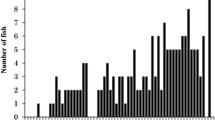
When acclimated to 20 °C, the three strains showed CTMs at 30.0–30.1 °C, which were significantly higher than CTMs of fish acclimated to 15 °C (P < 0.05) (Table 1; Fig. 1). In addition to this, the CTM of the thermally selected and Aomori strains acclimated to 20 °C was significantly higher than that of the Nikko strain acclimated to 20 °C (P < 0.05). The difference in the CTM between the thermally selected and Aomori strains acclimated to 20 °C was not significant at P > 0.05.

Critical thermal maximum (CTM) for the thermally selected strain (a, d), the Nikko strain (b, e) and the Aomori strain (c, f) rainbow trout Oncorhynchus mykiss juveniles acclimated to 15 °C (a–c) and 20 °C (d–f)
Coefficients of variation for CTMs of the thermally selected, Nikko and Aomori strains were 0.60–0.83% with no apparent difference among the three strains (Table 1). Coefficients of variation for CTMs of the three strains acclimated to 20 °C were 1.0–1.6% (Table 1), which were much higher than those of fish acclimated to 15 °C (0.6–0.8%). Figure 1 clearly shows that all the three strains acclimated to 20 °C had individual CTM values in the temperature range wider than that of fish acclimated to 15 °C. This was especially true for the Nikko strain acclimated to 20 °C, of which three individuals had CTMs of 28.5–29.2 °C. There were also two individuals of the thermally selected strain acclimated to 20 °C with a CTMs of 29.1 and 29.3 °C, respectively.
CTM of F1 and F2 generations
Since the Aomori strain was considered to have acquired thermal tolerance, probably due to the selective breeding during aquaculture in the coastal region around the Tsugaru Channel, F1 and F2 generations were produced only between the thermally selected and Nikko strains.
F1T from the thermally selected females crossed with the Nikko strain males and F1N from an opposite combination between the two strains were acclimated only to 20 °C and subjected to CTM analyses, because these parents showed differences in thermal tolerance only when acclimated at 20 °C, but not at 15 °C. Both F1 generations showed CTM at 30.1 °C and no significant difference was observed between the two F1 generations (P > 0.05) (Table 1; Fig. 2). However, CTMs of the F1 generations were significantly higher than that of the Nikko strain acclimated to 20 °C (P < 0.05) and comparable to that of the thermally selected strain acclimated to 20 °C.

CTM of rainbow trout O. mykiss juveniles acclimated to 20 °C for F1T hybrids produced by crossing thermally selected strain females with Nikko strain males (a) and F1N hybrids produced by crossing Nikko stain females with thermally selected strain males (b)
The distribution of CTM for both F1 generations, F1T and F1N, was compact compared to parent strains (Fig. 2) and gave coefficients of variation of 0.50–0.53%, which were much smaller than any coefficients of variation from parent strains acclimated either to 15 or 20 °C (0.60–1.58%) (Table 1).
F2T and F2N hybrids in the F2 generation showed CTM at 30.1 and 30.2 °C, respectively (Table 1, Fig. 3) with no significant difference between the two hybrid groups (P > 0.05). Coefficients of variation were 0.88 and 1.36% for F2T and F2N, respectively (Table 1), which were both comparable to those of their parental strains.

CTM of rainbow trout O. mykiss juveniles acclimated to 20 °C for F2T (a) and F2N (b) of the full-sib progeny of F1T and F1N, respectively. F1T and F1N were produced by crossing thermally selected strain females with Nikko strain males and by crossing Nikko strain females with thermally selected strain males, respectively
Discussion
The thermally selected strain of rainbow trout established in Miyazaki Prefecture, Japan, by selective breeding since 1966 was examined for tolerance to high temperatures, and this was compared with the temperature tolerance of normal strains. The normal strains originated from the Donaldson strain, which is one of the established pedigrees of rainbow trout produced for economic efficiency, growth and reproduction by the University of Washington (Donaldson and Olson 1955). One of the normal strains in the present study was bred at Nikko, Tochigi Prefecture, and the other in Aomori Prefecture, Japan.
CTM was used in the present study as an index of tolerance to high temperatures. In this method, fish are subjected to a constant dynamic temperature change until the temperature reaches a predefined sublethal endpoint at which the locomotory movements become disorganized and the animal loses its ability to escape from these conditions, which might ultimately lead to its death (Becker and Genoway 1979; Beitinger et al. 2000; Cox 1974).
According to our previous results (Ineno et al. 2005), DT is more sensitive than CTM. However, DT experiments are laborious and take a long time to obtain reproducible results. On the other hand, CTM experiments are much more convenient and many people employ this method, although these experiments are less sensitive than those using DT. Hartman and Porto (2014) also gave a similar opinion when referring to these results of ours. In the present CTM experiments there are significant differences between the Nikko strain and both the Aomori and thermally selected strain, although the differences are very small. The CTM experiments employed in the present study enabled us to perform many experiments using many individuals to ascertain the thermal tolerance trait for the F1 and F2 generations inherited from the thermally selected strain.
CTMs of the normal strains, including that bred in Miyazaki Prefecture, Japan, have been reported to be significantly lower than that of the thermally selected strain, although the difference was almost within 1 °C (Ineno et al. 2005). Based on the assumption that normal strains inhabiting northern Japan have a lower tolerance to high temperatures compared with the thermally selected strain, two pedigrees of the normal strains, the Nikko and Aomori strains, were subjected to CTM analyses in the present study.
Three strains, the thermally selected, Nikko and Aomori strains, showed CTMs of 29.1–29.2 °C and no significant difference was observed among the them when they were acclimated to 15 °C. When they were acclimated to 20 °C, however, CTMs were increased to 29.7–30.1 °C, which were significantly higher than those of fish acclimated to 15 °C (P < 0.05). Currie et al. (1998) reported a highly positive correlation between CTM and acclimation temperature for rainbow trout acclimated to 10, 15, and 20 °C at the rate of 0.18 °C CTM/1 °C increase in acclimation temperature. Among the three strains in the present study, the differences in CTMs were 0.9, 0.6, and 0.7 °C between fish acclimated to 15 and 20 °C in the thermally selected, Nikko and Aomori strains, respectively. Thus, the increased CTM in response to the increase of acclimation temperature for the thermally selected strain coincided well with the above-mentioned results. However, the increases in CTMs of the Nikko and Aomori strains, i.e., 0.12 and 0.14 °C, respectively, were lower than that reported by Currie et al. (1998), i.e., 0.18 °C. The CTM of the thermally selected strain acclimated to 20 °C (30.0 ± 0.30 °C) was higher than that reported by Currie et al. (1998), where CTMs of fish acclimated to 15 and 20 °C were 29.1 ± 0.27 °C and 29.8 ± 0.36 °C, respectively. While fish acclimated at 15 °C in the present study did not show any significant difference in CTM, those acclimated at 20 °C had significant differences between the thermally selected and Nikko strains and between the Aomori and Nikko strains (Table 1). Such effects of acclimation temperature on CTM have been observed in our previous experiments using the thermally selected, Donaldson and Nagano strain fry (Ineno et al. 2005). It seems that acclimation at 20 °C, but not at 15 °C, for more than 2 weeks is stressful for the Nikko strain, but not the Aomori and thermally selected strains. That is probably why we could observe significant differences between the Nikko strain and both the Aomori and thermally selected strain.
As described above, the thermally selected and Aomori strains showed significantly higher CTMs than the Nikko strain when fish were acclimated to 20 °C (P < 0.05), but not when fish were acclimated to 15 °C.
This thermally selected strain of rainbow trout has been preserved for reproductive breeding, although the FAO/UNEP (1981) emphasizes the importance of maintaining breeding populations with an effective size of at least 50 for short-term fitness and of at least 500 for long-term survival. However, maintaining breeding populations involves a great deal of effort and cost. Many strains of both plants and animals with beneficial traits have been established and their genetic resources have been preserved in seed vaults, for plants, and as frozen gametes or embryos for mammals (Börner 2006; Boettcher et al. 2005). Fish sperm have been successfully cryopreserved for many species (Leung and Jamieson 1991), including rainbow trout (Lahnsteiner 2000). Preservation of our thermally selected rainbow trout strain seems necessary if its sperm propagates the trait of high temperature tolerance.
CTMs of F1 hybrids showed no difference between the pedigrees F1T and F1N that were produced by crossbreeding between thermally selected strain females and Nikko strain males and between thermally selected strain males and Nikko strain females, respectively. CTMs of F1 showed no significant difference from that of the thermally selected strain, and were significant higher than that of the Nikko strain. These results indicate no apparent relation of a high temperature tolerance trait according to sex. Kudo et al. (2001) estimated the heritability for tolerance to high water temperature by using the average time of death of individuals exposed to high temperature (25 °C) in 120 diallel lots of progeny, which showed that high temperature tolerance of rainbow trout was influenced by a paternal trait. Anttila et al. (2013) also employed an experimental design involving half-sibling families from the same male, but with different females, in Atlantic salmon. The obtained data did not show a maternal effect, suggesting that there was a significant genetic component underlying CTM. In our present study, F1 was produced by crossing the thermally selected and the normal Nikko strains. F1 hybrids either from the thermally selected strain females crossed with the Nikko strain males or from an opposite combination between the two strains showed as high values of CTM as the thermally selected strain (Table 1). These results suggest that the thermal tolerance trait of our strain is inherited by offspring from the thermally selected strain irrespective of whether it is inherited from males and females.
While various factors, including genetic inheritance (Elliott 1991) and acclimation temperature (Becker and Genoway 1979; Elliott 1991), are considered to affect tolerance to high temperatures of fish, even the Nikko strain is more resistant to high temperatures than the other strains reported previously (e.g., North Carolina strain, Nimbus strain and PFRC strain acclimated to 15 °C, and Arizona strain acclimated 20 °C) (Galbreath et al. 2006; Myrick and Cech Jr 2005; Chen et al. 2015). Another salmonid, sockeye salmon Oncorhynchus nerka, showed local intraspecific differences in tolerance to high temperatures during their spawning migration (Lee et al. 2003; Eliason et al. 2011). The red band trout, a local population of rainbow trout in North America, has been reported to live at high temperatures in desert areas where temperature reaches 29 °C (Pickering 1992). These reports suggest that salmonid easily acquire a trait of tolerance to high temperatures during their adaptation to environments. It is thus assumed that the Aomori strain has acquired thermal tolerance during the breeding process in seawater where it grows faster than fish reared in freshwater.
The phenotypic variability is measured alternatively by the coefficient of variation, which is calculated by 100\(\sqrt {V_{\text{P}} }\)/µ, where µ is the mean and VP is the phenotypic variance (Hill and Mulder 2010). \(\sqrt {V_{\text{P}} }\) is expressed by the SD; therefore, the phenotypic variability is demonstrated as the formula of 100 × SD/µ. Kause et al. (2007) reported the coefficient of variation as 23% for body weight for 29,666 individuals of farmed rainbow trout (mean value, 1,158 g), after the growing season, in seawater from April to the subsequent winter at Åland Islands, Finland. Therefore, the coefficients of variation of CTMs for the three strains in the present study (0.50–1.58%) (Table 1) were markedly low, although the phenotypes compared (weight gain vs. thermal tolerance) are very different. Interestingly, the coefficients of variation for the CTMs of fish acclimated to 20 °C (0.97–1.58%) were higher than those for fish acclimated to 15 °C (0.60–0.83%), especially for the Nikko strain acclimated to 20 °C (0.65%).
Heterosis is often observed in F1 generations, where superior traits and phenotypic variations exceeding those of their parents are shown (Fujio et al. 1995). CTMs of F1 hybrids produced in the present study were higher than those of the Nikko strain, but not higher than those of the thermally selected strain. However, F1T and F1N showed coefficients of variation of 0.50 and 0.53% at an acclimation temperature of 20 °C, respectively, which were much lower than those of their parents, i.e., 1.0 and 1.6%, for the thermally selected and Nikko strains at the same acclimation temperature, respectively. These results suggest that heterotic effects on the thermal tolerance also occurred in the F1 generation in the present study. In this regard, clonal strains of ayu Plecoglossus altivelis have been reported to reduce phenotypic variation (Tsujimura and Taniguchi 1996). Although the coefficient of variation in total length of hatched larvae for normal diploid ayu was 4.3%, that of cloned ayu decreased to 1.5% (Tsujimura and Taniguchi 1996). Mice also showed a decreased coefficient of variation in weight gain during feeding between 21 and 42 days after selective breeding for 18 generations (approximately 40%) (Ibáñez-Escriche et al. 2008). The lower coefficients of variation of CTMs observed in the present study demonstrated that rainbow trout including the thermally selected strain have been selected for the long term or according to their high suitability for high-temperature treatments.
The F2 generation showed no difference in CTM between the two pedigrees, F2T and F2N, which were produced by segregating full-sib families of F1T and F1N, respectively. CTMs were also comparable between F2 and F1 hybrids, which showed no difference from those of the thermally selected strain, but were significantly higher than those of the Nikko strain (P < 0.05). It is generally accepted that phenotypic variation is enlarged in the F2 segregate generation, which was partly demonstrated for thermal tolerance in our experiments by the increased coefficients of variation, i.e., 0.88 and 1.36%, for F2T and F2N, respectively, compared with 0.50–0.53% for the F1 generation as described above.
The present results demonstrated that the Nikko strain was suitable for creating a reference family for selective breeding for tolerance to high temperatures in rainbow trout. In addition, CTM, employed in this study, was promising as a phenotypic index for tolerance to high temperatures. Previous researchers (Jackson et al. 1998; Danzmann et al. 1999; Perry et al. 2001) identified a phenotype having tolerance to high temperatures by measuring time for fish to lose their equilibrium around the incipient lethal temperature. The phenotype was determined by “effective time” or “resistance time” to death, which was over 10 min at temperatures in a range between the incipient lethal and ultimate lethal temperatures (Elliott 1981). Furthermore, Perry et al. (2001) reevaluated the effective time with regards the cumulative exposure value in degree (degree Celsius) minutes by multiplying the difference between acclimation and exposure temperatures (degrees Celsius) by time required to reach the loss of equilibrium (minutes) at all acclimation temperatures, considering that the effective time is affected by the rate of increase from acclimation to exposure temperatures. While CTM and effective time are the same measures from the viewpoint that they are employed to evaluate the stress responses to sub-lethal temperatures using loss of equilibrium as a parameter, it is interesting to re-evaluate tolerance to high temperatures in the thermally selected rainbow trout established in Miyazaki Prefecture. Nevertheless, the present study proposes a simplified method to evaluate tolerance to high temperatures of rainbow trout strains and their pedigrees, which is suggested to be useful for selective breeding of this thermal tolerance trait.
Taken together, the present study suggests that the thermal tolerance trait of our strain is inherited by F1 offspring from one parent, either male or female, and that the thermally selected strain and Nikko strain as a reference family provide a F2 generation for segregating phenotypes, which is required for genetic analysis for the selective breeding of thermal tolerance.
References
Anttila K, Dhillon RS, Boulding EG, Farrell AP, Glebe BD (2013) Variation in temperature tolerance among families of Atlantic salmon (Salmo salar) is associated with hypoxia tolerance, ventricle size and myoglobin level. J Exp Biol 216:1183–1190
Becker CD, Genoway RG (1979) Evaluation of the critical thermal maximum for determining thermal tolerance of freshwater fish. Environ Biol Fish 4:245–256
Beitinger TL, Bennett WA, McCauley RW (2000) Temperature tolerances of North American freshwater fishes exposed to dynamic changes in temperature. Environ Biol Fish 58:237–275
Boettcher PJ, Stella A, Pizzi F, Gandini G (2005) The combined use of embryos and semen for cryogenic conservation of mammalian livestock genetic resources. Genet Sel Evol 37:657–675
Börner A (2006) Preservation of plant genetic resources in the biotechnology era. Biotechnol J 1:1393–1404
Chen Z, Snow M, Lawrence CS, Church AR, Narum SR, Devlin RH, Farrell AP (2015) Selection for upper thermal tolerance in rainbow trout (Oncorhynchus mykiss Walbaum). J Exp Biol 218:803–812. https://doi.org/10.1242/jeb.113993
Cox DK (1974) Effects of three heating rates on the critical thermal maximum of bluegill. In: Gibbons JW, Sharitz RR (eds) Thermal ecology Conference 730505. National Technical Information Service, Springfield, pp 158–163
Currie RJ, Bennett WA, Beitinger TL (1998) Critical thermal minima and maxima of three freshwater game-fish species. Environ Biol Fish 51:187–200
Danzmann RG, Jackson TR, Ferguson MM (1999) Epistasis in allelic expression at upper temperature tolerance QTL in rainbow trout. Aquaculture 173:45–58
Donaldson LR, Olson PR (1955) Development of rainbow trout blood stock by selective breeding. Trans Am Fish Soc 85:93–101
Eliason EJ, Clark TD, Hague MJ, Hanson LM, Gallagher ZS, Jeffries KM, Gale MK, Patterson DA, Hinch SG, Farrell AP (2011) Differences in thermal tolerance among sockeye salmon populations. Science 332:109–112. https://doi.org/10.1126/sience.1199158
Elliott JM (1981) Some aspects of thermal stress on freshwater teleosts. In: Pickering AC (ed) Stress and fish. Academic, London, pp 209–245
Elliott JM (1991) Tolerance and resistance to thermal stress in juvenile Atlantic salmon, Salmo salar. Freshwater Biol 25:61–70
FAO/UNEP (1981) Conservation of the genetic resources of fish: problems and recommendations. Report of the Expert Consultation on the genetic resources of fish. FAO Fish Tech Pap 217:43
Fujio Y, Nakajima M, Nomura G (1995) Selection response on thermal resistance of the guppy Poecilia reticulata. Fish Sci 61:731–734
Galbreath PF, Adams ND, Sherrill LW, Martin TH (2006) Thermal tolerance of diploid versus triploid rainbow trout and brook trout assessed by time to chronic lethal maximum. Environ Biol Fish 75:183–193
Hartman KJ, Porto MA (2014) Thermal performance of three rainbow trout strains at above-optimal temperatures. Trans Am Fish Soc 143:1445–1454
Henryon M, Berg P, Olesen NJ, Kjaer TE, Slierendrecht WJ, Jokumsen A, Lund I (2005) Selective breeding provides an approach to increase resistance of rainbow trout (Oncorhynchus mykiss) to the diseases, enteric redmouth disease, rainbow trout fry syndrome, and viral haemorrhagic septicaemia. Aquaculture 250:621–636. https://doi.org/10.1016/j.aquaculture.2004.12.022
Hill WG, Mulder HA (2010) Genetic analysis of environmental variation. Genet Res Camb 92:381–395. https://doi.org/10.1017/S0016672310000546
Ibáñez-Escriche N, Moreno A, Nieto B, Piquears P, Salgado C, Gutiérrez JP (2008) Genetic parameters related to environmental variability of weight traits in a selection experiment for weight gain in mice: signs of correlated canalised response. Genet Sel Evol 40:279–293
Ikeguchi K, Ineno T, Itoi S, Kondo H, Kinoshita S, Watabe S (2006) Increased levels of mitochondrial gene transcripts in the thermally selected rainbow trout (Oncorhynchus mykiss) strain during embryonic development. Mar Biotechnol 8:178–188
Ineno T, Tsuchida S, Kanda M, Watabe S (2005) Thermal tolerance of a rainbow trout Oncorhynchus mykiss strain selected by high-temperature breeding. Fish Sci 71:767–775
Itoi S, Ineno T, Kinoshita S, Hirayama Y, Nakaya M, Kakinuma M, Watabe S (2001) Analysis on serum proteins from rainbow trout Oncorhynchus mykiss exposed to high temperature. Fish Sci 67:191–193
Jackson TR, Ferguson MM, Danzmann RG, Fishback AG, Ihssen PE, O’Connell M, Crease TJ (1998) Identification of two QTL influencing upper temperature tolerance in rainbow trout (Oncorhynchus mykiss). Heredity 80:143–151
Janhunen M, Kause A, Vehvilainen H, Jarvisalo O (2012) Genetics of microenvironmental sensitivity of body weight in rainbow trout (Oncorhynchus mykiss) selected for improved growth. PLoS One 7(6):e38766. https://doi.org/10.1371/journal.pone.0038766
Kause A, Paananen T, Ritola O, Koskinen H (2007) Direct and indirect selection of visceral lipid weight, fillet weight, and fillet percentage in a rainbow trout breeding program. J Anim Sci 85:3218–3227. https://doi.org/10.2527/jas.2007-0332
Kudo H, Inoguchi N, Kijima A (2001) Estimation of heritability of tolerance to high water temperature by factorial mating system in rainbow trout (Oncorhynchus mykiss). Suisanzoshoku 49:405–411 (in Japanese)
Lahnsteiner F (2000) Semen cryopreservation in the Salmonidae and in the northern pike. Aquac Res 31:245–258
Lee CG, Farrell AP, Lotto A, MacNutt MJ, Hinch SG, Healey MC (2003) The effect of temperature on swimming performance and oxygen consumption in adult sockeye (Oncorhynchus nerka) and coho (O. kisutch) salmon stocks. J Exp Biol 206:3239–3251
Leeds TD, Silverstein JT, Weber GM, Vallejo RL, Palti Y, Rexroad CE 3rd, Evenhuis J, Hadidi S, Welch TJ, Wiens GD (2010) Response to selection for bacterial cold water disease resistance in rainbow trout. J Anim Sci 88:1936–1946. https://doi.org/10.2527/jas.2009-2538
Leung LK, Jamieson BG (1991) Live preservation of fish gametes. In: Jamieson BG (ed) Fish evolution and systematics: evidence from spermatozoa. Cambridge University Press, Cambridge, pp 245–269
Lund SG, Caissie D, Cunjak RA, Vijayan MM, Tuft BL (2002) The effects of environmental heat stress on heat shock mRNA and protein expression in Miramichi Atlantic salmon (Salmo salar) Parr. Can J Fish Aquat Sci 59:1553–1562
Myrick CA, Cech JJ Jr (2005) Effects of temperature on the growth, food consumption, and thermal tolerance of age-0 nimbus-strain steelhead. N Am J Aquac 67:324–330
Ojima N, Mekuchi M, Ineno T, Tamaki K, Kera A, Kinoshita S, Asakawa S, Watabe S (2012) Differential expression of heat-shock proteins in F2 offspring from F1 hybrids produced between thermally selected and normal rainbow trout strains. Fish Sci 78:1051–1057
Perry GML, Danzmann RG, Ferguson MM, Gibson JP (2001) Quantitative trait loci for upper thermal tolerance in outbred strains of rainbow trout (Oncorhynchus mykiss). Heredity 86:333–341
Pickering AD (1992) Rainbow trout husbandry: management of the stress response. Aquaculture 100:125–139
Sae-Lim P, Komen H, Kause A, Van Arendonk JM, Barfoot AJ, Martin KE, Parsons JE (2012) Defining desired genetic gains for rainbow trout breeding objective using analytic hierarchy process. J Anim Sci 90:1766–1776
Schindler DW (2001) The cumulative effects of climate warming and other human stresses on Canadian freshwaters in the new millennium. Can J Fish Aquat Sci 58:18–29
Tan E, Kinoshita S, Suzuki Y, Ineno T, Tamaki K, Kera A, Muto K, Yada T, Kitamura S, Asakawa S, Watabe S (2016) Different gene expression profiles between normal and thermally selected strains of rainbow trout, Oncorhynchus mykiss, as revealed by comprehensive transcriptome analysis. Gene 576:637–643
Tang E, Wongwarangkana C, Kinoshita S, Suzuki Y, Oshima K, Hattori M, Ineno T, Tamaki K, Kera A, Muto K, Yada T, Kitamura S, Asakawa S, Watabe S (2012) Global gene expression analysis of gill tissues from normal and thermally selected strains of rainbow trout. Fish Sci 78:1041–1049
Tsujimura A, Taniguchi N (1996) Reduction of phenotypic variation in reproductive traits of clonal strains of ayu Plecoglossusa ltivelis. Nippon Suisan Gakkaishi 62:393–398
Acknowledgements
This study was supported in part by a grant from the Ministry of Agriculture, Forestry, and Fisheries of Japan. We are grateful to the researchers at Aomori Prefectural Industrial Technology Research Center, Freshwater Fisheries Research Institute, for providing the rainbow trout. We also thank Mr. T. Setoguma and Ms. Y. Yoshino of the Marine Ecology Research Institute Central Laboratory for rearing the rainbow trout.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ineno, T., Tamaki, K., Yamada, K. et al. Thermal tolerance of a thermally selected strain of rainbow trout Oncorhynchus mykiss and the pedigrees of its F1 and F2 generations indicated by their critical thermal maxima. Fish Sci 84, 671–679 (2018). https://doi.org/10.1007/s12562-018-1217-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12562-018-1217-2