
Abstract
The Upper Seine Valley sees during the Bronze Age an increase in settlements and cemeteries characterized by a variety of architectures and funerary practices. This variability originates from the coalescence of two cultural traditions, the Atlantic culture from the northwest and the Continental culture from the east, which find its roots in the Upper Rhine area. In this context, our research aims to understand the cultural identity of these communities by reconstructing subsistence economy and palaeodietary patterns and to trace human mobility, which influenced lifestyle and social–economic dynamics. Adding new 10 AMS dating and using multi-isotope (δ13C, δ15N, δ34S) and multi-proxy (botanical, animal and human remains) biochemical investigations, we detected changes in dietary habits throughout the Bronze Age linked to varying C3 and C4 plant consumption and diverse animal protein intake by humans. Three main outcomes are highlighted: (1) a variety of farming strategies with different manuring intensity according to the cereal species; (2) site-specific dietary patterns; (3) an increase in millet and animal protein intake during the Late Bronze Age. Moreover, results from one cemetery reveal that individual mobility is more significant for males. These dietary variations, differential agricultural strategies and mobility are consistent with other western Europe research, supporting major global changes in all of Europe at the end of the Bronze Age.
Similar content being viewed by others

Introduction
A recent overview of land occupation during the Middle Bronze Age and the beginning of the Late Bronze Age (16th–12th century BCE) in France has enabled the compilation of a great amount of archaeological data that bears witness to a significant increase in the number of settlements and cemeteries during this period (Lachenal et al. 2017). This increase is particularly apparent in the Upper Seine Valley to the southeast of Paris, where Late Bronze Age contexts largely outnumber earlier Early Bronze Age (EBA: 22nd–18th century BCE) and Middle Bronze Age (MBA: 18th–14th century BCE) sites by about ten to one (Peake et al. 2017a). The strong dynamic in this area could be due to its geographical position on the boundary between two competing cultural traditions, the Atlantic culture that covers northwest France, the British Isles and the Low Countries and the north Alpine Continental culture that covers northeast France, the Upper Rhine area of southern Germany and Switzerland (Mordant 1989; Peake et al. 2017a). Moreover, during the Bronze Age, the Upper Seine Valley was at the centre of an extensive hydrographic network used for the transit of people and goods between northeaster and southern Europe, the Atlantic coast and beyond (Mordant et al. 2021). The cultural contact and the dynamic trade network would have largely contributed to the economic growth seen in the area at the beginning of the Late Bronze Age (LBA: 14th–9th century BCE). This would account for the sharp rise in the number of sites and the increase in population numbers that seems to have happened within a couple of generations. One of the hypotheses put forward for this rapid growth is the arrival of new populations during the 14th century BCE from the northeast of France, bringing with them their own cultural traditions. There is effectively clear evidence of a switch from Atlantic to north Alpine-influenced material culture and funerary practices around this time (Mordant 1989; Peake et al. 2017b; Roscio 2018). Simple inhumations devoid of grave goods, characteristic of the EBA and MBA Atlantic tradition, are replaced by inhumations with grave goods of north Alpine influence (Mordant and Roscio 2010). Therefore, early LBA material culture and funerary practices in the Seine Valley do indeed suggest that some human groups originated from the north Alpine area where funerary contexts are characterized by abundant and well-preserved grave goods, elaborate tombs and monuments. These rich archaeological contexts showcase a variety of practices that are directed by cultural influence and community-based traditions (Peake et al. 2017b; Adroit and Peake 2021).
The data from funerary contexts led us to set up Bronz’Pal, a research project with the aim to reconstruct the subsistence economy and the palaeodietary patterns of the communities settled in the Upper Seine Valley during the Bronze Age and whose lifestyle and social–economic dynamics were likely affected by the arrival of these new human groups. To reconstruct the cultural identity of these societies that share different influences, we have investigated botanical, faunal and human remains from 17 Bronze Age sites from the Upper Seine Valley to explore their food production and consumption and to trace mobility events using a multi-proxy and multi-isotope biochemical approach. By combining isotopic data performed on different trophic chain levels, we aim to evaluate how food patterns changed during the Bronze Age, as well as investigate the mobility of individuals or the origin of non-local individuals. Our objective is to assess if the dynamic that we have observed in the area is focussed solely on the exchange of goods and the diffusion of material culture or if it affected lifestyle and if so in what way. Particularly, the investigation of mobility will provide further details about these large-scale contacts, if they regarded only the material culture or involved “permanent” human mobility.
Archaeological context
Over the last 60 years, large-scale development (gravel extraction, road building and industrial parks) has opened up areas to archaeological survey making the Upper Seine Valley a reference point for Bronze Age and Early Iron Age archaeology in France. This development covers over 5000 hectares and has led to the discovery of over 130 settlements and funerary sites dating to the Bronze Age and the Early Iron Age (Brunet et al. 2017). Of the 70 cemeteries of this inventory, 43 have been excavated; they vary from large prestigious cemeteries (which include burial mounds now unfortunately ploughed out), with numerous preserved tombs to much smaller groups of less than 10 burials. While this funerary landscape was continually used during the Bronze and Early Iron ages, the peak activity occurred during the first stages of the Late Bronze Age, a relatively short period during which most of the funerary groups in the valley were established and to which the majority of the burials can be dated.
Inhumation is the main burial practice for the Early and Middle Bronze Age. During this early period, inhumations in a flexed position form small groups, generally no greater than five, that are often located around a circular funerary monument. Devoid of any grave goods, these burials are solely dated by radiocarbon analysis. The rare discovery of three cremation burials in Atlantic style pottery vessels identical to those found in the few Early Bronze Age domestic contexts underlines the area’s adherence to the Atlantic cultural group. The Middle to Late Bronze Age transition period (14th–13th century BCE) is particularly prolific in funerary evidence, as 140 burials and 10 monumental funerary sites in the Marolles-sur-Seine area alone date to this period. The LBA sees the rapid transition from inhumation to cremation, and by the end of the 13th century BCE, most burials are cremations, housed either in pottery urns or in organic containers (Delattre and Peake 2012). Burials included a number of personal items, essentially jewellery and objects relating to day-to-day activities, such as daggers, knives, razors and tools, as well as pottery vessels, all of the north Alpine cultural tradition showing the shift from one cultural tradition to another. Even though cremation seems to be almost exclusive during the LBA, there are exceptions to the rule as illustrated by an inhumation found in the fill of a ditched enclosure at the “La Croix de la Mission” site (BP_H20, in this study). Radiocarbon analysis dated this burial, as there were no preserved grave goods other than a collection of flint blades probably in a leather pouch worn at the waist. The obtained date of 1220–900 BCE cal (2σ) (Ly-8278) was unexpected but illustrates the contemporaneity of this burial with cremation deposits in the same cemetery. Cremation remained the main funerary practice in the area until the end of the Early Iron Age when inhumations reappear sometime during the 7th century BCE (Peake et al. 2017a; Le Goff and Peake 2021).
Subsistence economy
Knowledge on Bronze Age subsistence strategies have benefited by large-scale studies of archaeozoological and archaeobotanical remains recently undertaken in the Seine Valley (Bãlãşescu et al. 2008; Le Goff 2008; Auxiette et al. 2015; Toulemonde et al. 2017, 2022). Investigations into food production and consumption highlighted significant differences at the site level, and this evidence seems to be linked to the fact that most of the sites in this area were small family farms where food production systems were maintained locally (Auxiette et al. 2015; Toulemonde et al. 2018). Nevertheless, very little data are available for EBA and MBA as consequence of the limited number of settlements, preventing inferences for the reconstruction of the subsistence economy of the area for these periods. In contrast, from LBA onwards, numerous large settlements were found all around the Seine Valley, allowing archaeologist to reconstruct husbandry strategies and agricultural activities. LBA faunal assemblages are dominated by domesticated species including cattle, pigs and ovicaprids, with sheep being more common than goat. Though all domesticates are present at each site, their frequencies vary (Auxiette et al. 2015). Indeed, in some settlements such as at Marolles-sur Seine, “Motteux”, the animal bone assemblage is dominated by cattle, and in others, like Grisy-sur-Seine, the main meat source is represented by sheep (Hermetey 1995). At the end of the LBA, an increase in pigs to the detriment of other species, in particular cattle, is recorded in many sites (e.g. Villiers-sur Seine) (Auxiette et al. 2015). These different patterns suggest site-specific livestock strategies developed according to the different needs. Game varies by site, but hunting was still an important activity, and the meat consumption from wild animals seems to be linked to the status of the site’s inhabitants or particular social events which implied for large number of people meeting (Auxiette et al. 2015).
As for the fauna records, archaeobotanical data are very few for the earlier phases of the Bronze Age, as settlements are practically unknown for these periods in the Upper Seine Valley. The subsistence economy is therefore difficult to document. Nevertheless, data from northeastern France, Moselle and Alsace, suggest that during this early period, farming mostly focussed on cereals, such as hulled wheats and barley (Toulemonde et al. 2022). Foraged foods may have been also an important resource, as pits filled with charred acorns are common at EBA settlements in Moselle. Starting at the end of MBA/initial stage of LBA, major changes occurred in crop production and subsistence economy, even though a large spectrum of cultivated species (e.g., hulled and naked barley, emmer, einkorn, spelt, new glume wheat, hexaploid and tetraploid types of naked wheat) and some gathered species make up the plant assemblages (Toulemonde 2010, 2013; Auxiette et al. 2015). Indeed, the numerous archaeobotanical data available for the Upper Seine Valley during LBA indicate the introduction of new crops such as millets (e.g. broomcorn millet, foxtail millet) in addition to more intense cultivation of the more comment cereal species like wheat and barley, as well as pulses and oil plants (Toulemonde 2013). Moreover, studies on weeds and cultivated crops attest that agriculture was based on an intensive system of small plots of land constantly maintained and cultivated, year in, year out (Toulemonde 2013; Auxiette et al. 2015).
Archaeological sites of the Upper Seine Valley
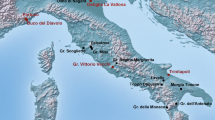
The 17 sites, considered here, date from the Early Bronze Age to the Middle of the Early Iron Age and are located in the floodplain of Upper Seine Valley, no more than 2 km from the river’s present course (Fig. 1). Settlements are regularly spaced along small tributaries and palaeochannels, built on the higher gravel domes less susceptible to flooding. They are generally short-lived installations, a family smallholding or farm, composed of several pits and posthole buildings (Peake et al. 2017a). Bronze and Early Iron Age cemeteries are located along the whole of this section of the Seine valley, with a concentration of funerary sites in the interfluvial plain between the Seine and Yonne rivers to the south of Marolles-sur-Seine. This part of the valley offers a unique topographical location, a broad flat plain set between two major rivers, thus making it possible for the funerary sites to truly dominate the surrounding countryside. Cemeteries mark the landscape more permanently than settlements (Adroit and Peake 2021). Most include large circular mounds that cover prestigious burials, which became the founding monuments of extensive funerary groups. Most cemeteries seem to correspond to family groups, their use being intrinsically linked to nearby settlements (Delattre and Peake 2012). However, some funerary spaces are used intermittently over several centuries as a systematic funerary space by local inhabitants, and perhaps by communities from farther afield. This is the case of two cemeteries included in this study, Marolles-sur-Seine “La Croix de la Mission” and Jaulnes “le Bas des Hauts Champs” (Peake and Delattre 2005, 2010). The earliest tombs dating to the Early Bronze Age are inhumations with cremation gradually becoming the dominant and the only practice during the initial phase of the Late Bronze Age (circa 1200 BCE). Inhumations therefore constitute a small minority of the corpus of Bronze Age burials available in the area, and care has been taken to sample as many as possible for this study.

Map of the sites investigated in this study: (1) Ville-Saint-Jacques “le Bois d’Echalas”, (2) Varennes-sur-Seine “la Maladrie”, (3) Cannes-Ecluse “le Petit Noyer”, (4) Marolles-sur-Seine “la Croix de la Mission”, (5) Marolles-sur-Seine “les Prés-Hauts”, (6) Marolles-sur-Seine “Bois de Marolles”, (7) La Tombe “la Cour des Lions”, (8) Marolles-sur-Seine “les Pâtures communales”, (9) Châtenay-sur-Seine “la Roche”, (10) Barbey “les Cents Arpents”, (11) Barbey “le Chemin de Montereau”, (12) Egligny “la Pêcherie”, (13) Balloy “la Haute Borne”, (14) Jaulnes “le Bas des Hauts Champs”, (15) Grisy-sur-Seine “les Champs Pineux”, (16) Grisy-sur-Seine “les Roqueux”, (17) Grisy-sur-Seine “la Ferme d’Isle” (map R. Peake, Inrap)
Seven cemeteries, located along the Upper Seine Valley, have provided human bone samples. Burials from Barbey, Châtenay-sur-Seine, Jaulnes and Varennes-sur-Seine date to the end of the Middle Bronze Age and the beginning of the Late Bronze Age, whereas the “millennium” cemetery at Marolles-sur-Seine “la Croix de la Mission” has provided burials dating to the Early, Middle and Late Bronze Age (Fig. 2). Samples dating to the Early Bronze and Middle Bronze Age come from the burial sites at Egligny and Marolles-sur-Seine “les Prés Hauts”. Animal bone and archaeobotanical samples have been selected from six settlement sites dating to the Late Bronze Age/Early Iron Age (1350–450 BCE) (Fig. 1 and Table 1).

A EBA burial from Marolles-sur-Seine (photo N. Ameye, Inrap); B LBA burial in a crouched position from the Barbey cemetery (sep. 7 photo C. Mordant, Univ. Bourgogne); C plan of the monumental cemetery Marolles-sur-Seine “la Croix de la Mission” (photo P. Pihuit, Inrap); D initial LBA inhumation at Varennes-sur-Seine “La Maladrie” (photo C. Valero, Inrap)
Stable carbon, nitrogen and sulphur analysis in bioarchaeology
Stable isotope analyses are routinely applied to reconstruct human and animal dietary patterns and agricultural practices to infer lifestyles, socio-economic organizations and trading activities (Katzenberg 2008; Knipper et al. 2014). Bone is constituted of a mineral component, hydroxyapatite and an organic component, collagen, whose compositions depend on the exploited environment and the type of food consumed (food and drinks). Collagen records the protein components of the diet, and its chemical composition reflects the food consumed over the last ca. 15 years of an individual’s life (Hedges et al. 2007). Bone collagen has a permanent turnover with a different rate according to the age and health status. According to the trophic chain, the plants at the bottom of the chain have isotopic values that vary according to several parameters (species, environment, type of photosynthesis). They are therefore a determining element for the isotopic values of the rest of the food chain. A fractionation in favour of the heavy isotope occurs from one consecutive link of the chain to the next, and the heavy isotope values increase (13C, 15N, 34S), at a rate estimated between 0 and 1‰ for carbon, between 3 and 5‰ for nitrogen and lower than 1‰ for sulphur (DeNiro and Epstein 1981; Minagawa and Wada 1984; Schoeninger and DeNiro 1984; Nehlich 2015). Stable carbon isotope values (δ13C) provide information about the environment where the foodstuffs originated (e.g. terrestrial vs. aquatic) and in particular the type of plants consumed. For example, C3 type plants (e.g. barley, wheat) are typical of a temperate environment, while C4 type plants (e.g. millet, sorghum) are widespread in open and warm environments. Stable nitrogen isotope values (δ15N) are used to infer on the trophic level occupied by an organism. Plants, being at the base of the trophic chain, present lower δ15N values, and as a consequence, δ15N values of predators are higher than that of their prey (DeNiro and Epstein 1981; Schoeninger and DeNiro 1984). Moreover, it has been demonstrated that the nitrogen analysis on plant remains (seeds, chaff) contributes to better understand agricultural strategies and changes across time, such as the use of the manure (Bogaard et al. 2007; Styring et al. 2014) and the hydric conditions in which plants grown (Ferrio et al. 2005; Wallace et al. 2013). Stable sulphur isotope values (δ34S) provide information on the origin of food sources—from a terrestrial, marine or freshwater ecosystem (e.g. Drucker et al. 2018). The δ34S values are related to the geology and can vary greatly from one region to another (Nehlich 2015), therefore, having the potential to provide information about mobility by detecting non-local individuals (Vika 2009; Varalli et al. 2016b). All the sites analysed here are placed in the Seine Valley on quaternary alluvial deposits (Fig. 1), but the surrounded area is dominated by Upper Cretaceous deposits (chalk) at the south of the Seine towards the Yonne area and by Oligocene and Eocene deposits at the north of the Seine river (https://www.geoportail.gouv.fr/). According to this, the bone collagen δ34S values can bear witness to the mobility of individuals within a population but only in the last 5–10 years of an individual’s life (Nehlich 2015). The application of Bayesian mixing model like FRUITS (Food Reconstruction Using Isotopic Transferred Signals) supports diet reconstruction studies. In this research, FRUITS is used to identify site and period-specific dietary patterns. Thanks to the combination of the different potential sources (plants, i.e. cereals and legumes, herbivores and omnivores) dated to the different periods, FRUITS incorporates the capability to account for dietary routing, that is, the contribution of different food fractions, such as the macronutrients, of the resources of each period towards a dietary proxy signal measured in the coeval consumer.
Material and methods
A total of 114 samples, human and animal skeletal material and archaeobotanical remains originating from 17 sites in the Upper Seine Valley were considered for this study (Table 2). The bioarchaeological dataset includes 41 humans of different age and sex. The age of the individuals was estimated according to Bruzek et al. (2005) and Schmitt (2008) for the adults and Birkner (1980), Ubelaker (1987) and Scheuer and Black (2000) for the infants and juveniles. The sex was estimated using Bruzek (1991). The strategy was to sample all of the inhumations from each site, with exception of Barbey where there was no skeletal material for about a third of the recorded burials. The animal assemblage is composed of 39 wild and domestic animals (pigs, wild boar, cattle, sheep, horse, deer, dogs). Wild and domestic animals were sampled from a large number of settlements located along the valley with a limited number of samples from each site and contemporary to the cemeteries. The strategy was to sample all available domestic species and consumed wild species. The archaeobotanical assemblage is made of 34 seeds from cultivated plants, mainly cereals like wheat (Triticum dicoccum), barley (Hordeum vulgare), broomcorn millet (Panicum miliaceum) and legumes like lentils (Lens culinaris). This sampling was more opportunistic due to the poor preservation of plant remains, and only two settlements provided samples from dated contexts.
The human and animal bone collagen extraction and the botanical remains pre-treatment were performed at the Laboratoire Méditerranéen de Préhistoire Europe Afrique, Aix Marseille University (LAMPEA, Aix-en-Provence. France), and the isotopic ratios were measured at the Iso-Analytical Limited Laboratory (Crewe, UK). 14C measurements on the collagen samples were performed at the Poznań Radiocarbon Laboratory.
Analytical method
The human and animal bone collagen extraction was performed according to the acid–base-acid protocol proposed by Longin method (1971) and modified by Bocherens et al. (1991). Each powdered sample (> 700 µm) was demineralised in HCl solution (1 M, 20′), then cleaned of contaminants in NaOH solution (0.125 M, < 18 h) and then solubilized in HCl solution (pH = 2, 17 h, 100 °C). The gelatinised collagen was then freeze-dried for 48 h.
A modified protocol as described by Aguilera and colleagues has been applied to purified the botanical remains (Aguilera et al. 2017). The grains were soaked in HCl (1 M, room temperature, for 24 h) and then rinsed three times with distilled water (24 h, 12 h and 6 h). Once dried in the oven, the samples were finely powdered.
For collagen, carbon and nitrogen isotope values are calibrated to V-PDB and AIR using IA-R068 (soy protein, δ13CV-PDB = − 25.22 ± 0.02‰, δ15NAIR = 0.99 ± 0.07‰) and sulphur isotope values to V-CDT using IA-R061 (barium sulphate, δ34SV-CDT = 20.33 ± 0.13‰). For seeds, carbon and nitrogen isotope values are calibrated to V-PDB and AIR using IA-R001 (wheat flour, δ13CV-PDB = − 26.43 ± 0.02‰, δ15NAir = 2.55 ± 0.02‰) for seed.
For collagen samples, carbon and nitrogen measurement uncertainty was monitored using tree check samples: IA-R038 (L-alanine, δ13CV-PDB = − 24.99 ± 0.06‰, δ15NAIR = − 0.65 ± 0.04‰), IA-R069 (tuna protein, δ13CV-PDB = − 18.88 ± 0.05‰, δ15NAIR = 11.60 ± 0.09‰) and a mixture of ammonium sulphate and IA-R046/IAEA-C7 (oxalic acid, δ13CV-PDB = − 14.48 ± 0.21‰, δ15NAIR = 22.04 ± 0.06‰). Sulphur measurement uncertainty was monitored using two check samples: IA-R068 (soy protein, δ34SV-CDT = 5.25 ± 0.10‰) and IA-R069 (tuna protein, δ34SV-CDT = 18.91 ± 0.22‰). For seed samples, carbon and nitrogen measurements uncertainty was monitored using two check samples: IA-R045/IA-R005 (mixture of ammonium sulphate and beet sugar, δ13CV-PDB = − 26.03 ± 0.11‰, δ15NAIR = − 4.71 ± 0.07‰) and IA-R046/IA-R006 (mixture of ammonium sulphate and cane sugar, δ13CV-PDB = − 11.64 ± 0.03‰, δ15NAIR = 22.04 ± 0.06‰).
For carbon and nitrogen, IA-R068, IA-R038, IA-R069 and IA-R001 are calibrated against and traceable to the international standards IAEA-CH-6 and IAEA-N-1. For sulphur, IA–R061 is calibrated using NBS-127, IAEA-S-1 international standards and IA–R068 and IA–R069 using NBS–127 and IAEA–SO–5.
The isotope ratios are reported as delta that has been defined according to IUPAC (International Union of Pure and Applied Chemistry): δ = (Rs/Rst) − 1 = 103[(Rs/Rst) − 1] ‰ where Rs and Rst are the isotope ratios 15N/14N, 13C/12C and 34S/32S of the isotopic abundances of 15N, 14N, 13C, 12C, 34S and 32S and ‰ = 10−3. Using calculation provided by Szpak et al. (2017) based on repeated measurements of calibration standards, check standards and sample replicates, the standard analytical uncertainty (Uc) is estimated, for collagen, to be ± 0.19‰ for δ13C and δ15N and ± 0.28‰ for δ34S. For seeds, due to the small sample size, no replicate measurements were performed; therefore, the uncertainty (U(bias)) was calculated with calibration standards and check standards which are ± 0.10‰ for δ13C and ± 0.13‰ for δ15N (SI1).
Collagen preservation quality was assessed considering the wt% collagen ≥ 1%, %C ≥ 30%, %N ≥ 15%, %S ≥ 0.15% and atomic C/N (2.9 < C:N < 3.6), atomic C/S (300 < C:S < 900) and atomic N/S (100 < N:S < 300) (DeNiro 1985; Ambrose 1990; van Klinken 1999; Nehlich and Richards 2009).
We applied the Bayesian model using the software FRUITS to reconstruct the dietary scenarios (Fernandes et al. 2014). Two offsets were used for carbon: (a) 4.8‰ between the δ13C values of the plants and the collagen of their consumers and (b) 0.8‰ between the carbon isotope values of the collagen of consumers of the previous trophic level (Styring et al. 2017). For nitrogen isotope compositions, a spacing of 4.0‰ was considered between the diet and the consumer of two different trophic levels (Styring et al. 2017). The values of the C3 plants, C4 plants, herbivore meat and omnivore meat used to make estimations are from this study, while the freshwater fish values are from the Lake of Bourget (Varalli et al. 2021). All details about the application of the models and the data are illustrated in SI3.
Statistical analyses were performed using the statistical program R (version 3.6.1), and data were visualized using RStudio (version 1.2.5033) (R Development Core Team 2005; R Core Team 2018; R Studio Team 2020) and Excel. Considering the small sample size of the group (n < 15), the non-parametric Wilcoxon and Kruskal–Wallis tests were used to determine differences between group means for δ13C, δ15N and δ34S values. A 0.05 probability (p < 0.05) is considered significant. Bayesian models using the software FRUITS (Beta version 2.1) were applied to reconstruct the dietary scenarios (Fernandes et al. 2014).
Calendar ages were determined using OxCal v 4.4 (Bronk Ramsey 2009) using the IntCal20 Northern Hemisphere atmospheric curve (Reimer et al. 2020). Date ranges are calibrated at the 2-sigma range.
Results
Preservation of the samples
No set criteria exist to verify seed preservation for stable isotope analyses. However, C:N ratios contribute to assess the reliability of the crop measurements. The C:N ratios of the cereals lie between 8.4 and 47.1 and the C:N of the lentils between 9.6 and 22.9. Comparing these data with other archaeobotanical and modern seed results from experimental research (Fraser et al. 2013; Vaiglova et al. 2014), our data fit in the range of the archaeological variability with the exceptions of two samples, BP_S04 (C:N = 47.1) and BP_S13 (C:N = 8.4). Considering that these samples show also the highest δ15N values, we exclude them from our analyses to avoid misinterpretations (Table 3).
Concerning the animals, four samples (BP_F03, BP_F06, BP_F18, BP_F32) showed a low collagen yield, and then they were excluded. Carbon and nitrogen isotopic measurements were then carried out on 35 samples. Of these, the bovine BP_F04 and the swine BP_F13 do not meet the %C, %N or C:N criteria, and they are then excluded in the discussion. Eight animals (BP_F05, BP_F16, BP_F17, BP_F26, BP_F27, BP_F31, BP_F33, BP_F35), having %C just below the threshold, are included in the discussion and considered with caution. No significant correlations exist between %C and δ13C, %N and δ15N suggesting they are reliable data (Fig. 1S2). Sulphur analysis was performed on a selection of 15 animals showing the highest collagen yield. One caprine from Grisy (BP_F35) has the %S at the limit of the threshold, and it has been considered with caution (Table 4).
Concerning the human remains, 10 individuals (BP_H18, BP_H27, BP_H28, BP_H29, BP_H100, BP_H101, BP_H102, BP_H108, BP_H111 and BP_H113) presented a low collagen yield, and they were excluded. The collagen was then isotopically measured for 31 individuals, and, among them, 2 individuals (BP_H16, BP_H25) did not meet the %N, %C or C:N conditions, and then they were excluded. Four individuals (BP_H03, BP_H04, BP_H06, BP_H17) have values at the thresholds of the limits; they are included in the study and considered with cautious. The C/N is included in the accepted range, and no significant correlations exist between %C and δ13C, %N and δ15N, suggesting they are reliable data (Fig. 1S2). The sulphur analyses have been performed on 27 individuals because four individuals (BP_H17, BP_H103, BP_H105, BP_H109) did not have enough collagen. Two individuals (BP_H16, BP_H25) have the %S below the accepted limit, and then they were excluded by the discussion. Two other individuals (BP_H04, BP_H06), being just below the accepted limit, were included in the study and considered with caution (Table 4; Fig. 1S2).
Radiocarbon dates
AMS 14C dates were performed on 10 human bone samples from various funerary sites across the valley: Varennes-sur-Seine, Marolles-sur-Seine, Jaulnes and Egligny. All the results, in addition to 9 humans, 7 animals and 2 botanical samples previously analysed (Gouge and Peake 2005; Peake and Delattre 2005; Rottier et al. 2012; Ameye et al. 2016; Toulemonde et al. 2022), are shown in Fig. 3 and Table 1SI2. Not all samples were radiocarbon dated for financial reasons; however, those samples that did not receive a 14C date do come from contexts reliably dated by their material culture (pottery and metal objects).

Calibrated AMS dates for 28 of the samples included in this study. New 14C dates are highlighted in green. Date ranges are calibrated at the 2-sigma range. Date calibrations were produced in OxCal v.4.4 (Bronk Ramsey 2009) using the IntCal20 Northern Hemisphere atmospheric curve (Reimer et al. 2020)
Botanical results
The wheat and barley (C3 plants, n = 16) raw data show a mean of − 24.1 ± 0.8‰ for δ13C and 5.6 ± 1.2 ‰ for δ15N. The broomcorn millet (C4 plants, n = 8) shows a mean of − 10.0 ± 0.2‰ for δ13C and 6.6 ± 0.9‰ for δ15N. The lentil mean is − 23.2 ± 5.5‰ (n = 8) for δ13C and 3.0 ± 1.7‰ for δ15N (Table 5). A correction of − 0.31‰ is made to the nitrogen isotopic values as a result of the carbonization phenomenon (Nitsch et al. 2015).
Animal results
The domestic and wild terrestrial animals' carbon and nitrogen values (n = 33) show a mean of − 21.7‰ ± 1.3 for δ13C and 7.0‰ ± 1.2 for δ15N. The δ34S values (n = 15) are widespread and show a mean of 3.6‰ ± 6.3 (min, − 6.6‰; max, 15.1‰) (Table 5). The non-parametric Kruskal–Wallis test indicates significant differences among the three main animal groups (pigs, cattle and sheep) for both δ13C and δ15N. Wilcoxon Mann–Whitney U-tests are shown in the online resource, and they highlight significant differences for both δ13C and δ15N between almost each pair of groups (Table 2SI2).
Human results
Human summary statistic is shown in Table 5. The δ13C and δ15N values for the 29 individuals have the δ13C mean of − 19.5 ± 1.0‰ and the δ15N mean of 11.0 ± 0.8‰. The δ34S values for 25 of those individuals have a mean of 7.9 ± 3.3‰ (min, − 1.9‰; max, 12.7‰). Statistical results by site are presented in the online resource (Tables 3 and 4SI2), and differences are highlighted between Marolles and Barbey for carbon and sulphur. Due to the small number of samples for each site, a statistical test for each period cannot be used. Nevertheless, clear dietary patterns according to the periods are distinguishable and they are widely described in the discussion section.
Discussion
The agricultural practices and livestock herding
Natural environmental factors (e.g. climate, water availability, salinity) influence the isotopic ratios of the plants that are at the bottom of the trophic system (e.g. Farquhar et al. 1989; Körner et al. 1991); however, experimental studies of traditional controlled agricultural systems have demonstrated that the carbon and nitrogen values of the plants provide information about the human management of crops, such as the use of manure (Bogaard et al. 2013).The use of manure to improve soil fertility causes an enrichment of 15N in plants, proportional to the duration and intensity of manuring. This can bee seen in the δ15N values of seeds, with those below 3% reflecting the absence of fertilizers, values between 3‰ and 6‰ intermediate level of fertility and values above 6‰ indicating intense and constant manure use. According to these ranges, the corrected results of the barley of this study span between 3.0‰ and 6.4‰ and the wheat results span between 4.8‰ and 7.5‰, suggesting a general intermediate/high level of fertilization of these plants (Fig. 4). Furthermore, wheat has higher δ15N values than barley (wheat δ15N = 5.8 ± 1.1‰; barley δ15N = 4.9 ± 1.2‰), suggesting a likely higher level of fertilization for this crop compared to the barley. Based on recent experimental studies on different millets species, the broomcorn millets δ15N values, higher than 6‰, seem to suggest they were fertilized as well (Christensen et al. 2022). Even the lentils that are less sensitive to the manuring and whose values when not amended are usually around 0‰ (Fraser et al. 2011) show δ15N around 3‰. This evidence highlights the wide use of manure and confirms how human activities affected the farming strategies.

Archaeobotanical and archaeozoological δ13C and δ15N results (animals: mean and standard deviation. EBA, Early Bronze Age; LBA, Late Bronze Age). The archaeobotanical δ15N values are corrected for carbonization
According to the dates of the different contexts, the barley from Balloy is dated to LBA, while the lentils, wheat and broomcorn millet from Ville St. Jacques are dated to the EIA (Toulemonde et al. 2022). Even though climatic and environmental changes which could have affected the isotopic values cannot be ignored and at present cannot be quantified (Magny et al. 2007; Kneisel et al. 2015; Capuzzo et al. 2018), it is interesting to highlight a possible slight increase in the use of the manure from the LBA to the EIA. Furthermore, this hypothesis may be supported by analyses on weed flora, using a nitrogen indicator for groups of settlements from different territories of Champagne and Île-de-France regions, dated to the LBA and EIA (Toulemonde 2013). These data indicate that during the LBA and EIA (1350–475 BCE), when the same farming system was in use, there was no significant change or decrease in cultivated soil fertility. However, to avoid speculations, further investigation on all the species from all the periods need to be analysed to confirm this hypothesis.
Estimated animal diets evaluated from the animal bone collagen isotopic ratios provide important information on animal husbandry (e.g. Varalli et al. 2021). The animal isotopic ratios show a predominant consumption of C3 temperate environment foodstuffs. Because some species are represented by a small sample size, statistical analyses could not be performed. However, variability in δ13C and δ15N values suggests differences in resources that animals consumed. Values may also reflect environmental shifts (Figs. 4 and 5). The EBA herbivores, represented by three red deer, have lower δ13C values compared to the LBA animals, while their δ15N values overlap with those of plants. These patterns do not seem linked to different husbandry practices during the two period. Instead, it is more likely that these differences are resulted to local environment shifts. Bronze Age environmental change in the Paris Basin, as recorded by pollen data, has been attributed to both climatic and anthropogenic influences (Jeune et al. 2012; Leroyer et al. 2012). Even though the small sample size for the EBA begs for cautious interpretation, similar isotopic ranges have already been recorded in other European regions, e.g. Switzerland, Italy (Masotti et al. 2019; Varalli et al. 2021, 2022).

Mean and standard deviation of the δ13C (A), δ15N (B) and δ34S (C) animal results of the Bronze Age and of local Neolithic sites of the Seine Valley (Balasse 1999; Cheung et al. 2021). For δ34S all available data from other periods have also been included (Bocherens et al. 2011; Cheung et al. 2021). In red Wilcoxon tests results
The LBA δ13C and δ15N values show wide ranges within each species group. This evidence suggests that no specific patterns are present at the site level, but eventually it may also indicate that animal management followed a broader region-wide practice, highlighting that local level strategies could have been replaced by shared management practices. When LBA faunal isotope values are considered together for the entire region, cattle have lower δ13C and δ15N values compared to sheep, pigs, and dog. This could suggest that cattle were mainly fed with wild plants. The domestic omnivores such as pigs and the dog show higher δ13C values than the other animals of this period. Their values, spanning from − 22.1 to − 18.8‰, suggest a mixed C3–C4 diet. Specifically, the dog shows the highest δ13C value, and this ratio falls in the range of the humans, implying that this animal closely lived with humans eating similar foodstuffs or the leftovers. Moreover, based on the animal bone collagen values, herbivores and omnivores seem to have mainly consumed none or little manured plants (Fig. 4). In particular, the herbivores would have expressed much higher carbon values if they had been fed domestic cereals. In contrast, the estimated diet of the pigs seems to be consistent with the consumption of cultivated seeds, confirming that these animals may have consumed the leftovers of the human meals.
Among animals, δ34S values show a significant variability, ranging from − 6.6 to 15.1‰ (n = 15). Previous research in the Seine Valley has already highlighted this wide animal range (Cheung et al. 2021). However, 8 animals (53.3%) are included in a relatively small range (4.1‰ < δ34S < 9.7‰), while 7 animals (46.7%) originating from Grisy-sur-Seine and Chatenay-sur-Seine are outside this range, with lower and higher values. These data are in line with the δ34S range of the Seine Valley, which correspond to the local bedrocks dated to the Late Cretaceous (https://www.geoportail.gouv.fr/) (Cheung et al. 2021). Considering the δ34S available data from the wild and domesticated terrestrial animals previously analysed (δ34S mean = 4.6 ± 3.1, n = 27; Bocherens et al. 2011; Cheung et al. 2021), many of the LBA animals fall within this terrestrial species range. However, as suggested by Cheung et al. (2021), it is likely that the wider domestic animal variability could be the result of a livestock management based on a “free-range” approach, where the animals were herded in a larger area with diverse geological bedrocks, compared to the other sites where the husbandry regime was more “hands-on” with the animals contained within a restricted area (Rey et al. 2019; Cheung et al. 2021). This hypothesis calls for further investigations seeing the high δ34S variability of the Paris Basin (i.e. Essonne, Marne, Seine and Yonne Valley). An alternative hypothesis is that the animal variability is the result of a different geographical origin of some animals, suggesting the on-site presence of non-local individuals as proposed by Rey et al. (2019) for animals of the Yonne Valley. Interactions among different areas resulting in the exchanges of animals as gifts or real trade could explain the presence of non-locally reared animals. For instance, animals reared elsewhere and then probably brought into the settlements for consumption has been proposed for the coeval site of Villiers-sur-Seine (Auxiette et al. 2015; Peake 2020). Furthermore, five of the LBA animals show negative values. Considering that the freshwater animals of the Seine Valley present negative values (δ34S mean = − 15.4 ± 3.2, n = 4; Bocherens et al. 2011; Cheung et al. 2021), these animals may have consumed some resources from an aquatic environment (Fig. 5). Even though it is difficult to confirm this interpretation, these results underline the great variability in husbandry—and probable intense trade of animals, in the Seine Valley during the Bronze Age.
The human diet
The δ15N, δ13C and δ34S values of the human samples from Barbey, Marolles-sur-Seine, Jaulnes and Varennes-sur-Seine suggest the presence of different dietary habits at inter- and intra-population levels, as confirmed by the significant statistical tests results (Tables 2 and 3SI, Fig. 6). All the isotopic values fall into a wide range, particularly carbon and sulphur (∆13C = 3.7‰, ∆34S = 11.6‰,). The variability in δ15N values indicates a consumption of different animal proteins (meat or dairy products) among the human groups, as well as a probable contribution of resources from different terrestrial and aquatic environments. Variability in δ13C values indicate a mixed consumption of C3 and C4 plants, which are less enriched in 13C such as barley and wheat (C3 plants) and more enriched in 13C such as foxtail and broomcorn millets (C4 plants). The isotopic values of humans dating from the Early to the Late Bronze seem to be site-specific and linked to the chronology.

Stable carbon and nitrogen isotopic values for human and animal remains of the present study
The analyses carried out on the samples from Marolles-sur-Seine show that diet was mainly based on terrestrial plant and animal resources from a C3 environment. Though most individuals from the site have similar values suggesting a homogeneous C3 diet, one individual, BP_H20, has higher δ15N (11.6‰) and δ13C values (− 18.0‰) compared to the rest of the community which would indicate an important contribution of C4 plants such as millet to the diet of this individual (Fig. 6). This difference could be linked to the fact that BP_H20 is a more recent burial dating to the LBA (1220–900 cal BCE), whereas the other burials are dated to the EBA (BP_H109, δ15N = 11.1‰, δ13C = − 20.7‰; BP_H110, δ15N = 11.3‰, δ13C = − 20.5‰) and MBA (BP_H21, BP_H22, BP_H23, BP_H24: δ15N mean = 10.2 ± 0.8‰, δ13C mean = − 20.7 ± 0.1‰). The difference in the diet between BP_H20 and the rest of the community is also confirmed by the FRUITS Bayesian model used to estimate the relative contribution from different sources (Fig. 7A and E). The EBA-MBA group and the LBA burial clearly show different patterns, with a contribution of C3 and C4 plants of 37.2% and 21.1%, respectively, for the EBA-MBA burials, and with a contribution of C3 and C4 plants of 19.9% and 39.8%, respectively, for the LBA burial BP_H20 (SI3). Based on the Bronze Age local animals, one may note a change in the origin of the animal protein intake between the two periods. Human data highlight similar levels of consumption of proteins from omnivores and herbivores during the EBA-MBA, in contrast with the LBA, which suggest a greater protein intake from omnivores than from herbivores.

Contributions of foodstuffs as estimated by the FRUITS Bayesian model using δ13C and δ15N values of the different resources (for the values entered refer to SI3). The boxes provide a 68% confidence interval (corresponding to the 16th and 84th percentiles), while the whiskers provide a 95% confidence interval (corresponding to the 2.5th and 97.5th percentiles)
Most human δ34S values fall in the same range as animals, likely reflecting a predominantly terrestrial diet in line with previous assessments. Indeed, no correlation between δ34S and δ15N values occurs (R = − 0.4, p = 0.419). Only the individual BP_H23, a child, has a higher δ34S value of 12.7‰ than the other individuals. The burial has no grave goods and even though the skeleton is too badly preserved to provide any information on health, it is difficult to reject the impact of pathologies whose aetiology would have affected the δ34S. Additional analysis (e.g. strontium, oxygen, amino acids) will provide information to reconstruct its life history.
The Barbey individuals all date to the LBA. Both C3 and C4 plants and terrestrial animals make up the human diet. Some individuals (BP_H02, BP_H03, BP_H05, BP_H06, BP_H10, BP_H13) show quite high δ13C values (mean = − 18.5 ± 0.3‰, n = 6), suggesting a greater contribution of C4 plants to the diet compared to the other individuals (BP_H01, BP_H04, BP_H07, BP_H08, BP_H9, BP_H11, BP_H12, BP_H14, BP_H15, BP_H17, BP_H18) whose δ13C mean value is − 19.6 ± 0.3‰ (n = 11) (Fig. 6). FRUITS Bayesian model confirms this trend attesting a consumption of C3 and C4 plants of 29.1% and 30.2%, respectively, with a slightly more important intake of proteins from the omnivores than herbivores (17.4% vs. 16.2%) (Fig. 7D). The δ13C and δ15N values do not highlight a different diet according to sex, as the isotopic ranges of males and females overlap. However, when considering δ34S values, a difference between the two groups is evident: males have significantly higher δ34S values than females (Mann–Whitney U-test, p < 0.05) (Fig. 6). Two competing hypotheses might explain this variation. Either δ34S values reflect dietary habits that differed between males and females, or they represent different origins of males compared to females.
The individual from Varennes-sur-Seine shows δ15N and δ13C values distributed within the range of the Barbey individuals, implying a mixed C3–C4 diet. Furthermore, herbivore and omnivore protein consumption are equivalent (Fig. 7C), which is consistent with the burials later date corresponding to the initial LBA. The δ34S value fits in the estimated local range, confirming a diet mainly based on local terrestrial resources.
The Jaulnes cemetery provides remarkable information about diet variability in the area during the Bronze Age. Indeed, while the δ15N values are contained within a narrow range (∆15N = 0.9‰), the δ13C values have a significantly wider range (∆13C = 3.7‰). The three humans with the lowest δ13C values are dated between 1440 and 1130 cal. BCE (MBA and Early LBA), whereas BP_H103 dating to 1220–900 cal. BCE has the highest δ15N and δ13C values of the group (δ13C = − 17.2‰ and δ13N = 11.3‰). We can therefore distinguish two different dietary patterns according to the dates of the burials: if C4 plant intake was quite low, no more than 20% of the entire diet at the end of the MBA-early LBA, C4 plants contribute up to 45.1% of the diet from the middle phase of the LBA. In addition, during this same period, a slight increase in omnivore protein consumption was detected compared to herbivore protein consumption from the MBA to the middle of the LBA (Fig. 7B and F). The δ34S values are available for only two individuals, BP_H106 and BP_H107. Interestingly, they both present negative values, which could suggest a consumption of freshwater resources from the Seine River according to the δ34S of freshwater animals (Bocherens et al. 2011; Cheung et al. 2021). No fauna from this site is available, however, animals from the nearby settlement of Grisy-sur-Seine show an important variability, making it difficult to evaluate the contribution of this resource to the diet.
To summarize, the human samples dating from the EBA to the LBA from four cemeteries along the Seine Valley that are just some kilometres apart clearly illustrate a change in food habits during the Bronze Age. An enrichment in the heavier carbon and nitrogen isotopes is attested, and a clear shift is evident for δ13C, suggesting an increase in the consumption of C4 plants from the EBA to the LBA. The highest δ13C values are recorded for the two most recent individuals of the dataset, BP_H20 from Marolles-sur-Seine and BP_H103 from Jaulnes (Fig. 6). This change in the main staple crop is linked to a greater omnivore consumption, such as pigs, in the form of meat or dairy products from cattle, sheep and goats.
Prehistoric diet change in the Seine Valley
Several isotopic studies have been undertaken in the Paris Basin especially for the Neolithic (Balasse 1999; Bocherens et al. 2011; Naito et al. 2013; Goude et al. 2017; Rey et al. 2017, 2019; Drucker et al. 2018; Cheung et al. 2021). However, only two isotopic studies have focused on cemeteries in the Upper Seine Valley, one study on human samples dating to Middle Neolithic (c. 4700–3500 BCE) from Balloy “Les Réaudins” (n = 36) (Cheung et al. 2021) and one study on samples from the LBA cemetery at Barbuise (n = 5) (Goude et al. 2017).
Considering first the LBA, the domesticated herbivores and omnivores from our study show similar values to those from Barbuise, despite the small number of individuals and the isotopic variability outlined by the authors (means of our study, δ13C = − 21.7 ± 1.0‰, δ15N = 7.0 ± 1.1‰, n = 26; means of Barbuise, δ13C = − 21.3 ± 1.6‰, δ15N = 6.6 ± 0.7‰, n = 3; Goude et al. 2017). This evidence suggests a C3 diet in line with the results from the other LBA sites and this similarity in values allows for a direct comparison between the human groups. The results obtained from the bulk collagen from the Barbuise human samples indicate that the protein intake originated from a C3 environment, likely animals fed a C3 diet. However, the bioapatite δ13C values from enamel show a low-protein C4 foods such as millets (Goude et al. 2017). This is in line with isotopic results from the other LBA sites of the area. Major differences occur in the δ15N values. The humans from Barbey, Marolles-sur-Seine, Jaulnes and Varennes-sur-Seine have higher δ15N values than Barbuise humans. According to the offsets between the human groups and the relative terrestrial animals, we note that values increase from Barbuise (∆15Nhuman-animal = 3.4‰) to Jaulnes (∆15Nhuman-animal = 3.9‰) and to Barbey (∆15Nhuman-animal = 4.6‰). This enrichment in 15N could be the result of a difference in the omnivore (pig) consumption, whose δ15N and δ13C values are higher as they were fed with human leftovers (Fig. 8). This result may suggest slightly different human dietary patterns based on a diverse animal protein intake, possibly resulting from different livestock strategies on the sites.

The Balloy cemetery for which isotopic data are available dates to the Middle Neolithic (c.4700–3500 BCE) and attributed to the Cerny culture (Chambon 1997). Between the Neolithic and the Bronze Age, climatic changes occurred in western Europe, particularly evident in the Mediterranean area (Magny et al. 2013). Indeed, with the passage from the two consequent climatic periods of Atlantic to the Subboreal, at the end of the Middle Neolithic, there was a generally colder and dryer climate. This resulted in changes to the vegetation cover due to the increase of the agro-pastoral activities that strongly affected the local environment and the broader landscape (Leroyer et al. 2012). Taking into account that environmental changes could have influenced the isotopic values at the bottom of the food web (vegetation cover), comparison between the different periods can only be done using the offsets between humans and the associated animals recovered at the site level. The terrestrial domestic animal assemblages isotopically analysed from the Neolithic and LBA are similar, made up of an analogous number of caprines, cattle and pig, assuring a good representation of all the potential resources consumed by humans (Fig. 8). At Balloy, individuals under 4 years old have higher δ15N values (12.6‰ ± 1.5, n = 8) than the other individuals (10.9‰ ± 10.6, n = 28) (Cheung et al. 2021), likely due to the breastfeeding effect (Fuller et al. 2006; Herrscher et al. 2017). They have therefore been excluded from the following interpretations.
First, Bronze Age humans have more variable δ15N and δ13C values compared to Neolithic humans. This evidence suggests an increase in dietary variability during Late Prehistory, with an isotopic signal typical of resources from a C3 environment during the Neolithic and the EBA and a consumption of mixed C3–C4 resources during the LBA. Second, the ∆15Nhuman-animal and the ∆13Chuman-animal for the Neolithic are, respectively, 3.5‰ and 0.3‰, both lower than the offsets for the LBA, which are 3.9‰ and 2.5‰. Considering only our dataset, the offset is even higher for the nitrogen, corresponding to 4.1‰, suggesting an increase in the animal protein intake from the Neolithic to LBA.
Concerning the carbon offset, the difference between the two periods is the result of C4 plants consumption joined with a diverse omnivore consumption. Indeed, one may note that both Neolithic and LBA pig δ13C ratios exhibit significantly higher values contrasting with the coeval animal groups (Fig. 8). These carbon offsets suggest a low contribution of pig meat to Neolithic human diets in contrast with a differential pig meat consumption for LBA humans correlated with their increasing δ13C values. Furthermore, for the LBA, such an elevated enrichment of 2.5‰ was likely induced by an important consumption of C4 plants, as also previously mentioned.
Human mobility
The high variability of the terrestrial animal δ34S values of the Upper Seine Valley and the other valleys of the Paris Basin (Bocherens et al. 2011; Rey et al. 2019; Cheung et al. 2021) makes it challenging using this technique along to reconstruct human palaeomobility in this area. Nevertheless, thanks to the additional terrestrial animal sulphur data provided by the present study that help define the local sulphur range of this region, we are able to formulate the hypothesis of the presence of non-local individuals that could have originated outside the Upper Seine Valley. Indeed, if the human δ34S values overlap with the local terrestrial animals, they should be reasonably from the same geological area and considered locals. The outcomes will be validated by further strontium analysis currently ongoing.
Combining results for the wild and domesticated terrestrial animals of our dataset (n = 15) with the animals from Noyen-sur-Seine (n = 13; Bocherens et al. 2011) and from Balloy (n = 14; Cheung et al. 2021), we define the local range for the Upper Seine Valley (Figs. 9 and 10). This range, even if large, could be likely considered as the local sulphur baseline for this area. Almost half of the human samples we analysed (44%) fall within this range. The individuals from Marolles-sur-Seine and Varennes-sur-Seine fall within the estimated local range, highlighting a probable local origin of these communities. The only outlier is the infant BP_H23 from Marolles-sur-Seine, whose higher value could indicate this infant is not from the Seine Valley. The strontium analysis will provide further details about this individual (Fig. 9).

Human and animal δ34S values combined with δ13C (A) and δ15N (B) values. The grey area has been estimated considering the mean and standard deviation of all wild and domesticates terrestrial animals available for the Seine Valley (n = 42, data from this study, Bocherens et al. 2011; Cheung et al. 2021). Neolithic humans of the Seine Valley are from Balloy (n = 22, data from Cheung et al. 2021)

Human δ34S values of Marolles-sur-Seine, Barbey, Jaulnes and Varennes-sur-Seine plotted with the local terrestrial animals (mean and standard deviation) divided according to the colour of the geological map of the region obtained from Geoportail (https://www.geoportail.gouv.fr/). Local terrestrial data are from Essonne (Orville, n = 5), Marne (Vignely, n = 16), Yonne (Beaumont and Gurgy, n = 25; Passy and Villeneuve, n = 9) and Seine valleys (this study and Balloy and Noyen-sur Seine, n = 42). Literature data are from Bocherens et al. (2011), Rey et al. (2017, 2019) and Cheung et al. (2021)
At Barbey, the values of the male samples are higher than the female samples and all lie outside the estimated local range. It is likely that the males had a different origin and consumed foods from an area, where the local geological sulphur values are higher than those of the Seine Valley. Interestingly, these δ34S values are consistent with the local animal range of the nearby Essonne (δ34S = 9.7 ± 1.5‰, n = 5, site: Orville) and Marne Valleys (δ34S = 8.2 ± 2.4 ‰, n = 16, site: Vignely) (Cheung et al. 2021) (Fig. 10) whose geological areas are different because they are more recent geological formation, i.e. Oligocene and Eocene formations for Essonne and Marne, respectively. Albeit impossible to infer the place of origin of the individuals, these similarities must be taken into consideration. Moreover, the archaeological evidence seems to support a probable presence of non-local individuals at the Barbey cemetery that distinguishes itself from other funerary groups in the area by its inhumations in a crouched position buried in small rectangular wooden chests or boxes. This unusual funerary practice is documented in only one other cemetery of the Upper Seine Valley located about 40 km east of Barbey at Barbuise (Paris et al. 1976; Rottier et al. 2012).
The two individuals from Jaulnes, with their negative values, are outside the range of possible local values, which could also be due to their non-local origin, even if the hypothesis of a different diet based on local freshwater resources cannot be fully rejected. The nearby Yonne Valley has the lowest δ34S values in the Paris basin (δ34S = 4.9 ± 6.1 ‰, n = 25, sites: Beaumont, Gurgy) (Rey et al. 2017, 2019; Cheung et al. 2021), and the δ34S values from Jaulnes are consistent with those from the Yonne, and it is possible that the individuals originate from there. Further strontium analyses will be undertaken in light of these cautious preliminary interpretations of palaeomobility.
Final remarks and conclusion
This study is a first attempt at reconstructing, through a multi-proxy and multi-isotope approach, the subsistence strategies, dietary behaviours and mobility for the Bronze Age in the southeast of the Paris Basin, which are critical for understanding changes in the global Bronze Age European economy. This area is placed in a strategic position at the frontier between two of the major cultures in western Europe, the Atlantic culture in the West and the Continental culture in the East.
On an archaeological level, this study provides new data that tie in with the occupation dynamics of the Upper Seine Valley observed in various contexts. During the Early and Middle Bronze Age, the area seems to have been quite sparsely populated with only a handful of sites, mainly funerary, that were thinly spread along the valley. The archaeological data show that funerary practices changed very little during this first period with inhumations in a lateral flexed position with no preserved grave goods and rare cremation burials in pottery urns of Atlantic tradition. No structured settlements are known for the EBA and MBA, and the material culture is poorly documented; nevertheless, the few known pottery assemblages and the metallic deposits testify to an Atlantic/Manche-Mer-du-Nord influence (Peake et al. 2017a). A major change occurs around the 14th century BCE (late MBA–early LBA) when the area sees a large and rapid increase in the number of sites, with over 50 funerary contexts and settlements dating to this period. Domestic pits and occupation levels provide a rich and varied array of finds that firmly place the area within the realms of the Continental cultural sphere of northeast France, southwest Germany and the north Alpine area. The increase in the number of sites and the change in cultural affinity suggest a quick and “uncompromising” adoption of new influences brought to the area by an influx of outsiders who settled in the Upper Seine Valley, bringing with them improved agricultural techniques and a new material culture (Peake et al. 2017a). This early dynamic instigates further growth of land occupation in the valley, which becomes the backdrop for the development of a hierarchical organization of communities, comprising of family farms, hamlets and high status settlements during the LBA and the beginning of the Early Iron Age. The preliminary results of this study clearly reflect the profound changes observed in LBA society of the Seine valley and in particular the presence of outsiders who seemed to have been well-established within the local communities. This also supports the idea of the mobility of certain individuals, either temporary or permanent, over short or longer distances who used the Seine Valley to provide the necessary means for the transport of raw materials and manufactured products while allowing the free movement of people.
These preliminary results show a wide variability in δ13C and δ15N values suggesting varying C3 and C4 plant consumption and diverse animal protein intake by humans. Four main outcomes are highlighted: (1) a variety of crop-growing strategies with different high or low levels of fertilization according to the chronological period; (2) site-specific dietary patterns; (3) the introduction of millet consumption at the end of MBA/beginning of the LBA; (4) the increase of omnivore protein intake at the end of the Bronze Age.
Despite the limited isotopic data for the Seine Valley, combining our dataset with available literature, we have been able to detect major changes in the subsistence economy during Late Prehistory. We highlighted that from the Neolithic to the Bronze Age subsistence strategies changed, supported by the introduction of new crops, the consumption of different animals and a general increase in dietary variability. Specifically, we noticed continuity in terms of the subsistence economy between the EBA and the Neolithic, before the major economic changes at the end of the Bronze Age that sees the introduction of new crops and production strategies, very similar to those documented in eastern Europe (Gamarra et al. 2018; Herrscher et al. 2018; Martin et al. 2021). These changes go hand in hand with the intensification of trade in order to supply the development of the first proto-urban societies that likely supported the spreading of new traditions, lifestyle and knowledge, in addition to the different goods and raw materials routinely transported. Particularly, we notice that during the EBA, the human diet was consistent and similar to the Neolithic diet because the main foodstuffs were from a C3 environment with an important animal protein intake. Thus, this evidence indicates that the subsistence strategies in the Seine Valley were oriented towards farming and animal husbandry. This result is also supported by other contemporary EBA sites in Auvergne (Herrscher et al. 2014; Herrscher and Goude 2017) and in the South of France (Le Roy et al. 2021). Dietary behaviours changed at the end of the Bronze Age, and it is evident that (1) the human diet was more varied at both intra- and inter-population levels; (2) millet started to become a staple crop, mainly for human subsistence; (3) pigs likely fed on leftovers, provided a significant source of protein. The impact and the significance of these results call for further investigation not only in the area but also in the nearby Champagne-Ardennes, Alsace and Lorraine regions, which are the main areas of passage between Central Europe and the Upper Seine Valley.
Even though the lack of data for this period highlighted the necessity to develop further investigations in France, similar dietary variations and differential agricultural strategies have been recorded in other coeval sites in western Europe, in Switzerland (Varalli et al. 2021) and in Italy (Varalli et al. 2016a, b, 2020; 2022; Tafuri et al. 2018; Masotti et al. 2019; Goude et al. 2020; Arena et al. 2020) and in eastern Europe, in Hungary (Gamarra et al. 2018) and in the Caucasus (Herrscher et al. 2018; Martin et al. 2021). There is evidence of strong economic ties between these areas and the Continental culture (Kristiansen and Larsson 2005; Cardarelli 2009), driven by the need to supply copper and especially tin ore, as deposits were rare and scattered throughout the continent. This trade was essential to sustain economic development (Earle et al. 2015; Vandkilde 2016; Kristiansen 2018). Future studies should interrogate additional lines of proxy data, including strontium isotope analyses and ancient DNA, to disentangle community relationships and population monument. These data have the potential to speak to our hypothesis about non-local identities and to reconstruct larger mobility events across Europe during the Bronze Age.
Data Availability
The authors confirm that the data supporting the findings of this study are available within the article [and/or] its supplementary materials.
References
Adroit S, Peake R (2021) Entre espace et temps. Regards sur les nécropoles de l’âge du Bronze et du premier âge du Fer à travers des études de cas. In: Marcigny C, Mordant C (eds) Bronze 2019, 20 ans de recherches actes du colloque international de Bayeux, juin 2019, supplément 7 au bulletin de l’APRAB. pp 439–452
Aguilera M, Zech-Matterne V, Lepetz S, Balasse M (2017) Crop fertility conditions in North-Eastern Gaul during the La Tène and Roman periods: a combined stable isotope analysis of archaeobotanical and archaeozoological remains. 23:323–337. https://doi.org/10.1080/14614103.2017.1291563
Ambrose SH (1990) Preparation and characterization of bone and tooth collagen for isotopic analysis. J Archaeol Sci 17:431–451
Ameye N, Delattre V, Peake R, et al (2016) L’ensemble funéraire du Bronze ancien à Mouy-sur-Seine, « Le Grand Gué » (Seine-et-Marne). Bull l’Association pour la Promot des Rech sur l’âge du Bronze 116–122
Arena F, Gualdi-Russo E, Olsen J et al (2020) New data on agro-pastoral diets in southern Italy from the Neolithic to the Bronze Age. Archaeol Anthropol Sci 12:245
Auxiette G, Peake R, Toulemonde F (2015) Food production and diet during the Late Bronze Age in the upper Seine valley ( France ). In: Kneisel J, Dal Corso M, Kirleis W et al (eds) The third food revolution? Setting the Bronze Age Table: Common trends in economic and subsistence strategies in Bronze age Europe. Verlag, Bonn, pp 48–61
Bãlãşescu A, Simonin D, Vigne J-D (2008) La faune du Bronze final IIIb du site fortifié de Boulancourt «le Châtelet» (Seine-et-Marne). 105:371–406
Balasse M (1999) De l’exploitation du lait au Néolithique moyen en Europe tempérée. Examen des modalités de sevrage des bovins par l’analyse isotopique des ossements archéologiques. Université Paris VI
Birkner R (1980) L’image radiologique typique du squelette : aspect normal et variantes chez l’adulte et l’enfant. Pour médecins, étudiante et manipulateurs. Maloine, Paris
Bocherens H, Drucker DG, Taubald H (2011) Preservation of bone collagen sulphur isotopic compositions in an early Holocene river-bank archaeological site. Palaeogeogr Palaeoclimatol Palaeoecol 310:32–38. https://doi.org/10.1016/j.palaeo.2011.05.016
Bocherens H, Fizet M, Mariotti A et al (1991) Application de la biogéochimie isotopique (13C, 15N) à la détermination du régime alimentaire des populations humaines et animales durant les périodes antique et médiévale. Arch Sci Genève 44:329–340
Bogaard A, Fraser R, Heaton THE et al (2013) Crop manuring and intensive land management by Europe’s first farmers. Proc Natl Acad Sci USA 110:12589–12594
Bogaard A, Heaton THE, Poulton P, Merbach I (2007) The impact of manuring on nitrogen isotope ratios in cereals: archaeological implications for reconstruction of diet and crop management practices. J Archaeol Sci 34:335–343. https://doi.org/10.1016/j.jas.2006.04.009
Bronk Ramsey C (2009) Bayesian analysis of radiocarbon dates. Radiocarbon 51:337–360. https://doi.org/10.1017/S0033822200033865
Brunet P, Gouge P, Peake R (2017) La notion d’espace étudié et d’occupation du sol à l’âge du Bronze et au premier âge du Fer dans l’Est de l’Ile-de-France. Bull La Société Archéologique Champenoise 110:229–242
Bruzek J (1991) Fiabilité des procédés de détermination du sexe à partir de l’os coxal Implications à l’étude du dimorphisme sexuel de l’Homme fossile. Ph.D. Thesis, Muséum National d’Histoire Naturelle
Bruzek J, Schmitt A, Murail P (2005) Identification biologique individuelle en paléoanthropologie. Détermination du sexe et estimation de l’âge au décès à partir du squelette. In: Dutour O, Hublin J. J, Vandermeersch B (eds) Objets et méthodes en paléoanthropologie. Paris, pp 217–246
Capuzzo G, Zanon M, Dal Corso M et al (2018) Highly diverse Bronze Age population dynamics in Central-Southern Europe and their response to regional climatic patterns. Plos One 8:e0200709
Cardarelli A (2009) The collapse of the Terramare culture and growth of new economic and social systems during the Late Bronze Age in Italy. Scienze Dell’antichità Stor Archeol Antropol 15:449–520
Chambon P (1997) La nécropole de Balloy “Les Réaudins”. Approche archéoanthropologique. In: La culture de Cerny Nouvelle économie, nouvelle société au Néolithique. APRAIF: Nemours, pp 489–498
Cheung C, Herrscher E, Andre G et al (2021) The grandeur of death – Monuments, societies, and diets in middle Neolithic Paris Basin. J Anthropol Archaeol 63:101332. https://doi.org/10.1016/J.JAA.2021.101332
Christensen BT, Jensen JL, Dong Y, Bogaard A (2022) Manure for millet: Grain δ15N values as indicators of prehistoric cropping intensity of Panicum miliaceum and Setaria italica. J Archaeol Sci 139:105554. https://doi.org/10.1016/J.JAS.2022.105554
Delattre V, Peake R (2012) The role of the individual in the Bronze Age funerary sphere : an archaeo-anthropological reading of data from the Seine Valley (Seine-et-Marne, France). In: Bérenger D, Bourgeois J, Talon M, Wirth S (eds) Gräberlandschaften der Bronzezeit = Paysages funéraires de l’âge du Bronze. Verlag Philipp von Zabern, Darmstadt, pp 295–316
DeNiro MJ (1985) Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to paleodietary reconstruction. Nature 317:806–809
DeNiro MJ, Epstein S (1981) Influence of diet on the distribution of nitrogen isotopes in animals. Geochim Cosmochim Acta 45:341–351
Drucker DG, Valentin F, Thevenet C et al (2018) Aquatic resources in human diet in the Late Mesolithic in Northern France and Luxembourg: insights from carbon, nitrogen and sulphur isotope ratios. Archaeol Anthropol Sci 10:351–368. https://doi.org/10.1007/s12520-016-0356-6
Earle T, Ling J, Uhnér C et al (2015) The political economy and metal trade in Bronze Age Europe: understanding regional variability in terms of comparative advantages and articulations. Eur J Archaeol 18:633–657
Farquhar GD, Hubick HT, Condon AG, Richards RA (1989) Carbon isotope fractionation and water use efficiency. In: Rundel PW, Ehleringer JR, Nagy KA (eds) Stable isotopes in ecological research. Springer-Verlag, New York, pp 22–40
Fernandes R, Millard AR, Brabec M et al (2014) Food reconstruction using isotopic transferred signals (FRUITS): a Bayesian model for diet reconstruction. PLoS One 9:e87436. https://doi.org/10.1371/JOURNAL.PONE.0087436
Ferrio JP, Araus JL, Buxó R et al (2005) Water management practices and climate in ancient agriculture: Inferences from the stable isotope composition of archaeobotanical remains. Veg Hist Archaeobot 14:510–517
Fraser RA, Bogaard A, Charles M et al (2013) Assessing natural variation and the effects of charring, burial and pre-treatment on the stable carbon and nitrogen isotope values of archaeobotanical cereals and pulses. J Archaeol Sci 40:4754–4766. https://doi.org/10.1016/j.jas.2013.01.032
Fraser RA, Bogaard A, Heaton T et al (2011) Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices. J Archaeol Sci 38:2790–2804. https://doi.org/10.1016/j.jas.2011.06.024
Fuller BT, Molleson TI, Harris D, a, et al (2006) Isotopic evidence for breastfeeding and possible adult dietary differences from Late/Sub-Roman Britain. Am J Phys Anthropol 129:45–54. https://doi.org/10.1002/ajpa.20244
Gamarra B, Howcroft R, McCall A et al (2018) 5000 years of dietary variations of prehistoric farmers in the Great Hungarian Plain. PLoS One 13:e0197214
Goude G, Dori I, Sparacello VS et al (2020) Multi-proxy stable isotope analyses of dentine microsections reveal diachronic changes in life history adaptations, mobility, and tuberculosis-induced wasting in prehistoric Liguria (Finale Ligure, Italy, northwestern Mediterranean). Int J Paleopathol 28:99–11. https://doi.org/10.1016/j.ijpp.2019.12.007
Goude G, Rey L, Toulemonde F et al (2017) Dietary changes and millet consumption in northern France at the end of Prehistory: evidence from archaeobotanical and stable isotope data. Environ Archaeol 22:268–282
Gouge P, Peake R (2005) Aux marges du Bronze atlantique, sites et chronologie de la région du confluent Seine-Yonne. In: Bourgeois L, Talon M (eds) L’âge du Bronze dans le nord de la France dans son contexte européen. CTHS, Paris, pp 333–359
Hedges REM, Clement JG, Thomas CDL, O’connell TC (2007) Collagen turnover in the adult femoral mid-shaft: modeled from anthropogenic radiocarbon tracer measurements. Am J Phys Anthropol 133:808–816
Hermetey C (1995) Potentialités informatives des petits assemblages de faune de l’Age du Bronze: l’exemple de la Bassée, Seine-et-Marne (IXème- VIIème siècle avant J.-C.). PhD Thesis, Univ. Paris I, Panthéon-Sorbonne
Herrscher E, Goude G (2017) Impact des facteurs socio-culturels sur l’alimentation: confrontation des données isotopiques, biologiques et archéologiques. In: Lisfranc R, Vital J (eds) La nécropole Bronze ancien de Gerzat, Chantemerle (Puy-de-Dôme). ALPARA, pp 394–406
Herrscher E, Goude G, Lisfranc R et al (2014) Pratiques alimentaires au Bronze ancien en Auvergne: Essai de restitution par l’analyse isotopique. Préhistoires Méditerranéennes 4:2003
Herrscher E, Goude G, Metz L (2017) Longitudinal study of stable isotope compositions of maternal milk and implications for the palaeodiet of infants. Bull Mem Soc Anthropol Paris 29:131–139
Herrscher E, Poulmarc’h M, Pecqueur L et al (2018) Dietary inferences through stable isotope analysis at the Neolithic and Bronze Age in the southern Caucasus (sixth to first millenium BC, Azerbaijan): from environmental adaptation to social impacts. Am J Phys Anthropol 167:856–875
Jeune YL, Leroyer C, Pastre J (2012) Lʼévolution holocène de la basse vallée de la Marne ( Bassin parisien, France ) entre influences climatiques et anthropiques Holocene evolution of the lower Marne valley ( Paris basin, France ): mixed climatic and anthropogenic influences. Géomorphologie Reli Process Environ 4:459–476. https://doi.org/10.4000/geomorphologie.10060
Katzenberg MA (2008) Stable isotope analysis: a tool of studying past diet, demography, and life history. In: Katzenberg M, Saunders S (eds) Biological Anthropology of the Human Skeleton. John Wiley & Sons Inc, Hoboken, NJ, pp 413–441
Kneisel J, Dal Corso M, Kirleis W et al (2015) The third food revolution? Common trends in economic and subsistence strategies in Bronze Age Europe. In: Kneisel J, Dal Corso M, Kirleis W et al (eds) The third food revolution? Setting the Bronze Age Table: Common trends in economic and subsistence strategies in Bronze age Europe. Universitätsforschungen zur prähistorischen Archäologie, Bonn, pp 257–286
Knipper C, Meyer C, Jacobi F et al (2014) Social differentiation and land use at an Early Iron Age “princely seat”: bioarchaeological investigations at the Glauberg (Germany). J Archaeol Sci 41:818–835
Körner C, Farquhar GD, Wong SC (1991) Carbon isotope discrimination by plants follows latitudinal and altitudinal trends. Oecologia 88:30–40
Kristiansen K (2018) Warfare and the political economy: Europe 1500–1100 bc. In: Horn C, Kristiansen K (eds) Warfare in Bronze Age Society. Cambridges University Press, Cambridge, pp 23–46
Kristiansen K, Larsson TB (2005) The rise of Bronze Age society: travels, transmissions and transformations. Cambridge University Press
Lachenal T, Mordant C, Nicolas T, Veber C (2017) Le Bronze moyen et l’origine du Bronze final en Europe occidentale, XVIIe-XIIIe siècle avant notre ère. In: Actes du Colloque international APRAB, Strasbourg 17–20 Juin 2014. Mémoire d’Archéologie du Grand Est: Strasbourg
Le Goff C (2008) La faune. In: Valero C (ed) Grez-sur-Loing «L’Epine»Seine-et-Marne, Vestiges du Néolithique, habitats del’âge du bronze final. INRAP, Saint-Denis: Service Régional de l’Archéologie d’Ile-de-France, pp 29–41
Le Goff I, Peake R (2021) Inhumation versus incinération ou incinération versus inhumation? Dualité du traitement du corps à l’âge du Bronze en France. In: Marcigny C, Mordant C (eds) Bronze 2019, 20 ans de recherches actes du colloque international de Bayeux, juin 2019, supplément 7 au bulletin de l’APRAB. pp 423–438
Le Roy M, Magniez P, Goude | Gwenaëlle, et al (2021) Stable-isotope analysis of collective burial sites in Southern France at late Neolithic/early Bronze Age transition. Int J Osteoarchaeol. https://doi.org/10.1002/OA.3074
Leroyer C, David R, Mazier F, et al (2012) Environnement et anthropisation du milieu durant l’âge du Bronze dans le Bassin parisien: l’apport des données polliniques et de la modélisation du couvert végétal. In: Melin M, Mougne C (eds) L’Homme, ses ressources et son environnement, dans le Nord-Ouest de la France à l’âge du Bronze: actualités de la recherche. Géosciences Rennes
Longin R (1971) New Method of Collagen Extraction for Radiocarbon Dating. Nature 230:241–242
Magny M, Bossuet G, Gauthier E et al (2007) Variation du climat pendnat l’Age du Bronze au centre-ouest de l’Europe: vers l’établissement d’une chronologie à haute résolution. In: Richard H, Magny M, Mordant C (eds) Environnements et cultures à l’Age du Bronze en Europe Occidentale. CTHS, Paris, pp 13–28
Magny M, Combourieu-Nebout N, De BJL et al (2013) North-south palaeohydrological contrasts in the central Mediterranean during the Holocene: tentative synthesis and working hypotheses. Clim past Discuss 9:1901–1967
Martin L, Messager E, Bedianashvili G et al (2021) The place of millet in food globalization during Late Prehistory as evidenced by new bioarchaeological data from the Caucasus. Sci Rep 11:13124. https://doi.org/10.1038/s41598-021-92392-9
Masotti S, Varalli A, Goude G et al (2019) A combined analysis of dietary habits in the Bronze Age site of Ballabio (northern Italy). Archaeol Anthropol Sci 11:1029–1047
Minagawa M, Wada E (1984) Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochim Cosmochim Acta 48:1135–1140
Mordant C (1989) Transgression culturelle et mouvements de population aux XIVe- XIIIe siècles avant notre ère dans le Bassin parisien, compétition culturelle et phénomène de lisière. Dynamique du Bronze moyen en Europe occidentale, Actes du Colloque de Strasbourg-Haguenau, 113e Congrès national des Sociétés savantes Strasbourg 1988. CTHS, Paris, pp 283–303
Mordant C, Peake R, Roscio M (2021) Weighing equipment in Late Bronze Age graves in the Seine and Yonne valleys. In: Rahmstorf L, Barjamovic G, Ialongo N (eds) Merchants, Measures and Money. Understanding technologies of early trade in a comparative perspective. Proceedings of two Workshops funded by the European Research Council (ERC), pp 159–172
Mordant C, Roscio M (2010) Variabilité des pratiques funéraires à la fin du Bronze moyen/Bronze final initial (XIVe-XIIIe s. av. J.-C.) en France orientale, de l’Ile-de-France à l’Alsace. In: Studia Celtica Classica et Romana Nicolae Szabo Septuagesismo Dedicata. Pyhtéas, Budapest, pp 169–192
Naito YI, Chikaraishi Y, Ohkouchi N et al (2013) Nitrogen isotopic composition of collagen amino acids as an indicator of aquatic resource consumption: insights from Mesolithic and Epipalaeolithic archaeological sites in France. World Archaeol 45:338–359. https://doi.org/10.1080/00438243.2013.820650
Nehlich O (2015) The application of sulphur isotope analyses in archaeological research: a review. Earth-Science Rev 142:1–17
Nehlich O, Richards MP (2009) Establishing collagen quality criteria for sulphur isotope analysis of archaeological bone collagen. Archaeol Anthropol Sci 1:59–75
Nitsch EK, Charles M, Bogaard A (2015) Calculating a statistically robust δ 13 C and δ 15 N offset for charred cereal and pulse seeds. Sci Technol Archaeol Res 1:STAR20152054892315Y.0000000001
Paris J, Mordant C, Mordant D, Bontillot J (1976) La nécropole du Bronze final des Cents Arpents à Barbey (Seine-et-Marne). Montereau-fault-Yonne et Bray-sur-Seine
Peake R (2020) Villiers-sur-Seine, un habitat aristocratique entre deux rives du 9e siècle avant notre ère. Inrap-CNRS
Peake R, Delattre V (2010) Monumentalité de la mort : la nécropole diachronique de Jaulnes « Le Bas des Hauts Champs » (Seine-et-Marne). Bull L’association Pour La Promot Des Rech Sur L’âge Du Bronze 7:19–22
Peake R, Delattre V (2005) L’apport des analyses 14C à l’étude de la nécropole de l’âge du Bronze de Marolles-sur-Seine. La Croix de la Mission. Rev Archeol Centre France 44:5–25
Peake R, Delattre V, Le Goff I, Billand G (2017a) Lecture des pratiques funéraires du Bronze moyen et du début du Bronze final dans le Centre-Nord de la France. In: Lachenal T, Mordant C, Nicolas T, Véber C (eds) Le Bronze moyen et l’origine du Bronze final en Europe occidentale, de la Méditerranée aux pays nordiques (XVIIe-XIIIe s. av. J.-C.), actes du colloque international de Strasbourg, APRAB, 2014. Monographies d’Archéologie du Grand Est, pp 763–786
Peake R, Roscio M, Issenmann R, et al (2017b) Le Bronze moyen et le début du Bronze final dans le sud du Bassin parisien. In: Le Bronze moyen et l’origine du Bronze final en Europe occidentale, de la Méditerranée aux pays nordiques (XVIIe-XIIIe s. av. J.-C.), actes du colloque international de Strasbourg, APRAB, 2014. Monographies d’Archéologie du Grand Est, Strasbourg, pp 193–214
R Core Team (2018) R vesion 3.6.1 (R foundation for Statistical Computing). Vienna, Austria
R Development Core Team (2005) A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria: ISBN 3–900051–07–0, URL: http://www.R-project.org
R Studio Team (2020) RStudio: Integrated Development for R. RStudio. Boston, MA URL http://www.rstudio.com/
Reimer PJ, Austin WEN, Bard E et al (2020) The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62:725–757. https://doi.org/10.1017/RDC.2020.41
Rey L, Goude G, Rottier S (2017) Comportements alimentaires au Néolithique : nouveaux résultats dans le Bassin parisien à partir de l’étude isotopique (δ13C, δ15N) de la nécropole de Gurgy « Les Noisats » (Yonne, Ve millénaire av. J.-C.). Bull Mem Soc Anthropol Paris 29:54–69. https://doi.org/10.1007/S13219-016-0170-9
Rey L, Salazar-García DC, Chambon P, h, et al (2019) A multi-isotope analysis of Neolithic human groups in the Yonne valley, Northern France: insights into dietary patterns and social structure. Archaeol Anthropol Sci 11:5591–5616. https://doi.org/10.1007/s12520-019-00885-6
Roscio M (2018) Les nécropoles de l’étape ancienne du Bronze final du Bassin parisien au Jura souabe (XIVe- XIIe siècle avant notre ère). Editions universitaires de Dijon, Dijon
Rottier S, Piette J, Mordant C (2012) Archéologie funéraire du Bronze final dans les vallées de l’Yonne et de la haute Seine : Les nécropoles de Barbey. Editions universitaires de Dijon, Dijon, Barbuise et La Saulsotte
Scheuer L, Black S (2000) Developmental juvenile osteology. Elsevier Academic Press, Oxford
Schmitt A (2008) Une nouvelle méthode pour discriminer les individus décédés avant ou après 40 ans à partir de la symphyse pubienne. J Médecine Légale Droit Médical
Schoeninger MJ, DeNiro MJ (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Acta 48:625–639
Styring AK, Charles M, Fantone F et al (2017) Isotope evidence for agricultural extensification reveals how the world’s first cities were fed. Nat Plants 3:17076
Styring AK, Fraser RA, Bogaard A, Evershed RP (2014) Cereal grain, rachis and pulse seed amino acid δ15N values as indicators of plant nitrogen metabolism. Phytochemistry 97. https://doi.org/10.1016/j.phytochem.2013.05.009
Szpak P, Metcalfe JZ, Macdonald RA (2017) Best practices for calibrating and reporting stable isotope measurements in archaeology. J Archaeol Sci Reports 13:609–616. https://doi.org/10.1016/J.JASREP.2017.05.007
Tafuri MA, Rottoli M, Cupito M et al (2018) Estimating C4 plant consumption in Bronze Age Northeastern Italy through stable carbon and nitrogen isotopes in bone collagen. Int J Osteoarchaeol 28:131–142
Toulemonde F (2013) Économie végétale et pratiques agricoles au Bronze final et au premier âge du Fer, de la côte de l’Île-de-France à la côte de Champagne. Université de Paris 1
Toulemonde F (2010) L’alimentation végétale durant la Protohistoire ancienne en Île-de- France. Étude carpologique des sites de Gif-sur- Yvette « Rond-point de Corbeville » (Essonne) et Villiers-sur-Seine « Le Gros Buisson » (Seine-et- Marne). Rev Archéologique D’île-De-France 3:253–272
Toulemonde F, Auxiette G, Bouby L, et al (2018) L’alimentation à l’âge du Bronze en France. In: Guilaine J, Garcia D (eds) La protohistoire de la France. Hermann, pp 297–309
Toulemonde F, Wiethold J, Bonnaire E, et al (2022) Millets in Bronze Age agriculture and food consumption in north-eastern France. In: Kirleis W, Dal Corso M, Filipović D (eds) Millet and what else? The wider context of the adoption of millet cultivation in Europe. Scales of transformation in Prehistoric and Archaic Societies 14, Sidestone
Toulemonde F, Zech-Matterne V, Wiethold J, et al (2017) Reconstitution des pratiques agricoles du Ier millénaire a.C. en France orientale, d’après le croisement des données carpologiques et archéologiques. In: Marion S, Deffressigne S, Kaurin J, Bataille G (eds) Production et proto-industrialisation aux âges du Fer. Perspectives sociales et environnementales, Actes du 39e colloque international de l’Association Française pour l’Étude de l’Âge du Fer (Nancy 14–17 mai 2015). pp 29–50
Ubelaker DH (1987) Estimating age at death from immature human skeleton: an overview. J Forensic Sci 32:1254–1263
Vaiglova P, Bogaard A, Collins M et al (2014) An integrated stable isotope study of plants and animals from Kouphovouno, southern Greece: a new look at Neolithic farming. J Archaeol Sci 42:201–215
van Klinken GJ (1999) Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J Archaeol Sci 26:687–695
Vandkilde H (2016) Bronzization: the bronze age as pre-modern globalization. Prahistorische Zeitschrift 91:103–123
Varalli A, Desideri J, David-Elbiali M et al (2021) Bronze Age innovations and impact on human diet: a multi-isotopic and multi-proxy study of western Switzerland. PLoS One 16:e0245726. https://doi.org/10.1371/journal.pone.0245726
Varalli A, Moggi-Cecchi J, Goude G (2022) A multi-proxy bioarchaeological approach reveals new trends in Bronze Age diet in Italy. Sci Rep 121(12):1–20. https://doi.org/10.1038/s41598-022-15581-0
Varalli A, Moggi-Cecchi J, Dori I et al (2016a) Dietary continuity vs. discontinuity in Bronze Age Italy. The isotopic evidence from Arano di Cellore (Illasi, Verona, Italy). J Archaeol Sci Rep 7:104–113. https://doi.org/10.1016/j.jasrep.2016.03.047
Varalli A, Moggi-Cecchi J, Moroni A, Goude G (2016b) Dietary variability during Bronze Age in Central Italy: first results. Int J Osteoarchaeol 26:431–446. https://doi.org/10.1002/oa.2434
Varalli A, Villotte S, Dori I, Sparacello V (2020) New insights into activity-related functional bone adaptations and alterations in Neolithic Liguria (Northwestern Italy). BMSAP 32:34–58. https://doi.org/10.3166/bmsap-2020-0072
Vika E (2009) Strangers in the grave? Investigating local provenance in a Greek Bronze Age mass burial using d34S analysis. J Archaeol Sci 36:2024–2028
Wallace M, Jones G, Charles M et al (2013) Stable carbon isotope analysis as a direct means of inferring crop water status and water management practices. World Archaeol 45:388–409
Acknowledgements
The authors would like to thank Leia Mion (LAMPEA, Aix-Marseille University) for their help with the isotopic analyses. Isotopic measurements were supervised by Steve Brooks and Ian Begley at Iso-Analytical Ltd (Crew, Cheshire, UK).
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This work was funded by the Ile-de-France regional council and the Institut National de Recherches Archéologiques Préventives (Inrap) as a multidisciplinary research program “PCR Bronz’Pal”.
Author information
Authors and Affiliations
Contributions
Conceptualization, AV, EH, RP; data curation, AV, EH, RP; formal analysis, AV, EH; funding, RP; resources, EH; investigation, GA; writing—original draft, AV, RP, EH; writing—review and editing, AV, RP, EH, CM, VD, VB, GA, PG, MR, FT.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Varalli, A., Peake, R., Auxiette, G. et al. Insights into the frontier zone of Upper Seine Valley (France) during the Bronze Age through subsistence strategies and dietary patterns. Archaeol Anthropol Sci 15, 26 (2023). https://doi.org/10.1007/s12520-023-01721-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-023-01721-8