
Abstract
This meta-study of animal semantics is anchored in two claims, seemingly creating a fuzzy mismatch, that animal utterances generally appear to be simple in structure and content variation and that animals’ communicative understanding seems disproportionally more advanced. A set of excerpted, new studies is chosen as basis to discuss whether the semantics of animal uttering and understanding can be fused into one. Studies are prioritised due to their relatively complex designs, giving priority to dynamics between syntax, semantics, pragmatics, and between utterers and receivers in context. A communicational framework based on utterance theory is applied as a lens for inspection of how these aspects relate to the assumed mismatch. Inspection and discussions of the studies bring several features to surface of which five are stressed in the following. Firstly, both syntactic structures and possible semantic content are seen as lean, although richer than earlier believed, and research continues to reveal new complexities in utterances. Secondly, there is a clear willingness to broaden the perception of animals’ semantic capacity to comprehend communication both by arguing theoretically and by generating empirical research in new contexts. Thirdly, the ambition to make sense of these tendencies is still often motivated by an evolutionary search for early building blocks for verbal language, with the pro et cons that such a position can have. Fourthly, the ‘allowed’ scientific frame for studying semantic capacity among animals is extended to new fields and contexts challenging the only-in-the-wild norm. Fifthly, the dilemma of integrating uttering and understanding as aspects of an after all functional communicational system, calls for new epistemological concepts to make sense of the claimed mismatch. Affordances, abduction, life-genre, and lifeworld are suggested.
Similar content being viewed by others


Introduction
Match and/or mismatch between semantic capacities?
It might be an illusionary effect of language that there exists a prevalent conception of a functional, fairly balanced match between uttering and understanding, in spite of the common insight that there often occurs a mismatch between what for instance toddlers, brain-damaged people, foreigners, blind and deaf people can utter and what they can understand. Regarding animals, and great apes in particular, the idea that they by learning to ‘speak’ verbal language could increase their capacity to utter, and hence their active semantic repertoire, has now mostly been given up (Terrace, 2019). One thing is that chimpanzees’ oral organs do not seem well fitted for sounds that make up our phonemes. Another recognition is that even among our closest relatives there seem to be an imbalance, a semantic mismatch, between what orally can be uttered and what can be understood: “The core argument is that, just as a person’s utterances reveal only a subset of what they know, animal communication signals express an intrinsically limited subset of that species’ conceptual storehouse” (Fitch, 2019, p. 2).
Hence, seen from the perspective of receivers, Fischer (2017, p. 76) holds that it is important to focus on to what degree animal calls can be used to make inferences about happenings in concrete contexts. She too is preoccupied with the assumed mismatch and wishes future research for instance could ask: “[…] how do nonhuman primates get away with such limited repertoires, although they have such rich representations of the world?” (Fischer, 2017, p.76). An aim for this meta-study is to investigate this claimed fuzziness.
A strong imbalance raises a crucial question for any ‘overall’ communicational system for a specific taxon in the animal world: To which degree and in which sense might a sub-system for uttering, in our case sending, and a sub-system for interpreting, in our case receiving, be both different and the same? A hypothesis is implied in the question. The visual shape of letter Y, seen as a tree, can model a simplistic answer – it is both/and, neither just the same nor just different: The trunk of the Y- may illustrate what is shared, the two branches what differs. Verbal communication might have a relatively long trunk, meaning that quite much of what is uttered can be understood. Dogs might have a short one due to a mismatch between the semantics of urinating and sniffing, just to hint disproportional imbalances of what can be uttered and what can be interpreted. The point here is not just to focus the mismatch as such, but rather to investigate, based on the assumption that a communicational system incorporating the both/and perspective, might exist, and, at the end of the day, be fairly functional despite that most animals’ repertoire for expressing/uttering is seen as stereotyped and simplistic: “Studies on the variation in calling between different species of the same genus, or between subspecies, also suggest that the structure of nonhuman primate calls is highly conserved” (Fischer, 2017, p. 75). Fischer further concludes:
Taken together, nonhuman primates do not have the capacity (nor necessity) to acquire their vocalizations; and hence they do not meet the minimal preconditions for developing a socially transmitted conventionalized communication system. (Fischer, 2017, p. 75).
In other words, given that there nevertheless might exist some kind of communicational system for any species – what are the main patterns of its parts and how could it be conceptualised as a whole?
There are recent studies, now more often based on more advanced methods, of different taxa’s repertoire for uttering, such as Marconi et al. (2020) on mice’s ultrasonic communication, Fedurek, Zuberbühler, and Dahl (2016) on chimpanzees’ pant hoot calls, and Prior et al. (2018) on finches’ song repertoire. Documentation of more differentiated patterns has accordingly led to an increased interest in possible semantic implications of these more variable patterns for ‘both’ sub-systems, that is, uttering and interpreting, which in turn has contributed to an increased interest in animal cognition (Fitch, 2019), and hence content and semantics. Both is her put in inverted commas to make aware that the two are even seen as one.
This paper will not discuss semantic or syntactic compositionality (Gabrić, 2021; Suzuki, 2021), although relevant for a discussion of content richness in animal utterances. There is much research on supposedly differentiated and more advanced structuring of vocal forms, especially on non-human primates and birds (Suzuki, 2021; Townsend et al., 2018). It seems premature though to conclude on implications for the semanticity of animals in general. Regarding bird song Beecher (2021, p. 1) concludes that although the song seems “[…]nuanced and complex, and has the acoustic potential for productivity, it is not productive – it cannot be used to say many different things”. In any case, as several studies have pointed out, possible content elements in animal utterances need to be correlated with their documented impact on receivers’ reactions.
Clarifications of key concepts
Studying potential animal semanticities, their capacities, from a meta-position, necessitates some principal, conceptual clarifications, epistemological positionings, and methodological reflexions. Below I clarify what could be meant by semiotic semantics, capacities, and contexts.
In 1985 Barbieri published The Semantic Theory of Evolution (Barbieri, 1985) where the term semantics was given almost the same meaning as in “a semantic theory of language”. In 2020 he argues that the purpose was to show that semantic mechanisms are at work in living systems: “The semantic theory […] tells us that the rules of language are generated by human societies, not by individual human beings, and that they are acquired by children with interpretive neural networks that we have inherited from our animal ancestors” (Barbieri, 2020, p. 3). Hence, he holds that semantics plays a crucial role in the evolutionary line between related species, even in the case of the final leap to Homo sapiens. In addition he hints that semantics has to be seen in a social, and therefore systemic perspective.
Any semantics should take into consideration semantic cooperation between levels. While Barbieri’s interests primarily concern the lower, the micro-levels of life, such as cells, my focus in this study is mainly on macro levels, that is, more general kinds of communication, such as utterances, life-genres, and lifeworlds. My scope is therefore even wider than the critical and clarifying work on animal semantics done by for instance Fitch (2016), Manser (2013), Wheeler and Fischer (2012), Fischer (2017), Lyn (2017), and Lyn and Christopher (2020). In Ongstad (2021a) I have given a critical review of key concepts applied in recent studies of animal semantics. One of the conclusions is that important disagreements over the scientific functionality of crucial semantic terms seem rooted in designs generated in somewhat restricted communicational models. Nevertheless, some few newer studies did apply a more open scope and more complex methodologies, implicating both a more comprehensive view of animal communication and hence a semiotic semantics as well as a more systemic view on communication. This current meta-study will therefore focus both on the semantic content and content as related to form, function, and on context, which implies interrelating semantic studies critically with semiotic studies of syntax, pragmatics, and context theory. It further means prioritising higher communicational levels, such as utterance, genre, and life-world/Umwelt at the dispense of signs, in short a more macro-oriented systemic perspective.
Next, the study is focused on animal semantic capacities. Epistemologically, studying capacity may imply a break with the more purist approach to study animals in the wild only, in a species’ so-called natural habitat. By including studies from other contexts I extend the scope of the study from what animals actually do, to what they might be able to comprehend, semantically in new environments. Therefore, studies from contexts other than animal communication in the wild will be inspected, such as Lyn and Christopher (2020), a work titled How Environment Can Reveal Semantic Capacities in Nonhuman Animals where they argue:
Frequently dismissed or ignored in the discussion of the usefulness of animal communication to the study of human language evolution is the wealth of data from the language research projects that show remarkable levels of symbolic behavior in animals reared in enriched, enculturated environments. These data show that the capacity for semantic communication systems is not unique to humans (Lyn and Christopher, 2020, p. 159).
Yet, their point of departure differs somewhat from mine on a crucial point, as they, as quite a few others, discuss findings in the light of animal semanticity as a possible origin of human verbality. I do not. There is no search for a missing link. The strong drive behind hypotheses about and empirical search for connections between animal vocalities and human language is understandable though. However, it seems worth listening to a recent conclusion from a key figure in this search for evolutionary clues:
In summary, nonhuman primate communication shares fewer similarities with human speech than many scholars may have hoped for, but we would never have learned so much about primate vocal communication were it not for the breakthrough study by Seyfarth, Cheney, and Marler (Fischer, 2020, p. 108).
Since my view is not linguistic, but rather semiotic, it implies a broader perspective. A broad, but not explicitly semiotic conception of semantics is found in Dezecache and Berthet (2018, p. 5):
Animal vocalizations possess a semantics in the simple sense that one can potentially identify a set of circumstances in which a given call is appropriate or often observed. Thus, calls need not correspond to a natural class of objects in the world (like “leopard” or “terrestrial predator”) to possess a semantics.
This more general definition should imply a farewell to a linguistic epistemology in favour of a semiotic one. Regarding concepts, semantics here means the branch of semiotics that deals with meanings that particular signs can have for any user (Morris, 1938). This wide definition ranges, on the one hand, from the weakest connections between structured form and content, between for instance a particular zebra’s stripe-pattern which other zebras might see as a sign for the specific other, to, on the other hand, a full-fledged arbitrary, semiotic symbol (Deacon, 2013; Peirce, 1992). The perspective implicates that, according to Morris (1938) semantics is seen as a key constitutive semiotic aspect, along with syntax, studying forms, and pragmatics, studying acts. Semantics is not just associated with human, verbal language and thus linguistics and human psychology (Jackendoff, 1983). It would embrace various kinds of signs, such as icons, indexes, and symbols, to use Peirce’s notions (Peirce, 1992). With Bühler (1934) this view would include the sign, not only as a referential symbol but even as a symptom of an utterer’s inner state and as a signal to others. In the utterance “Ruuun!”the word’s extended, enforced form, its expressivity, is a symptom of the utterer’s inner state. “Run” refers semantically to an activity and is thus a symbol. The exclamation mark hints an imperative and works hence as an addressed signal to receivers.
Although this meta-study does not search a missing link to language, the overall, main project which this sub-study is a part of, nevertheless does have an evolutionary perspective (Ongstad, 2019). It is assumed that there might exist a connection between animals’ and humans’ ability to understand broad, basic kinds of communication (life-genres), as they share some basic life-functions and thus even some general patterns of communication serving these functions (Ongstad, 2019). This sub-study will restrict the use of empirical studies of the semantics to vertebrates. Still, according to a biosemiotic perspective ‘semantic’ pre-elements might already have been established at an early evolutionary state (Barbieri, 2020, Dittrich, 2018). We know, for instance, that non-vertebrates such as slime fungus and octopuses have remarkable ‘cognitive’ capacities (Jabr and Rothschild, 2012; Godfrey-Smith, 2016). This argument implies that concepts such as semantics, mind, meaning, mentality, and cognition are seen as closely related, both in a synchronic and a diachronic, evolutionary, perspective (Zlatev, 2012).
For the time being a general evolutionary hypothesis is, that when verbal language as an additional semiotic system emerged in the first place, probably less than a couple of hundred thousands years ago (Perreault and Mathew, 2012), it could only work as a system based on more general premisses of communication evolution already had established through in the first place (Tomasello, 2010). In other words, non-human animals and humans are believed to share the ability to utter and interpret basic kinds of utterances, what I have called life-genres, in a variety of contexts as parts of an integrated system. In Ongstad (2021a), an investigation of recent semantic studies, it is therefore suggested that a move of research interests from uttering to receiving should be followed even by a focus on integration and interplay, that is, on the nature of a species’ communicational system as a whole (Fuller, 2013). An implication would be to include empirical studies of semantic capacities of animals in other circumstances and environments, such as domesticated animals, dogs in particular (Pilley and Reid, 2011; Fugazza et al. 2012).
This extension finally turns focus to the third concept, which is context. A simple hypothesis is that a species’ capacity to ‘passively’ extract meaning from contexts is far more advanced than expected and can compensate for a species’ restricted repertoire to actively utter. This position necessitates a deeper theoretical understanding of animals’ communicational contexts that can be sufficiently operationalised. It further asks for developing more specific theories of how or to which degree animal sensing and storing of impressions (input) eventually can lead to concepts, if at all. The applied framework, presented in next section, will see context as a dynamic set of aspects that are placed in the background by the very focus an utterance generates (Ongstad, 2021a, 2021b). In this sense an utterer, for instance using an alarm call, creates the communicational context. However, although the utterer and the receiver share some contextual aspects, they do not find themselves in the exact same context. A context is considered as dynamic and, although partly shared, even personal. Hence, it is in addition situational and episodic (Ongstad, 2021b).
Chosen empirical studies, applied theory, and approach
Excerpting newer studies of semantic capacities
What is at stake is animal semantics as such, how it is conceptualised and researched. Meta-studies seem to agree on a need for more research in this subfield, among other reasons, due to lack of common scientific views and shared concepts (Manser, 2013; Ongstad, 2021a). In Ongstad (2021a), which focused animals’ ability to refer, two underlying patterns came to surface.
First, project-designs were found mostly dyadic, prioritising form/content, content/function, or function/form, and rarely a triad of form/content/function. While such dyadic approaches might increase the chance for presenting well documented, ‘proofed’ results, the number of unsolved epistemological questions seem to increase. Triadic, or more complex and broad study designs, integrating several key communicational aspects, seem to achieve more holistic, and thus systemic, understanding of animal semantics, but risked being more dependent on interpretation.
Second, some recent studies that favor more complex designs, in fact focused less on references, the meaning as such and more on types of communication, semantic levels, and context. I have already mentioned in this respect Fedurek, Zuberbühler, and Dahl (2016). This work seems characteristic for a quite new trend allowing aspects of animal communication of particular species to be studied in a broader as well as a more integrative perspective. They examined the complex chimpanzee (Pan troglodytes schweinfurthii) ‘pant hoot’ call assuming that a reason for its complexity is that receivers may gain different types of information from a single call, what they term the multiple information hypothesis. Concluding, they found their detailed empirical study consistent with the hypothesis:
By virtue of being a complex, multi-phased call, different phases of the call seem to co-vary differently with specific attributes of the caller (such as identity, age, or social status) or with context of call production. This consistent covariance might then allow receivers to make decisions about subsequent responses (Fedurek, Zuberbühler, and Dahl, 2016, p. 7).
Hence, although animals’ registers for uttering seem lean, they might be richer than first believed. I have searched for recent studies, roughly from the last decade, which in some sense deal with or can help enlighten the mismatch dilemma and have ended up with a set of relevant core studies. I present them briefly below and flag issues that seem important for the inquiry.
Fitch (2019) discusses whether animals have concepts. He studies evolutionary lines for human language, claiming that one cannot focus solely on communication. Animals’ cognitive capacities should be brought into picture too. While Fitch here seems to keep communication and cognition conceptually separated, the framework that is applied places utterance at the centre, and cognitive-referential elements are seen as more integrated, (Bakhtin, 1986; Ongstad, 2019).
Dezecache and Berthet (2018) review studies on perceptions of alarm calls. Their meta-study or review, on the meaning of general alarm calls seems appropriate for my critical discussion of key terms used in the field, such as call, versus utterance, and alarm call, versus life-genre.
Kalkman (2019) discusses varieties of meaning and their role in explaining communication both among animals and humans, while placing himself on the information-side of the semantic debate. He finds it necessary to include all kinds of content, that is, semantic concepts and references, and study how these different kinds relate to their possible functionality for receivers. A study of this relation is of course relevant to the mismatch-dilemma.
Lyn and Christopher (2020), building partly on Lyn (2017) argue that other domains should be investigated to trace animal semantic capacities. Earlier Lyn (2012) had summarised, systematically, 40 years of research on apes and language, in which semantics was one of the key investigated topics. In this context I see these three works partly as a whole since they move from an interest in content element of other primates’ language toward animal semantic capacity more in general. As for me, this extension leads to the inclusion of studies of animals in non-wild contexts.
Fischer (2017) is preoccupied with how specific traits of animal communication relate to the evolution of human, verbal language. She dwells on the relationship between animal semantics and newer, more detailed studies of the structure of male vervet vocalisations, to understand how perceptions of environment is linked to calls. As pointed to above, she besides hopes that further research will deal with the puzzling mismatch between apes’ limited vocal repertoire and their assumed rich representation of their world. It seems natural in this connection also to touch upon Fischer (2020), entitled Nonhuman Primate Alarm Calls Then and Now.
Fuller (2013) is chosen not only because he consequently applies an explicit triadic design by combining form, content, and function in the title of his thesis. He even insists on including as much as possible of a species’ (vocal) total communicative repertoire. In other words he applies a full systemic view.
Prior et al. (2018), study zebra finches’ calls, applying a multifunctional perspective. Besides, their work opens for a discussion of the semantic/neural relationship between impression and concept, which is at the inner, neurological core of the mismatch dilemma, helping us to explain how impressions might be transformed to expressions.
Since some researchers are puzzled with the semantic capacities of animals also in their interaction with humans, my study includes even this type of studies. I have chosen two studies of dogs’ ability to learn and remember names and terms for concrete objects (Pilley and Reid, 2011; Fugazza et al. 2021) as well as Lyn and Christopher’s (2020) studies of great apes.
Highlighting and grouping these works aim at bringing the question of animal semantics some steps further. Although these studies’ final destinations and goals may differ, they all apply a more developed theoretical design in order to embrace and interrelate more aspects of animal communication. Hence, the main aim of my own article is to seek, critically, among these works for expanded common grounds, if any, for understanding animals’ semantic capacities in a systemic perspective. The inquiry of the studies is not treated one by one in a certain order. Rather, elements from the studies are extracted and clustered around particular key topics, and issues that hopefully can bring discussions on animal semantics some steps further, and perhaps can generate new, relevant, researchable questions. Although my investigation is rooted in these researchers’ empirical work, it seems fair to say that my own approach is mainly theoretical.
Utterance theory as a framework for inquiry
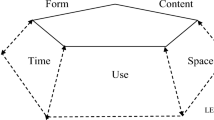
Between the above lines one can sense the assumption that a theory or a framework aiming to grasp the variety of these studies should be broad, multidimensional, multi-leveled, and integrated. A key concept for this effort is utterance. Utterances are expressed in any semiotics, or any communication channel, and uttering is believed to be a faculty possessed by all animals (Ongstad, 2019). It is seen as a joint construction of the five necessary, integrated, constitutive aspects – structured form, referential content, functional act, contextual time, and contextual space, including place. With Habermas (1986) they are seen as simultaneous, and with Bakhtin (1986) seen as combining expressivity, referenciality, and addressivity in a chronotopical context. Following Morris (1938) communication with signs is considered as basically semiotic, and is studied separately by syntax, semantics, and pragmatics in a context as well as a whole.
The framework further operates with four interrelated levels – sign, utterance, genre, and lifeworld, where signs are seen as dependent elements within utterances. Utterances are independent phenomena but in general often tend to be interpreted as possible, recognizable kinds of macro communication tightly related to a species’ particular life-functions, so-called life-genres (Luckmann, 2009; Ongstad, 2019). The highest or deepest level is the more abstract lifeworld (Umwelt) in which an overall sense or meaning is generated and interpreted (Habermas, 1986; Uexküll, 1921). Lifeworld-related decisions hence come close to outcomes from an active mind (Call and Tomasello, 2011). A fuller version of the framework, which even includes necessary processes, is found in Ongstad (2019, 2021a) and a recent discussion of a generalised genre concept is found in Ongstad (2021b). Methodological implications and consequences are discussed in Ongstad (2014).
This sketched version of the framework has its strength in embracing and relating most key aspects that might be at work in animal communication and in studies thereof, and its ditto weakness in increased dependency on more open-ended interpretations (Ongstad, 2021b). Also, it has proved workable when comparing different approaches and epistemologies in a variety of other domains and fields (Ongstad, 2014, 2021b). Since the framework is not a proper method it will be used more loosely as a set of different optics, offering different lenses to give alternative perspectives on concepts, designs, and empirical results in the excerpted cluster of studies. The inspected aspects are thus positioned epistemologically (Ongstad, 2021b) regarding animal semantic capacities in a variety of contexts. The reading and interpretation of the studies do not follow a certain methodology. Hence, focus is both on how semantics might relate to the other aspects and on the relationship between the semantics of uttering and receiving as a system.
Critical inspections of semantic studies
Paradigmatic shifts in the history of studying animal semantics
In the history of empirical research on animal communication semantics hardly had a place until the first classical studies of honey-bees waggle-dance and vervet monkeys’ alarm system (von Frisch, 1967; Seyfarth et al, 1980a, b). The debates in the wake of these findings have led to somewhat different positions regarding animals’ capacity to transmit information, to refer, to convey meaning or the like, in short, what is meant by semantics and semantic capacity (Manser, 2013; Lyn and Christopher, 2020). The first period was characterised by sender-receiver models, with the underlying idea of something being transmitted, with shifting foci on which participants might play the main part. For instance, according to Font and Carazo (2010, p. e3) Krebs & Dawkins (1984) put receivers in focus contending that communication generates from “[…] a coevolutionary arms race between senders playing the role of manipulators and receivers playing the role of mind-readers of sender’s intentions and possible behavior”. Font and Carazo see this positioning in a wider context and emphasise that:
“[…] newer studies have revealed that receivers can and indeed do obtain other types of information about the sender, not just its intentions. Again, it is expressions like ‘information transmission’ or ‘information sharing’, not the concept of information, that are to be blamed for generating confusion” (Font and Carazo, 2010, p. e3).
Later there has been, on the one hand, an increased search for different forms of uttering that might contain content in different species’ communication (Manser, 2013; Fitch, 2019; Kalkman, 2019). On the other hand there are sceptics that doubt animals’ ability to convey, at least arbitrary referential meanings (Wheeler and Fischer, 2012) as well as an increasing number of researchers concluding that there exists a problematic semantic mismatch between systems for uttering and systems for interpreting (Fitch, 2019). The puzzle has led to studies contrasting a claimed simplicity of the repertoire for uttering and a seemingly fairly functional. and thus a pragmatic communicational system for making sense of signalling (Fischer, 2017). Attention has thus moved from a focus on studies of utterers toward receivers, from the semantics of expressing toward the semantics and pragmatics of interpreting and comprehending (Lyn, 2017). Also, it has been claimed that signaller-receiver relations often are asymmetric. However, Bar-On and Moore (2017) argue that the claims are confused by applying ambiguous perceptions of pragmatics. Anyway, in a strongly dichotomized perspective, it is easy to downplay the obvious that any individual sender is also receiver, and vice versa. In other words – how are the roles combined in the processes of input and output? And is there a system that transforms inputs to outputs?
The fallacy of conceptualising concept as verbal
For vertebrates in general and mammals in particular, active, and observable semantics starts working at the moment of the offspring’s birth. A simple hypothesis is that impressions, created by the continuous storing and uptake of segments by a species’ senses, end up as general concepts (Janik and Slater, 1997). By experience and over the years these impressions become more individually differentiated, by explicit learning. Semantic elements or parts are thus not only elements, but also wholes. Wholes and parts continue to interfere and develop throughout life. This view implies that even the most general, abstract parts of a web of experiences should be seen as semantic, not just its specific, observable elements. The semantic patchwork that over time is generated by perception may eventually become part of an animal’s life-world (Ongstad, 2021a).
In parallel all species make experiences by active communication. They have to relate to the uttering of others by which perception and communication become intertwined (Green et al., 2020). What-is-what becomes blurred. In the inner world (Habermas, 1981), a possible mind, impressions might develop into workable concepts. However, an animal concept can in no way be comprehended in linguistic terms or be associated with human verbality (Fitch, 2019). In the animal world concepts should be directly related to processed and stored impressions stemming from each of their specific senses and different modes used for communication (Hertzog & Scharnowski, 2016). In other words, animal concepts are sign based and thus semiotic. For all animals there is a potential mismatch between the two abilities, to percept and to utter, between impression and expression:
Thus, an organism can have a concept […] independently of any words, sentences or other signals that express this concept. Referential links between real-world objects or events and non-verbal mental concepts (representations) can exist even if an organism has no means in its communication system to express those concepts (Fitch, 2019, p. 3).
In the same article Fitch gives a well-argued defence for the view that animals’ semantic capacity should include their, in my words, ‘passive’ repertoire built up by cognition. To his question Do animals have concepts? he gives a row of empirical examples that leads to the conclusion, that they do, but their (active) signals are not concepts in philosophical and linguistic terms.
The term concept and the concept of concept are nevertheless often contaminated by the tacit ideology of verbal language tempting us to believe that the nomenclature language reflects other species’ realities. Hence, as already claimed, studying animal semantics should at the outset be based on zoo- or biosemiotics, and should not be considered a restricted linguistic semantics, by giving primacy to animals’ own sign systems (based on their ways of sensing). An additional terminological challenge is of course the anthropomorphic infection from English. Concept is an English word, hence an Englishness. English, the new Latin of science, can hardly be avoided though, but a linguistic world-view can (Doyle, 2013). Yet, a semiotic priority does not escape the problem of describing animals’ semantic capacity, since what is empirically researched will eventually be expressed verbally. Following Fitch (2019, p. 3) concept can hence be seen as a non-linguistic (i.e., delimited) mental representation.
The semantics of alarm calls and the problem of calling a call a ‘call’
Calls are by far the most researched animal life-genre and should thus be well suited for meta-studying animal semantics. Regarding the basic semantic question of denotation, Fischer (2017, p. 72) reports that similar structures to alarm calls were used in within- and between-group aggression. She holds that this finding is difficult to reconcile with the idea that calls denote objects in the environment. In addition, she stresses that actual structures are often highly conserved. Nevertheless, it is likely that animals from a semiotic perspective are able to differentiate phenomenologically between kinds of dangers and to store and restore such impressions for future communicational purposes.
While Fischer’s research interest is closely related to finding or refuting evolutionary connections between animal vocalisation and human speech, my own interest is on animals’ ability to utter, and which role animal semantics might play in sending and perceiving utterances. Fischer suggests different strands for further research, of which one is studies of ”[…] the developmental processes involved in the integration of production and perception of vocalizations” (Fischer, 2017, p. 72, my italics). To utter and to interpret utterances include the integration of production and perception of semantic elements.
Although an alarm call perhaps not necessarily points out a concrete object or a specific dangerous predator, it might refer to a phenomenon that implies a more general semantic category. Pell et al. (2018) found that Australian magpies, Cracticus tibicen, extract relevant information about the type of threat from functionally referential alarm calls given by noisy miners, Manorina melanocephala, signalling a predator in flight. This heterospecific eavesdropping is proved by measuring the magpies’ bill angle when listening to taped calls of miners. The semantic element might here simply be direction (upwards), a space-aspect (where) that connects utterance and context. It is already demonstrated that honeybees signal food in certain directions by performing a dance. This means that the actual percept still could be seen as a semantic phenomenon, only just a more general and contextual one, in this case a particular part or aspect of space of place of the magpies’ lifeworld.
The difference between call and utterance is not just a matter of terms. The Englishness of call might tempt users to restrict the meaning to an act, an isolated sign function which Bühler (1934) might have termed signal or to a dynamic utterance aspect which Bakhtin (1986) termed addressivity. According to a theory of utterance to utter would imply three possibilities at once (Habermas, 1981), that is, the simultaneity of expressing an inner state, referring to something the utterance is about, and addressing as an act to others, always uttered in a dynamic context of time and space. This dyadic contextual chronotope is established by the very focusing, a particular circumstance a specific utterance will generate by uttering (Ongstad, 2019 and 2021a). These five aspects create on the one hand trouble for a monadic/categorial as well as a dyadic approach/methodology but might on the other hand open for broader and more specific discussions of so-called calls. In other words, both for utterer and receivers each utterance is in principle always multifunctional, with ditto high risks for misunderstanding, misconception, or dysfunctional communication.
Moreover, utterance theory includes the level of genre. Since the overall hypothesis is that animals can differentiate appropriately between their basic life-functions, they have, to different degrees for different taxa, developed ditto recognisable kinds or forms of communication, partly genetical, partly cultural (by learning and enculturation). These are seen as so-called life-genres (Ongstad, 2019). They reduce uncertainty and make communication for a species more predictable and thus (more) functional.
Dezecache and Berthet on the ‘meaning’ of general alarm calls. A study ‘revisited’
Dezecache and Berthet (2018) review studies of perceptions of alarm calls, focusing on alert (or alarm) contexts documenting a lack of agreement on the definition of calls. This absence motivates their claim for further search for mechanisms underlying vocal production and comprehension. They make it clear though that they present working hypotheses. I find their discussion both informative and relevant and will apply what they have sketched, with the aim to extend the scope for discussions on animal semantics.
There are several observations, statements, and suggestions in Dezecache and Berthet’s article that are worthwhile to consider when dealing with clarification of alarm calls. As they weigh different hypotheses up against each other they suggest that a hypothesis might be appropriate to a specific species, but not necessary to others (Dezecache and Berthet, 2018, p. 1). Further, general calls are found in different (in my word life-function) contexts, concerning for instance alarming, feeding, and socialising. Also, with support in Schlenker et al. (2016) calls do not need to match a concrete object to be semantical. And finally, with support in Wheeler and Fischer (2012), the existence of calls is not regarded as context-specific hints of semantic-cognitive capabilities in animals (Dezecache and Berthet, 2018, p. 13).
In stead of presenting Dezecache and Berthet’s individual arguments I paraphrase some key points concerning semantics capacity in their article in the light of utterance theory. Their point of departure is a review of the long-lasting discussion over the concept “functional referenciality”. (See also Manser, 2016.) They conclude that the status of general calls remains unclear. Since I in Ongstad (2021a) came to similar conclusions, I go straight to their intro for discussing “specific and general calls”:
What are general calls? As seen above, general calls are found in alarm contexts (Fichtel & Kappeler, 2002), feeding contexts (Clay et al., 2012) and may also be present in social contexts (Townsend & Manser, 2013). For the sake of simplicity, we will focus on general alarm calls in the rest of our argumentation (Dezecache and Berthet, 2018, p. 4).
Alarming, feeding, and socialising can hence be seen as examples of life-functions. Stereotype utterances in particular contexts, at times and within spaces, where such processes are at work, could be seen as life-genres. Utterances, in principle seen as multifunctional by theory and thus potentially ambiguous for users, need to be interpreted as a kind, first generally, and probably even more specific, as a sub-genre, depending on how delicate or fine-grained a taxon’s repertoire might be. In the perspective of utterance theory, general calls would just be termed utterance. A call, however, would be seen as focusing addressivity more in general. That is, it is an utterance directed to some other/s. If it turns out to concern a particular life-function, for instance protecting, alarm call seems like an adequate term for the sub-genre in question. While an utterance’s physical vocalisation is concrete and external, the genre is more abstract, and hence internal, which implies that an utterance has to be interpreted by receivers as a possible genre or sub-genre.
They further theorise on the semantics of general alarm calls, discussing especially a caller’s choice between the use of a general and a specific call confronted with different dangers. Theoretically they apply Venn’s diagram(s) to sort out the logic of the semantics and empirically data on natural observations of male blue monkeys’ alarm behavior (Fuller, 2013). The logic of this diagram is similar to the logic of the Y-figure introduced in the beginning to exemplify the part-whole paradox of uttering and understanding as an integral system. As a conclusion the benefit of relating utterance to life-genres and lifeworld when studying alarm calls is the possibility of seeing the elements in a systemic perspective.
To distinguish between call types
Much of the early research on animal signalling, transmittance of information, and functional reference has been performed based on an explicit or implicit sender-receiver-model. Fischer (2017) characterises for instance a particular design with these terms: “The diagnostic criteria for functional reference were acoustic distinctiveness and production specificity at the side of the sender, and differential responses at the side of the receiver” (Fischer, 2017, p. 73, my italics). The implicit methodological solution is to compare a closed category with an observable, delimitable, and describable behavioral activity in a sufficiently controlled context.
As a contrast, an utterance’s nature, as part of a systemic view of semiotic communication, when seen as consisting of form, content, act, time, and space, is multi-dimensional. An utterance is thus both sent and interpreted as a mixture of the form’s expressivity, the content’s referenciality, the act’s addressivity, the time’s temporality, and the space’s topology.
Regarding expressivity, which concerns forms and structures, more delicate methods can reveal whether there is more to call sounds than just being a general alarm call. For instance Prior et al. (2018) use spectrograms to decide to which degree fine-grained communicational structures in finches’ calls which the human ear can not hear, could carry complex information. In their discussion they argue that some signals, or in my words – some utterances, can carry at least three different aspects, categories, or information bits:
There is a significant amount of behavioral and neurobiological research that has demonstrated that zebra finches are able to discriminate the calls of males and females […]. Additionally, zebra finches have 10 call types in their call repertoire, and there is strong physiological evidence that perceptual categories exist for many of those call types […]. Furthermore, there is also independent evidence that zebra finch calls carry individual identity, which is at least accessible to an individual’s pair-bonded mate […] Prior et al. (2018, p. 8) (my italics).
In other words, as Prior et al. themselves point to, zebra finches’ utterances are potentially multifunctional, and there are therefore reasons to believe that the different call types in my terms could be seen as life-genres as they might have different functions in their lifeworld.
Besides, an additional interest here is their claim that some of the types have content or what they term perceptual categories that might function as a basis for references in utterances. With Hertzog et al. (2018) one could call these mind-located category-bits percepts. A reason for using Firth’s outdated notion (Firth, 1949) is to mark a difference between human and animal in order to reduce impact from human conceptualisation, contaminated by the logic and ideology of language. Regarding referencialities in animal utterances there is a need to differentiate between kinds of signs as well as between generating and comprehending meaning, respectively the active and seemingly passive relation to semanticity. Nieder (2009) claims that animals do not use symbolic referencing, while some researchers seem to continue searching (Ongstad, 2021a). A way to deal with content inconsistencies between studies is to relate semantics to form and function, working triadically.
A triadic approach studying a species’ vocal system
The title of Fuller (2013) should indicate his thesis’ relevance for my discussion: Diversity of form, content, and function in the vocal signals of adult male blue monkeys (Cercopithecus mitis stuhlmanni): An evolutionary approach to understanding a signal repertoire. One of the strengths of this research, not just for my purpose, is the ambition to cover most of one specific species’ entire communicational repertoire, its signal system as a whole, a position he finds support for in Gros-Louis et al. (2008). Another is the consequent triadic view he takes on signals, by investigating thoroughly the variety in the three signal elements, acoustic structure, signal contents, and adaptive functions. This multifunctional approach puts his work partly in line even with my framework, seeing utterances as triadic in a context.
A great benefit of his work for the different fields that study animal communication, is his theoretical review and empirical documentation of many sub-categorisations of each of the three signal elements. For content, or information, in vocal signals of several primate species Fuller finds that the following six elements are most frequently identified: species; age-sex class; body size; condition and social dominance; identity; external referents, context, and behavioral commitment.
That said, it nevertheless becomes clear that the thesis of his thesis epistemologically has home in the realm of philosophy and is closer to mainstream ethology than to (bio-)semiotics and different functional theories of communication. This difference raises the question of compatibility with my own approach. Compared, Fuller uses signal where I use utterance. In utterance theory a signal is a sign aspect (Bühler, 1934) where signalling is addressing a receiver (Bakhtin, 1986), and is hence functional. Further, Fuller does not operate with an overarching genre concept, although he does talk about alarm calls as signal classes (Fuller, 2013, p. 33) and generally even sees them as typified communication. As far as I have registered the notions signal and signal classes are not seen as directly interrelated. As a contrast life-genres are seen as crucial resources for interpreting utterances.
In addition my article focuses on the so-called mismatch between supposedly lean uttering and rich understanding, primarily regarding semantics. However, I have stated that semantics should be seen in the light of semiotic syntax and semiotic pragmatics as well as contextual time and space. What Fuller’s study offers in this respect, without necessarily using such terms is a highly differentiated theoretical understanding of syntactically structured forms of vocalities, as well as a discussion of the various pragmatic sub-functions signals have and can have. The documented patterns thus lead to a major epistemological question: How is function to be understood systemically?
First, function seems mainly perceived as a causality phenomenon, since a signal leads to a reaction, the signal has an effect. The more often the same signal matches the same documented effect in receivers, the more significant a function is taken to be. On the other hand, a semiotic, communicational view, based in utterance theory may avoid function and primarily speak about signalling or addressing on behalf of the utterer, and describe receivers’ re-/actions or behavior, in other word as a communicational act, a response. The behavior of receivers is an effect or a consequence. To understand what the reactions are or represent, third parts, such as observers should have sufficient overall knowledge of the animal’s life-functions, anticipating that reactions or lack of reactions are results of the animal’s use of ditto life-genres.
Further, signal content is described briefly by Fuller (2013, p. 108) as attributes of signallers that are reliably indicated by features of signals. He examined vocal behavior of 32 adult male blue monkeys (C. m. stuhlmanni) to identify content in six call types in the male repertoire. At least the three boom, katrain, and pyow were reliable indicators of identity, and features of pyow were correlated with body size. He also found that social status was content in all calls except nasal screams. Further, ka and katrain “[…] were strongly associated with […] aerial predators, and one call (ant) had a similar relationship to terrestrial predators (Fuller, 2013, p. 109). Pyow was associated with terrestrial predators and boom with multiple external variables. Nasal screams were used in rather aggressive situations.
This rough outline from Fuller’s abstract shall just make clear that, based on his intricate empirical research, we firstly can refute a one-to-one relationship between a call and a function at least for this species. Alternatively or instead, the five described utterances (nasal screams excluded) could, in the ears and minds of receivers, be seen as possible kinds of communication, as life-genres, utterances that need to be interpreted as types (‘classes’). For the observing researcher precise claims are possible due to the empirical coverage of much of the blue monkey’s life-world, although Fuller warns that the content identified in his study does not constitute an exhaustive list for male blue monkey vocalisations (Fuller, 2013, p. 151). Secondly, and most important for my study, the mismatch has here lost some of its fuzziness since the relationship between utterance capacities and receptive use (function) for this taxon is investigated in great detail. Communication seems to take the shape of a possible, comprehensible system.
However, we are still left with the problem of treating both references, defined as true symbols and information about the sending utterer achieved by indication, as semiotic semantics. My answer here is, for the time being, that a broader concept of animal semantics seems more promising for further studies of semantic capacities in an evolutionary perspective, and hence in a contextual perspective, than a more restricted one. Which leads to the inclusion of studies of semantics in non-wild contexts.
Dogs’ capacity for human verbal terms and concepts?
Humans can force a percept upon an animal’s mind. Pillay and Reid (2011:184) claim that their studies of the border collie Chaser indicated that “[…] Chaser acquired referential understanding of nouns.” They further hold that it is proved that Chaser learned more than 1000 nouns for objects, and even understood (or was not confused by) that an object may have more than one name or term (Pillay and Reid 2011:195). These studies are just used by me as one of several indications of the likelihood that so-called higher order animals can have specific percepts on their mind, that is, that they possess a mental faculty to manage ‘content’ in a broad sense. Since dogs by sniffing have obtained a highly advanced ability to differentiate between countless smells, what I have called percepts, it is not unlikely that a parallel or similar mental network can be established by signs, by semiosis, whether this happens as an innate process and/or is created by humans (cf. Pavlov).
Pillay and Reid’s studies have been criticised for drawing too general conclusions based on the design of the studies (for instance lack of control group). Fugazza et al. (2021) have done similar studies with a more vigorous design investigating dog’s rapid learning of object names in a social context, finding that two particular dogs, Whiskey and Vicky Nina, were able to learn and remember words over time for objects based on four different types of tests (Fugazza et al., 2021, p.3). They self-critically used the same procedure with 20 typical family dogs as with the two exceptional dogs with a high-performance rate. They found that these family dogs did not demonstrate any evidence of learning remembering over time the object names at the same rate as the high achievers: ”This suggests that only a few subjects show this ability. Future studies should investigate whether this outstanding capacity stems from the exceptional talent of some individuals or whether it emerges from previous experience with object name learning” (Fugazza et al. 2021, p. 1). Although their study focuses mainly on learning and memory of concepts or terms for objects, it shows that our understanding of animal’s semantic capacity will on the one hand gain from studying them in different contexts. On the other hand, it turns out that the notion of context is problematic. Although the two dogs showed increased semantic competence, the researchers associated their ability with the specific social play-with-words-context the owners already had established from early on.
Apes’ extended, learned semantic repertoire
According to Lyn (2012) studies based of so-called Wh-questions directed to apes made clear that apes by means of sign language were cognitively able to manage such questions adequately. While the researchers behind this research, Gardner and Gardner (1969), saw this ability both as an indication of multidimensional representation of symbols and as a grammatical capacity, Lyn concludes more carefully: “However, all these findings, taken together, certainly suggest that some categorical information is part of the apes’ representations of the symbols”.
It seems generally accepted that what apes can do, both cognitively and communicationally, in tests by far surpasses what they are observed to do in the wild. Lyn (2012) suggests that this increase in achievement with a Vygotskyan notion could be seen as within their “zone of proximal development”. For her the discrepancy is due to missing cultural development, rather than a missing biological development. This view is a tacit attack on the idea of natural habitat for each species as natural and given. In an evolutionary perspective environmental shifts contribute significantly to new varieties and eventually to new species with new competences. Lyn thus concludes, based on a row of meta-studies of apes, that environment has been shown to be the strongest modifier of communicative capability in apes (Lyn, 2012).
In a personal conversation with me Richard Moore points out that regarding mismatch between production and comprehension of great apes’ semantic capacities in a non-wild environment a striking illustration can be found by looking at the famous bonobo Kanzi’s communicative repertoire, well illustrated in Shanker, Savage-Rumbaugh, and Taylor (1999).
These views put the ideas of animal learning and culture, and hence life-genres in focus. Although most capacities probably are gene-based it is recognised that for instance certain song-birds songs, their utterances, are explicitly taught (Beecher, 2008), that some bird-dances are exercised, and that among killer whales hunting strategies, and hence communications are trained. These and similar animal life-genres are taught and learned and hence part a species culture (Schuppli and van Schaik, 2019). If or when a habitat changes, the culture, and thus the life-genres, may change with it and therefore even the set of percepts/concepts that are associated with it, and hence the system as such.
The hypothesis of animal interpretation
Summing up
The above meta-inquiries have brought to surface some key patterns in contemporary research on animal semantic capacities:
-
1.
Both syntactic structures and possible semantic content of utterances are seen as richer than earlier believed. Research continues to reveal new complexities in utterances and in ability to interpret.
-
2.
There is both a theoretical and empirical willingness to broaden the perception of animals’ semantic capacity to comprehend communication.
-
3.
The ambition to make sense of these tendencies and to continue to explore these questions is often motivated by search for evolutionary links to verbal language, but several researchers hesitate to draw clear lines.
-
4.
The scientific frame for studying semantic capacity among animals includes new contexts challenging the norm of studying animals in their natural habitats and add to the understanding of both semantics and capacities.
-
5.
There is a general growing awareness of the challenge of finding a proper term for so-called animal concepts. Most studies seem to refute a pure linguistic view on semantics, and some few distinguish between input and output by applying the notion percept for the mental storing or uptake of impressions.
-
6.
None of the studies discuss the principal question of how results from studies of semantic competencies or capacities in the wild and the none-wild can be amalgamated, although some argue that the different approaches should be seen as compatible when the criterion is capacity.
-
7.
For historical reasons, many studies investigate the semantics of alarm calls because of the richness of studies in this field. However, few studies problematise the concept of call or the anthropomorphic risk of using English language in classifying kinds of calls.
-
8.
Some studies argue that one should not generalise too far about semantic capacities from one species to another. One study, of dogs, even warns that there are major variations in capacity within a species.
-
9.
Several of the studies are explicitly multifunctional in different ways but are mostly reluctant to extend discussions to include systemic implications a multifunctional view may have on animal communication in general and on animal semantics in particular.
-
10.
Although most studies bring together different aspects or problematise the multifunctional nature of utterances and receivers’ interpretations, reactions and behavior, none explicitly seems to discuss such elements as aspects in a possible overarching system for each taxon or species.
Even if the semantics of uttering still is seen as quite restricted, animals’ semantic readiness for understanding could nevertheless be seen as fairly rich. Given that animals do consider utterances, raises the question – by means of what? To integrate uttering and understanding as aspects (parts) to a functional communicational whole, calls for concepts that can explain communicational patterns in new contexts, can show how animals infer communicationally, and can incorporate levels, aspects, and processes. What follows are arguments for some suggested additional concepts that can back up the hypothesis that animals do interpret, based on semiotic semantics, and that a relatively broad semantic repertoire is a key requisite in an animal’s and a species’ communicational system.
Animals’ communicative understanding. On abduction, affordances, life-genre, and lifeworld
Cognitively it is proved that some animals are able to draw rational conclusions from experiences both in the wild and in test situations (Healy, 2019). Generally we think of this mental, cognitive capacity in terms of inference, as induction and/or deduction. A more radical idea is that due to so-called higher order animals’ general lack of a well developed and differentiated system for active uttering, their fairly rational and differentiated reactions to such forms of communication as alarm calls might work by abduction (Magnani, 2007; Park, 2017). Based on the idea of abduction (Peirce, 1992) the sum of impressions of different senders’ utterances and a fairly rational understanding of the specific context can be seen as a basis for an interpretative abductional understanding. The older and thus the more experienced an animal is, the better abduction in the long run might work as behavioral reactions to warnings of different kinds and of developing the sense for different degrees of specificity. Animal life-genres could be seen as systemic, which in this context roughly means partly given, partly open communicational systems. Accordingly, consideration and interpretation are key process patterns and the mental way of managing them could be seen as abductional. Abduction is hence not necessarily a more advanced way of reasoning than induction and deduction, but perhaps more basic? Hence, it is perhaps not illogical that abduction as a form of inference occurs early in evolution? In an evolutionary perspective abductional capacities might cover much of a species need for a functional semiotic system.
Hence, close to abduction is the idea of affordances (Gibson, 1979; Dennett, 2018), what contextual environments cognitively offer an animal for good and bad. This is close both to Lyn’s idea of Vygotsky’s principle of scaffolding (Lyn, 2012, p. 17) and to the notion of social context in Fugazza et al. (2021). Environment can further be associated with two tightly connected perspectives, the material world in which a species bodily lives, and its overall mental storing of it, the mental umwelt, or in my terms lifeworld, by which each single animal try to make sense of communications in its mind (Deacon, 2013). Two key mental tools that can help to establish this kind of lifeworld in which animals can abduce, are utterances and genres (Habermas, 1981; Bakhtin, 1986). Animal utterances are probably not rich, when seen as a separate sub-system for semantics. However, when combined with life-genre (Ongstad, 2019, 2021a; Luckmann, 2009) as a source for interpreting and hence seen as a whole, as a dynamic, overarching system, more of the fuzziness could be explained and hence be seen more as a match than as a mismatch. The relationship between fairly simple utterances and quite impressive mental maps of animals’ lifeworld built up by experiences with life-genres and used for interpretation, is hence no longer necessarily a mismatch.
However, the claimed relationship can probably not make sense unless a couple of other theoretical assumptions related to the framework also are considered. Animals’ cognition has been internalised by semiotic communication, by the dynamics between uttering and the interpreting by life-genres (Ongstad, 2019). On this point I agree with Fitch’s insistence on taking cognition into account (Fitch, 2019), though I still prefer the Habermasian/Bakhtinian view that all utterance aspects arrive at a receiver at once. When a specific genre is applied to interpret which aspects of the utterance are dominant (Jakobson, 1935) in a given situation, it is seen as applied abductionally. Even if inefficient communication of course may occur for individuals and for a species, such as faults and misconceptions, most systems seem to work fairly functionally after all. Yet, when the environmental, contextual conditions change significantly, genres can simply die out being too dysfunctional or be further specialised and enhanced which is a key point in Lyn and Christopher (2020).
To establish a grand theory of wholeness in communication is out of question, but further advances in the connectedness of key aspects in animal communication seem feasible. In this meta-study of semantic capacities in different contexts it is suggested that to move from micro toward macro combining semantics, syntax, and pragmatics in different contexts is a worthwhile option. Yet, it should be admitted that further progress even rests on future advances in studies of animals’ inner neurological processes when interpreting utterances and activities related to contexts, as pointed to in several of the inspected studies. On the day of finishing this article a Norwegian newspaper announced that a research group (Gardner et al., 2021) at the Norwegian University of Science and Technology, NTNU, lead by the Nobel prize winners E. and M. B. Moser, has revealed how thousands of nerve cells in the brain work together to form an animal’s sense of place.
References
Bakhtin, M. (1986). Speech genres and other late essays. Austin: University of Texas Press
Bar-On, D., & Moore, R. (2017). Pragmatic interpretation and signaller-receiver asymmetries in animal communication. In K. Andrews, & J. Beck (Eds.), The Routledge handbook of philosophy of animal minds (pp. 299–300). London: Taylor & Francis
Barbieri, M. (2020). The semantic theory of evolution. Routledge
Barbieri, M. (2020). The semantic theory of language. BioSystems, 190, 104100. homepage:www.elsevier.com/locate/biosystems
Beecher, M. D. (2008). Function and mechanisms of song learning in song sparrows. Advances in the Study of Behavior, 38, 167–225
Beecher, M. D. (2021). Why Are No Animal Communication Systems Simple Languages? Frontiers in Psychology, 12, 701
Bühler, K. (1934). Sprachtheorie. Stuttgart: Fischer
Call, J., & Tomasello, M. (2011). Does the chimpanzee have a theory of mind? 30 years later.Human Nature and Self Design,83–96
Clay, Z., Smith, C. L., & Blumstein, D. T. (2012). Food-associated vocalizations in mammals and birds: what do these calls really mean? Animal Behaviour, 83(2), 323–330
Deacon, T. W. (2013). Incomplete nature: How mind emerged from matter. New York: Norton
Dezecache, G., & Berthet, M. (2018). Working hypotheses on the meaning of general alarm calls. Animal Behaviour, 142, 113–118
Dennett, D. (2018). From Bacteria to Bach and Back. The evolution of minds. London: Penguin Books
Dittrich, P. (2018). Towards measuring the semantic capacity of a physical medium demonstrated with elementary cellular automata. Biosystems, 164, 177–185
Doyle, B. (2013). English and Englishness. London: Routledge
Fedurek, P., Zuberbühler, K., & Dahl, C. D. (2016). Sequential information in a great ape utterance. Scientific reports, 6, 38226. doi: https://doi.org/10.1038/srep38226 (2016)
Fichtel, C., & Kappeler, P. M. (2002). Anti-predator behavior of group-living Malagasy primates: mixed evidence for a referential alarm call system. Behavioral Ecology and Sociobiology, 51(3), 262–275
Firth, R. (1949). Sense-data and the percept theory. Mind, 58(232), 434–465
Fischer, J. (2017). Primate vocal production and the riddle of language evolution. Psychonomic Bulletin & Review, 24(1), 72–78
Fischer, J. (2020). Nonhuman primate alarm calls then and now.Anim. Behav. Cogn.7(2)108–116. https://doi.org/10.26451/abc.07.02.05.2020
Fitch, W. T. (2019). Animal cognition and the evolution of human language: why we cannot focus solely on communication. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 375(1789), 20190046–20190046
Font, E., & Carazo, P. (2010). Animals in translation: Why there is meaning (but probably no message) in animal communication. Animal Behaviour, 80(2), e1–e6. https://doi.org/10.1016/j.anbehav.2010.05.015
Fugazza, C., Andics, A., Magyari, L., et al. (2021). Rapid learning of object names in dogs. Sci Rep, 11, 2222. https://doi.org/10.1038/s41598-021-81699-2
Fuller, J. L. R. (2013). Diversity of form, content, and function in the vocal signals of adult male blue monkeys (Cercopithecus mitis stuhlmanni): An evolutionary approach to understanding a signal repertoire. New York: Columbia University
Gardner, R. A., & Gardner, B. T. (1969). Teaching sign language to a chimpanzee. Science, 165, 664–672
Gardner, R. J., Hermansen, E., Pachitariu, M., Burak, Y., Baas, N. A., Dunn, B. J. … Moser, E. I. (2021). Toroidal topology of population activity in grid cells. bioRxiv, 1–26. Published online 12th January 2022 by Nature. https://doi.org/10.1038/s41586-021-04268-7
Gabrić, P. (2021). Overlooked evidence for semantic compositionality and signal reduction in wild chimpanzees (pan troglodytes). An. Acad. Bras. Ciênc, 76(2), 325–334
Gibson, J. J. (1979). The Ecological Approach to Visual Perception. Boston: Houghton Mifflin
Godfrey-Smith, P. (2016). Other minds: The octopus, the sea, and the deep origins of consciousness. Farrar, Straus and Giroux
Green, P. A., Brandley, N. C., & Nowicki, S. (2020). Categorical perception in animal communication and decision-making. Behavioral Ecology, 31(4), 859–867
Gros-Louis, J. J., Perry, S. E., Fichtel, C., Wikberg, E., Gilkenson, H., Wofsy, S., & Fuentes, A. (2008). Vocal repertoire of Cebus capucinus: acoustic structure, context, and usage. International Journal of Primatology, 29(3), 641–670
Habermas, J. (1981). The theory of communicative action. London: Beacon Press
Habermas, J. (1986). Kommunikativt handlande. Texter om språkrationalitet och samhälle. Göteborg: Daidalos
Healy, S. D. (2019). The face of animal cognition. Integrative zoology, 14(2), 132–144
Herzog, M. H. et al. (2016). Time slices: what is the duration of a percept? PLoS Biology, 14(4), 1–12
Jackendoff, R. (1983). Semantics and cognition (8 vol.). MIT press
Jabr, F., & Rothschild, A. (2012). How brainless slime molds redefine intelligence. Scientific American, 7, p. 1. https://www.nature.com/articles/nature.2012.11811.pdf?origin=ppub visited 07.07. 2021
Jakobson, R. (1935). The dominant. In L. Mateijka, & P. Krystyna (Eds.), Readings in Russian poetics: Formalist and structuralist views (pp. 82–87). Cambridge: The MIT Press
Janik, V. M., & Slater, P. J. (1997). Vocal learning in mammals. Advances in the Study of Behaviour, 26, 59–100
Kalkman, D. (2019). Meaning in Animal Communication: Varieties of meaning and their roles in explaining communication. Unpublished Ph.D. thesis. Canberra: The Australian National University. https://openresearch-repository.anu.edu.au/bitstream/1885/151932/1/David%20Kalkman%20Thesis%202019.pdf
Krebs, J. R., & Dawkins., R. (1984). Animal Signals: Mind-reading and Manipulation.Behavioural Ecology: An Ew) lutionary Approach. Blackwell. Boston,380–402
Luckmann, T. (2009). Observations on the structure and function of communicative genres. Semiotica, (173), 267–282
Lyn, H. (2012). Apes and the Evolution of Language: Taking Stock of 40 Years of Research. In J. Vonk, & T. K. Shackelford (Eds.), The Oxford handbook of comparative evolutionary psychology (pp. 356–380). Oxford: Oxford University Press
Lyn, H. (2017). The question of capacity: Why enculturated and trained animals have much to tell us about the evolution of language. Psychonomic Bulletin and Review, 24, 85–90
Lyn, H., & Christopher, J. L. (2020). How environment can reveal semantic capacities in nonhuman animals. Animal Behavior and Cognition, 7(2), 159–167. https://doi.org/10.26451/abc.07.02.10.2020
Magnani, L. (2007). Animal abduction. From mindless organisms to artefactual mediators. In L. Magnani, & P. Li (Eds.), Model-Based Reasoning in Science, Technology, and Medicine (pp. 3–38). Berlin: Springer
Manser, M. B. (2013). Semantic communication in vervet monkeys and other animals. Animal Behaviour, 86(3), 491–496
Manser, M. B. (2016). Referents and semantics in animal vocalizations. Psychological mechanisms in animal communication (pp. 223–249). Cham: Springer
Marconi, M. A., Nicolakis, D., Abbasi, R., Penn, D. J., & Zala, S. M. (2020). Ultrasonic courtship vocalizations of male house mice contain distinct individual signatures. Animal Behaviour, 169, 169–197
Morris, C. W. (1938). Foundations of the Theory of Signs. In O. Neurath, R. Carnap, & C. W. Morris (Eds.), Foundations of the unity of science: toward an international encyclopedia of unified science (pp. 1–59). Chicago: University of Chicago Press
Nieder, A. (2009). Prefrontal cortex and the evolution of symbolic reference. Current opinion in neurobiology, 19(1), 99–108
Ongstad, S. (2014). The Blindness of Focusing. Pragmatic theories of communication and the challenge of validation. Reconceptualizing Educational Research Methodology, 5(2), 128–144
Ongstad, S. (2019). A conceptual framework for studying evolutionary origins of life-genres. Biosemiotics, 12(2), 245–266
Ongstad, S. (2021a). Can Animals Refer? Meta-Positioning Studies of Animal Semantics. Biosemiotics, 14(2), 433–457
Ongstad, S. V. (2021b). Genre Constituents in “Reflections on Genre as Social Action”–in the Light of 1980s’ Genre Research? Discourse and Writing/Rédactologie, 31, 86–108
Park, W. (2017). Magnani’s Manipulative Abduction. In L. Magnani, & T. Bertolotti (Eds.), Springer Handbook of Model-Based Science (pp. 41–66). Cham: Springer. https://doi.org/10.1007/978-3-319-30526-4_9
Peirce, C. S. (1992). The essential Peirce: selected philosophical writings (2 vol.). Indiana University Press
Pell, F. S. D. et al. (2018). Birds orient their heads appropriately in response to functionally referential alarm calls of heterospecifics. Animal Behaviour, 140, 109–118
Perreault, C., & Mathew, S. (2012). Dating the origin of language using phonemic diversity.PloS one, 7(4), e35289
Pilley, J. W., & Reid, A. K. (2011). Border collie comprehends object names as verbal referents. Behavioural processes, 86(2), 184–195
Prior, N. H., Smith, E., Lawson, S., Ball, G. F., & Dooling, R. J. (2018). Acoustic fine structure may encode biologically relevant information for zebra finches. Scientific reports, 8(1), 6212. DOI:https://doi.org/10.1038/s41598-018-24307-0
Schlenker, P., Chemla, E., Schel, A. M., Fuller, J., Gautier, J. P., Kuhn, J. … Keenan, S. (2016). Formal monkey linguistics. Theoretical Linguistics, 42(1–2), 1–90. https://doi.org/10.1515/tl-2016-0001
Schuppli, C., & van Schaik, C. P. (2019). Animal cultures: how we’ve only seen the tip of the iceberg.Evolutionary Human Sciences, 1
Seyfarth, R. M., Cheney, D. L., & Marler, P. (1980a). Monkey responses to three different alarm calls: evidence of predator classification and semantic communication. Science, 210, 801–803
Seyfarth, R. M., Cheney, D. L., & Marler, P. (1980b). Vervet monkey alarm calls: semantic communication in a free-ranging primate. Animal Behavior, 28, 1070–1094
Shanker, S. G., Savage-Rumbaugh, E. S., & Taylor, T. J. (1999). Kanzi: A new beginning. Animal Learning & Behavior, 27(1), 24–25
Suzuki, T. N. (2021). Animal linguistics: exploring referentiality and compositionality in bird
calls.Ecol. Res.36(2),221–231. https://doi.org/10.1111/1440-1703.12200
Terrace, H. S. (2019). Why chimpanzees can’t learn language and only humans can. New York: Columbia University Press
Townsend, S. W., & Manser, M. B. (2013). Functionally referential communication in mammals: the past, present and the future. Ethology, 119(1), 1–11
Townsend, S. W., Engesser, S., Stoll, S., Zuberbühler, K., & Bickel, B. (2018). Compositionality in animals and humans. PLoS Biol, 16(8), e2006425. https://doi.org/10.1371/journal.pbio.2006425
Uexküll, J. (1921). Umwelt und Innerwelt der Tiere. Berlin: Springer
von Frisch, K. (1967). The dance language and orientation of bees. Cambridge, MA: Harvard University Press
Wheeler, B. C., & Fischer, J. (2012). Functionally referential signals: a promising paradigm whose time has passed. Evolutionary Anthropology: Issues, News, and Reviews, 21, 195–205
Zlatev, J. (2012). Cognitive semiotics: An emerging field for the transdisciplinary study of meaning. Public Journal of Semiotics, 4(1), 2–24
Acknowledgements
I would like to thank prof. em. Jon Smidt and dr. Richard Moore as well as the two anonymous reviewers for comments on a draft of this article.
Funding
Not applicable.
Open access funding provided by OsloMet - Oslo Metropolitan University
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ongstad, S. Simple Utterances but Complex Understanding? Meta-studying the Fuzzy Mismatch between Animal Semantic Capacities in Varied Contexts. Biosemiotics 15, 85–108 (2022). https://doi.org/10.1007/s12304-022-09477-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12304-022-09477-9