
Abstract
Key message
Oxalotrophic Stenotrophomonas isolated from tomato rhizosphere are able to protect plants against oxalate-producing pathogens by a combination of actions including induction of plant defence signalling callose deposition and the strengthening of plant cell walls and probably the degradation of oxalic acid.
Abstract
Oxalic acid plays a pivotal role in the virulence of the necrotrophic fungi Botrytis cinerea and Sclerotinia sclerotiorum. In this work, we isolated two oxalotrophic strains (OxA and OxB) belonging to the bacterial genus Stenotrophomonas from the rhizosphere of tomato plants. Both strains were capable to colonise endophytically Arabidopsis plants and protect them from the damage caused by high doses of oxalic acid. Furthermore, OxA and OxB protected Arabidopsis from S. sclerotiorum and B. cinerea infections. Bacterial inoculation induced the production of phenolic compounds and the expression of PR-1. Besides, both isolates exerted a protective effect against fungal pathogens in Arabidopsis mutants affected in the synthesis pathway of salicylic acid (sid2-2) and jasmonate perception (coi1). Callose deposition induced by OxA and OxB was required for protection against phytopathogens. Moreover, B. cinerea and S. sclerotiorum mycelial growth was reduced in culture media containing cell wall polysaccharides from leaves inoculated with each bacterial strain. These findings suggest that cell walls from Arabidopsis leaves colonised by these bacteria would be less susceptible to pathogen attack. Our results indicate that these oxalotrophic bacteria can protect plants against oxalate-producing pathogens by a combination of actions and show their potential for use as biological control agents against fungal diseases.












Similar content being viewed by others

References
Asselbergh B, Höfte M (2007) Basal tomato defences to Botrytis cinerea include abscisic acid-dependent callose formation. Physiol Mol Plant Pathol 71:33–40
Bateman DF, Beer SV (1965) Simultaneous production and synergistic action of oxalic acid and polygalacturonase during pathogenesis by Sclerotiorum rolfsii. Phytopathol 55:204–211
Bravo D, Braissant O, Cailleau G, Verrecchia E, Junier P (2015) Isolation and characterization of oxalotrophic bacteria from tropical soils. Arch Microbiol 197:65–77
Cantu D, Vicente AR, Labavitch JM, Bennett AB, Powell ALT (2008) Strangers in the matrix: plant cell walls and pathogen susceptibility. Trends Plant Sci 13:610–617
Carpita NC, McCann M (2000) The cell wall. In: Buchanan BB, Gruissem W, Jones RL (eds) Biochemistry and molecular biology of plants. American Society of Plant Biologists, Rockville, pp 45–110
Cecchini NM, Monteoliva MI, Alvarez ME (2011) Proline dehydrogenase contributes to pathogen defense in Arabidopsis. Plant Physiol 155(4):1947–1959
Cessna SG, Sears VE, Dickman MB, Low PS (2000) Oxalic acid, a pathogenicity factor for Sclerotinia sclerotiorum, suppresses the oxidative burst of the host plant. Plant Cell 12:2191–2199
Chipps TJ, Gilmore B, Myers JR, Stotz HU (2005) Relationship between oxalate, oxalate oxidase activity, oxalate sensitivity, and white mold susceptibility in Phaseolus coccineus. Phytopathol 95(31):292–299
da Silva LF, Dias CV, Cidade LC et al (2011) Expression of an oxalate decarboxylase impairs the necrotic effect induced by Nep1-like protein (NLP) of Moniliophthora perniciosa in transgenic tobacco. Mol Plant Microbe Interact 24(7):839–848
de Nogueira EM, Vinagre F, Masuda HP, Vargas C, de Pádua VLM, da Silva FR, dos Santos RV, Baldani JI, Ferreira PCG, Hemerley (2001) Expression of sugarcane genes induced by inoculation with Gluconacetobacter diazotrophicus and Herbaspirillum rubrisubalbicans. Genet Mol Biol 24:99–206
Dias B, Cunha W, Morais L (2006) Expression of an oxalate decarboxylase gene from Flammulina sp. in transgenic lettuce (Lactuca sativa) plants and resistance to Sclerotinia sclerotiorum. Plant Pathol 55:187–193
Dutton MV, Evans CS (1996) Oxalate production by fungi: its role in pathogenicity and ecology in the soil environment. Can J Microbiol 42:881–895
Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39:783–791
Ferrari S, Plotnikova JM, De Lorenzo G, Ausubel FM (2003) Arabidopsis local resistance to Botrytis cinerea involves salicylic acid and camalexin and requires EDS4 and PAD2, but Not SID2, EDS5 or PAD4. Plant J 35:193–205
Feys BJ, Benedetti CE, Penfold CN, Turner JG (1994) Arabidopsis mutants selected for resistance to the phytotoxin coronatine are male sterile, insensitive to methyl jasmonate, and resistant to a bacterial pathogen. Plant Cell 6:751–759
Franceschi VR, Loewus FA (1995) Oxalate biosynthesis and function in plants and fungi. In: Khan SR (ed) Calcium oxalate in biological systems. CRC Press, Boca Raton, pp 113–130
Godoy G, Steadman JR, Dickman MB, Dam R (1990) Use of mutants to demonstrate the role of oxalic acid in pathogenicity of Sclerotinia sclerotiorum on Phaseolus vulgaris. Physiol Mol Plant Pathol 37:179–191
Guimarães RL, Stotz HU (2004) Oxalate production by Sclerotinia sclerotiorum deregulates guard cells during infection. Plant Physiol 136:3703–3711
Hallmann J, Quadt-Hallmann A, Mahaffee WF, Kloepper JW (1997) Bacterial endophytes in agricultural crops. Can J Microbiol 43:895–914
Heller A, Witt-Geiges T (2013) Oxalic acid has an additional, detoxifying function in Sclerotinia Sclerotiorum pathogenesis. PLoS ONE 8:e72292. https://doi.org/10.1371/journal.pone.0072292
Hu X, Bidney DL, Yalpani N, Duvick JP, Crasta O, Folkerts O, Lu G (2003) Overexpression of a gene encoding hydrogen peroxide-generating oxalate oxidase evokes defense responses in sunflower. Plant Physiol 133:170–181
Iniguez AL, Dong Y, Carter HD, Ahmer BMM, Stone JM, Triplett EW (2005) Regulation of enteric endophytic bacterial colonization by plant defenses. Mol Plant Microb Interact 18:169–178
Kabbage M, Williams B, Dickman MB (2013) Cell death control: the interplay of apoptosis and autophagy in the pathogenicity of Sclerotinia sclerotiorum. PLoS Pathog 9:e1003287. https://doi.org/10.1371/journal.ppat.1003287
Kesarwani M, Azam M, Natarajan K, Mehta A, Datta A (2000) Oxalate decarboxylase from Collybia velutipes. Molecular cloning and its overexpression to confer resistance to fungal infection in transgenic tobacco and tomato. J Biolog Chem 275:7230–7238
Kim KS, Min JY, Dickman MB (2008) Oxalic acid is an elicitor of plant programmed cell death during Sclerotinia sclerotiorum disease development. Mol Plant Microbe Interact 21:605–612
Kloepper JW, Ryu CM, Zhang S (2004) Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathol 94:1259–1266
Kost TNS, Agnoli K, Eberl L, Weisskopf L (2014) Oxalotrophy, a widespread trait of plant-associated burkholderia species, is involved in successful root colonization of lupin and maize by Burkholderia phytofirmans. Front Microbiol 4:1–9
Kumar V, Chattopadhyay A, Ghosh S, Irfan M, Chakraborty N, Chakraborty S, Datta A (2015) Improving nutritional quality and fungal tolerance in soya bean and grass pea by expressing an oxalate decarboxylase. Plant Biotechnol J. https://doi.org/10.1111/pbi.12503
Kunz C, Vandelle E, Rolland S et al (2006) Characterization of a new, nonpathogenic mutant of Botrytis cinerea with impaired plant colonization capacity. New Phytol 170:537–550
Le Gall H, Philippe F, Domon JM, Gillet F, Pelloux J, Rayon C (2015) Cell wall metabolism in response to abiotic stress. Plants 4:112–166
Marciano P, Di Lenna P, Magro P (1983) Oxalic acid, cell wall degrading enzymes and pH in pathogenesis and their significance in the virulence of two Sclerotinia sclerotiorum isolates on sunflower. Physiol Plant Pathol 22:339–345
Marina M, Maiale SJ, Rossi FR, Romero MF, Rivas EI, Gárriz A, Ruiz OA, Pieckenstain FL (2008) Apoplastic polyamine oxidation plays different roles in local responses of tobacco to infection by the necrotrophic fungus Sclerotinia sclerotiorum and the biotrophic bacterium Pseudomonas viridiflava. Plant Physiol 147:2164–2178
Marina M, Vera Sirera F, Rambla JL, Gonzalez ME, Blázquez MA, Carbonell J, Pieckenstain FL, Ruiz OA (2013) Thermospermine catabolism increases the resistance to Pseudomonas viridiflava in Arabidopsis thaliana. J Exp Bot 64(5):1393–1402
Maxwell DP, Lumsden RD (1970) Oxalic acid production by Sclerotinia sclerotiorum in infected bean and in culture. Phytopathol 60:1395–1398
Miché L, Battistoni F, Gemmer S, Belghazi M, Reinhold-Hurek B (2006) Upregulation of jasmonate-inducible defense proteins and differential colonization of roots of Oryza sativa cultivars with the endophyte Azoarcus sp. Mol Plant Microbe Interact 19:502–511
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Nardi CF, Villarreal NM, Rossi FR, Martínez S, Martínez GA, Civello PM (2015) Overexpression of the carbohydrate binding module of strawberry expansin2 in Arabidopsis thaliana modifies plant growth and cell wall metabolism. Plant Mol Biol 88:101–117
Nei M, Kumar S (2000) Molecular evolution and phylogenetics. Oxford University Press, New York
Noyes RD, Hancock JG (1981) Role of oxalic acid in the Sclerotinia sclerotiorum wilt of sunflower Helianthus annuus. Physiol Plant Pathol 18:123–132
Partridge-Telenko DE, Hu J, Livingstone DM, Shew BB, Phipps PM, Grabau E (2011) Sclerotinia blight resistance in Virginia-type peanut transformed with a barley oxalate oxidase gene. Phytopathol 101:786–793
Perini MA, Sin IN, Villarreal NM, Marina M, Powell ALT, Martínez GA, Civello PM (2017) Overexpression of the carbohydrate 1 binding module from Solanum lycopersicum Expansin 1 (Sl-EXP1) modifies tomato fruit firmness and Botrytis cinerea susceptibility. Plant Physiol Biochem 113:122–132
Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29:e45
Pfaffl M, Horgan G, Dempfle L (2002) Relative expression software tool 24 (REST(C)) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res 30:e36
Ramirez V, Agorio A, Coego A, Garcia-Andrade J, Hernandez MJ, Balaguer B, Ouwerkerk PBF, Zarra I, Vera P (2011) MYB46 modulates disease susceptibility to Botrytis cinerea in Arabidopsis. Plant Physiol 155(4):1920–1935
Romero FM, Marina M, Pieckenstain FL (2016) Novel components of leaf bacterial communities of field-grown tomato plants and their potential for plant growth promotion and biocontrol of tomato diseases. Res Microbiol 167:222–233
Romero FM, Rossi FR, Gárriz A, Carrasco P, Ruíz OA (2019) A bacterial endophyte from apoplast fluids protects canola plants from different phytopathogens via antibiosis and induction of host resistance. Phytopathol 109(3):375–383
Rosenblueth M, Martinez-Romero E (2006) Bacterial endophytes and their interactions with hosts. Mol Plant Microbe Interact 19:827–837
Rossi FR, Gárriz A, Marina M, Romero FM, Gonzalez ME, Collado I, Pieckenstain FL (2011) The sesquiterpene botrydial produced by Botrytis cinerea induces the hypersensitive response and its toxicity is modulated by host signaling pathways mediated by salicylic acid and jasmonic acid. Mol Plant Microbe Interact 24:888–896
Rossi FR, Marina M, Pieckenstain FL (2015) Role of arginine decarboxylase (ADC) in Arabidopsis thaliana defence against the pathogenic bacterium Pseudomonas viridiflava. Plant Biol 17:831–839
Ryan RP, Monchy S, Cardinale M, Taghavi S, Crossman L, Avison MB, Berg G, van der Lelie D, Dow JM (2009) The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat Rev Microbiol 7:514–525
Sahin N (2003) Oxalotrophic bacteria. Res Microbiol 154:399–407
Schoonbeek H, Jacquat-Bovet AC, Mascher F, Métraux JP (2007) Oxalate-degrading bacteria can protect Arabidopsis thaliana and crop plants against Botrytis cinerea. Mol Plant Microbe Interact 20:1535–1544
Sneath PH, Sokal RR (1973) Numerical taxonomy. W. H. Freeman and Co, San Francisco
Stone JM, Heard JE, Asai T, Ausubel FM (2000) Simulation of fungal-mediated cell death by fumonisin B1 and selection of fumonisin B1-resistant (fbr) Arabidopsis mutants. Plant Cell 12:1811–1822
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599
Thomma BP, Eggermont K, Penninckx IAMA, Mauch-Mani B, Vogelsang R, Cammue BPA, Broekaert WF (1998) Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. PNAS 95:15107–15111
Thomma BP, Eggermont K, Tierens KFMJ, Broekaert WF (1999) Requirement of functional Ethylene-Insensitive 2 gene for efficient resistance of Arabidopsis to infection by Botrytis cinerea. Plant Physiol 121:1093–1101
Versalovic J, Schneider M, De Bruijn F, Lupski J (1994) Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol Cell Biol 5:25e40
Villarreal NM, Marina M, Nardi CF, Civello PM, Martínez GA (2016) Novel insights of ethylene role in strawberry cell wall metabolism. Plant Sci 252:1–11
Vogel J, Somerville S (2000) Isolation and characterization of powdery mildew-resistant Arabidopsis mutants. PNAS 97:1897–1902
Wildermuth MC, Dewdney J, Wu G, Ausubel FM (2001) Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 414:562–565
Williams B, Kabbage M, Kim HJ, Britt R, Dickman MB (2011) Tipping the balance: Sclerotinia sclerotiorum secreted oxalic acid suppresses host defenses by manipulating the host redox environment. PLoS Pathog 7:e1002107. https://doi.org/10.1371/journal.ppat.1002107
Zhao-Xia J, Wang C, Chen W, Chen X, Li X (2007) Induction of oxalate decarboxylase by oxalate in a newly isolated Pandoraea Sp. OXJ-11 and its ability to protect against Sclerotinia sclerotiorum infection. Can J Microbiol 53:1316–1322
Zhou T, Boland GJ (1999) Mycelial growth and production of oxalic acid by virulent and hypovirulent isolates of Sclerotinia sclerotiorum. Can J Plant Pathol 21:93–99
Acknowledgements
This work was supported by grants of Consejo Nacional de Ciencia y Técnica (PIP 0256 and PIP 0440, Argentina), Agencia Nacional de Promoción Científica y Tecnológica (PICT 2147, Argentina). F.M.R has a post doctoral fellowship of Agencia Nacional de Promoción Científica y Tecnológica. M.M, N.M.V, AG, F.R.R, G.A.M and F.L.P are members of the Research Career of CONICET. The authors are grateful to José Luis Burgos (CIC) and Patricia Alejandra Uchiya (CIC) for their valuable technical assistance.
Author information
Authors and Affiliations
Contributions
MM designed the study. MM, FMR, NMV, AJM, FRR and AG performed the experimental studies. All results and data were analyzed and interpreted by MM, FMR, NMV, AG, GAM and FLP. MM wrote the manuscript with the contribution of GAM and FLP.
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
11103_2019_888_MOESM2_ESM.pdf
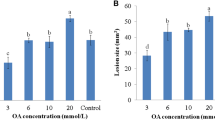
Supplementary Supplemental Fig. 1: Potassium oxalate toxicity on A. thaliana leaves. Five-μl aliquots of potassium oxalate solutions (3, 6 and 20 μM) in water (pH 4.0) were dispensed on both sides of the central vein of the adaxial surface of detached A. thaliana Col-0 leaves, which were placed in Petri dishes containing 0.8% (p/v) water-agar and incubated in a plant growth chamber. Leaves treated with water (pH 4.0) were used as controls. Figure shows the development of the lesions 72 h after potassium oxalate application observed with the naked eye (PDF 467 kb)
11103_2019_888_MOESM3_ESM.pdf
Supplemental Fig. 2: Lesions caused by B. cinerea on tomato leaves inoculated by OxA and OxB. Detached tomato leaves previously inoculated with OxA or OxB cell suspensions were inoculated with B. cinerea conidea (5 x 104 conidia ml−1). The size of the lesion area around the inoculation site was determined at 48 and 72 hpi using the Image-ProPlus V 4.1 software (Media Cybernetics). Results are means of 30 replicates ± SD and different letters indicate statistically significant differences between plant treatment at each time, according to one-way ANOVA and Tukey’s test (PDF 38 kb)
11103_2019_888_MOESM4_ESM.pdf
Supplemental Fig. 3: Direct in vitro confrontation assay between B. cinerea and OxA or OxB. Direct confrontation cultures on PDA medium between fungal pathogen B. cinerea and bacterial isolates OxA or OxB. The cultures were incubated at 25 ºC until 3 and 7 days post-inoculation (PDF 50 kb)
11103_2019_888_MOESM5_ESM.pdf
Supplemental Fig. 4: Lesions caused by B. cinerea and S. sclerotiorum on Arabidopsis Col-0, sid2-2 and coi1 leaves. Detached Arabidopsis leaves were inoculated with (A)B. cinerea conidea (5 x 104 conidia ml−1) or (B)S. sclerotiorum mycelium. Col-0 leaves were used as controls. The size of the lesion area around the inoculation site was determined at 24, 48 and 72 hpi using the Image-ProPlus V 4.1 software (Media Cybernetics). Results are means of 30 replicates ± SD and different letters indicate statistically significant differences between plant lines, according to one-way ANOVA and Tukey’s test (PDF 50 kb)
11103_2019_888_MOESM6_ESM.pdf
Supplemental Fig. 5: Lesions caused by 20 mM potassium oxalate on Arabidopsis Col-0, sid2-2 and coi1 leaves. Five-μl aliquots of potassium oxalate solution (20 μM) in water (pH 4.0) were dispensed on both sides of the central vein of the adaxial surface of detached A. thaliana Col-0, sid2-2 and coi1 leaves, which were placed in Petri dishes containing 0.8% (p/v) water-agar and incubated in a plant growth chamber. The size of the lesion area around the inoculation site was determined at 48 hpt using the Image-ProPlus V 4.1 software (Media Cybernetics). Results are means of 24 replicates ± SD and statistically analyzed by to one-way ANOVA and Tukey’s test (PDF 5138 kb)
11103_2019_888_MOESM7_ESM.pdf
Supplemental Fig. 6: A. thaliana leaves previously inoculated by OxA or OxB. (A) Fluorescence microscopy was used to analyse the accumulation of phenolic compounds (autofluorescence) in Arabidopsis leaves 24 h post-inoculation with OxA or OxB. (B) Fluorescence microscopy was used to analyse callose deposition after aniline-blue staining 6 h post inoculation (hpi). Control leaves were mock-inoculated with MgCl2 10 mM pH 7.0 (PDF 4652 kb)
11103_2019_888_MOESM8_ESM.pdf
Supplemental Fig. 7: AIRs extracted from Arabidopsis leaves inoculated with OxA or OxB. Cell wall polysaccharides were obtained as alcohol insoluble residues (AIR) from Arabidopsis leaves previously (48 h before) inoculated with OxA or OxB. Leaves treated with MgCl2 10 mM pH 7.0 were used as controls. Results are means of 3 replicates ± SEM no statistically significant differences between treatments and controls were found, according to one-way ANOVA and Tukey’s test (PDF 21 kb)
Rights and permissions
About this article

Cite this article
Marina, M., Romero, F.M., Villarreal, N.M. et al. Mechanisms of plant protection against two oxalate-producing fungal pathogens by oxalotrophic strains of Stenotrophomonas spp.. Plant Mol Biol 100, 659–674 (2019). https://doi.org/10.1007/s11103-019-00888-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11103-019-00888-w