
Abstract
Jezioro Lake is the only natural lake in southern Poland outside mountainous areas to have existed continuously since the Pleistocene. The record of environmental change in the Late Vistulian (Weichselian) and Holocene is preserved in the deposits and landforms around the lake. This paper presents the results of paleogeographical and paleoecological research that emabled us to reconstruct the history of the Jezioro Lake. At the end of the Vistulian period, the outlet of the lake was blocked by a parabolic dune moving in from the west. Limnic sedimentation was evident in the sediment core at all levels from the Holocene, with remains of Cladocera, Chironomidae larvae, and aquatic plants. The lake did not disappear at that time, although its area decreased by a factor of 12 by the end of the period. Paleobotanical research permitted the reconstruction of sequences of plant communities and changes in nutrient status and water level. An initial oligotrophic lake, as indicated by the presence of Isoëtes lacustris L., changed to a eutrophic lake, as indicated by the presence of Potamogeton natans L. and Nuphar sp., then the lake progressed to the present-day dystrophic lake that is surrounded by a swamp. The profile of organic deposits contains a record of environmental change at least since the Younger Dryas in southern Poland.
Similar content being viewed by others

Introduction
In Central and Eastern Europe, the majority of lakes that hold a record of changes in environmental conditions from the late Pleistocene until modern times in their sediments are young glacial lakes (Błaszkiewicz 2007). Such lakes can be found from the Mecklenburg Lake District in Germany through northern Poland, Lithuania and north-western Russia to Finland. One of the best studied sediment profiles of this type of lake is from Gościąż Lake (Ralska-Jasiewiczowa et al. 1998, 2003; Goslar et al. 1999). Sediments that are also valuable from the point of view of Late Pleistocene/Early Holocene paleogeography include those of karst lakes situated north of Lublin in Poland (Bałaga 2007, 2010) and in western Belarus, as well as ox-bow lake sediments, e.g. in the Sandomierz Basin (Nalepka 1994; Kołaczek 2010), and those of single lakes left behind by mountain glaciers (Obidowicz 1996). Profiles of those lake sediments have been compared to profiles of peat bogs situated in various regions of Poland (Czyżewska 2005).
The further south we move in Poland, the fewer natural water bodies are present. A peculiar type of lake found there is the aeolian barrier lake. These lakes formed in the late Pleistocene as a result of small valleys being blocked by dunes that moved in from the west and they preserve a rich record of environmental change in their sediments. They formed in sandy areas with poor soils and thus remained outside the settlement zone for a long time. As a result, anthropogenic impact was only reflected in their sediment records at a late stage. Most of these, usually shallow, lakes rapidly became eutrophic. Jezioro Lake on the Silesian-Kraków Upland is one of the few lakes that still remain.
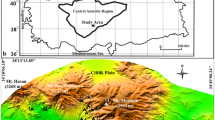
Jezioro Lake is located west of Częstochowa (Fig. 1a). This splendid lake is the only such lake of non-anthropogenic origin in southern Poland outside mountainous areas (Czylok et al. 2004). It lies in forested terrain (Fig. 1b) within the “Lasy nad Górną Liswartą” (“Forests on the Upper Liswarta”) Landscape Park, which is only partly used by traditional agriculture.

Location of study site: a location of Jezioro Lake (limestone structural thresholds—hatched, sandstone structural threshold—dotted); b Jezioro Lake environs; c Bathymetric plan of lake
Earlier research, carried out in 2004, was mainly concerned with the contemporary condition of the lake. That research had an interdisciplinary character that involved surface geological and geomorphological investigations, hydrological mapping, physico-chemical analyses of water, analysis of shallow lake sediments and mapping of contemporary plant communities (Czylok et al. 2004).
The present paper interprets the evolution of the lake based on a study that was carried out in 2006–2009.
The importance of Jezioro Lake deposits in providing complete information on the Late-Vistulian and Holocene transformations of the environment in southern Poland is emphasised by the fact that the nearest site from which similar organic deposits have been described lies in the area of Jaworzno (Szczepanek and Stachowicz-Rybka 2004), located 180 km to the south and this contains a slightly shorter record, reaching only down to the Younger Dryas, i.e. GS-1 according to the stratigraphic division proposed for the North Atlantic region (Björck et al. 1998).
Study area
Jezioro Lake (N 50°47′ 21.33″, E 18°50′ 20.00″) is a natural water body located in the northern part of the Silesian Upland, on the margin between the Herby Ridge, formed by ferruginous sandstones of the Middle Jurassic and the Liswarta Depression formed in Lower Jurassic clays, loams and sands. The surface is formed of Pleistocene glacial, fluvioglacial, fluvial and aeolian deposits and Holocene fluvial deposits 10–20 m thick. Their thickness is locally reduced to a few metres where the Jurassic substratum is elevated (Klimek 1966; Haisig et al. 1983).
The swampy topographic depression in which the lake is located (Figs. 1b, 2a, 2b) drains into a stream that flows into the Liswarta River through a system of ditches. The lake is situated in a coniferous forest that occupies almost 1/3 of its 2.5 km2 catchment (Fig. 1b).

Geomorphic sketch of the Jezioro Lake environs: a Geomorphic sketch (1 fluvioglacial-morainic plateau and kame terrace, 2 fluvioglacial terrace with aeolian cover, 3 Holocene valley floors, 4 dunes, 5 depression without drainage, 6 organogenic plain, 7 slopes, 8 summit of elevations, 9 lake, 10 old spring niche, 11 old bog-spring, 12 topographic profile lines). b Schematic morphological profiles of the Jezioro Lake area; location of morphological profiles—see figure a (L1 old shoreline of lake, L2 modern shoreline of lake). c Schematic morpho-geological cross-section of the shore of Jezioro Lake—without horizontal scale, e–e′ cross-section line is marked on figure a, after Czylok et al. (2004) (a dune, b fragment of outwash plain with aeolian sand covers with different thicknesses, c old lake shore with micro-cliff, d peat-plain, e floating mat, f open water, 1 variable-grained sand, 2 fine-grained sand, 3 sand with humus, 4 organic horizon of forest soil, 5 peat, 6 floating mat, 7 bottom deposits, 8 open water)
Jezioro Lake is a dystrophic lake with water area of 0.027 km2 and a total depth of 1.2–1.5 m (Fig. 1c), of which more than 0.4 m is made up of semiliquid bottom deposits (Fig. 2c). Significant amounts of humic substances in the water give it a brown hue.
In the vicinity of the lake, plant communities are present that are unique within the region and rare in Poland. Among the most valuable are patches of mesotrophic and oligotrophic peat as well as birch wood swamp (Betuletum pubescentis R. Tx.) and adjacent coniferous swamp forest communities (Fig. 3).

Distribution of the main areas of natural vegetation at Jezioro Lake
Methods
Fieldwork
The landforms in the lake catchment were mapped using transects. The geomorphology of the deposits was examined using exploratory boreholes. The relationships between landforms and the deposits forming them were investigated through field observations, and by relating these to the Topographic map of Poland at 1:10,000 scale and the Detailed Geological Map of Poland at 1:50,000 scale. On the transect lines and at other selected points, exploratory boreholes were made to a depth of 3 m, and the macroscopic structural and textural features of the deposits were examined.
The maximum historic extent of the waterline was determined by investigating old micro-cliff and sub-cliff sediment zones. Where the relief of lake banks was less articulated, the waterline was determined by projecting the bases of the micro-cliff across the lake using land surveying equipment.
A bathymetric plan of the contemporary lake was generated using an echo-sounder and range pole. The positions of the measurements were determined by the use of a GPS receiver.
Granulometric composition of the mineral sediments was attained by using a sieve method. Analysis of roundness and degree of matting of quartz grains was conducted using the morphoscopic method of A. Cailleux as modified by Goździk (1980) on the 0.8–1.0 mm fraction. The content of calcium carbonate was determined using a field methodology involving 10 % hydrochloric acid poured on the samples. It was assumed that episodes when the mineral sediments had a better oxygen penetration were marked with an orange-reddish colour from iron compounds.
Plant macroremains and pollen analysis
Core J18 was obtained for peat and pollen analysis using an Instorf Ø 80 peat borer at a location that is presently dry but which is situated along the axis of the former lake basin (Fig. 3). The analysis of macroscopic remains was conducted from samples of 100 cm3 taken every 5 cm along the core. Microscope slides were prepared from this material in order to identify the presence of remains of Cladocera and to identify accurately the features of the deposit. Fresh sediment was soaked with water to separate remains and then washed through a 0.2-mm mesh sieve. This permitted individual remains to be segregated and preserved in a solution of water, glycerine, and alcohol (1:1:1) and some drops of thymol. Some plant remains were washed with alcohol and dried before identification for better demonstration. Determinations were mainly made with a stereoscopic microscope. Paleobotanical keys (Berggren 1969; Cappers et al. 2006; Grosse-Brauckmann 1992; Katz et al. 1965; Velichkevich and Zastawniak 2006; Aalto 1970; Wasylikowa 1986) were used to identify macroremains. The results were compiled in the form of a diagram of macroremains.
Palynology samples were taken from the J18 core at intervals of 2–2.5 cm. Macerated samples were 1 cm3 in volume and typically taken at 5-cm intervals. Depending on the results obtained these intervals were shorter in certain profiles of the core. Maceration agents included 10 % KOH, 10 % HCl, ZnCl2 and Erdtman’s acetolysis was applied (Faegri and Iversen 1989). Two tablets of Lycopodium were added to the samples at the beginning of the maceration process. Calculations were performed on the basis of the sum total of arboreal pollen (AP) and of land-growing non-arboreal pollen (NAP). Results of the pollen analysis are shown on a pollen diagram created using the POLPAL package (Nalepka and Walanus 2003). The main criterion for determining the limits of macrofossil assemblage zones was the appearance or disappearance of one or several species typical for a given level, or the frequent occurrence of species with similar ecological requirements. The layers identified reflect the transformation of vegetation and the phases of development of the water body and its immediate environment that followed.
The pollen diagram has been divided into 6 local pollen assemblage zones (LPAZ) according to Birks (1986) and Janczyk-Kopikowa (1987) supported by numerical analysis (ConSLink).
Radiocarbon dating
A total of 7 samples from the J18 cross-profile were radiocarbon dated (Table 1). The 6 samples taken from gyttja were dated at the Radiocarbon Laboratories in Gliwice and in Skała (Poland). Radiocarbon dates were calibrated with OxCal v4.0.5 (Bronk Ramsey 2001) and the IntCal04 atmospheric curve (Reimer et al. 2004). A sample of clay and silt was dated at the Radiocarbon Laboratory in Kiev (Ukraine).
The J18 profile stratigraphy was based on radiocarbon dating, palynological research and on analyses of plant remains. Chronozones were determined according to Mangerud et al. (1974).
Results
Geological and geomorphological conditions
The most significant surface deposits in the vicinity of Jezioro Lake are sandy to gravely fluvioglacial deposits from the Oder glaciation that are located on a large sandy plain stretching along the Liswarta valley at an elevation of 250–280 m a.s.l. The top layers of the sandy deposits are composed of coarse-grained sands, with a local admixture of gravels.
Sandy fluvioglacial and fluvial deposits, easily blown by wind, provided the source material for fields of aeolian sands that developed into sand dunes of different types. In addition to the plain of aeolian sands, parabolic dunes and longitudinal dunes forming long ridges occur in areas to the west, south, and southeast of Jezioro Lake (Figs. 2a, 2b). Dunes reach heights of 2.5–6 m up to 13–15 m, and the thickness of the aeolian sand cover exceeds 2 m.
An analysis of the remains of old micro-cliffs in the south and west part of the lake revealed that the maximum water level could have reached 272.4 m a.s.l., i.e. 1 m higher than today (Figs. 2a, 2b). In the north and east the former waterline is often indistinguishable. A contour was drawn corresponding to the old waterline, which helped to estimate the maximum water surface of the lake at about 0.32 km2, or 12 times the present surface area.
Among drillings made within the historic lake bowl that is currently filled with sediment, profile J18 (Fig. 3) was selected as the one offering the best potential for plant remains and palynological analyses.
At its lowest level the J18 well contained various-grained sands with an admixture of silt and clay, as well as trace quantities of organic matter. From a depth of 3.61–3.75 m upwards, clay and silty clay with organic matter are found. Sediments from 3.65 to 3.68 m were radiocarbon dated to 12,000 ± 90 BP, i.e. 12,400–11,600 cal. BC (Ki-13020) (Table 1). No pollen was found. Analyses of samples taken from the entire sediment did not contain any calcium carbonate. The clay and silt layers as well as gyttjas directly above them contain 0.3–1.0 cm thick sandy laminae, as well as scattered grains of sand. They are particularly numerous at depths between 3.45 and 3.75 m. The layer at the depth of 3.55 m was particularly pronounced.
The clay and silt layer becomes rusty in colour when it borders on a sandy lamina, which would suggest higher degrees of iron oxidation. Above this layer is clayey and loamy gyttja as well as fine-detritus gyttja 2.34 m thick with no mineral matter. At the top level these gyttjas are covered with a thin layer of peat formed as a result of a floating mat encroaching inwards from the banks. A limnic origin is suggested for the entire deposit series found under the peat layer by the presence of remains of water vegetation and numerous remains of Cladocera and of Chironomidae larvae, which are indicators of lacustrine sedimentation.
The characteristics of the deposits in core J18 are presented in Table 2, and Figs. 4 and 5.
Analysis of plant macroremains
In total, 72 samples were analysed in which 61 plant taxa were recognized. It was possible to identify 43 of these plant taxa down to species (including 5 bryophyte species).
The list of plant macroremains (Fig. 4) enabled the researchers to distinguish 7 local macrofossil assemblage zones (L MAZ) (Table 3).

Plant macrofossil diagram from Jezioro Lake (profile J18): 1 living vegetation, 2 peat, 3 gyttja, 4 clayey gyttja, 5 silty gyttja with laminae of sand, 6 clayey and silty gyttja, 7 clayey silt with laminae of sand, 8 clay and silty clay with laminae of sand, 9 sand with laminae of silt
Within the J18-1 L MAZ level, endocarps of Potamogeton filiformis Pers. (26 in sample No. 72) and P. praelongus Wulfen were the most numerous among aquatic plant remains. Oospores of Chara sp. were also very frequent (more than 900 specimens in sample No. 72). Among rush plant remains, Carex nuts were the most numerous. Within this level, trees and shrubs were represented by Betula remains (including Betula nana L.) and Pinus sylvestris L. Fragments of a Chamaedaphne calyculata (L.) Moench leaf were also found.
Within the J18-2 L MAZ level, Potamogeton natans L. endocarps were the most frequent aquatic plant remains. There were very few P. praelongus, P. filiformis and P. pusillus L. endocarps. Remains of Nymphaea candida C. Presl. testa and Batrachium sp. fruits were also found. Among rush plant remains, Carex rostrata Stokes nuts dominated. Menyanthes trifoliata L., Comarum palustre L. and Schoenoplectus lacustris (L.) Palla seeds were also present. Trees and shrubs were represented by remains of Pinus sylvestris and Betula pubescens Ehrh. (and B. sect. Albae L.). A Betula nana seed and fruit scale as well as a single B. sect. Humilis Schrank fruit were also found.
Within the J18-3 L MAZ level, Potamogeton pusillus endocarps were the most frequent aquatic plant remains (76 in sample No. 63). P. natans, P. praelongus, P. filiformis endocarps, Nuphar pumila (Timm) D.C and Nymphaea candida seeds and testa fragments, single Myriophyllum verticillatum L., Myriophyllum alterniflorum DC. and Ceratophyllum demersum L. fruits as well as single Isoëtes lacustris L. megaspores were also found. Rush plants were few and mostly represented by Carex nuts. Trees were represented by numerous Pinus sylvestris bark fragments and Betula nana and B. pubescens fruits and fruit scales. Single Rhynchospora alba (L.) Vahl seeds and Eriophorum vaginatum L. spindles were also found.
The J18-4 L MAZ level was made distinct by the presence of numerous Isoëtes lacustris megaspores (more than 300 in sample No. 55) and the increase in the number of Chara sp. oospores (more than 900 in sample No. 55). Among the remains of vascular aquatic plants, Batrachium sp. fruits and single endocarps of Potamogeton natans, P. pusillus and P. praelongus were present. Najas minor All., Nuphar pumila and Nymphaea candida seeds and Nymphaea alba L. testa fragments were present as well. Rush plants were represented by Carex nuts. Trees were represented by Pinus sylvestris bark fragments, bud scales, one Pinus sylvestris seed and one Pinus sylvestris cone as well as by single Betula pubescens and B. sect. Albae fruits.
Within the J18-5 L MAZ level, numerous Potamogeton natans endocarps and Chara sp. oospores were found. Single Potamogeton pusillus and P. praelongus endocarps and Najas minor, Nuphar pumila, Nymphaea candida and N. alba seeds were found. Among rush plant remains, Schoenoplectus lacustris seeds were the most numerous. Eleocharis palustris (L.) Roem. and Schult and Sagittaria sagittifolia L. fruits as well as Carex nuts, including C. rostrata, were also found. Peat plants were, inter alia, represented by Scheuchzeria palustris L. leaves, Eriophorum vaginatum spindles, Rhynchospora alba fruits and Menyanthes trifoliata seeds. Trees were represented by diverse and numerous Pinus sylvestris and Betula remains. A single Populus tremula L. bud scale was also found.
Within the J18-6 L MAZ level, aquatic plant macroremains were only present in the bottom part. These were Potamogeton (among others P. natans) endocarps and Chara sp. oospores. Rush plants were represented by single Carex limosa L., C. canescens L., C. acuta L. and C. rostrata nuts. Among peat plant macroremains, numerous Eriophorum vaginatum spindles, Scheuchzeria palustris L. leaf remains and Oxycoccus palustris Pers. leaves were found. Trees were represented by numerous Pinus sylvestris bark fragments and bud scales and Betula (including B. nana) seeds and fruit scales.
Within the J18-7 L MAZ level, no aquatic plant remains were found. Among macroremains, Carex (mainly C. rostrata) nuts generally prevailed. Carex nigra Reichard, C. canescens and C. sect. Paludosae G. Don nuts were also found. Among peat plants, Scheuchzeria palustris, Eriophorum angustifolium Honck and Oxycoccus palustris leaf remains were found. Trees were represented by rare Pinus sylvestris bud scales and Betula pubescens fruits.
Pollen analysis
Within the J18-1 L PAZ level, herbaceous and grass communities with Artemisia, Helianthemum nummularium (L.) Mill. type, Rumex acetosella L. were found, sometimes with Chenopodiaceae pollen and Hippophaë rhamnoides L. In sandy areas, which are common in the vicinity, Juniperus shrubs were found, but also Betula alba type and Pinus sylvestris type In all 5 % of the pollen found was Pinus cembra L., sometimes accompanied by Larix. In diverse habitats in the vicinity of the lake, Salix pentranda L. and Populus, Betula nana and Salix (S. polaris type and Salix undiff.) were present (Table 4).
Within the J18-2 L PAZ level, in Pinus-NAP subzone a, the percentage of forest species sporomorphs (Pinus sylvestris type and Betula alba type) increases, while the percentage of open grass communities falls and Juniperus disappears. Poaceae and Artemisia are still present to some extent.
Within the J18-3 L PAZ level, in Pinus subzone b, pine forest dominated in the vicinity, while Betula alba type was a sporadic element in the forest. Its percentage only rose in Corylus-Ulmus-Betula subzone c, while the percentage of Pinus decreased. Ulmus gradually appeared and reached 4 % of pollen in the late c subphase.
Within the J18-3 L PAZ level, more significant changes can be seen in the vicinity of Jezioro Lake. Forest species dominate—first Pinus sylvestris type and Betula alba type, but with Corylus avellana L. subsequently becoming increasingly significant, gradually reaching a percentage of 27 %. Ulmus also appears with a percentage of more than 6 % alongside Quercus.
Within the J18-4 L PAZ level, mixed forest species are present with Quercus, Tilia (T. cordata type and T. platyphyllos type), Ulmus, Acer, Corylus, Fraxinus excelsior L. and Alnus. Taxus baccata L. is sporadic. Corylus has a significant representation, while there is little Pinus sylvestris type and Betula alba type pollen (2–3 % in the younger part of the level).
Within the J18-5 L PAZ level, no major changes in vegetation are recorded. Quercus is present alongside Tilia cordata type, Taxus, Fraxinus, Acer, Ulmus, Alnus and an admixture of Corylus. New components reaching percentages of pollen of respectively 2 and 2.5 % are Carpinus betulus L. and Fagus sylvatica L. Picea abies (L.) H. Karst. constitutes 5 % of pollen.
Within the J18-6 L PAZ level, Quercus still dominates and percentages of Carpinus betulus and Fagus sylvatica increase, while that of Abies alba Mill is maintained. Alnus is present with a slight downward trend in the upper part of the level. The percentage of Corylus decreases, there is little Ulmus and Tilia (T. cordata type) pollen, and the percentage of NAP increases slightly in the upper part of the profile.
Discussion
History of the environmental changes in Jezioro Lake
The results obtained from the analysis of the cores of organogenic sediment and from the geomorphological studies allowed us to reconstruct the Late Vistulian and Holocene history of Jezioro Lake.
An analysis of the bottom sediments found in J18 reveals that they may have begun to accumulate intermittently, but that the Jezioro Lake never entirely disappeared, although its area has decreased by a factor of 12 between its formation and the present day. The water level in the lake was probably lowered as a result of the construction of drainage ditches, not earlier than the end of the eighteenth century, during the period of the so-called Fryderycjanska colonization.
Late Vistulian period
During the Late Vistulian period, two stages of increased aeolian activity in the Older Dryas (i.e. GI-1d) and Younger Dryas (i.e. GS-1) played a considerable role in the development of the land relief in the Liswarta Depression and beyond in the Silesian Upland and in an eastern part of the Silesian Lowland (Szczypek 1977; Waga 1994). During the Older Dryas period, very strong northwestern winds created sand dunes, some of which moved up valleys that opened to the west. Numerous small valleys became blocked and their drainage hampered. This is likely to have been the case with the basin-shaped valley where the Jezioro Lake is found.
The age of layers at 3.65–3.68 m was determined as 12,400–11,600 cal. BC (Ki-13020) (Fig. 4), which indicates that the initial phase of lake development, i.e. the period of clay and partly silty clay accumulation, could reach as far back as the end of the Bölling or the beginning of the Older Dryas. The varied character of the deposits at the base of the core, sand and admixtures of finer mineral material, suggests that its deposition occurred in an environment of variable supply of terrigenous matter. The sources of this material were fluvioglacial and fluvial sediments of the Liswarta Depression, assisted by aeolian processes of the Late Glacial. This is indicated by the abrasion of quartz grains in medium- and fine-grained sands at the base of the sediment. A significant contribution of rounded and matted grains (RM)—14–26 % and transitional grains of EM type—60–68 % is typical. The predominance of grains of RM and EM types, indicates brisk aeolian activity and short sand transport during the period of sedimentation at the base of the profile.
In the early stages of lake development (J18-1 L MAZ; 3.60–3.75 m) (Fig. 4), Chara sp., Potamogeton filiformis and Potamogeton praelongus algae were abundant and Potamogeton pusillus, Potamogeton natans and Batrachium sp. were scarce. The drop in the count of the P. filiformis and P. praelongus endocarps at a later stage shows that phytocenoses composed of these species declined. The presence, especially in the older profiles, of Potamogeton filiformis suggests that the lake was cool and mesotrophic (Velichkevich and Zastawniak 2006; Kolstrup 1979; Matuszkiewicz 2001) with clear water, abundant in CaCO3 (Bennike et al. 1994) and with a pH higher than 7 (Lang 1994).
In lakes situated in areas where carbonates are abundant in sediments (e.g. mesozoic limestone or morainic clays), cycles of increasing concentration of calcium carbonate in profiles usually coincide with warm and wet periods and with intensive development of aquatic vegetation (De Klerk et al. 2008; Lauterbach et al. 2011). In the case of the Jezioro Lake, calcium carbonate is scarce in local sandy-gravelly fluvioglacial and glacial sediments originating from the Oder glaciation. Carbonates are easily removed from this type of sediment (Bukowska-Jania and Pulina 1999). On the other hand, the presence of CaCO3 within the water body is recorded in the composition of aquatic plant species that grew in the lake in the Older Dryas. This apparent chronological anomaly may be explained by the fact that carbonates only began to be supplied in abundance to the water body during the “stormy”, cold period of the Old Dryas. They came from soil layers near the surface that underwent soil formation processes during the Bölling period (Kowalkowski 1991). Organic soil particles were probably redeposited at the same time, which may explain the presence of material dated to 12,000 ± 90 BP, i.e. 12,400–11,600 cal. BC (Ki-13020) in the sediments of the J-18 profile (correlated with Older Dryas). A similar phenomenon whereby carbonates were redeposited has been observed in Lake Perespilno in eastern Poland. An increase of CaCO3 content in the early Younger Dryas section of the sediments from the lake was an effect of the drop in the groundwater level which stimulated the oxidation of soils in the lake catchment (Goslar et al. 1999). During the late Older Dryas/early Alleröd period the phase when sediments were enriched with CaCO3 was more clearly recorded, inter alia, in Karaśne Lake sediments in eastern Poland (Bałaga 2007).
The water clarity of Jezioro Lake is evidenced by the development of chlorophyta of the genus Pediastrum. At the time, the most numerous were colonies of Pediastrum boryanum var. boryanum (Turp.) Meneghini, characterised by its wide ecological spectrum. Two other species, i.e. P. duplex var. rugulosum Raciborski, considered indicative of large and sparsely vegetated lakes (Komárek and Jankovská 2001), and Pediastrum boryanum var. longicorne Reinsch, known to prefer dystrophic waters (Komárek and Jankovská 2001), were less numerous.
A complex of moss and sedge communities, probably still of an initial nature, developed on the banks. Among the species present, the major role was played by Rhynchospora alba represented mainly by numerous bracts and far fewer seeds. Other plants included Carex canescens and mosses, including Cinclidium stygium Sw. noteworthy for its cold-climate associations (Kłosowski and Kłosowski 2006). Subshrubs were represented by Chamedaphne calyculata (L.) Moench and Oxycoccus palustris. Isolated stands of Betula nana could also be found. Reedbeds consisted mainly of Carex acuta. There were also some minor phytocenoses of Eleocharis palustris and clumps of Juncus on the banks.
The decline of phytocenoses consisting of Potamogeton filiformis and of the Chara sp. algae (J18-2 L MAZ; 3.45–3.60 m) correlated with a growth in importance of Potamogeton natans and Nymphaea candida. A regression of Potamogeton filiformis with a simultaneous broad expansion of Potamogeton natans, a species that often appears at the onset of eutrophication (Arts et al. 1990), would suggest a slight trend towards more eutrophic conditions. However, the most likely cause of the change observed was a drop in CaCO3 concentration and a decline of pH. Within the Jezioro Lake catchment, which mostly includes varieties of silicate, the carbonates that could be leached and subsequently redeposited were exhausted relatively quickly. This is corroborated by the presence of Nymphaea candida which is known in modern times to reach its optimum in mesotrophic water bodies low in calcium, and the appearance of species often associated with the littoral zone of inter-bog lakes and with acid bogs, including Menyanthes trifoliata and Comarum palustre (Kłosowski and Kłosowski 2006).
In reedbed communities the dominating Carex acuta began to give way to the spreading Carex rostrata. New reedbed species included Schoenoplectus lacustris, Iris sp., Sparganium sp. and Equisetum sp., the existence of the latter supported by the presence of its spores. The proportion of remains of Sphagnum sp., which is higher than at lower levels, and the presence of Andromeda polifolia L. together with the presence of Betula nana suggest a continued development of fen in the banks of the lake. The occurrence of mixed-species reedbeds, including Carex rostrata, and the development of floating-leaved plants would suggest a shallow lake, the waters of which might become humified as a result of the surrounding fens.
In the youngest profile of J18-2 L MAZ there is a change to the species composition in the lake vegetation that suggests a change in the habitat conditions. The importance of Chara sp. increased again at the expense of floating-leaved plants. At the time, according to pollen analysis, the lake floristic diversity increased with the occurrence of Myriophyllum spicatum L. and Isoëtes lacustris. A lack of macro-sized remains of reedbed and fen plants indicates that their phytocenoses had undergone a considerable regression. This change suggests rising water level together with increasing oligotrophy.
At 3.55 m a thin layer of sand appears confirming a restart of more intensive aeolian processes. During the Younger Dryas, numerous old dunes were blown out. Newly formed dunes often stopped at vegetation barriers (Waga 1994). This could explain the raising of the water level in the Jezioro Lake.
In addition this part of the J18 profile (the upper profile of the J18-1 NAP L PAZ; 3.365–3.63 m) was correlated with the Younger Dryas in the palynological record (Figs. 4, 5) due to the occurrence of pine and birch pollen spectrums with Betula nana type, Salix polaris Wahlenb. type and Juniperus, as well as a high concentration of Poaceae, Cyperaceae and Artemisia comparable with well dated cores from Wolbrom (Latałowa 1989) and Jaworzno (Szczepanek and Stachowicz-Rybka 2004) nearby. Also noteworthy is a high proportion of chlorophyta of the genus Pediastrum, which is characteristic of late glacial sediments (Leroy et al. 2000; Komárek and Jankovská 2001; Sarmaja-Korjonen et al. 2006).

Simplified pollen diagram from Jezioro Lake (profile J18). Key to lithology as on Fig. 4
The boundary between the Late Glacial (Younger Dryas) and the Holocene (Preboreal chronozone) periods is identified by a drop in non-arboreal pollen (NAP) and Betula nana type pollen and an increase in pollen counts of Pinus sylvestris type and Betula alba L. type. Criteria adopted in identifying this boundary are compatible with criteria adopted for other Polish core profiles (Ralska-Jasiewiczowa and Latałowa 1996).
Preboreal chronozone
Sediments of the J18-2 L PAZ Pinus-Betula level (2.525–3.365 m) were correlated with the Preboreal chronozone (Fig. 5), as pine-birch pollen spectra, containing also small proportions of Poaceae and Artemisia, are typical for the period (Ralska-Jasiewiczowa and Latałowa 1996, Latałowa 1989). In the initial phase of sediment accumulation in the bottom section of the J-18 profile, older humus, which contained little plant pollen, could have been brought in from adjacent areas by wind or water. For these reasons the radiocarbon dating (11,900 ± 290 BP, i.e. 12,810–11,250 cal BC) carried out for fine-detritus sediments found at 3.22–3.25 m was regarded as controversial and was excluded from the paleoecological analysis (Nita and Szymczyk 2010). The age of sediments in this level, at a depth of 2.78–2.85 m, is better described as 9,180 ± 180 BP, i.e. 9,200–7,800 cal BC.
During the early Preboreal chronozone (the middle and upper profile of J18-3 L MAZ; 3.10–3.45 m) the lake was dominated by phytocenoses with Potamogeton pusillus, although initially their contribution was small. At the time, phytocenoses consisting of Myriophyllum spicatum played the leading role as evidenced by the increased occurrence of its pollen. There was also Potamogeton praelongus, as well as Myriophyllum alterniflorum regarded as an indicator of water acidity (Vöge 1988, 1993; Arts 2002; Arts et al. 1990; Eriksson et al. 1983). Its presence indicates a water low in CaCO3 with a pH below 6 (Brandrud 2002).
A subsequent drop in the occurrence of Myriophyllum spicatum pollen indicates that its phytocenoses declined in significance as they were replaced by communities including Potamogeton natans and then P. pusillus. Ceratophyllum demersum also appeared in the lake. The dominance of Potamogeton pusillus and the presence of Ceratophyllum demersum, as well as the expansion of Potamogeton natans earlier on, make it plausible that the level of nutrients in the lake became probably the highest in the lake’s history. Species which are typical for mesotrophic lakes, e.g. Myriophyllum verticillatum, Nymphaea candida and Nuphar pumila, appear in the youngest profile of the horizon. These species would suggest that the lake water became more oligotrophic again. Numerous colonies of Pediastrum, primarily P. boryanym var. boryanym, continue to be present in the lowest profile attributed to the earliest period of the Preboreal chronozone, but then their significance declined rapidly. Reedbed communities began to re-establish themselves along the banks, but their species composition changed and diversity decreased. In the lowest profile of the horizon Carex species, including Carex rostrata, played a significant role, while only a minor role was played by Typha sp., Phragmites australis (Cav.) Trin. ex Steud., Equisetum fluviatile and Cladium mariscus (L.) Pohl. The occurrence of C. mariscus, which is currently normally found in reedbeds of mesotrophic lakes and less frequently in oligotrophic or dystrophic lakes (Kłosowski and Kłosowski 2006), may confirm a lower water level at that stage. The appearance on the banks of more temperature demanding species such as Typha sp. (Kolstrup 1979; Isarin and Bohncke 1999) indicates better climatic conditions.
Initially, the fen surrounding the lake consisted primarily of Eriophorum vaginatum and Rhynchospora alba. The lack of Sphagnum remains and of characteristic subshrubs in the oldest section of the horizon may mean that the fens were of loose, clump-like nature. Mosses only appear to gain in significance in the fen in a younger profile of the level.
Significant changes to the Jezioro Lake biocenosis occurred in the oldest profile of the level (J18-4 L MAZ; 2.65–3.10 m). Phytocenoses with Potamogeton pusillus declined while Isoëtes lacustris, a species represented here by mega and micro spores, expanded rapidly. It reached its maximum count in the middle of the horizon. The advent of Isoëtes brought with it a disappearance of Pediastrum and an increased role of Chara sp. algae. Small numbers of Potamogeton praelongus, Najas minor and Batrachium sp. were found, and in shallower sections Potamogeton natans, Nuphar pumila, Nymphaea candida, were also found as well as, at a later stage, Nymphaea alba.
The expansion of Isoëtes lacustris suggests a trend towards oligotrophy and the beginning of a soft-water oligotrophic lake phase. The water could have had an acidity similar to modern-day lakes with similar phytocenoses at 5–6.5 pH (Arts 2002). In the youngest profile of the horizon there was a breakdown in the population of Isoëtes lacustris and an expansion of Potamogeton natans. This signifies yet another change in the habitat conditions at the end of the Preboreal and beginning of the Boreal periods.
Reedbed vegetation was probably poorly developed and did not form a continuous strip. The prime roles were played by Phragmites australis and Carex sedges. The lack of any remains of fen-specific vascular plants means that the fen lobes existing at the time on the banks of the lake had severely deteriorated.
In the upper profile of the horizon (subhorizon J18-2c L PAZ Corylus-Ulmus-Betula) pollen of Corylus increased to 5 % and of Ulmus to 4 %, which suggests that the sediments date back to the transition between the Preboreal and Boreal chronozones (Miotk-Szpiganowicz et al. 2004).
Boreal chronozone
Accumulation of homogenous fine-detritus gyttja without any soil-forming admixture (J18-5 L MAZ; 2.15–2.65 m) (Fig. 4) reflects stabilised sedimentation conditions in the lake. At this horizon Isoëtes lacustris was only sporadic. One of the possible causes for its regression could be an expansion of communities involving Potamogeton natans and other floating-leaved plants (Nuphar pumila, N. lutea Sm., Nymphaea alba and N. candida). As they grew, they left the bottom in the shallow parts of the lake in their shadow thus possibly eliminating I. lacustris from its previous habitats. Submergent macrophytes were sparsely represented by Potamogeton pusillus, P. praelongus and Najas minor. The presence in the lake of species typical of oligotrophic waters (Isoëtes lacustris), mesotrophic waters (Najas minor, Nuphar pumila and Nymphaea candida) and plants with wider ecological spectrums (Potamogeton pusillus and Nymphaea alba) leads to the conclusion that the lake water was low in calcium carbonate and mesotrophic.
Reedbeds were expanding with Phragmites australis, Schoenoplectus lacustris and Carex plants playing a significant role. Other species included Eleocharis palustris, Sagittaria sagittifolia together with the Equisetum sp. and Sparganium sp. found in pollen analysis. The expansion of varied reedbed communities and the growth of floating-leaved plants may indicate a shallowing phase in the lake, possibly as a result of gyttja sedimentation.
A comparison of pollen curves from the J18-3 L PAZ Corylus-Ulmus profile and the floor profile of the J18-4 L PAZ Corylus-Alnus-Quercus-Tilia (1.82–2.375 m) with the curves from Wolbrom (Latałowa 1989) and Jaworzno (Szczepanek and Stachowicz-Rybka 2004) helps to correlate the former two with the Boreal chronozone. The evidence that leads to this conclusion mainly includes the high pollen counts of Corylus (27 %) in the J18-3 L PAZ horizon and the existence of continuous pollen curves of Quercus and Alnus in the single percentage range.
The boundary between the Boreal and Atlantic chronozones in the profile studied is marked by the radiocarbon date of 8,000 ± 240 BP (at a depth of 2.26–2.29 m) and by a marked increase in the pollen of Quercus, Alnus and Tilia cordata Mill. type in an underlying sample at 2.35 m. A comparison with other Polish sites (Latałowa 1989; Ralska-Jasiewiczowa et al. 2004) would suggest that these sediments belong to the Atlantic chronozone.
Atlantic chronozone
In a younger part of the zone J18-5 L MAZ (2.15–2.65 m), correlated with the Atlantic chronozone (Fig. 4), the lake plant life is clearly less abundant. The only phytocenoses of high significance include those composed of the Chara sp. algae accompanied with sparse occurrence of Potamogeton natans, Nymphaea alba and Nuphar lutea.
The surrounding fen grew extensively, as is evident from the existence of species such, as Rhynchospora alba, Scheuchzeria palustris, and Eriophorum vaginatum. Other species included Menyanthes trifoliata, Comarum palustre and Potentilla sp. The appearance of Menyanthes trifoliata communities, probably in the contact zone between the fen and the lake water, suggests that towards the end of the accumulation of this horizon the fen expanded, by the extension of floating mats, and the lake underwent a process of progressive dystrophy. The dating of the sample from the depth of about 1.90 m (4,740 ± 80, i.e. 3,660–3,360 cal BC) suggests that these sediments had already accumulated during the Subboreal chronozone.
According to palynological assessment, the boundary between the Atlantic and Subboreal chronozones, which coincides with the upper boundary of the J18-4 L PAZ Corylus-Alnus-Quercus-Tilia (1.82–2.375 m), is marked by a decline in the pollen of Ulmus and the beginning of a continuous percentage curve of Carpinus and Fagus (Nita and Szymczyk 2010), similar to that occurring in other profiles from this part of Poland (Latałowa 1989; Szczepanek and Stachowicz-Rybka 2004).
Subboreal chronozone
Palynological analysis and an analysis of macro-sized remains show that during the Subboreal chronozone (J18-6 L MAZ excluding the oldest profile; 0.40–2.15 m) (Fig. 4), the shallowest profile of the lake was only populated by communities containing Nuphar lutea and phytocenoses based on Chara sp. A growing proportion of remains of Sphagnum sp. and the occurrence of species such, as Carex limosa, Eriophorum vaginatum, E. angustifolium, Scheuchzeria palustris and Oxycoccus palustris attests to a continued development and expansion of fen phytocenoses and the growth of floating mats that gradually encroached on the lake. This is also confirmed by the decline of reedbed communities composed of Phragmites australis, Sparganium sp. and Typha latifolia L. which are underrepresented here. The expansion of fen was accompanied by a progressive dystrophy of the lake.
The correlation of the J18-6 L PAZ Carpinus-Fagus-Abies (0.50–1.525 m) with the Subboreal chronozone proposed by Nita and Szymczyk (2010) is confirmed by radiocarbon dating of sediments at about 1.65 m (at 3,550 ± 90, i.e. 2,140–1,660 cal BC) and at about 1.20 m (at 3,650 ± 230, i.e. 2,670–1,490 cal BC).
Sediments of the youngest horizon (J18-7 L MAZ; 0.15–0.40 m), represented by peat, show that in the part of the lake where the profile was taken, the expanding floating mats filled the lake completely and fen flourished with species, such as Eriophorum vaginatum, E. angustifolium, Rhynchospora alba, Scheuchzeria palustris, Oxycoccus microcarpus Turcz. ex Rupr. and O. palustris.
Conclusions
The rich fossil material, including pollen and macro-sized plant remains, demonstrates that the accumulation of deposits in the Jezioro Lake has continued ever since its beginning in the Late Glacial period. Research carried out on macroremains allows demonstrated 7 phases of development of Jezioro Lake, depending on the composition of the phytocoenoses present on the site, degree of eutrophication, and pH-reaction.
The lake sediments also contain a record of extreme geomorphologic activity, i.e. two periods of aeolian activity. The accumulation of sands at the mouth of the valley made the original development of the lake possible. The existence of sandy layers on the bottom of the existing water body and the presence of a sizable parabolic dune retaining the lake to the west seem to point to the cause of the increase in the water level during the Younger Dryas.
The lithological and paleobotanical record found in the sediments, which is continuous since the Younger Dryas, makes this site important for the history of the Holocene in southern Poland and beyond.
During the Holocene, the Jezioro Lake watershed was not deforested, and the water balance of the lake did not change at least until the so-called Fryderycjanska colonization (late eighteenth century). The transformation related to colonisation was limited, however, because no significant traces of anthropogenic influence have been found in the profiles, perhaps because settlement in the vicinity proved unsuccessful.
Throughout the Holocene, the forest and the fen that surrounded the lake played an important role from the point of view of its water and shores, forming biogeochemical buffers that enabled certain species to survive for a long time, inter alia Isoëtes lacustris (until the end of the Atlantic chronozone) and Betula nana (until the beginning of the Subboreal chronozone). The dystrophication process affecting the lake started during the Atlantic chronozone.
References
Aalto M (1970) Potamogetonaceae fruits. I: Recent and subfossil endokarps of the Fennoscandian species. Acta Bot Fenn 88:1–85
Arts GHP (2002) Deterioration of Atlantic soft water macrophyte communities by acidification, eutrophication and alkalinisation. Aquat Bot 73:337–393
Arts GHP, van der Velde G, Roelofos JGM, van Swaay CAM (1990) Successional changes in the soft water macrophyte vegetation of (sub)Atlantic, sandy lowland regions during this century. Freshw Biol 24:287–294
Bałaga K (2007) Changes in the natural environment recorded in the sediments of the Karaśne Lake-Mire complex (Lublin Polesie, E Poland). Geochronometria 29:1–21
Bałaga K (2010) Przemiany krajobrazu Pojezierza Łęczyńsko-Włodawskiego w okresie 13 000 lat: wyniki badań palinologicznych. In: Chmielewski T, Piasecki D (eds) The future of hydrogenic landscapes in European Biosphere Reserves. Przyszłość krajobrazów hydrogenicznych w rezerwatach biosfery Europy. UP Lublin, Poleski PN, PAN Oddz. Lublin, Komitet Narodowy UNESKO-MAB Warszawa. Lublin, pp 163–190
Bennike O, Houmark-Nielsen M, Bocher J, Heinberg EO (1994) A multi-disciplinary macrofossil study of Middle Weichselian sediments at Kobbelgard, Mon, Denmark. Paleogeogr Paleoclimatol Paleoecol 111:1–15
Berggren G (1969) Atlas of seeds and small fruits of Northwest-European plant species with morphological descriptions. Part 2, Cyperaceae. Swedish Museum Natural History, Stockholm
Birks HJB (1986) Numerical zonation, comparison and correlation of Quaternary pollen-stratigraphical data. In: Berglund BE (ed) Handbook of Holocene Paleoecology and Paleohydrology. Wiley, Chichester, pp 743–774
Björck S, Walker MJC, Cwynar LC, Johnsen S, Knudsen KL, Lowe JJ, Wohlfarth B, INTIMATE Members (1998) An event stratigraphy for the last termination in the North Atlantic region based on the Greenland ice-core record: a proposal by the INTIMATE group. J Quat Sci 13(4):283–292
Błaszkiewicz M (2007) Geneza i ewolucja mis jeziornych na młodoglacjalnym obszarze Polski—wybrane problemy. Studia Limnologica et Telmatologica 1(1):5–16. [Origin and evolution of lake basins on early post-glacial landscape—selected issue (in Polish)]
Brandrud TE (2002) Effects of liming on aquatic macrophytes, with emphasis on Scandinavia. Aquat Bot 73:395–404
Bronk Ramsey C (2001) Development of the radiocarbon calibration program OxCal. Radiocarbon 43(2A):355–363
Bukowska-Jania E, Pulina M (1999) Calcium carbonate in deposits of the last Scandinavian glaciation and contemporary chemical denudation in western Pomerania—NW Poland, in the light of modern processes in Spitsbergen. Z Geomorphol Suppl 119:27–41
Cappers RTJ, Bekker R. M, Jans JEA (2006) Digitale zadenatlas van Nederland (digital seed atlas of The Netherlands). Groningen Archaeol Stud 3, Barkhuis and Groningen University Library, XXVI, Groningen
Czylok A, Fajer M, Machowski R, Szymczyk A, Waga JM (2004) Naturalny zbiornik wodny w Jeziorze (zlewnia górnej Liswarty)—charakterystyka uwarunkowań środowiskowych In: Jankowski AT, Rzetala M (eds) Lakes and artificial water reservoirs—functioning, revitalization and protection. University of Silesia—Faculty of Earth Sciences, Pol Limnol Soc, Pol Geogr Soc—Branch Katowice. Sosnowiec, pp 39–50. [Natural water reservoir in Jezioro (upper Liswarta catchment)—characteristics of the environmental conditions (in Polish)]
Czyżewska K (ed) (2005) Postglacial vegetation changes in the development of raised mires in Poland. Monogr Bot 94, Łódź
De Klerk P, Helbig H, Janke W (2008) Vegetation and environment in and around the Reinberg basin (Vorpommern, NE Germany) during the Weichselian late Pleniglacial, Lateglacial, and Early Holocene. Acta Palaebot 48(2):301–324
Eriksson F, Hörnström E, Mossberg P, Nyberg P (1983) Ecological effects of lime treatment of acidified lakes and rivers in Sweden. Hydrobiologia 101:145–164
Faegri K, Iversen J (1989) Textbook of pollen analysis. Wiley, Chichester
Goslar T, Ralska-Jasiewiczowa M, van Geel B, Łącka B, Szeroczyńska K, Chróst L, Walanus A (1999) Anthropogenic changes in the sediment composition of Lake Gościąż (central Poland) during the last 330 yrs. J Paleolimnol 22:171–185
Goździk J (1980) Zastosowanie morfometrii i graniformametrii do badań osadów w kopalni węgla brunatnego Bełchatów. Studia Regionalne IV:101–114 (in Polish)
Grosse-Brauckmann G (1992) Über pflanzliche Makrofossilien mitteleuropäischer Torfe. III. Früchte, Samen und einige Gewebe (Fotos von fossilen Pflanzenresten). Telma 22:53–102
Haisig J, Kotlicki S, Wilanowski S, Żurek W (1983) Objaśnienia do Szczegółowej Mapy Geologicznej Polski, ark. Boronów, 1:50 000. Wydawnictwa Geologiczne, Warszawa. [Explanations for the detailed geological map of Poland, sheet Boronów, 1:50,000 (in Polish)]
Isarin RFB, Bohncke SJP (1999) Mean July temperatures during the Younger Dryas in north-western and central Europe inferred from climate indicator plant species. Quat Res 51:158–173
Janczyk-Kopikowa Z (1987) Uwagi na temat palinostratygrafii czwartorzędu. Kwart Geol 31(1):155–162 (in Polish)
Katz NJ, Katz SV, Kipiani MG (1965) Atlas i opredelitel plodov i semyan, vstrechayushchikhsya v chetvertichnykh otlozheniyakh SSSR. Nauka, Moscow. [Atlas and key to fruits and seeds occurring in the quaternary deposits of the Soviet Union (in Russian)]
Klimek K (1966) Deglacjacja północnej części Wyżyny Śląsko-Krakowskiej w okresie zlodowacenia środkowopolskiego. Pr Geogr IG PAN 53:1–136. [Deglaciation of the northern part of the Silesian-Cracow Upland in the period of the Middle Polish Glaciation (in Polish)]
Kłosowski S, Kłosowski G (2006) Rośliny wodne i bagienne. Multico Oficyna Wydawnicza, Warszawa. [Water and marsh plants (in Polish)]
Kołaczek P (2010) The development of Late Glacial and Holocene vegetation and human impact near Grodzisko Nowe in the Lower San Valley (Sandomierz Basin, Poland SE). Acta Palaeobot 50(2):101–117
Kolstrup E (1979) Herbs as July temperature indicators for parts of the Pleniglacial and the Late-Glacial in The Netherlands. Geol Mijnb 59:337–380
Komárek J, Jankowská V (2001) Review of green algal genus Pediastrum, implication for pollen-analytical research. Bibl Phycol 108. Berlin, Stuttgart
Kowalkowski A (1991) Ewolucja gleb w holocenie. In: Starkel L (ed) Geografia Polski. Środowisko przyrodnicze. PWN, Warszawa, pp 127–139 (in Polish)
Lang G (1994) Quartare Vegetationsgeschichte Europas. Gustav Fischer Verlag, Jena
Latałowa M (1989) Type region P-h: the Silesia-Cracow Upland. Acta Palaeobot 29(2):45–49
Lauterbach S, Brauer A, Andersen N, Danielopol DL, Dulski P, Hüls M, Milecka K, Namiotko T, Plessen B, von Grafenstein U, DecLakes participants (2011) Multi-proxy evidence for early to mid-Holocene environmental and climatic changes in northeastern Poland. Boreas 40(1):57–72
Leroy SAG, Zolitschka B, Negendank JFW, Seret G (2000) Palynological analyses in the laminated sediment of Lake Holzmaar (Eifel, Germany): duration of Lateglacial and Preboreal biozones. Boreas 29:52–71
Mangerud J, Andersen ST, Berglund BE, Donner J (1974) Quaternary stratigraphy of Norden, a proposal for terminology and classification. Boreas 3:109–126
Matuszkiewicz W (2001) Przewodnik do oznaczania zbiorowisk roślinnych Polski, Vademecum Geobotanicum. Polish Scientific Publishers PWN, Warszawa. [Guide for determination of plant communities of Poland, Vademecum Geobotanicum (in Polish)]
Miotk-Szpiganowicz G, Zachowicz J, Ralska-Jasiewiczowa M, Nalepka D (2004) Corylus avellana L.—Hazel. In: Ralska-Jasiewiczowa M, Latałowa M, Wasylikowa K, Tobolski K, Madeyska E, Wright HE Jr, Turner C (eds) Late Glacial and Holocene history of vegetation in Poland based on isopollen maps. Szafer Institute of Botany, Polish Academy Science, Cracow, pp 79–87
Nalepka D (1994) Historia roślinności w zachodniej części Kotliny Sandomierskiej w czasie ostatnich 15 000 lat. Wiad Bot 38(3–4):95–105. [The history of vegetation in the western part of sandomierz basin during the last 15,000 years (in Polish with summary in English)]
Nalepka D, Walanus A (2003) Data processing in pollen analysis. Acta Palaebot 43(1):125–134
Nita M, Szymczyk A (2010) Vegetation changes in the Jezioro Lake on the background of the Holocene history of forests, Woźniki-Wieluń Upland. Poland. Acta Palaeobot 50(2):119–132
Obidowicz A (1996) A Late Glacial-Holocene history of the formation of vegetation belts in the Tatra Mts. Acta Palaeobot 36(2):159–206
Ralska-Jasiewiczowa M, Latałowa M (1996) Poland. Paleoecological events during the last 15,000 years. In: Berglund BE, Birks HJB, Ralska-Jasiewiczowa M, Wright HE (eds) Regional synthesis of paleoecological studies of lakes and mires in Europe, Wiley, Chichester, pp 403–472
Ralska-Jasiewiczowa M, Goslar T, Madeyska T, Starkel L (eds) (1998) Lake Gościąż, central Poland. A monographic study. Part 1. Szafer Institute of Botany, Polish Academy Science, Cracow
Ralska-Jasiewiczowa M, Goslar T, Różanski K, Wacnik A, Czernik J, Chrost L (2003) Very fast environmental changes at the Pleistocene/Holocene boundary, recorded in laminated sediments of Lake Gościąż, Poland. Paleogeogr Paleoclimatol Paleoecol 193:225–247
Ralska-Jasiewiczowa M, Latałowa M, Wasylikowa K, Tobolski K, Madeyska E, Wright HE Jr, Turner CH (eds) (2004) Late Glacial and Holocene history of vegetation in Poland based on isopollen maps. Szafer Institute of Botany, Polish Academy Science, Cracow
Reimer PJ, Baillie MGL, Bard E, Bayliss A, Beck JW, Bertrand C, Blackwell PG, Buck CE, Burr G, Cutler KB, Damon PE, Edwards RL, Fairbanks RG, Friedrich M, Guilderson TP, Hughen KA, Kromer B, McCormac FG, Manning S, Bronk Ramsey C, Reimer RW, Remmele S, Southon JR, Stuiver M, Talamo S, Taylor FW, van der Plicht J, Weyhenmeyer CE (2004) IntCal04 terrestrial radiocarbon age calibration, 0–26 cal kyr BP. Radiocarbon 46:1029–1058
Sarmaja-Korjonen K, Seppänen A, Bennike O (2006) Pediastrum algae from the classic late glacial Bølling Sø site, Denmark: response of aquatic biota to climate change. Rev Paleobot Palynol 138(2):95–107
Szczepanek K, Stachowicz-Rybka R (2004) Late Glacial and Holocene vegetation history of the “Little Desert” dune area of the south-eastern Silesian Upland, southern Poland. Acta Palaeobot 44(2):217–237
Szczypek T (1977) Utwory i procesy eoliczne w północnej części Wyżyny Śląskiej. University of Silesia Press, Katowice. [Aeolian processes and deposits in the northern part of the Silesian Upland (in Polish)]
Velichkevich FY, Zastawniak E (2006) Atlas of the Pleistocene vascular plant macrofossils of Central and Eastern Europe (Part 1—Pteridophytes and monocotyledons). Szafer Institute of Botany, Polish Academy Science, Cracow
Vöge M (1988) Tauchuntersuchungen der submersen Vegetation in skandinavischen Seen unter Berücksichtigung der Isoetiden-Vegetation. Limnologica 19:89–107
Vöge M (1993) Tauchexkursionen zu Standorten von Myriophyllum alternifolium DC. Tuexenia 13:91–108
Waga JM (1994) Rzeźba eoliczna na obszarze wschodniej części Niecki Kozielskiej. Scripta Rudensia, 2. Park Krajobrazowy “Cysterskie Kompozycje Krajobrazowe Rud Wielkich”, Rudy Wielkie. [Aeolian relief in the eastern part of the Koźle Basin area (in Polish)]
Wasylikowa K (1986) Analysis of fossil fruits and seeds. In: Berglund BE (ed) Handbook of Holocene paleoecology and paleohydrology. Wiley and Sons Ltd, Chichester-New York, pp 571–590
Acknowledgments
We would like to thank Dr Martin Cahn for correcting the English language of the manuscript. We also thank the editors and anonymous reviewers for constructive remarks. The scientific work in this research project no. N N306 071740 was financed by research funds allocated in the years 2011–2013.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Fajer, M., Waga, J.M., Rzetala, M. et al. The Late Vistulian and Holocene evolution of Jezioro Lake: a record of environmental change in southern Poland found in deposits and landforms. J Paleolimnol 48, 651–667 (2012). https://doi.org/10.1007/s10933-012-9634-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-012-9634-1