
Abstract
The analysis of microbotanical remains (starch grains and phytoliths) from food-related domestic contexts and artefacts has the potential to provide insights into daily plant preparation and consumption activities. In particular, pottery vessels offer an unparalleled comparative framework for the study of food preparation and consumption, since pottery is used for a variety of domestic food-related activities, including storage, processing and serving. This study illustrates the potential of microbotanical remains to provide a holistic approach to the plant food preparation and consumption cycle—from storage to disposal—through the analysis of starch grains and phytoliths from Late Minoan storage, cooking and serving vessels recently uncovered at the Knossian ‘neighbourhood’ of Lower Gypsades. The results show that starch grains and phytoliths are more abundant and diverse in cooking vessels, presumably reflecting a higher deposition of microbotanical remains as a result of the disturbance caused by cooking. The results further offer insights into Minoan storage and cooking practices at Gypsades, suggesting that cereals were stored de-husked in pithoi and, possibly, in palm leaf baskets, and later cooked together with non-staple plants. The virtual absence of microbotanical remains from serving vessels may be due to depositional factors and/or the fact that some of the vessels analysed in this study were used for serving/consuming liquids or animal products, and not plants.
Similar content being viewed by others

Introduction
Domestic plant food–related activities can be divided into initial preparation (storage, de-husking), preconsumption preparation (grinding, soaking, cooking, etc.), consumption and disposal (Twiss 2012), although the latter often takes place outside the domestic domain. Macrobotanical research has mostly focused on the initial preparation of plant foods, for example through the analysis of grain–chaff–weed ratios in domestic contexts (e.g. Alonso et al. 2008). Since plant remains are virtually always destroyed during consumption (Samuel 1996: 488), macroscopic evidence of preconsumption preparation and consumption is rarely preserved in the archaeological record, thus impeding a holistic archaeobotanical approach to domestic food–related activities.
Plant preparation and consumption activities, however, also leave microscopic traces in domestic contexts and on artefacts used for food storage, processing and consumption. Thus, the analysis of microbotanical remains (starch grains and phytoliths) from food-related domestic contexts (i.e. areas used for preparing and consuming food) and artefacts such as grinding stones and cooking vessels has the potential to provide a more nuanced understanding of daily plant preparation and consumption activities. In particular, pottery vessels offer an unparalleled comparative framework for the study of food preparation and consumption, since pottery is used for a variety of domestic food–related activities, including storage, processing and serving.
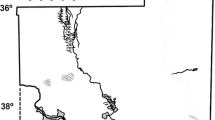
Most microbotanical studies from archaeological pottery focus exclusively on the analysis of cooking vessels (Boyd et al. 2006, 2008; Crowther 2005; García-Granero et al. 2018; Hart et al. 2003, 2007; Musaubach and Berón 2017; Perry et al. 2007; Saul et al. 2012, 2013; Staller and Thompson 2002; Thompson et al. 2004; Yang and Jiang 2010; Yang et al. 2012a, 2014a; Zarrillo et al. 2008), mostly due to the exceptional preservational environment provided by charred food remains (food crust) attached to the vessels (Saul et al. 2012). Pottery vessels used for other food-processing activities such as grinding, grating or sieving have received comparatively less attention (for an example see Dickau et al. 2012), and microbotanical analyses from storage, serving and consumption vessels are extremely rare (but see Pető et al. 2013 for an exception), thus limiting the potential of microbotanical remains to provide a holistic approach to domestic food–related activities. This study attempts to illustrate the potential of microbotanical remains to provide a holistic approach to the plant food preparation and consumption cycle—from storage to disposal. To this end, we analyse microbotanical remains from storage, cooking and serving vessels recovered from a Late Minoan IA (c. 1550–1500 BC) midden deposit in the Knossian ‘neighbourhood’ of Lower Gypsades (Crete) (Fig. 1). The Gypsades hill is an area of the Bronze Age city of Knossos located between the palace to the north and the cemetery area to the south, marking the southern limits of habitation. The samples analysed in this study come from excavations conducted in 2014 by members of the Knossos-Gypsades project, a synergasia between the Herakleion Ephoreia and the British School at Athens directed by Ioanna Sepretsidaki, Eleni Hatzaki, Amy Bogaard and Gianna Ayala under the auspices of the British School at Athens (Morgan 2015: 34–35).

Map of Crete showing the location of the main cities and the site of Knossos
A Holistic Approach to Domestic Food–Related Activities Through the Analysis of Microbotanical Remains from Pottery Vessels
Starch grains are produced in the edible plant parts (seeds, fruits, underground storage organs, etc.) and are usually regarded as a direct evidence of the consumption of starchy plants, including, among other, cereals, pulses and tubers (Torrence and Barton 2006). Size and shape of starch granules are highly variable and influenced by factors such as the part of the plant where they originated, their age and the general nutritional status of the plant. Regardless of this variability, size, shape and other features of starch granules (e.g. the presence of lamellae, the position of the hilum, etc.; see García-Granero et al. 2017: Table 2) can be diagnostic of the plant species in which they occur, facilitating their identification and classification. Experimental work shows that different processing and cooking techniques (brewing, grinding, soaking, boiling, etc.) produce different types of damage to starch grains from domestic cereals and pulses (Henry et al. 2009; Valamoti et al. 2008; Wang et al. 2017). Although more experimental work is needed to confirm this trend in these and other plant taxa, the existing experimental research suggests that starch grains could be a proxy to identify not only the ingredients of ancient cuisines but also the culinary methods employed by prehistoric populations.
Phytoliths form as a result of the deposition of soluble silica in living plants and are liberated when plants die and decay (Piperno 2006). Phytoliths are found mostly in aerial plant structures (leaves, culms, inflorescences, etc.), although certain plant taxa also silicify their underground storage organs (e.g. manioc, see Chandler-Ezell et al. 2006). Not all plants produce phytoliths, and not all plants that produce phytoliths do so in the same way. For example, grasses (Poaceae) produce phytoliths in non-edible parts, especially in the epidermis of the inflorescence bracts (glumes, lemmas and paleae), but not in the edible caryopses, whereas squashes and gourds (Cucurbitaceae) produce diagnostic phytoliths in fruit rinds (Ball et al. 2016: Table 1). Therefore, phytoliths from different plant species have the potential to inform about past culinary practices in different ways, from the initial preparation of cereals (e.g. de-husking) to the consumption of fruits such as squashes and gourds, among others.
In order to reconstruct domestic food–related activities through the study of microbotanical remains from pottery vessels, we must first consider how the different use of each type of vessel (storage, processing, serving) may affect the deposition and preservation of microbotanical remains. It is often assumed that mechanical actions such as grinding or physical disturbances such as boiling trap microbotanical remains in the pores and crevices of artefacts, thus protecting them from enzymatic degradation (Haslam 2004). Hypothetically, the higher the porosity of the artefact and the more intense the disturbance (e.g. longer grinding), the more residues will be trapped in the artefact. At the same time, however, microbotanical remains subjected to physical pressures for a long period of time may lose diagnostic features or even become completely unrecognisable (Henry et al. 2009: Figs. 2 and 3; Portillo et al. 2017). Table 1 describes the hypothetical deposition of microbotanical remains in pottery vessels used for different domestic food–related activities. Since a great number of phytoliths are produced in the epidermis of inflorescence bracts of grasses, a distinction has been made in Table 1 between hulled cereals, on the one hand, and clean (de-husked) cereals and other taxa, on the other. Even after de-husking, cereal grains retain a certain amount of phytoliths (Portillo et al. 2017), so the mere presence of grass phytoliths does not necessarily imply that de-husking has taken place. We may expect higher quantities and proportions of phytoliths from cereal inflorescences in vessels used to store, process or consume hulled cereals, whereas vessels used for clean cereals or other taxa should contain comparatively less phytoliths, and from a more varied taxonomical and anatomical origin.

Starch grains recovered from samples analysed in this study. a Triticeae type A and type B starch grains from sample 30055-2. b cf. Panicoideae starch grain from sample 30055-5. c UNID type starch grains from sample 30055-2. All pictures are under unpolarised (left) and polarised (right) light. Scale bars 20 μm

Length of Triticeae starch grains from Gypsades (vessel 30055-2) compared with starch grains from modern Hordeum vulgare (barley) and Triticum spp. (wheat): a boxplot and b results of the one-way ANOVA (statistically significant differences are highlighted in bold)
In storage vessels plants would not be exposed routinely to mechanical damage, and therefore very few microbotanical remains would be deposited in the pores of the vessel. If cereals were stored hulled, however, a certain amount of grass inflorescence phytoliths may be present. In vessels used for cooking and other plant-processing activities (such as grating or grinding, e.g. Dickau et al. 2012), on the other hand, the repeated disturbance caused by such activities would account for a comparatively richer microbotanical assemblage. A relatively high proportion of the starch assemblage, however, would have been damaged, diminishing the taxonomic value of the starch grains (Henry et al. 2009). We must also consider the effect of multiple processing episodes on the starch assemblage, particularly when a single vessel is used to process different foodstuffs. If starch grains were damaged as a result of e.g. boiling, further boiling is likely to completely destroy the grains. Therefore, the starch assemblage from a vessel used for multiple processing episodes should reflect the plants processed in the last (or few last) episode(s), but this should be specifically addressed through experimentation in order to test this hypothesis. Finally, vessels used to serve and consume food would contain a certain amount of microbotanical remains transferred directly from the food, but since no mechanical disturbance happens on the vessel itself, only a few microbotanical remains would be deposited on the pores of serving/consumption vessels. To conclude, we hypothesise that vessels directly used to process (cook, grind, grate, etc.) plant foodstuffs should have distinctive microbotanical assemblages (i.e. comparatively rich and including damaged starch grains), but it may be challenging to distinguish between serving and storage vessels based merely on the presence/absence of microbotanical remains due to the lack of mechanical disturbance, which would result in similar depositional patterns.
Materials and Methods
As part of the intensive recording of material culture and systematic sampling of bioarchaeological proxies implemented within the Knossos-Gypsades project, potsherds from cooking, storage and presentation/serving vessels were carefully collected in the trench. Potsherds were later sub-selected by EH based on their morphology and contextual information and sampled in order to explore food preparation and consumption patterns. In this study, we analysed starch grains and phytoliths from a total of 28 archaeological samples: 24 samples from ceramic vessels, plus four sediment samples from the archaeological matrix in order to provide a benchmark to assess contamination from the burial environment (Table 2). Pottery types included 13 cooking vessels (tripod cooking pots), four storage vessels (pithoi) and seven presentation/serving vessels (four conical cups, an open decorated vessel, an egg cup and a bridge-spouted jar). The selection of pottery types included the most commonly found pottery types at Gypsades (tripod cooking pots, pithoi and conical cups), which are well-known in Minoan archaeology (Momigliano 2007), as well as three elaborated types thought to have been used for serving purposes. In order to check for modern starch contamination (see Crowther et al. 2014), we also analysed starch grains from laboratory consumables used to process and analyse starch samples: two tubes, a beaker and the glycerine used to mount starch slides.
Sampling, processing and analysis of microbotanical remains followed the protocols described in García-Granero et al. (2017). The recovery of microbotanical remains from potsherds took place in a controlled environment—a closed room with no airstream at the Stratigraphic Museum, Knossos, Crete. Residue recovery consisted of a two-step process in which the outer layer of sediment was first dry brushed from the inner surface of the vessel (dry sample), and then the inner layer of sediment was brushed with deionised water (wet sample) (Hart 2011). Processing and analysis of the wet samples took place at the BioGeoPal Laboratory, IMF-CSIC, Barcelona, Spain. Gloves were not used during the sampling of potsherds or the extraction and analysis of starch grains to prevent starch contamination (Crowther et al. 2014). Microbotanical remains were analysed at ×200 and ×630 magnifications using a Leica DM 2500 microscope equipped with plain and cross-polarised transmitted light and a Leica DF 470 camera for microphotography. Starch slides were wholly scanned, and all grains were photographed for posterior analysis. Phytolith slides were scanned until 250 phytoliths were identified or until 10% of the slide was covered; only phytoliths with diagnostic taxonomic value were photographed. When necessary, slides were revisited to further photograph selected phytoliths for morphometric analysis.
Measurements of all starch grains and selected phytoliths were acquired with the software ImageJ 10.2 and compared with morphometric data from modern plant reference material hosted at the Archaeobotany Laboratory, School of Archaeology, University of Oxford (UK). Morphometric data were expressed as boxplots for visualisation. The statistical significance of the differences between archaeological and modern samples was tested using a multivariate analysis of variance (MANOVA). Each individual variable was further analysed using a one-way analysis of variance (ANOVA) to explore the differences between groups in detail through a series of Tukey honestly significant difference (HSD) multiple comparison tests. All the statistical analyses were computed using the software R 3.4.2 on RStudio 1.1.383 using the package ‘ggplot2’ for plotting the boxplots and the package ‘dplyr’ for computing the MANOVAs and the one-way ANOVAs. The raw outcome data of the statistical analyses can be found in the Online Supplementary Material (ESM1 and ESM2).
Results
Starch Grains
Starch grains were recovered from all types of sample but were generally scarce (Table 3). Most grains were simple (found in isolation from each other), with the exception of two compounds (starch aggregates) found in sample 30055-2. Four different morphotypes were identified (Fig. 2):
-
a)
37 medium/large grains (range 10.079–32.597 μm, average 20.407 μm, SD 5.148 μm) characteristic of the Triticeae tribe (Pooideae, Poaceae) labelled ‘Triticeae Type A’. Grains presented a smooth surface and had central hila, and were rounded or ovate in two-dimensional shape and discoidal in 3D. Most grains presented some degree of damage (Figure S1), including cracks along the perimeter, a central depression and loss of birefringence, all of which have been associated with cooking (Henry et al. 2009). Some grains presented lamellae, although it is not possible to ascertain whether these are naturally occurring or the result of cooking (see Henry et al. 2009: Fig. 2).
-
b)
11 very small/small grains (range 3.308–7.581 μm, average 5.717 μm, SD 1.278 μm) also characteristic of the Triticeae tribe labelled ‘Triticeae Type B’. Grains presented a smooth surface and had central hila, and were rounded in two-dimensional shape and spherical in 3D.
-
c)
9 small/medium grains (range 8.738–18.639 μm, average 14.199 μm, SD 3.184 μm) resembling taxa within the Andropogoneae tribe (Panicoideae, Poaceae) labelled ‘Andropogoneae-type’. Grains presented a wrinkle surface and had central hila, and were polygonal in two-dimensional shape and polyhedral in 3D.
-
d)
6 small/medium grains (range 8.696–17.106 μm, average 12.305 μm, SD 3.359 μm) from an unknown origin labelled ‘UNID Type’. Grains presented a smooth surface and had central hila, and were ovate in two-dimensional shape and ovoid in 3D.
Triticeae starch grains were recovered from both archaeological and laboratory control samples, whereas Andropogoneae-type and UNID-type grains were recovered only from archaeological samples. Very few starch grains (≤ 4 grains) were recovered from most samples, with the exception of sample 30055-2 (a cooking vessel), which was relatively rich in type A Triticeae starch grains.
The measurement of the major axis (length) of the type A Triticeae starch grains from sample 30055-2 was compared statistically to the length of starch grains from modern domestic cereals (Table 4). Modern taxa were selected on the basis of previous archaeobotanical research in Crete, which has identified free-threshing wheat (Triticum turgidum ssp. durum/T. aestivum), emmer (T. turgidum ssp. dicoccum), einkorn (T. monococcum ssp. monococcum) hulled barley (Hordeum vulgare) and naked barley (H. vulgare var. nudum) as the main cereal crops during the Neolithic and the Bronze Age (Livarda and Kotzamani 2013 and references therein). Significant statistical differences were only found between the length of the type A Triticeae starch grains from sample 30055-2 and the length of type A starch grains from modern einkorn; all other modern specimens were statistically indistinguishable from the archaeological granules (Fig. 3). A one-way ANOVA showed that the modern taxa most resembling the starch assemblage from Gypsades are naked barley and durum wheat (Fig. 3).
Phytoliths
Phytoliths were relatively abundant in cooking vessels and sediment samples, but virtually absent from other types of vessels (Table 5, Table S2). Samples with less than 100 identified phytoliths were considered sterile and are not further discussed—sterile samples included all storage and serving vessels and five cooking vessels. Most encountered phytoliths are attributable to grasses (Poaceae), in particular from the Pooideae subfamily, which includes domestic cereals such as wheat and barley, among other taxa. Pooideae phytoliths included rondels (Fig. 4a) and several types of trapeziforms (Fig. 4b). A more precise taxonomic identification of the grass phytoliths is not possible due to the absence of silica skeletons (compound phytoliths) from grass inflorescences, which can be identified to species level when analysed morphometrically (see e.g. Ball et al. 2009), but the morphology of certain grass inflorescence long cells is reminiscent of Triticum spp. (Fig. 4c)—with dendritic protuberances and a width comparable to inflorescence phytoliths from modern T. aestivum and T. turgidum ssp. dicoccum (Ball et al. 1999: Table 5). Grass phytoliths from the Chloroideae (saddles) and the Panicoideae (e.g. bilobates) subfamilies, which in this region are attributable to weeds or wild grasses, were very scarce or completely absent, as were phytoliths from dicotyledons (e.g. globular irregulars). The phytolith assemblage further included globular echinate phytoliths characteristic of palms (Arecaceae) (Fig. 4d), which formed a small part of the assemblage but were present in most samples.

Phytoliths recovered from samples analysed in this study: a, b Pooideae short cells from sample 30059-3, c grass (Poaceae) inflorescence long cell (Triticum-type) from sample 30057-7 and d palm (Arecaceae) phytoliths from sample 30055-20. Scale bars 20 μm
The palm phytoliths from cooking vessels and sediment samples were morphometrically compared with modern globular phytoliths from Cretan date palm (Phoenix theophrasti), which is native to the island, and date palm (Phoenix dactylifera), which is not native to Crete but is widely distributed along the eastern Mediterranean coast (Barrow 1998). Both the major (length) and minor (breadth) axes were measured, as well of the ratio between them (globularity). No statistically significant differences were found between the length and breadth of palm phytoliths from cooking vessels and sediment samples from Gypsades, or between phytoliths from these two groups and phytoliths from date palm leaves (Table 6; Fig. 5).

Length and breadth measurement of the Arecaceae phytoliths from sediment samples and vessels from Gypsades compared with fruit and leaf phytoliths from modern Phoenix dactylifera and Phoenix theophrasti: a boxplot and b results of the one-way ANOVA of each variable (statistically significant differences are highlighted in bold)
Discussion
Taphonomic Processes Affecting the Microbotanical Assemblage
Before discussing the interpretation of the results of this study in terms of food preparation and consumption, it is essential to assess critically the depositional and post-depositional processes that may have affected the microbotanical assemblage from Gypsades.
As discussed above, the different uses of each category of pottery vessel (storage, cooking and serving) may affect the deposition of microbotanical remains. At Gypsades, plants would not have been exposed to mechanical damage in storage and serving vessels, which may explain the virtual absence of microbotanical remains in these samples. On the other hand, the repeated disturbance caused by cooking methods such as boiling could also account for the comparatively richer microbotanical assemblage recovered from cooking vessels—among all the samples from potsherds, cooking vessels contained most starch grains (91%) and were the only ones with 100 or more identified phytoliths.
The post-depositional processes potentially affecting the microbotanical assemblage from Gypsades can be divided into two stages: those occurring between the time of deposition and the time of retrieval from the archaeological matrix (stage 1), and those occurring between the time of excavation and the analysis (stage 2). Phytolith contamination is more likely to occur during stage 1 as a result of a transfer from the archaeological matrix (burial environment) to the artefacts, whereas starch contamination is more likely to occur during stage 2 as a result of a transfer from sources of modern starch to the archaeological samples. Since all of the pottery vessels analysed in this study come from the same archaeological context, it can be assumed that any potential stage 1 process had a similar effect on the preservation of microbotanical remains in all samples from potsherds. Therefore, differences in the microbotanical assemblage recovered from pottery vessels are unlikely to be due to stage 1 post-depositional processes. Samples from the archaeological matrix (sediment samples), however, would have been subjected to other post-depositional pressures. In particular, without the protective environment provided by the pores of pottery vessels, starch grains would have been exposed to enzymatic activity and other environmental destructive factors (Haslam 2004), which probably account for their absence from most sediment samples. Regarding phytoliths, the similarities between the assemblages recovered from pottery vessels and those recovered from sediment samples indicate that (some of) the phytoliths recovered from the artefacts may not reflect their actual use but the plants generally present at the site. However, we must take into account that all the samples come from a very particular context—a midden—where pottery and food remains were deposited. It is therefore possible that the phytolith assemblage from the sediment samples reflects the same plants that were stored, cooked and/or served in pottery vessels. Further microbotanical analyses from Gypsades (currently ongoing) will help to disentangle the potential transfer of phytoliths from the archaeological matrix to artefacts.
Between their recovery from the archaeological matrix and their analysis (stage 2), samples could have been exposed to modern starch contamination from a series of environmental and laboratory sources. Starch contamination may occur at any point between recovery, processing and analysis (Crowther et al. 2014; Mercader et al. 2017), and it is not always possible to detect it once it has occurred. Two plant taxa are most likely to appear in archaeological assemblages as a result of modern starch contamination: maize (Zea mays) and wheat (Triticum spp.) (Crowther et al. 2014). The starch assemblage from Gypsades includes grains resembling both maize—the size and shape of the Andropogoneae-type (a tribe of grasses which includes maize, among several other plants) starch grains fall within the range of modern maize starch grains (Piperno et al. 2009: Table S2)—and wheat, and therefore we must consider whether these taxa entered the starch assemblage as consumed foodstuffs or modern contaminants.
Gloves are the most common source of modern maize starch grains, although these can also be found in other laboratory consumables (Crowther et al. 2014: Table 3). For this reason, the use of gloves was avoided during the whole study of starch samples from Gypsades, from field recovery to microscopic analysis. Moreover, no maize-like starch grains were recovered from laboratory consumables. The possibility of modern maize starch contamination is by no means excluded, but other scenarios should also be considered.
Despite not being a common crop, there is some evidence of millet crops (c.f. Panicum sp., Panicoideae, Poaceae) in Bronze Age Crete (Livarda and Kotzamani 2013: 11), although this evidence is currently being reviewed and it is likely that they represent intrusive material (Alexandra Livarda pers. comm.). In any case, Panicum spp. produce starch morphologically comparable with the Andropogoneae-type grains recovered from Gypsades but somewhat smaller (Yang et al. 2012b). Starch grains morphologically and morphometrically comparable to the Andropogoneae-type grains from Gypsades have been found in the stems of several cereal species including wheat (Yang et al. 2014b), although the similarity of these grains with maize starch raises questions regarding potential contamination.
The Andropogoneae-type grains from Gypsades could also belong to a completely different plant taxon (see the discussion in García-Granero et al. 2018: ESM1). Indeed, similar starch grains have been recovered from other prehistoric and early historic contexts in the Aegean (García-Granero et al. 2018; García-Granero unpublished research), which may be indicative of the widespread consumption of a plant taxon not yet identified. In any case, the taxonomical adscription of the Andropogoneae-type starch grains from Gypsades must be considered with caution, and until more evidence (from Gypsades or elsewhere) is available contamination from modern sources cannot be ruled out.
The presence of a small amount (≤ 2 grains) of wheat-like (Triticeae type A) starch grains in the laboratory control samples analysed in this study, on the other hand, strongly suggests that at least part of the Triticeae starch grains from Gypsades were incorporated in the archaeological assemblage as a result of contamination during laboratory processing. For this reason, all the samples in which only a small amount (≤ 2 grains) of Triticeae type A starch grains were recovered are considered potentially contaminated and Triticeae type A starch grains from these samples are not further discussed. Consequently, the discussion of the results of the Triticeae Type A starch grains focuses exclusively on the assemblage from sample 30,055–2 (a cooking vessel).
A Holistic Microbotanical Approach to Foodways at Late Minoan Gypsades
Bearing in mind the depositional and post-depositional processes that may have affected the starch and phytolith assemblages from Gypsades, it is now possible to evaluate how the microbotanical evidence informs about Late Minoan foodways (Table 7). Despite the possible contamination of part of the starch assemblage, the predominance of pooid phytoliths in all samples and the starch evidence from vessel 30055-2 suggest that cereals were an important part of the diet of the inhabitants of LM IA Gypsades. Phytoliths could not be attributed to a lower taxonomic category (genus/species) due to the lack of silica skeletons, but the morphometric data from Triticeae type A starch grains suggests that the remains from sample 30055-2 belong to either naked barley or durum wheat, although it is also statistically possible that the archaeological starch grains belong to bread wheat, emmer or hulled barley, all of which have been identified in the macrobotanical assemblage from Gypsades (Gypsades project unpublished research).
The phytolith data further suggest that the cereals consumed by the Late Minoan inhabitants of Gypsades were thoroughly cleaned before being stored and cooked. Phytoliths representing weeds and wild taxa—in this case, short cells from chloridoid and panicoid grasses and morphotypes from dicots—were virtually absent, suggesting the agricultural produce had undergone sieving and winnowing (Jones 1990). Moreover, the scarcity of grass inflorescence phytoliths in all samples, including storage vessels, suggests that cereals were de-husked prior to storage and consumption.
The presence of cereals in the diet of the inhabitants of Gypsades is not surprising considering the macrobotanical data available from Crete and elsewhere in the Bronze Age Aegean, but the absence of pulses is somewhat unexpected. Pulse taxa are not great phytolith producers, but the seeds are rich in starch grains, which can be easily identified to family (Fabaceae) or even subfamily (Faboideae) level (see examples in Henry et al. 2009: Figs. 2 and 3), but no Faboideae starch grains were recovered from the Gypsades samples. Charred seeds from a variety of pulses have been commonly recovered from prehistoric archaeological contexts in Crete (Livarda and Kotzamani 2013), including deposits (separate and later in date than the ‘midden’ analysed here) excavated during the 2014–2015 field seasons at Gypsades (Gypsades project unpublished research). Moreover, several pulses stored as separate crops—including clean winged vetchling (Lathyrus ochrus) and Celtic bean (Vicia faba)—alongside cereals were recovered from the Late Minoan II (c. 1450–1400 BC) burned destruction layer of Storeroom P in the Unexplored Mansion, Knossos (Jones 1984, 1992). The evidence for the consumption of pulses in Minoan Crete seems conclusive, but their absence from the Gypsades microbotanical samples raises questions regarding the preparation and consumption of pulses in Minoan Gypsades. It is possible that cereals and pulses followed different culinary pathways, and that pithoi and tripod cooking vessels were used for the former but not the latter. We must also consider, however, that the samples analysed in this study come from a single archaeological context, which does not represent the temporal and spatial variability of the culinary practices at Minoan Gypsades. It is thus possible that the absence of pulses in the samples from Gypsades responds to the specificities of the archaeological context.
Beyond cereal crops, the microbotanical assemblage from Gypsades includes a few starch grains that could not be assigned to a specific taxon (UNID type). These starch grains do not belong to agricultural staples (cereals and pulses) and only appear in cooking vessels, suggesting that they could belong to plants consumed as complements to agricultural staples or as flavour enhancers. A number of herbs and spices such as mint and coriander are mentioned in Linear B texts (Isaakidou 2007 and references therein). Moreover, macrobotanical remains from several non-agricultural plants have been recovered from Bronze Age contexts in Crete, including fruits, nuts, condiments and leafy greens, several of which have been traditionally used for human consumption (Livarda and Kotzamani 2013: Table 3).
Date Palm Basketry: an Exotic Import
The presence of date palm leaf phytoliths in Gypsades warrants a separate discussion. Date palms (P. dactylifera) are not native to Crete (Barrow 1998), and available archaeobotanical evidence suggests that they were not cultivated in the island in prehistoric times. Charred date palm kernels have not been recovered from Cretan Neolithic or Bronze Age contexts (Livarda and Kotzamani 2013), and Phoenix sp. charcoal—which could belong to either P. dactylifera or P. theophrasti—is extremely rare in Minoan Crete, and has not been found in the analysed samples from Gypsades (Gypsades project unpublished research). Palm leaves produce millions of phytoliths per gram of dried plant matter (Albert et al. 2009). However, very few date palm leaf phytoliths were recovered in the samples from Gypsades, suggesting that these phytoliths were indirectly incorporated into the midden in Trench 3—i.e. not after the in situ decay of a date palm leaf. More likely, phytoliths were transferred to pottery vessels and sediment samples from a finished product made of date palm leaves, such as a basket, which would have been in direct contact with foodstuffs that were later processed in pottery vessels. Unlike pollen grains, phytoliths are not transported over long distances due to their relative ‘heaviness’ (Madella and Lancelotti 2012: 78), and therefore, phytolith contamination from modern sources is very unlikely because archaeological samples were not in contact with or in close proximity to palm leaves at any point. The evidence thus suggests that the inhabitants of Gypsades used imported baskets made of date palm leaves, perhaps as (crop) storage containers.
Cretan date palms (P. theophrasti) are widely distributed along the Cretan coast (Barrow 1998), and were potentially even more widespread during Minoan times. The leaves of Cretan date palms are still weaved by traditional Cretan populations for crafting (García-Granero et al. 2020), a practice that could also have taken place in prehistoric times. However, the phytolith evidence presented here suggests that the inhabitants of Gypsades imported baskets made of date palm leaves but did not use a locally and widely available resource (Cretan date palm leaves) to produce their own.
Imported goods made of date palms were also identified through phytolith analyses at Mycenaean Tiryns, in the Peloponnese (Vetters et al. 2016), but not from the Neolithic deposits under the Central Court of the Knossos palace (Madella 2013), suggesting that goods made of date palms were imported during the (Late) Bronze Age but not in earlier times. We must take into account, however, that Madella’s (2013) study considered a limited number of samples from a very small excavation trench. Moreover, no phytolith studies have been conducted thus far on Early and Middle Minoan deposits. Therefore, more evidence is needed to time the arrival of goods made of date palm leaves into Crete.
Conclusions
This study illustrates how microbobotanical remains can provide direct evidence of past plant preparation and consumption. The microbotanical analyses of Late Minoan pottery from Gypsades document not only the consumption of de-husked cereals in Late Minoan Gypsades but also, interestingly, the absence of pulses from pottery vessels from the midden in Trench 3. The results also suggest that non-agricultural plants were incorporated into Minoan foodways, a topic that will be further explored through extended starch and phytolith reference collections.
This study aimed at reconstructing the whole domestic lifespan of food crops, from storeroom to midden. The study of microbotanical remains from pottery vessels provided evidence of food storage (cereals stored de-husked in pithoi and possible use of palm leaf baskets as storage containers) and culinary practices (cooking of clean cereal product and non-agricultural plants). However, microbotanical remains were virtually absent from serving vessels. As discussed above, this may be due to depositional factors or simply the fact that some of the vessels analysed in this study were used for serving/consuming liquids or animal products, and not plants. In order to identify the full scope of foodstuffs consumed by the inhabitants of Gypsades, future research will integrate the analysis of microbotanical remains with the chemical analysis of the residue trapped in pottery vessels.
Finally, it is worth highlighting the importance of (a) putting in place preventive mechanisms for minimising modern starch contamination and (b) critically assessing the integrity of the ancient starch assemblage. In this study, we decided to be particularly cautious and treat the starch assemblage from most archaeological samples as potentially contaminated to ensure that our interpretation took potential modern starch contamination into account. We believe that a critical assessment of archaeological starch assemblages should be a fundamental part of future studies considering ancient starch.
References
Albert, R. M., Bamford, M. K., & Cabanes, D. (2009). Palaeoecological significance of palms at Olduvai Gorge, Tanzania, based on phytolith remains. Quaternary International, 193(1-2), 41–48.
Alonso, N., Junyent, E., Lafuente, A., & López, J. B. (2008). Plant remains, storage and crop processing inside the Iron Age fort of Els Vilars d’Arbeca (Catalonia, Spain). Vegetation History and Archaeobotany, 17(S1), 149–158.
Ball, T. B., Gardner, J. S., & Anderson, N. (1999). Identifying inflorescence phytoliths from selected species of wheat ( Triticum monococcum, T. dicoccon, T. dicoccoides, and T. aestivum) and barley (Hordeum vulgare and H. spontaneum) (Gramineae). American Journal of Botany, 86(11), 1615–1623.
Ball, T. B., Ehlers, R., & Standing, M. D. (2009). Review of typologic and morphometric analysis of phytoliths produced by wheat and barley. Breeding Science, 59(5), 505–512.
Ball, T., Chandler-Ezell, K., Dickau, R., Duncan, N., Hart, T. C., Iriarte, J., Lentfer, C., Logan, A., Lu, H., Madella, M., Pearsall, D. M., Piperno, D. R., Rosen, A. M., Vrydaghs, L., Weisskopf, A., & Zhang, J. (2016). Phytoliths as a tool for investigations of agricultural origins and dispersals around the world. Journal of Archaeological Science, 68, 32–45.
Barrow, S. C. (1998). A monograph of Phoenix L. (Palmae: Coryphoideae). Kew Bulletin, 53(3), 513–575.
Boyd, M., Surette, C., & Nicholson, B. A. (2006). Archaeobotanical evidence of prehistoric maize (Zea mays) consumption at the northern edge of the Great Plains. Journal of Archaeological Science, 33(8), 1129–1140.
Boyd, M., Varney, T., Surette, C., & Surette, J. (2008). Reassessing the northern limit of maize consumption in North America: Stable isotope, plant microfossil, and trace element content of carbonized food residue. Journal of Archaeological Science, 35(9), 2545–2556.
Chandler-Ezell, K., Pearsall, D. M., & Zeidler, J. A. (2006). Root and tuber phytoliths and starch grains document manioc (Manihot esculenta), arrowroot (Maranta arundinacea), and llerén (Calathea sp.) at the Real Alto site, Ecuador. Economic Botany, 60(2), 103–120.
Crowther, A. (2005). Starch residues on undecorated Lapita pottery from Anir, New Ireland. Archaeology in Oceania, 40(2), 62–66.
Crowther, A., Haslam, M., Oakden, N., Walde, D., & Mercader, J. (2014). Documenting contamination in ancient starch laboratories. Journal of Archaeological Science, 49, 90–104.
Dickau, R., Bruno, M. C., Iriarte, J., Prümers, H., Betancourt, C. J., Holst, I., & Mayle, F. E. (2012). Diversity of cultivars and other plant resources used at habitation sites in the Llanos de Mojos, Beni, Bolivia: Evidence from macrobotanical remains, starch grains, and phytoliths. Journal of Archaeological Science, 39(2), 357–370.
García-Granero, J. J., Lancelotti, C., & Madella, M. (2017). A methodological approach to the study of microbotanical remains from grinding stones: A case study in northern Gujarat (India). Vegetation History and Archaeobotany, 26(1), 43–57.
García-Granero, J. J., Urem-Kotsou, D., Bogaard, A., & Kotsos, S. (2018). Cooking plant foods in the northern Aegean: Microbotanical evidence from Neolithic Stavroupoli (Thessaloniki, Greece). Quaternary International, 496, 140–151.
García-Granero, J. J., Sarpaki, A., Skoula, M., Cárdenas, M., Madella, M., & Bogaard, A. (2020). A long-term assessment of the use of Phoenix theophrasti Greuter (Cretan date palm): The ethnobotany and archaeobotany of a neglected palm. Journal of Ethnobiology, 40(1), 101–114.
Hart, T. C. (2011). Evaluating the usefulness of phytoliths and starch grains found on survey artifacts. Journal of Archaeological Science, 38(12), 3244–3253.
Hart, J. P., Thompson, R. G., & Brumbach, H. J. (2003). Phytolith evidence for early maize (Zea mays) in the northern Finger Lakes region of New York. American Antiquity, 68(4), 619–640.
Hart, J. P., Brumbach, H. J., & Lusteck, R. (2007). Extending the phytolith evidence for early maize (Zea mays ssp. mays) and squash (Cucurbita sp.) in Central New York. American Antiquity, 72(3), 563–583.
Haslam, M. (2004). The decomposition of starch grains in soils: Implications for archaeological residue analyses. Journal of Archaeological Science, 31(12), 1715–1734.
Henry, A. G., Hudson, H. F., & Piperno, D. R. (2009). Changes in starch grain morphologies from cooking. Journal of Archaeological Science, 36(3), 915–922.
Isaakidou, V. (2007). Cooking in the labyrinth: Exploring ‘cuisine’ at Bronze Age Knossos. In C. Mee & J. Renard (Eds.), Cooking up the past (pp. 5–24). Oxford: Oxbow Books.
Jones, G. (1984). The LM II plant remains. In M. R. Popham (Ed.), The Minoan unexplored mansion at Knossos: Text (pp. 303–306). London: British School at Athens.
Jones, G. (1990). The application of present-day cereal processing studies to charred archaeobotanical remains. Circaea, 6, 91–96.
Jones, G. (1992). Ancient and modern cultivation of Lathyrus ochrus (L.) DC in the Greek islands. Annual of the British School at Athens, 87, 211–217.
Livarda, A., & Kotzamani, G. (2013). The archaeobotany of Neolithic and Bronze Age Crete: Synthesis and prospects. Annual of the British School at Athens, 108, 1–29.
Madella, M. (2013). Plant economy and the use of space: Evidence from the opal phytolith. In N. Efstratiou, A. Karetsou, & M. Ntinou (Eds.), The Neolithic settlement of Knossos in Crete: New evidence for the early occupation of Crete and the Aegean Islands (pp. 119–132). Philadelphia: INSTAP Academic Press.
Madella, M., & Lancelotti, C. (2012). Taphonomy and phytoliths: A user manual. Quaternary International, 275, 76–83.
Mercader, J., Abtosway, M., Baquedano, E., Bird, R. W., Díez-Martín, F., Domínguez-Rodrigo, M., Favreau, J., Itambu, M., Lee, P., Mabulla, A., Patalano, R., Pérez-González, A., Santonja, M., Tucker, L., & Walde, D. (2017). Starch contamination landscapes in field archaeology: Olduvai Gorge, Tanzania. Boreas, 46(4), 918–934.
Momigliano, N. (Ed.). (2007). Knossos pottery handbook: Neolithic and Bronze Age (Minoan). London: British School at Athens.
Morgan, C. (2015). The work of the British School at Athens, 2014–2015. Archaeological Reports, 61, 34–48.
Musaubach, M. G., & Berón, M. A. (2017). Food residues as indicators of processed plants in hunter-gatherers’ pottery from La Pampa (Argentina). Vegetation History and Archaeobotany, 26(1), 111–123.
Perry, L., Dickau, R., Zarrillo, S., Holst, I., Pearsall, D. M., Piperno, D. R., Berman, M. J., Cooke, R. G., Rademaker, K., Ranere, A. J., Raymond, J. S., Sandweiss, D. H., Scaramelli, F., Tarble, K., & Zeidler, J. A. (2007). Starch fossils and the domestication and dispersal of chili peppers (Capsicum spp. L.) in the Americas. Science, 315(5814), 986–988.
Pető, Á., Gyulai, F., Pópity, D., & Kenéz, Á. (2013). Macro- and micro-archaeobotanical study of a vessel content from a Late Neolithic structured deposition from southeastern Hungary. Journal of Archaeological Science, 40(1), 58–71.
Piperno, D. R. (2006). Phytoliths: A comprehensive guide for archaeologists and paleoecologists. Rowman Altamira Press.
Piperno, D. R., Ranere, A. J., Holst, I., Iriarte, J., & Dickau, R. (2009). Starch grain and phytolith evidence for early ninth millennium BP maize from the Central Balsas River Valley, Mexico. Proceedings of the National Academy of Sciences, 106, 5019–5024.
Portillo, M., Llergo, Y., Ferrer, A., & Albert, R. M. (2017). Tracing microfossil residues of cereal processing in the archaeobotanical record: An experimental approach. Vegetation History and Archaeobotany, 26(1), 59–74.
Samuel, D. (1996). Investigation of ancient Egyptian baking and brewing methods by correlative microscopy. Science, 273(5274), 488–490.
Saul, H., Wilson, J., Heron, C. P., Glykou, A., Hartz, S., & Craig, O. E. (2012). A systematic approach to the recovery and identification of starches from carbonised deposits on ceramic vessels. Journal of Archaeological Science, 39(12), 3483–3492.
Saul, H., Madella, M., Fischer, A., Glykou, A., Hartz, S., & Craig, O. E. (2013). Phytoliths in pottery reveal the use of spice in European prehistoric cuisine. PLoS ONE, 8(8), e70583.
Staller, J. E., & Thompson, R. G. (2002). A multidisciplinary approach to understanding the initial introduction of maize into coastal Ecuador. Journal of Archaeological Science, 29(1), 33–50.
Thompson, R. G., Hart, J. P., Brumbach, H. J., & Lusteck, R. (2004). Phytolith evidence for twentieth-century BP maize in northern Iroquoia. Northeast Anthropology, 68, 25–40.
Torrence, R., & Barton, H. (Eds.). (2006). Ancient starch research. Walnut Creek: Left Coast Press.
Twiss, K. (2012). The archaeology of food and social diversity. Journal of Archaeological Research, 20(4), 357–395.
Valamoti, S. M., Samuel, D., Bayram, M., & Marinova, E. (2008). Prehistoric cereal foods from Greece and Bulgaria: Investigation of starch microstructure in experimental and archaeological charred remains. Vegetation History and Archaeobotany, 17(S1), 265–276.
Vetters, M., Brysbaert, A., Ntinou, M., Tsartsidou, G., & Margaritis, E. (2016). People and plants: Piecing together archaeological and archaeobotanical data to reconstruct plant use and craft activities in Mycenaean Tiryns. Opuscula, 9, 93–132.
Wang, J., Liu, L., Georgescu, A., Le, V. V., Ota, M. H., Tang, S., & Vanderbilt, M. (2017). Identifying ancient beer brewing through starch analysis: A methodology. Journal of Archaeological Science: Reports, 15, 150–160.
Yang, X., & Jiang, L. (2010). Starch grain analysis reveals ancient diet at Kuahuqiao site, Zhejiang Province. Chinese Science Bulletin, 55(12), 1150–1156.
Yang, X., Wan, Z., Perry, L., et al. (2012a). Early millet use in northern China. Proceedings of the National Academy of Sciences, 109, 3726–3730.
Yang, X., Zhang, J., Perry, L., Ma, Z., Wan, Z., Li, M., Diao, X., & Lu, H. (2012b). From the modern to the archaeological: Starch grains from millets and their wild relatives in China. Journal of Archaeological Science, 39(2), 247–254.
Yang, X., Ma, Z., Wang, T., Perry, L., Li, Q., Huan, X., & Yu, J. (2014a). Starch grain evidence reveals early pottery function cooking plant foods in North China. Chinese Science Bulletin, 59(32), 4352–4358.
Yang, X., Ma, Z., Li, Q., Perry, L., Huan, X., Wan, Z., Li, M., & Zheng, J. (2014b). Experiments with lithic tools: Understanding starch residues from crop harvesting. Archaeometry, 56(5), 828–840.
Zarrillo, S., Pearsall, D. M., Raymond, J. S., Tisdale, M. A., & Quon, D. J. (2008). Directly dated starch residues document early formative maize (Zea mays L.) in tropical Ecuador. Proceedings of the National Academy of Sciences, 105, 5006–5011.
Acknowledgments
We are grateful to the Greek Ministry of Culture and Sports for granting us permission to excavate (ΥΠΠΟ/ΓΔΑΠΚ/ΔΙΠΚΑ/ΤΕΞΕΙΟΔΘ/Α2/Φ1/103929/595) and to export the archaeological samples analysed in this study (ΥΠΠΟ/ΓΔΑΠΚ/ΔΣΑNM/ΤΕΕ/Φ77/235442/141123/3619), and to the staff of the Ephorate of Herakleion for generously facilitating the project. Special thanks are due to the following Ephorate of Herakleion staff: Maria Rousaki (archaeologist responsible for the Knossos valley), Manolis Chatzimanolis (conservator) and Stavros Amanakis (antiquities guard). Day-to-day fieldwork operations at Gypsades were overseen on behalf of the Ephorate of Herakleion by archaeologist Marina Velegraki. We are extremely thankful to all the members of the Knossos-Gypsades project, especially Vasiliki Choleva for her help with sampling the potsherds, Dr. Valasia Isaakidou for her inestimable help during the fieldwork and sampling process and for her helpful comments on a previous version of this manuscript, and Charlotte Diffey and Maria Ntinou for sharing unpublished archaeobotanical and anthracological data. We are also grateful to Gerald Cadogan, Dr. Kostis Christakis, Prof. Paul Halstead, Dr. Matthew Haysom (former BSA Knossos Curator), and Prof. Cathy Morgan for their help at various stages of the project, and to Prof. Marco Madella for discussion during the initial stages of the microbotanical study.
Funding
The project was made possible with funds provided by INSTAP (EH, GA, AB); Semple Fund, Department of Classics, University of Cincinnati (EH); ERC Grant No. 312785 (AB); Fell Fund, University of Oxford (AB); the British Academy Grant No. SG152915 (AB); and the BSA. While working on this manuscript, JJGG was funded by a Marie Skłodowska-Curie Individual Fellowship (MSCA 2015, European Commission Grant No. 704867) and a Juan de la Cierva Incorporación 2018 Fellowship (Spanish Ministry of Science, Innovation and Universities), and ET is currently a F.S.R. Research Fellow funded by the Universitè Catholique de Louvain.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflicts of interest.
Ethical Approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
Outcome of the statistical analysis of modern and archaeological starch grains. (TXT 2 kb)
ESM 2
Outcome of the statistical analysis of modern and archaeological phytoliths. (TXT 3 kb)
Table S1
Morphometric data of modern and archaeological starch grains (summarised in Table 4). (XLSX 13 kb)
Table S2
Phytoliths recovered from pottery vessels and sediment samples, morphotype by morphotype. (XLSX 39 kb)
Table S3
Morphometric data of modern and archaeological phytoliths (summarised in Table 6). (XLSX 30 kb)
Figure S1
Examples of damaged Triticeae Type A starch grains recovered from sample 30055-2 (cooking vessel) from Gypsades: a) swollen grain that has almost completely lost birefringence and other diagnostic features, b) grain with a central depression that has partially lost birefringence and c) grain showing small pits on the surface that has almost completely lost birefringence. Scale bars 20 μm. (PNG 4223 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
García-Granero, J.J., Hatzaki, E., Tsafou, E. et al. From Storage to Disposal: a Holistic Microbotanical Approach to Domestic Plant Preparation and Consumption Activities in Late Minoan Gypsades, Crete. J Archaeol Method Theory 28, 307–331 (2021). https://doi.org/10.1007/s10816-020-09456-9
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10816-020-09456-9