
Abstract
Osteoarthritis (OA), a chronic joint disease, is characterized by cartilage surface erosion, subchondral bone rebuilding, and formation of osteophytes. To date, the nosogenesis and underlying mechanisms of OA have not yet been elucidated. However, it is widely accepted that TNF-α is a crucial cytokine in the development of OA. Glycitin, a natural isoflavone extracted from legumes, affects physiological reactions and pathological responses. Recently, the anti-inflammatory effect of glycitin has been reported. However, the function of glycitin in cartilage degeneration in OA remains to be investigated. In the current study, primary murine chondrocytes were isolated and stimulated by TNF-α to evaluate the anti-inflammatory effects and protective function of glycitin in chondrocytes. In vivo, the ACLT mouse model, a frequently-used OA model, was used to further examine the therapeutic role of glycitin in cartilage degeneration and inflammation in OA. Consequently, glycitin functions were examined both in vivo and in vitro. Moreover, the underlying mechanism of action of glycitin was investigated and was found to involve the NF-κB signaling pathway. Collectively, this study suggests that glycitin can be potentially used for the treatment of joint degenerative diseases, including OA.
Similar content being viewed by others

INTRODUCTION
Osteoarthritis (OA), characterized by cartilage surface erosion, subchondral bone rebuilding, and formation of osteophytes, is estimated to become the fourth leading cause of disability by 2020 [1, 2]. It is a chronic joint disease that leads to oppressive pain, stiffness, and movement disorders. As previously reported, 12% of the people in Western countries suffer from OA and 25% (over 55 years old) are afflicted by joint pain [3, 4]. It is a widely held view that progressive degeneration of articular cartilage, synovitis, and formation of osteophytes are major pathologic lesions in OA [5].
Although the pathogenesis and associated underlying mechanism of OA are still unclear, it is well accepted that inflammatory reactions and inflammatory cytokines such as TNF-α, IL-1β, and IL-6 play a crucial role in the development of OA [6,7,8]. We have previously shown that inhibition of TNF-α may affect the release of IL-1β, IL-6, and other cytokines closely associated with OA, such as MMPs and ADAMTS [9,10,11]. The expression of matrix metalloproteinases (MMPs) and a disintegrin and metalloproteinase with thrombospondin motifs (ADAMTS) may inhibit the production of collagen and proteoglycan, which are vital components of cartilage. Moreover, it has been reported that inflammatory mediators such as prostaglandin E2 (PGE2) and nitric oxide (NO) are closely associated with the degeneration of cartilage, and are released in chondrocytes particularly when stimulated with TNF-α [12,13,14].
To date, there are still limited therapeutic approaches for OA, such as nonsteroidal anti-inflammatory drug (NSAIDs), physiotherapy, and surgery [15]. However, the therapeutic effect of NSAIDs and physiotherapy is limited and their side effects, including coronary heart disease, angina pectoris, cerebrovascular accident, and amyotrophy, should not be neglected [15, 16]. As has been recently reported, several therapeutic approaches for OA, including the use of functional extracts from various herbs, have shown a protective effect in OA [17, 18]. However, the mechanism of action of these therapeutics still requires further investigation. Thus, a new therapeutic treatment with a clear mechanism of action and less side effects is necessary.
Glycitin (4′-hydroxy-6-methoxyisoflavone-7-D-glucoside; CAS:40246–10-4), also named glycitin 7-O-β-glucoside, a natural isoflavone extracted from legumes, affects many physiological and pathological processes [19, 20]. It has been reported that glycitin plays a substantial role in cardiovascular diseases, obesity, and wound healing [21,22,23]. Moreover, antioxidant and anti-inflammatory effects have been reported in recently published studies [21, 24, 25]. Chen et al. have recently reported that glycitin has a therapeutic effect on LPS-induced acute lung injury in mice [20]. Furthermore, they confirmed the anti-inflammatory effect of glycitin in RAW264.7 cells stimulated by LPS [20]. In addition, the NF-κB signaling pathway was inhibited by glycitin in many pathological conditions [26, 27]. It is well accepted that the NF-κB signaling pathway plays a significant role in inflammation and cartilage degeneration [12, 27]. These findings prompted us to investigate whether glycitin plays a role in cartilage degeneration and OA progression and provide a potential therapeutic target for OA.
MATERIALS AND METHODS
Animals
Male BL6/C57 wild-type (WT) mice (10 weeks) were purchased from the Animal Center of Shandong University Laboratory (Jinan, Shandong, China). The experimental procedures performed on animals were approved by the Animal Research Ethics Committee at Shandong University.
Isolation and Culture of Primary Murine Chondrocytes
The primary chondrocyte cells were isolated from the knee joint cartilage and costicartilage of 10-week-old mice according to our previous work [28]. Briefly, the cartilage tissue from at least 10 mice was digested using 0.2% collagenase II (Sigma-Aldrich, USA) in DMEM (Gibco, USA) for 4 h at 37 °C. Next, the cartilage of mice was filtered through a filter mesh (74 μm), followed by centrifugation with 300×g for 5 min at 4 °C. Next, the cells were resuspended by DMEM containing 10% FBS (Gibco, USA), 100 U/ml penicillin (HyClone, UT, USA) and 100 μg/ml streptomycin (HyClone, UT, USA) into 1 × 106 cells/ml. Then 1 × 107 cells were planted in culture plastics (Thermo Scientific, Shanghai, China) in a 5% CO2 atmosphere at 37 °C for expansion. After three generations, the chondrocytes were planted in 24-well plates (2 × 105/well) or 96-well plates (5 × 104/well) or 12-well plates (5 × 105/well) for next test.
LAL Test and Cell Viability
Glycitin used in this study was purchased from MedchemExpress Company. First, glycitin was dissolved in bacteria-free PBS and filtered using a 0.22-μm filter membrane. The LAL test was used to examine endotoxin contamination of the glycitin solution (10 mg/ml in vivo and 25 μg/ml, 50 μg/ml, 75 μg/ml, 100 μg/ml, and 150 μg/ml in vitro) according to the protocol of ToxinSensor Chromogenic LAL Endotoxin Assay Kit (GenScript; limit of detection < 0.01 EU/ml). The results showed that the endotoxin contamination of glycitin was lower than 0.01 EU/ml. Furthermore, the effect of various concentrations of glycitin on cell viability was examined by using the CCK-8. In brief, chondrocytes were inoculated in 96-well plates (5 × 104/well) and cultured with different concentrations of glycitin for 24 h. Next, the CCK-8 reagent was added and incubation continued for 4 h at 37 °C. The Thermo Scientific Microplate Reader (Thermo Scientific, Shanghai, China) was used for the analysis and the data were analyzed by GraphPad Prism. All the samples were assayed in triplicate and the experiments were repeated at least three times.
Stimulation of Chondrocytes by TNF-α
Mice chondrocytes were used to evaluate the anti-inflammatory effect of glycitin in vitro. The chondrocytes were divided into 5 groups: 1. Control group (untreated), 2. TNF-α (10 ng/ml) without glycitin, 3. TNF-α (10 ng/ml) with glycitin (low dose 10 μg/ml), 4. TNF-α (10 ng/ml) with glycitin (high dose 40 μg/ml), 5. only glycitin (high dose 40 μg/ml). We seeded the chondrocytes into 12-well plates (5 × 105/well) with complete culture medium. When the cell density reaches 95%, complete culture medium was replaced by conditional culture medium (DMEM without FBS and any antibiotic) for 12 h. Then the cells in all groups were treated as described above for 24 h and next supernatant from the five groups were collected for NO analysis. Moreover, the protein and mRNA were extracted for further analyses.
OA Surgical Models
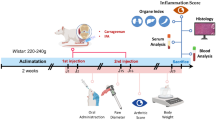
We established an Anterior Cruciate Ligament Transection (ACLT) mouse model to investigate the anti-inflammatory effect of glycitin in vivo. The mice were divided into three groups (n = 6): sham-operation group, ACLT group, and treatment group. In the sham operation group, the mice underwent the same operation without ligament transaction, followed by treatment with 0.2 ml PBS intraperitoneally once a day for 4 weeks. In the ACLT group, the anterior cruciate ligament (ACL) was transected, followed by intraperitoneal injection with 0.2 ml PBS once a day for 4 weeks. In the glycitin treatment group, the mice underwent the same operation but were also treated with glycitin (10 mg/ml) once a day for 4 weeks. The animals were sacrificed 4 weeks later and the knee joint tissue and serum of each mouse were collected for further evaluation.
Histological and Immunostaining Analysis
Samples collected from each group were fixed in 4% PFA and decalcified in 10% EDTA. Following embedding in paraffin, samples were cut in 5-μm sections and then stained with Safranin O/Fast Green. The proteoglycan content of cartilage, which reflects the histopathology, was graded by the OARSI scoring system for OA. For immunocytochemistry (IHC), after dewaxing and rehydration, sections were incubated in goat serum for at least 45 min to block nonspecific protein binding. Next, the sections were incubated with the antibody (p-IκB antibody (1:150 dilution, Santa Cruz Biotechnology, CA, USA.); MMP13 antibody (diluted 1:100; ProteinTech Group, Chicago, IL, USA); ADAMTS-5 antibody (diluted 1:100; ProteinTech Group, Chicago, IL, USA); collagen II antibody (diluted 1:100; ProteinTech Group, Chicago, IL, USA); iNOS antibody (diluted 1:150; ProteinTech Group, Chicago, IL, USA); COX2 antibody (diluted 1:150; ProteinTech Group, Chicago, IL, USA)) at 4 °C overnight. Finally, we used the enhanced polymer detection system kit (Boster Biological Technology, China) for detection, followed by counterstaining with 1% hematoxylin.
Western Blotting
Proteins extracted from the knee joint tissue and chondrocytes were analyzed by western blot. Membranes were incubated with antibodies against p-p65(diluted 1:3000 Affinity Biosciences, USA) P65 (diluted 1:3000; ProteinTech Group, Chicago, IL, USA), p-IκBα (diluted 1:1000; Santa Cruz Biotechnology, Santa Cruz, CA, USA), IκBα (diluted 1:1000; Santa Cruz Biotechnology, Santa Cruz, CA, USA) COX-2 (diluted 1:1000; Boster Biological Technology, Wuhan, China), iNOS (diluted 1:3000; ProteinTech Group, Chicago, IL, USA); Aggrecan (diluted 1:3000; ProteinTech Group, Chicago, IL, USA) at 4 °C overnight. We used an enhanced chemiluminescence system (Thermo Scientific, Shanghai, China) to visualize the bound antibodies.
Real-Time Polymerase Chain Reaction
Total RNA was extracted from tissues and chondrocytes from each group. As for chondrocytes, cells in each group were collected by cell scraper and their mRNA of was isolated by RNAeasy™ Animal RNA Isolation Kit with Spin Column (R0027, Beyotime Biotechnology, Beijing, China). As for tissues, knee cartilage of mice was cut by lancet and their mRNA of was isolated by RNAeasy™ Animal RNA Isolation Kit with Spin Column. Next, the RNA quality and integrity were tested by ultraviolet spectrophotometer (Thermo Scientific, Shanghai, China), and only qualified mRNA (A260/280 > 2; A260/230 > 2) was used for next test. Next, the RT-PCR kit (Vazyme Corporation, Nanjing, China) was used to reverse transcribe the mRNA into cDNA. Next, we performed real-time polymerase chain reaction (PCR) using the Thermal Cycler Dice Real-Time System with SYBR Green I dye (Stage 1: Reps:1, 95 °C for 5 min; Stage 2: Reps:40, 95 °C for 10 s, 60 °C for 30 s; Stage 3: Reps:1, 95 °C for 15 s, 60 °C for 60 s; 95 °C for 15 s). The primer sequences used for the analyses are presented in Table 1. The mean Ct values of each gene were normalized against GAPDH. The results were quantified using the ΔΔCt method. All experiments were repeated at least three times.
ELISA Assays both in vivo and in vitro
Serum from each mouse was harvested and analyzed for IL-1β, TNF-α, IL-6, and PGE2 using ELISA kits (Boster Biological Technology, Wuhan, China). Moreover, culture supernatants from chondrocytes in each group were collected for analysis using ELISA (IL-1β and IL-6). Sandwich ELISA assay was performed according to the manufacturer’s instructions as follows: 96-well plates were incubated with samples (100 μl) and standards (100 μl) for 90 min at 37 °C without any wash. Next, the plates were incubated with detection Abs (100 μl per well) at 37 °C for 60 min followed by 4 washes by 0.01 M TBS. Next, 100 μl ABC complex (per well) were added into plates, incubated at 37 °C for 30 min, and then washed with 0.01 M TBS for 5 times. Finally, the plates were incubated with TMB substrate solution and the reaction was stopped by the addition of 100 μl of 1 N HCl. All results were tested using the Thermo Scientific Microplate Reader (Thermo Scientific, Shanghai, China), and all the samples were analyzed at least three times.
Statistical Analysis
Results are shown as average values ± SEM. One-way ANOVA and nonparametric comparison (Bonferroni’s multiple comparison test) were performed using GraphPad Prism 7 software. P values less than 0.05 were considered significant.
RESULTS
Glycitin-Inhibited TNF-α Induced Inflammatory Response and Degeneration of Chondrocytes
As an aseptic inflammatory disease, OA is caused by an inflammatory response induced by mechanical stress and other physical and chemical factors and characterized by cartilage degeneration. Among the chemical factors, inflammatory cytokines, especially TNF-α, which was used in this study to stimulate chondrocyte, play a key role in inflammatory cartilage degeneration. In this study, before stimulation with TNF-α, viability of chondrocytes cultured with various concentrations of glycitin was examined by CCK8. As shown in Fig. 1a, low doses of glycitin (0–100 μg/ml) had no significant effect on chondrocyte viability. Thus, in this study, chondrocytes were treated with 25 μg/ml or 75 μg/ml glycitin. Nitric oxide (NO), produced and released by nitric oxide synthase (NOS) upon inflammation, was also analyzed by the Griess experiment. As indicated in Fig. 1b, less nitrite was produced in chondrocytes when stimulated with TNF-α in the presence of 0–100 μg/ml glycitin. As has been previously reported, inflammatory cytokines such as IL-1β, iNOS, MMP13, ADAMTS-5, COX-2, and IL-6 play crucial roles in the progression of OA. As described above, chondrocytes were stimulated with 10 ng/ml TNF-α in the presence or absence of glycitin, and total proteins were extracted and analyzed by western blot. As Fig. 1c shows, the expression levels of iNOS and COX-2 were significantly increased when chondrocytes were stimulated with TNF-α, and this effect was inhibited by the addition of glycitin. Moreover, mRNA was extracted from each group and real-time PCR was performed. The mRNA levels of IL-1β, IL-6, iNOS, and COX-2 were also increased after treatment of chondrocytes with TNF-α, and this effect was inhibited by the addition of glycitin (Fig. 1d–g). Furthermore, the levels of released IL-1β and IL-6 were in line with their gene transcription levels (Fig. 1l–m). The above results indicated that the inflammatory reaction caused by TNF-α in chondrocyte was significantly repressed by the treatment of glycitin. To assay for the extent of cartilage degeneration, we also measured the levels of collagen II and aggrecan, which are components of the cartilage matrix. As shown in Fig. 1c and j–k, glycitin treatment restored their levels that were reduced following treatment with TNF-α. Moreover, MMP13 and ADAMTS-5 have been shown to closely associate with cartilage degeneration. In this study, the mRNA levels of MMP13 and ADAMTS-5 were also examined by real-time PCR. Figure 1h–i indicates that MMP13 and ADAMTS-5 gene transcription of was increased following TNF-α stimulation but repressed by glycitin. Collectively, these results suggest that glycitin may suppress TNF-α-induced inflammatory response and associated cartilage degeneration in chondrocytes.

Glycitin antagonized cartilage degeneration and inflammation induced by TNF-α in vitro. a Cell viability was assayed by the CCK8 test. The values are mean ± S.E.M of at least three independent experiments. b Analysis of NO production in culture medium was performed by Griess experiments. c The protein levels of iNOS, COX2, and aggrecan in each group were assayed by western blot (relative quantitation was shown in Fig. S1). d–k mRNA levels of inflammatory biomarkers including IL-1β, iNOS, MMP13, ADAMTS-5, COX-2, and IL-6 were assayed in each group by real-time PCR. l–m The levels of IL-1β and IL-6 in culture medium of all groups were examined by ELISA. #P < 0.05; ##P < 0.01 versus CTL group; *P < 0.05; **P < 0.01 versus TNF-α stimulated group; NS, not significant versus CTL group.
Glycitin Attenuated Cartilage Degeneration in ACLT Mice Model
ACTL mouse model was established as above to investigate the therapeutic effect of glycitin in the knee joint of mice. One hundred and fifty microliters of PBS or glycitin (10 mg/ml) were intraperitoneally injected into mice of the ACLT group and the treatment group. Serum and knee joints of each mouse were collected and analyzed by ELISA, western blot, histological assays, etc. Figure 2a is a representative staining from each group, which was performed by Safranin O and Fast Green. As shown in Fig. 2a, glycitin significantly alleviated the degree of cartilage degeneration of knee joints after ACLT surgery. To further quantify the degree of cartilage degeneration, the OARSI score was calculated as described above for histological grading analysis. As Fig. 2b indicates, the OARSI scores were markedly higher in the ACLT surgery group compared to CTL group. However, the comparison of the ACLT group and the glycitin treatment group indicated that cartilage degeneration was inhibited following treatment with glycitin Moreover, cartilage PG depletion, cartilage surface erosion, and chondrocyte death in the three groups were in line with OARSI score (Fig. 2c–e). Apart from histological analysis, the biomarkers of cartilage degeneration were also examined in each group. Collagenase 3, which is also named Matrix metalloproteinase-13 (MMP13), could decompose the cartilage by degrading collage II. Moreover, as an extracellular matrix (ECM) degrading enzyme, Aggrecanase-2, also called A disintegrin and metalloproteinase with thrombospondin motifs 5 (ADAMTS-5), cleaves aggrecan in cartilage. Collagen II and aggrecan are considered the most important ingredients of cartilage. Thus, the expression of collagen II, MMP13, and ADAMTS-5 in knee joint tissues in all groups were examined by IHC. The results indicated that glycitin had a therapeutic effect on cartilage degeneration (Fig. 2f). Furthermore, the mRNA levels of collagen II, aggrecan, MMP13, and ADAMTS-5 were examined by real-time PCR, and identical results are presented in Fig. 2g–j.

Glycitin protected against cartilage degeneration in the ACLT mouse model in vivo. a Cartilage degeneration present in the CTL, ACLT, and glycitin treatment groups, assayed by Safranin O staining (black arrow, cartilage PG depletion; red arrow, cartilage surface erosion; yellow arrow, chondrocyte death). b the OARSI score of osteoarthritis in the two groups was calculated based on the results of Safranin O staining. c–e Treatment with glycitin protected cartilage stability in the ACLT model, assayed by histological score analysis according to the score of cartilage PG depletion, cartilage surface erosion, and chondrocyte death. f the expression levels of collagen II, aggrecan, and MMP13 were assayed by immunohistochemistry (relative quantitation was shown in Fig. S2F). g–j the mRNA levels of collagen II, aggrecan, MMP13, and ADAMTS-5 were examined by real-time PCR. The values are mean ± S.E.M of at least three independent experiments. *P < 0.05; **P < 0.01. Scale bar at 50 μm. N = 6 for each group.
Glycitin Suppressed Inflammatory Response in vivo
The inflammatory reaction of these three groups was also assayed. Serum was collected from all mice to measure the levels of circulating IL-1β, TNF-α, IL-6, and PGE2 by ELISA. The levels of the circulating inflammatory cytokines mentioned above were increased after ACLT surgery but decreased after glycitin treatment (Fig. 3a–d). Knee joint tissues were collected from each mouse and total mRNA and protein were extracted and followed by real-time PCR and western blot analysis. As shown in Fig. 3e–j, the mRNA levels of the inflammatory cytokines IL-1β, iNOS, TNF-α, COX-2, IL-6, and PEG2 were significantly inhibited when mice that underwent ACLT surgery were treated with glycitin. Moreover, the expression levels of iNOS and COX-2 were examined by immunostaining (Fig. 3h) and western blot analysis (Fig. 3i). The results were in line with those regarding mRNA levels. Collectively, our results indicate that treatment with glycitin decreased cartilage degeneration and reduced inflammation in arthritis in ACLT osteoarthritis mouse model.

Glycitin protected against the inflammatory response in ACLT mouse model in vivo. a–d The circulating levels of IL-1β, TNF-α, IL-6, and PGE2 in serum in each group were assayed by ELISA. e–j the mRNA levels of IL-1β, iNOS, TNF-α, COX-2, IL-6, and PGE2 in the knee joint were assayed by real-time PCR. The protein expression levels of iNOS and COX2 in the cartilage of the knee in all groups was tested by i western blot and j immunostaining (relative quantitation was shown in Fig. S3I-J). The values are mean ± S.E.M of at least three independent experiments. *P < 0.05; **P < 0.01. Scale bar at 50 μm. N = 6 for each group.
Glycitin Antagonized Activation of NF-κB Signaling Pathways in OA
As has been previously reported, NF-κB signaling pathway plays a crucial role in cartilage degeneration and inflammation. Glycitin was shown to inhibit the activation of NF-κB signaling pathway, which suggested that the positive effect of glycitin in OA may be achieved through the repression of NF-κB signaling pathway. When NF-κB signaling pathway is activated, the phosphorylation of IκBα and P65 is markedly upregulated, and nuclear translocation of NF-κB-P65 is accelerated. In the current study, the activation of NF-κB signaling pathway was examined both in vivo and in vitro. As shown in Fig. 4a–b, the mRNA levels of NF-κB1 and the phosphorylation of P65 and IκBα were significantly upregulated upon treatment of chondrocytes with TNF-α. However, treatment with glycitin restored the levels of NF-κB1 mRNA and reversed the phosphorylation of P65 and IκBα. Moreover, the subcellular localization of P65 was examined by immunofluorescence. TNF-α-induced nuclear translocation of P65 was inhibited by glycitin (Fig. 4e). Identical results were obtained in vivo (Fig. 4c, d). Furthermore, the phosphorylation of IκBα was investigated by immunohistochemistry in vivo. As Fig. 4f indicates, compared with the control group, IκBα phosphorylation was upregulated in the ACLT group and restored by glycitin treatment. Collectively, our data suggested that glycitin may affect the inflammatory response and cartilage degeneration via the NF-κB signaling pathways.

Glycitin antagonizes the activation of NF-κB signaling pathway both in vivo and in vitro the mRNA levels of NF-κΒ1 were assayed by real-time PCR both ain vitro and cin vivo. b The phosphorylation of IκBα and P65 in the control group, TNF-α group, and glycitin treatment group were examined by western blotting (relative quantitation was shown in Fig. S4B). d The phosphorylation of IκBα and P65 in the control group, ACLT, and glycitin treatment group were tested by western blotting (relative quantitation was shown in Fig. S4D). e Immunofluorescent staining was performed in chondrocytes to further investigate nuclear translocation of NF-κB-P65. f (Relative quantitation was shown in Fig. S4E), the expression of P-IκB in the ACLT mouse model were assayed by (relative quantitation was shown in Fig. S4F). The values are mean ± S.E.M of at least three independent experiments. *P < 0.05; **P < 0.01.
DISCUSSION
Cartilage, mainly comprised of cartilage matrix and chondrocytes, is a crucial component of the joint. Osteoarthritis, predominantly characterized by cartilage destruction and inflammatory osteogenesis in the whole joint, is a chronic disease that results in enormous financial loss because of the absence of effective treatment except for total joint replacement surgery in late periods [29, 30]. Thus, further studies are needed to understand its pathogenesis and to develop a curative treatment. It has been reported that TNF-α is a key inflammatory cytokine which could accelerate cartilage degeneration and inflammation in osteoarthritis [7, 31]. Collagen II and aggrecan are crucial ingredients of the cartilage matrix, and their levels are controlled by proteases such as MMPs and ADAMTS. Interestingly, it has been shown that TNF-α upregulated the mRNA and protein levels of MMPs and ADAMTS [32, 33]. Thus, TNF-α is a novel target for OA therapy. Moreover, it has already been shown that NF-κB signaling pathway plays a critical role in the occurrence and development of osteoarthritis [34, 35]. Additionally, in cartilage degeneration, activation of NF-κB signaling pathway by TNF-α has been reported previously [36].
Glycitin is a soy isoflavone, which is a flavonoid compound and a secondary metabolite formed during the growth of soybeans. Nowadays, an increasing number of studies demonstrate that components of soybeans have antibacterial, antiviral, and estrogenic effects [21, 24, 25]. Moreover, anti-inflammatory effects have recently been shown. Furthermore, it has been reported that glycitin played a positive role in osteoblasts in bone marrow stem cells [19, 37]. However, the unique effect of glycitin on OA and its associated underlying mechanism still need to be investigated. In this study, we demonstrated the protective effect of glycitin in osteoarthritis and clarified the underlying mechanism.
As previously reported, nitric oxide (NO), regulated by nitric oxide synthase (NOS), especially inducible nitric oxide synthase (iNOS), is a crucial cytotoxic mediator of inflammatory processes [10]. It is widely accepted that the production and activation of MMPs are promoted by NO and that the synthesis of collagen II and proteoglycans are suppressed by NO, resulting in ECM degradation in osteoarthritis [13]. In normal conditions, iNOS expression is very low, but its expression is increased upon inflammation. Cyclo-oxygenase 2 (COX2), which is also expressed at high levels upon inflammation, is able to activate MMPs to promote cartilage degeneration. Interestingly, the regulation of the mRNA and protein levels of COX2 is still unclear, but it has been reported that NO could upregulate COX2 expression [38]. Moreover, NF-κB could specifically bind to promoters of iNOS and COX2, while iNOS and COX2 could activate NF-κB signaling pathway, forming a positive feedback mechanism [34]. Furthermore, as has been previously reported, expression of iNOS is activated by inflammatory cytokines, such as IL-1β, TNF-α, and IL-6, and COX2 could increase the expression of IL-1β, TNF-α, IL-6, and PGE2 [13]. In this study, the production of NO in chondrocytes was stimulated by TNF-α and restored by glycitin. Moreover, the increased expression of detrimental inflammatory cytokines such as IL-1β, iNOS, ADAMTS-5, COX-2, and IL-6, in chondrocytes was significantly antagonized by the addition of glycitin at both low and high doses. Collectively, the above results indicated that glycitin has an anti-inflammatory effect in vitro.
The OA mouse model of anterior cruciate ligament transection (ACLT) has been previously established to investigate the pathogenesis and treatment of osteoarthritis because of significant similarities between the ACLT mouse model and human OA, including subchondral changes, articular cartilage damage, and synovitis [39]. The inflammatory response in the ACLT mouse model was remarkably inhibited following glycitin treatment. Moreover, the degeneration and destruction of articular cartilage damage and the subchondral changes progressed slowly in the glycitin treatment group.
In this study, the positive effect of glycitin in cartilage degeneration and inflammation in OA was verified both in vivo and in vitro. However, the effect in patients is still unknown. Moreover, the security of daily treatment with glycitin via i.p. injection still needs to further confirm not only in mouse but also in humans. Currently, there are no studies on the effectiveness of glycitin in patients with osteoarthritis. However, elucidation of the effective mechanism of glycitin treatment in OA in mice provides the basis to investigate the effect of glycitin in patients in the future. As previously reported, glycitin alleviates lipopolysaccharide-induced acute lung injury via inhibiting NF-κB and MAPKs pathway activation. Thus, not only NF-κB pathway activation but also MAPKs pathway were measured in this study. As Fig. S1 indicated, the phosphorylation of ERK1/2 was upregulated in vitro but downregulated with the treatment of glycitin, which indicated that the effects of glycitin are not NFkB-specific. Therefore, maybe there is another mechanism of the glycitin treatment in osteoarthritis. We will further investigate it in the future.
At present, the widely used medicines are still the NSAIDs, which are effective against inflammation but have various side effects, such as peptic ulcer and cardiovascular events [40]. Moreover, NSAIDs do not significantly inhibit cartilage degeneration. Thus, glycitin, which inhibits both inflammation and cartilage degeneration, is a potential novel and suitable target for medicinal development in OA.
References
Thielen, N.G.M., P.M. van der Kraan, and A.P.M. van Caam. 2019. TGFbeta/BMP signaling pathway in cartilage homeostasis. Cells 8.
Zhu, S., D. Makosa, B. Miller, and T.M. Griffin. 2019. Glutathione as a mediator of cartilage oxidative stress resistance and resilience during aging and osteoarthritis. Connective Tissue Research: 1–14.
Teirlinck, C.H., D.M.J. Dorleijn, P.K. Bos, J.B.M. Rijkels-Otters, S.M.A. Bierma-Zeinstra, and P.A.J. Luijsterburg. 2019. Prognostic factors for progression of osteoarthritis of the hip: a systematic review. Arthritis Research & Therapy 21: 192.
Lane, N.E., K. Shidara, and B.L. Wise. 2017. Osteoarthritis year in review 2016: clinical. Osteoarthritis and Cartilage 25: 209–215.
Flemming, D.J., and C.N. Gustas-French. 2017. Rapidly progressive osteoarthritis: a review of the clinical and radiologic presentation. Current Rheumatology Reports 19: 42.
Haj-Mirzaian A, Mohajer B, Guermazi A, Conaghan PG, Lima JAC, Blaha MJ, et al. 2019. Statin use and knee osteoarthritis outcome measures according to the presence of Heberden nodes: results from the osteoarthritis initiative. Radiology 190557.
Kapoor, M., J. Martel-Pelletier, D. Lajeunesse, J.P. Pelletier, and H. Fahmi. 2011. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nature Reviews Rheumatology 7: 33–42.
Zhao, Y.P., B. Liu, Q.Y. Tian, J.L. Wei, B. Richbourgh, and C.J. Liu. 2015. Progranulin protects against osteoarthritis through interacting with TNF-alpha and beta-Catenin signalling. Annals of the Rheumatic Diseases 74: 2244–2253.
Hossain, F.M., Y. Hong, Y. Jin, J. Choi, and Y. Hong. 2019. Physiological and pathological role of circadian hormones in osteoarthritis: dose-dependent or time-dependent? Journal of Clinical Medicine 8.
Lai, Y., X. Bai, Y. Zhao, Q. Tian, B. Liu, E.A. Lin, Y. Chen, B. Lee, C.T. Appleton, F. Beier, X.P. Yu, and C.J. Liu. 2014. ADAMTS-7 forms a positive feedback loop with TNF-alpha in the pathogenesis of osteoarthritis. Annals of the Rheumatic Diseases 73: 1575–1584.
Zhao, Y., Y. Li, R. Qu, X. Chen, W. Wang, C. Qiu, B. Liu, X. Pan, L. Liu, K. Vasilev, J. Hayball, S. Dong, and W. Li. 2019. Cortistatin binds to TNF-alpha receptors and protects against osteoarthritis. Ebiomedicine 41: 556–570.
Mei, J., J. Sun, J. Wu, and X. Zheng. 2019. Liraglutide suppresses TNF-alpha-induced degradation of extracellular matrix in human chondrocytes: a therapeutic implication in osteoarthritis. American Journal of Translational Research 11: 4800–4808.
Martinez-Moreno, D., G. Jimenez, P. Galvez-Martin, G. Rus, and J.A. Marchal. 1865. Cartilage biomechanics: a key factor for osteoarthritis regenerative medicine. Biochimica et Biophysica Acta - Molecular Basis of Disease 2019: 1067–1075.
Hermann, J., R.W. Lipp, A. Dunzinger, C. Spreizer, G. Schaffler, H. Kvaternik, et al. 2014. Anti-TNF scintigraphy to assess TNF-alpha-associated joint inflammation in rheumatoid arthritis and osteoarthritis. Clinical and Experimental Rheumatology 32: 614.
Nelson, A.E. 2018. Osteoarthritis year in review 2017: Clinical. Osteoarthritis and Cartilage 26: 319–325.
Ibi, M. 2019. Inflammation and temporomandibular joint derangement. Biological & Pharmaceutical Bulletin 42: 538–542.
Wang, A., D.J. Leong, L. Cardoso, and H.B. Sun. 2018. Nutraceuticals and osteoarthritis pain. Pharmacology & Therapeutics 187: 167–179.
Leong, D.J., M. Choudhury, D.M. Hirsh, J.A. Hardin, N.J. Cobelli, and H.B. Sun. 2013. Nutraceuticals: potential for chondroprotection and molecular targeting of osteoarthritis. International Journal of Molecular Sciences 14: 23063–23085.
Zhang, L., J. Chen, W. Chai, M. Ni, X. Sun, and D. Tian. 2016. Glycitin regulates osteoblasts through TGF-beta or AKT signaling pathways in bone marrow stem cells. Experimental and Therapeutic Medicine 12: 3063–3067.
Chen, Y., S. Guo, K. Jiang, Y. Wang, M. Yang, and M. Guo. 2019. Glycitin alleviates lipopolysaccharide-induced acute lung injury via inhibiting NF-kappaB and MAPKs pathway activation in mice. International Immunopharmacology 75: 105749.
van der Velpen, V., A. Geelen, P.C. Hollman, E.G. Schouten, P. van’t Veer, and L.A. Afman. 2014. Isoflavone supplement composition and equol producer status affect gene expression in adipose tissue: a double-blind, randomized, placebo-controlled crossover trial in postmenopausal women. The American Journal of Clinical Nutrition 100: 1269–1277.
Choi, I., Y. Kim, Y. Park, H. Seog, and H. Choi. 2007. Anti-obesity activities of fermented soygerm isoflavones by Bifidobacterium breve. Biofactors 29: 105–112.
Seo, G.Y., Y. Lim, D. Koh, J.S. Huh, C. Hyun, Y.M. Kim, et al. 2017. TMF and glycitin act synergistically on keratinocytes and fibroblasts to promote wound healing and anti-scarring activity. Experimental & Molecular Medicine 49: e302.
Kim, E.Y., K.B. Hong, H.J. Suh, and H.S. Choi. 2015. Protective effects of germinated and fermented soybean extract against tert-butyl hydroperoxide-induced hepatotoxicity in HepG2 cells and in rats. Food & Function 6: 3512–3521.
Kang, K.A., R. Zhang, M.J. Piao, K.H. Lee, B.J. Kim, S.Y. Kim, H.S. Kim, D.H. Kim, H.J. You, and J.W. Hyun. 2007. Inhibitory effects of glycitein on hydrogen peroxide induced cell damage by scavenging reactive oxygen species and inhibiting c-Jun N-terminal kinase. Free Radical Research 41: 720–729.
Valachovicova, T., V. Slivova, H. Bergman, J. Shuherk, and D. Sliva. 2004. Soy isoflavones suppress invasiveness of breast cancer cells by the inhibition of NF-kappaB/AP-1-dependent and -independent pathways. International Journal of Oncology 25: 1389–1395.
Zhang, B., H. Chen, J. Ouyang, Y. Xie, L. Chen, Q. Tan, et al. 2019. SQSTM1-dependent autophagic degradation of PKM2 inhibits the production of mature IL1B/IL-1beta and contributes to LIPUS-mediated anti-inflammatory effect. Autophagy: 1–17.
Li, W., W. Wang, L. Liu, R. Qu, X. Chen, C. Qiu, J. Li, J. Hayball, L. Liu, J. Chen, X. Wang, X. Pan, and Y. Zhao. 2019. GDF11 antagonizes TNF-alpha-induced inflammation and protects against the development of inflammatory arthritis in mice. The FASEB Journal 33: 3317–3329.
Hosseinzadeh, A., S.K. Kamrava, M.T. Joghataei, R. Darabi, A. Shakeri-Zadeh, M. Shahriari, R.J. Reiter, H. Ghaznavi, and S. Mehrzadi. 2016. Apoptosis signaling pathways in osteoarthritis and possible protective role of melatonin. Journal of Pineal Research 61: 411–425.
Dieppe, P., S. Goldingay, and M. Greville-Harris. 2016. The power and value of placebo and nocebo in painful osteoarthritis. Osteoarthritis and Cartilage 24: 1850–1857.
Nees, T.A., N. Rosshirt, T. Reiner, M. Schiltenwolf, and B. Moradi. 2019. Inflammation and osteoarthritis-related pain. Schmerz 33: 4–12.
Rainsford, K.D., A.L. Parke, M. Clifford-Rashotte, and W.F. Kean. 2015. Therapy and pharmacological properties of hydroxychloroquine and chloroquine in treatment of systemic lupus erythematosus, rheumatoid arthritis and related diseases. Inflammopharmacology 23: 231–269.
Mobasheri, A., Y. Henrotin, H.K. Biesalski, and M. Shakibaei. 2012. Scientific evidence and rationale for the development of curcumin and resveratrol as nutraceutricals for joint health. International Journal of Molecular Sciences 13: 4202–4232.
Rigoglou, S., and A.G. Papavassiliou. 2013. The NF-kappaB signalling pathway in osteoarthritis. The International Journal of Biochemistry & Cell Biology 45: 2580–2584.
Liang, H., H. Wang, L. Luo, S. Fan, L. Zhou, Z. Liu, S. Yao, X. Zhang, K. Zhong, H. Zhao, and Z. Zha. 2019. Toll-like receptor 4 promotes high glucose-induced catabolic and inflammatory responses in chondrocytes in an NF-kappaB-dependent manner. Life Sciences 228: 258–265.
Xie, L., H. Xie, C. Chen, Z. Tao, C. Zhang, and L. Cai. 2019. Inhibiting the PI3K/AKT/NF-kappaB signal pathway with nobiletin for attenuating the development of osteoarthritis: in vitro and in vivo studies. Food & Function 10: 2161–2175.
Li, X.H., J.C. Zhang, S.F. Sui, and M.S. Yang. 2005. Effect of daidzin, genistin, and glycitin on osteogenic and adipogenic differentiation of bone marrow stromal cells and adipocytic transdifferentiation of osteoblasts. Acta Pharmacologica Sinica 26: 1081–1086.
Li, H., D. Wang, Y. Yuan, and J. Min. 2017. New insights on the MMP-13 regulatory network in the pathogenesis of early osteoarthritis. Arthritis Research & Therapy 19: 248.
Guo, L., X. Wei, Z. Zhang, X. Wang, C. Wang, P. Li, et al. 2019. Ipriflavone attenuates the degeneration of cartilage by blocking the Indian hedgehog pathway. Arthritis Research & Therapy 21: 109.
Honvo, G., V. Leclercq, A. Geerinck, T. Thomas, N. Veronese, A. Charles, V. Rabenda, C. Beaudart, C. Cooper, J.Y. Reginster, and O. Bruyère. 2019. Safety of topical non-steroidal anti-inflammatory drugs in osteoarthritis: outcomes of a systematic review and meta-analysis. Drugs & Aging 36: 45–64.
Funding
This work was supported by the Natural Science Foundation of Shandong Province (Grant Nos. ZR2019MH05, ZR2019MH075, and BS2015SW028), the Key Research and Development Projects of Shandong Province (Nos. 2015GSF118115 and 2019GSF108152), the Cross-disciplinary Fund of Shandong University (Grant No. 2018JC007), and the National Natural Science Foundation of China (Grant Nos. 81501880 and 81602761).
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: Hao Li, Xin Pan.
Performed the experiments: Wenhan Wang, Ruitong Yang, Minfa Zhang, Jiayi Li, Jiangfan Peng, Mingyang Xu, Yunpeng Zhao.
Corresponding authors
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.




Rights and permissions
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Wang, W., Yang, R., Zhang, M. et al. Glycitin Suppresses Cartilage Destruction of Osteoarthritis in Mice. Inflammation 43, 1312–1322 (2020). https://doi.org/10.1007/s10753-020-01210-3
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10753-020-01210-3