
Abstract
Taxonomic diversity of desmids and other coccoid green algae is discussed in relation to different species concepts. For want of unambiguous criteria about species delimitation, no reliable estimations of global species richness can be given. Application of the biological species concept is seriously hampered by lack of sexual reproduction in many species. Molecular analyses demonstrated cases of close affiliation between morphologically highly different taxa and, contrary, examples of little relationship between morphologically similar taxa. Despite the fact that desmids and chlorococcal algae, because of their microbial nature, can be readily distributed, cosmopolitan species are relatively scarce. The geographic distribution of some well-recognizable morphospecies is discussed in detail. Of some species a recent extension of their area could be established, e.g., in the desmids Micrasterias americana and Euastrum germanicum, and in the chlorococcaleans Desmodesmus perforatus and Pediastrum simplex.
Similar content being viewed by others

Introduction
This review focuses on the diversity and geographic distribution of some groups of green algae showing a coccoid level of organization but belonging to different taxonomic units. According to modern systematic views, the desmids (Desmidiales) are placed in the divison Charophya (‘Streptophyta’), class Zygnemophyceae (Lewis and McCourt 2004). Desmids are coccoid and have a striking morphology characterized by two symmetrical halves (semicells). They comprise both solitary and colonial taxa. The other coccoid green algae studied here (in the following text designated as ‘chlorococcal algae’), were formerly artificially classified under the Chlorococcales sensu lato and are now grouped in several orders of Chlorophyceae, Trebouxiophyceae and Prasinophyceae (Krienitz et al. 2003; Lewis and McCourt 2004). These orders mainly contain solitary or colonial algae with a spherical, ellipsoidal or needle-shaped morphology. We selected such diverse groups of green micro-algae in order to show the different state of the art in research on diversity and geography of these tiny protists.
Geographically the best studied group of green algae is that of the desmids, due to their often appealing appearance. Samples from far abroad revealed a lot of astonishing, exotic forms. Reliable knowledge of geographical distribution patterns, of course, is confined to those taxa that cannot be confused with any other ones. Fortunately, among the desmids quite a number of such taxa may be designated. Particularly the genus Micrasterias is marked by a high percentage of well recognizable species the distribution of which is confined to relatively small parts of the world (Figs. 1–5). In a previous paper, Coesel (1996) distinguished 10 desmid floral regions: Indo-Malaysia/Northern Australia, Equatorial Africa, Tropical South and Central America, North America, Extratropical South America, Eastern Asia, Southern Australia and New Zealand, South Africa, Temperate Eurasia and, finally, the circumpolar and high mountain regions. The number of species supposed to be endemic to any of those regions roughly speaking decreases from over a hundred to less than 10 and presumably goes hand in hand with the total desmid species diversity to be encountered.

Desmids with limited geographical distribution (1–5) and cryptic (6, 7) and synonymous (8–13) species. 1: M. hardyi (S.E. Australia; after Krieger 1939); 2: M. sudanensis (tropical Africa; after Thomasson 1960); 3: M. depauperata (American continents; after Krieger 1939); 4: M. muricata (North America; after Krieger 1939); 5: M. ceratophora (S.E. Asia and N. Australia: after Scott and Prescott 1961). 6, 7: Closterium ehrenbergii (vegetative cell and sexual reproduction stage), a morphospecies consisting of at least 12 different biological species (original). 8–13: Six Cosmarium species which are probably synonymous 8: C. wallichii West and West (after Wallich 1860, as Euastrum clepsydra Wall.); 9: C. seelyanum Wolle (after Wolle 1884); 10: Cosmarium nobile (Turner) Krieg. (after Turner 1892, as Euastrum nobile Turner); 11: C. naivashensis Rich (after Rich 1932); 12: C. divergens Krieg. (after Krieger 1932); 13: C. subnobile Hinode (after Hinode 1966)
As compared to the desmids, the study of diversity and distribution of chlorococcal algae presents more difficulties because of a high degree of morphological uniformity (‘green balls’) on the one hand and an extreme phenotypic variability of colony structure and cell wall equipment such as bristles, spines, ornamentations and incrustations on the other. Therefore, the conventional morphological species concept does not reflect the real diversity. Most of the classical phycologists describing chlorococcal algal species from tropical and polar regions (for details see Komárek and Fott 1983) did not collect the material themselves. They examined fixed samples taken from scientific travellers and their species descriptions are usually difficult to link to modern, DNA-based views of chlorococcal taxonomy.
Both for desmids and chlorococcal algae it holds that experimental systematic studies, including ecophysiological tests and molecular sequence analyses on ‘exotic’ species are badly needed.
Species concepts in desmids and chlorococcal algae
Problems concerning species definition in microalgae considerably hamper the interpretation of algal biodiversity and biogeography. The morphologic species concept considers species as groups of morphologically identical or similar organisms (Futuyma 1998). The biologic species concept defines species as groups of interbreeding populations which are reproductively isolated from other groups (Mayr 1942). However, both species concepts give problems when applied to algal species. In this context, John and Maggs (1997) come to the conclusion that at present no operational species concept is available for eukaryotic algae.
Traditional, morphology-based desmid taxonomy is overloaded with synonyms and suffers from a high rate of splitting (see Diversity chapter below). Unfortunately, sexual reproduction—essential for applying the biologic species concept—is a relatively rare phenomenon in this algal group (of many species no sexual stages are known at all). In addition, desmids are haploid organisms, so most mutations are immediately expressed. Consequently, by predominant lack of sexual reproduction (so possible exchange of genes during meiotic cell division) genotypically determined morphological variation is not wiped out. Therefore, exclusively clonal reproduction (particularly in euplanktic species) may readily result in the formation of desmid microspecies, just like in apomictically reproducing macrophytes (Coesel and Joosten 1996).
One of the possible reasons that sexual reproduction stages (zygospores) are usually encountered only incidentally is that but few clonal populations are homothallic (i.e., self-fertile). In experiments, out of some 120 randomly selected desmid strains belonging to 16 genera and over 80 species, only three showed homothallic sexual reproduction (Coesel and Teixeira 1974). On the other hand, mating experiments between different clones, often originating from different sites, revealed the phenomenon of heterothallism in, e.g., Closterium ehrenbergii Ralfs (Ichimura 1981; Coesel 1988), Closterium strigosum Bréb. (Watanabe and Ichimura 1978), Pleurotaenium mamillatum West (Ling and Tyler 1976) and Micrasterias thomasiana Archer (Blackburn and Tyler 1987). Especially in Closterium ehrenbergii (Figs. 6, 7), many mating types have been demonstrated, that is, populations which mutually show (almost) complete sexual isolation (Ichimura 1981). Such mating types can be considered syngens or biological species. Often, but not necessarily, mating types of one and the same morphospecies slightly differ in morphology, ecology and/or geographical distribution (Ichimura and Kasai 1990; Ichimura et al. 1997). No doubt, such sibling species—also to be traced from DNA analyses (Denboh et al. 2003)—will occur in many more morphospecies, and it is clear that they will substantially increase the diversity of this algal group.
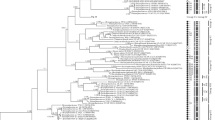
Also in chlorococcal algal taxonomy, the morphologic species concept is burdened with a high degree of misinterpretation. For example, one and the same morphotype, such as the globular ‘green ball’, has evolved in different phylogenetic lineages. On the other hand, highly diverse morphotypes can belong to one and the same lineage (Luo et al. 2005). This ambivalent interpretation of morphotypes was documented on the archetypical green ball Chlorella and relatives. In freshwater and brackish habitats several distinct lineages of spherical ‘Chlorella-like’ green algae have been found and designated as separate clades: Mychonastes/Pseudodictyosphaerium, now placed in the Chlorophyceae; and Choricystis/Nanochlorum/Chlorella, now placed in the Trebouxiophyceae (Krienitz et al. 1999; Hepperle and Krienitz 2001). In marine habitats, several other lineages of globular green algae have been reported, e.g., in the prasinophytes Pycnococcus provasolii Guillard, now placed in the Pseudoscourfieldiales (Daugbjerg et al. 1995); and Ostreococcus tauri Courties et Chrétiennot-Dinet, now placed in the Mamiellales (Courties et al. 1998). Further members of the Chlorella morphotype await taxonomic treatment and a transfer to other lineages of the Chlorophyta (Komárek and Fott 1983). The multiple origin of ‘Chlorella-like’ algae may be explained by an adaptive advantage of the coccoid morphotype in ecosystems (Potter et al. 1997). On the other hand, sequence analyses revealed that several morphologically different algae, that were grouped in different lineages, such as Closteriopsis, Actinastrum, Dictyosphaerium, Didymogenes and Micractinium cluster close to globular ‘true’ Chlorella sensu stricto species (Fig. 14).

Phylogenetic tree of Chlorellaceae derived from 18S rRNA gene sequences. The true, spherical Chlorella species, C. vulgaris, C. sorokiniana and C. lobophora, are intermixed by morphologically different taxa which were formerly placed within other families of coccoid green algae. Chlorella kessleri is transferred to the genus Parachlorella which is closely related to species of the needle-shaped Closteriopsis and Dicloster (according to Krienitz et al. 2004 and Luo et al. 2005)
In numerous chlorococcal algal species sexual reproduction is unknown. Therefore, the biologic species concept is not applicable. Fortunately, molecular analyses offer an alternative to get insight in their relationships. After the introduction of DNA sequencing and phylogenetic analyses, the systematics of algae is going through a dramatic phase of change. The recent situation is marked by the quest for a compromise between the conventional (artificial) and the phylogenetic system. This upheaval will have significant consequences on biodiversity and biogeography of algal taxa. Some examples of these changing views will be given in the following two chapters.
Diversity
According to Gerrath (1993), estimations of global desmid species richness highly vary, but for the most part will be too high in view of the fair number of current synonyms. To give here a single example of possible synonymy: Cosmarium seelyanum Wolle, C. wallichii West and West, C. divergens Krieg., C. naivashensis Rich, C. nobile (Turner) Krieg., and C. subnobile Hinode are possibly the same species (Figs. 8–13), which occur in the literature also under the name of C. abruptum Lundell. Minor morphologic differences often are within the phenotypic variability of a species, and are thus not of taxonomic significance. Many forms, at any time described as separate varieties or even species, appear to be interconnected by transitional forms, so most likely belong to one and the same taxon, e.g, the common species Staurastrum furcatum (Ralfs) Bréb. and Staurastrum aciculiferum (West) Andersson (Figs. 15–17). However, such causes for overestimation of species diversity are counterbalanced by reasons for underestimation. Different species the vegetative cells of which are very much alike in practice usually will not be recognized. Certain small-sized, smooth-walled Cosmarium species can only be identified reliably when zygospores are encountered (Figs. 18–23). As zygospores usually are met only incidentally, new species can be expected on account of zygospore morphology. Apart from possible under- and overestimations of species diversity, related to insufficient observations or failures in knowledge of relevant literature, a reliable assessment of species diversity is also hindered by the experience that polymorphism in a ‘species’ is not always phenotypic but also may have got a genetic base, like in sibling species.

Problems in species identification (15–23), flagship species (24), and species with restricted distribution (25–31). 15–17: Staurastrum furcatum (left) and S. aciculiferum (right) are interconnected by a transitional form (after Péterfi 1973). 18–23: Some morphologically similar, small-sized, smooth-walled Cosmarium species that can only be reliably identified by their zygospore. 18: C. majae K.Strøm (after Coesel and Meesters 2002); 19: C. asterosporum Coesel (after Coesel 1989); 20: C. pseudobicuneatum Jao (after Jao 1940); 21, 22, 23: unknown species still to be described (originals). 24. Cosmarium barramundiense Coesel, a recently described flagship species from northern Australia (after Coesel 2004). 25–28: Staurastrum species that have, within Europe, a distinctly atlantic-subarctic distribution (originals). 25: S. arctiscon; 26: S. ophiura (in top view); 27: S. maamense; 28: S. elongatum. 29, 30: Two desmids (Euastrum germanicum, Cosmarium striolatum) with a predominantly continental distribution within Europe (originals). 31: Micrasterias americana, a fast expanding desmid species in The Netherlands (original)
Taking into account all these complications it is fully understandable that estimations of total desmid species richness in (rather recent) literature range from 1,500 to 12,000 (Gerrath 1993). Gerrath (1993) thinks that there are approximately 3,000 ‘good’ desmid species worldwide. When extrapolating the number of morphospecies distinguished in an ongoing inventory of European Staurastra, the first author comes to a comparable number. Anyhow, it is remarkable that hardly any new desmid flagship species are found. A recent, rather extensive investigation in northern Australia revealed but a single ‘brand-new’ species, quite different from all taxa described before (Fig. 24). This could be an indication that the number of desmid species endemic to a relatively small geographical area is limited, which for a group of readily to be transported micro-organisms is not really surprising.
The most recent comprehensive monograph on chlorococcal algae (Komárek and Fott 1983) contains about 1,200 species and subspecific taxa. Estimations of the real number of species are extremely vague because of the pending situation in species concepts. To illustrate this, the former genus Scenedesmus (now split into the genera Scenedesmus, Desmodesmus, Acutodesmus) may be used as an example. As a result of the high variability in morphologic characters (shape and organization of coenobia, spines, incrustations, cell wall ornamentations), more than 1,300 (morpho)species and subspecies have been desribed (Hegewald and Silva 1988). Studies using unialgal cultures to estimate the morphological variability revealed a severe overestimation of species number (Hegewald et al. 1990; Trainor 1998; Hegewald 1999). These observations were supported by molecular studies (Hegewald 2000). On the other hand, combined studies on fine structure and gene sequence (ITS2) of 22 clones identified as Desmodesmus costato-granulatus (Skuja) Hegewald indicated a higher diversity than expected. According to this interdisciplinary approach, it was split into five species (Vanormelingen et al. 2007).
Another example of the ambivalent situation in estimation of species diversity is Botryococcus braunii Kützing considered to be a ‘well-known’ microplanktont of inland waters, showing typical, large colonies like bunches of grape. Komárek and Marvan (1992) collected 74 populations of Botryococcus-like algae worldwide, studied their morphologic characteristics, and found 13 different morphotypes which fulfilled the species status according to the commonly used criteria in chlorococcal algal taxonomy. Unfortunately, molecular studies on these algae are deficient. The only known study on the phylogenetic placement of Botryococcus is supporting a polyphyletic origin (Senousy et al. 2004).
Also taxon delimitation within the Selenastraceae, a family of needle-shaped and lunate chlorococcal algae, has experienced considerable changes in recent times. Komárková-Legnerová (1969) and Marvan et al. (1984) provided revisions of the Selenastraceae based on morphotypes. Hindák (1984) described several new species which are of intermediate morphology with respect to described species, e.g., Monoraphidium intermedium Hindák as an intermediate taxon of M. griffithii (Berk.) Kom.-Legn. and M. obtusum (Korsh.) Kom.-Legn. The first molecular phylogenetic study on the Selenastraceae (Krienitz et al. 2001) revealed an intermixing of common members of the genera Ankistrodesmus, Monoraphidium and Selenastrum which contradicts the traditional way of circumscription of genera and species in this family. Finally, Fawley et al. (2005) discovered cryptic species in the Selenastraceae. It was found that isolates of one and the same morphotype can differ in 18S rDNA sequences, whereas isolates with identical or similar 18S rDNA sequence can exhibit different morphologies. These results give further arguments for the necessity of interdisciplinary work in algal systematics and diversity.
Geographic distribution
Examples of peculiar, well-defined distribution patterns in desmids were already shown by Donat (1926) and refined by Heimans (1969). Most striking is the occurrence of a number of flagship species that, within Europe, are characterized by a marked atlantic-subarctic distribution: Staurastrum elongatum Barker, S. maamense Archer, S. arctiscon (Ralfs) Lundell, S. cerastes Lundell, S. ophiura Lundell, S. brasiliense Nordst., and S. sexangulare (Bulnh.) Lundell (Figs. 25–28). As we have to do with aquatic organisms, it is difficult to imagine which climatic factor(s) could be responsible for such a remarkable distribution pattern. Likely it is some ecological parameter, linked to the nearness of seawater, that is decisive for their occurrence. The above-mentioned species are also known from the North American continent, but their distribution over there seems to be less distinct (Prescott et al. 1982). Anyhow, even in the atlantic and subarctic regions of Europe none of these species is really common. From The Netherlands, out of the seven above-mentioned species, S. elongatum, S. ophiura, S. arctiscon, S. brasiliense and S. cerastes have been reported, but only from one or a few sites and only in the first half of the last century. Some of them were regularly found during a longer period, e.g., S. ophiura between 1912 and 1930. From the fact that none of these species succeeded in enlarging its regional area, it is suggested that ecological demands rather than climatic factors or dispersal abilities are limiting.
In Europe, versus atlantic-subarctic species, also continental desmid species can be distinguished. Striking examples are Cosmarium striolatum (Nägeli) Archer [synonymous with C. tesselatum (Delponte) Nordst.] and Euastrum germanicum (Schmidle) Krieg. (Figs. 29, 30). Both Cosmarium striolatum and Euastrum germanicum are widely distributed on the continent (Heimans 1969; Coesel 1978), but are absent from Great Britain (Brook and Williamson 1991). In this case, the lack might be attributed to the isolated position of the British Islands. A few decades ago, these species were extremely rare in The Netherlands, too. Yet, in recent years Dutch records, particularly of Euastrum germanicum, are remarkably increasing in number. Obviously, both species are advancing in western direction and it may be only a question of (relatively little) time that they reach England.
Possibly, the fast increasing number of Dutch records of Euastrum germanicum in the last decade has to do with increasing average year temperatures. In this context, also the expansion of another conspicuous Dutch desmid species in The Netherlands has to be stressed, i.e., of Micrasterias americana Ralfs (Fig. 31). Although The Netherlands have been intensively inventoried for desmids already from the beginning of the 20th century, this species was not recorded before 1952 (Heimans 1969). Since then the number of Dutch records steadily increased, but it is only in the last decade that it has become one of the most common Micrasterias species of The Netherlands, also found in disturbed habitats. This latter phenomenon, though, might be an indication of a changed genetic constitution enabling the filling of another (larger) ecological niche.
In contrast to the desmids, the chlorococcal algae are generally supposed to be ubiquitous and to have a cosmopolitan distribution. As such they would serve as a good example of Beijerinck’s metaphor, taken up by Baas-Becking and finally accentuated by Fenchel et al. (1997), Finlay (2002) and Fenchel and Finlay (2004): ‘in micro-organisms, everything is everywhere, the environment selects’. This statement has evoked a heated discussion focusing on the species concepts. In recent times, numerous articles have been published which contradict the ubiquity hypothesis (reviewed by Foissner 2006 and Logares 2006). For micro-algae this statement is still under disputation (Coleman 2002, Finlay and Fenchel 2002). In diatoms, Hillebrandt et al. (2001) revealed a decreasing similarity of species composition with increasing geographic distance, and thus they reject strict ubiquity of unicellular taxa. Coleman (2001) found local adaptation and endemism in phytoflagellates of the genera Pandorina and Volvulina.
As for the chlorococcal algae, there are indications of endemism both in classical, morphological and in modern, molecular approaches. Wille (1924) described the genus Soropediastrum which contains two species only found on the Kergueles. From the same Antarctic region Pediastrum marvillense Théréz. and Couté (Fig. 32) was discovered, which also seems to be endemic (Komárek and Jankovská (2001). Detailed studies on chlorococcal algae of Cuba demonstrated in 20% of the taxa slight morphological differences in comparison to the original descriptions based on material from the temperate zone (Komárek 1983; Comas 1996). The latter authors found 21 taxa only recorded from Cuba. Several morphospecies of Botryococcus studied by Komárek and Marvan (1992) are only known from a few localities, e.g., B. fernandoi Komárek and Marvan (Fig. 33). The large oocystacean Makinoella tosaensis Okada (Fig. 34) has been reported only from Japan and Korea (Hegewald et al. 1999). Amphikrikos variabilis Krienitz (Fig. 35), with a distinct pattern of cell wall incrustation, has been found only in swamps and rivers of Namibia (Krienitz 1998).

Some examples of chlorococcal algae with limited geographic distribution. 32: Pediastrum marvillense Théréz. and Couté (Antarctic; after Thérézien and Couté 1977); 33: Botryococcus fernandoi Komárek and Marvan (Ethiopia; after Komárek and Marvan 1992); 34: Makinoella tosaensis Okada (S.E. Asia; after Okada 1949); 35: Amphikrikos variabilis Krienitz (Namibia; after Krienitz 1998)
Though, there are also indications that several species enlarged their distribution area considerably during the last decades (‘invading species’), e.g., the formerly ‘tropical/subtropical’ species Desmodesmus perforatus (Lemmerm.) Hegewald and Pediastrum simplex Meyen. Nowadays, these species are encountered regularly in the temperate zone (Jeon and Hegewald 2006; Geissler and Kies 2003).
Slapeta et al. (2006) performed molecular analyses in the morphospecies Micromonas pusilla Butcher, a marine, picoplanktic prasinophyte. This morphospecies appeared to be a complex of morphologically indistinguishable phylogenetic lineages, representing cryptic species. Although some of these entities were shown to have a global, oceanic distribution, a more restricted distribution of other ones could not be precluded.
References
Blackburn SI, Tyler PA (1987) On the nature of eclectic species—a tiered approach to genetic compatibility in the desmid Micrasterias thomasiana. Br phycol J 22:277–298
Brook AJ, Williamson DB (1991) A check-list of desmids of the British Isles. Occasional publication 28, Freshwater Biological Association, Ambleside
Coesel PFM (1978) Taxonomical, geographical and ecological notes on Euastrum mononcylum var. germanicum Schmidle (Chlorophyta, Desmidiaceae). Arch Protistenk 120:436–445
Coesel PFM (1988) Biosystematic studies on the Closterium moniliferum/ehrenbergii complex (Chlorophyta, Conjugatophyceae) in western Europe. II. Sexual compatibility. Phycologia 27:421–424
Coesel PFM (1989) Taxonomic notes on Dutch desmids. Cryptogam Algol 10:181–193
Coesel PFM (1996) Biogeography of desmids. Hydrobiologia 336:41–53
Coesel PFM (2004) Some new and otherwise interesting desmid species from Kakadu National Park (Northern Australia). Quekett J Microsc 39:779–782
Coesel PFM, Joosten AMT (1996) Three new planktic Staurastrum taxa (Chlorophyta, Desmidiaceae) from eutrophic water bodies and the significance of microspecies in desmid taxonomy. Algol Stud 80:9–20
Coesel PFM, Meesters J (2002) Signalen van sieralgen. Natura 99:68–70
Coesel PFM, Teixeira RMV (1974) Notes on sexual reproduction in desmids. II. Experiences with conjugation experiments in uni-algal cultures. Acta Bot Neerl 23:603–611
Coleman AW (2001) Biogeography and speciation in the Pandorina/Volvulina (Chlorophyta) superclade. J Phycol 37:836–851
Coleman AW (2002) Microbial eukaryote species. Science 297:337
Comas AG (1996) Las Chlorococcales dulciacuícolas de Cuba. Biblthca phycol 99:1–192
Courties C, Perasso R, Chretiennot-Dinet MJ et al (1998) Phylogenetic analysis and genome size of Ostreococcus tauri (Chlorophyta, Prasinophyceae). J Phycol 34:844–849
Daugbjerg N, Moestrup O, Arctander P (1995) Phylogeny of genera of Prasinophyceae and Pedinophyceae (Chlorophyta) deduced from molecular analysis of the rbcL gene. Phycol Res 43:203–213
Denboh T, Ichimura T, Hendrayanti D et al (2003) The Closterium moniliferum-ehrenbergii (Charophyceae, Chlorophyta) species complex viewed from the 1506 group I intron and ITS2 of nuclear rDNA. J Phycol 39:960–977
Donat A (1926) Zur Kenntnis der Desmidiaceen des norddeutschen Flachlandes. Pflanzenforschung 5. Fischer, Jena
Fawley MW, Dean ML, Dimmer SK et al (2005) Evaluating the morphospecies concept in the Selenastraceae (Chlorophyceae, Chlorophyta). J Phycol 42:142–154
Fenchel T, Esteban GF, Finlay BJ (1997) Local versus global diversity of microorganisms: cryptic diversity of ciliated protozoa. Oikos 80:220–225
Fenchel T, Finlay BJ (2004) The ubiquity of small species: patterns of local and global diversity. BioScience 54:777–784
Finlay BJ (2002) Global dispersal of free-living microbial eukaryote species. Science 296:1061–1063
Finlay BJ, Fenchel T (2002) Microbial eukaryote species. Science 297:337
Foissner W (2006) Biogeography and dispersal of micro-organisms: a review emphasizing protists. Acta Protozoologica 45:111–136
Futuyma DJ (1998) Evolutionary biology. Sinauer Assoc, Massachusetts
Geissler U, Kies L (2003) Artendiversität und Veränderungen in der Algenflora zweier städtischer Ballungsgebiete Deutschlands: Berlin und Hamburg. Nova Hedwigia, Beih 126:1–777
Gerrath JF (1993) The biology of desmids: a decade of progress. In: Round FE, Chapman DJ (eds) Progress in phycological research 9. Biopress, Bristol, pp 79–192
Hegewald E (1999) Polymorphismus und Variabilität in der Grünalgengattung Scenedesmus Meyen, 1829 (Chlorophyta, Chlorococcales). Cour Forsch-Inst Senckenberg 215:123–128
Hegewald E (2000) New combinations in the genus Desmodesmus (Chlorophyceae, Scenedesmaceae). Algol Stud 96:1–18
Hegewald E, Hindák F, Schnepf E (1990) Sudies on the genus Scenedesmus Meyen (Chlorophyceae, Chlorococcales) from South India, with special reference to the cell wall ultrastructure. Beih Nova Hedwigia 99:1–75
Hegewald E, Schnepf E, Jeon SL (1999) Report on Makionella tosaensis Okada (Chlorophyta, Oocystaceae). Algae 14:87–90
Hegewald E, Silva P (1988) Annotated catalogue of Scenedesmus and nomenclaturally related genera including original desriptions and figures. Biblthca phycol 80:1–587
Heimans J (1969) Ecological, phytogeographical and taxonomic problems with desmids. Vegetatio 17:50–82
Hepperle D, Krienitz L (2001) Systematics and ecology of chlorophyte picoplankton in German inland waters along a nutrient gradient. Internat Rev Hydrobiol 86:269–284
Hillebrandt H, Watermann F, Karez R et al (2001) Differences in species richness patterns between unicellular and multicellular organisms. Oecologia 126:114–124
Hindák F (1984) Studies on the chlorococcal algae (Chlorophyceae). III. VEDA, Slovak Acad Science, Bratislava
Hinode T (1966) Desmids from the northern district of Tokushima Prefecture (3). J Jap Bot 41:305–316
Ichimura T (1981) Mating types and reproductive isolation in Closterium ehrenbergii Meneghini. Bot Mag Tokyo 94:325–334
Ichimura T, Kasai F (1990) Mating systems and speciation in haplontic unicellular algae, desmids. In: Kawano S (eds) Biological approaches and evolutionary trends in plants. Academic Press, London, pp 309–332
Ichimura T, Kasai F, Coesel PFM (1997) Geographical and ecological distribution of highly polyploid populations of the Closterium ehrenbergii species complex (Chlorophyta). Phycologia 36:157–163
Jao CC (1940) Studies on the freshwater algae of China. IV. Subaerial and aquatic algae from Nanyoh, Hunan. II. Sinensia 11:241–361
Jeon SL, Hegewald E (2006) A revision of the species Desmodesmus perforatus and D. tropicus (Scenedesmaceae, Chlorophyceae, Chlorophyta). Phycologia 45:567–584
John DM, Maggs CA (1997) Species problems in eukaryotic algae: a modern perspective. In: Claridge MF, Dawah HA, Wilson MR (eds) Species: the units of biodiversity. Chapman and Hall, New York, pp 83–107
Komárek J (1983) Contribution to the chlorococcal algae of Cuba. Nova Hedwigia 37:65–180
Komárek J, Fott B (1983) Chlorophyceae (Grünalgen) Ordnung: Chlorococcales. In: Huber-Pestalozzi G (ed) Das Phytoplankton des Süßwassers 7. Teil, 1. Hälfte. Schweizerbart, Stuttgart
Komárek J, Jankovská V (2001) Review of the green algal genus Pediastrum; implication for pollenanalytical research. Biblthca phycol 108:1–127
Komárek J, Marvan P (1992) Morphological differences in natural populations of the genus Botryococcus (Chlorophyceae). Arch Protistenk 141:65–100
Komárková-Legnerová J. (1969) The systematics and ontogenesis of the genera Ankistrodesmus Corda and Monoraphidium gen. nov. In: Fott B (ed) Studies in phycology. Academia Praha, pp 262–292
Krieger W (1932) Die Desmidiaceen der Deutschen Limnologischen Sunda-Expedition. Arch Hydrobiol, Suppl 11:129–230
Krieger W (1939) Die Desmidiaceen Europas mit Berücksichtigung der aussereuropäischen Arten. Rabenhorst’s Kryptogamen-Flora 13, 1, 2. Akad Verlagsges, Leipzig
Krienitz L (1998) Amphikrikos variabilis sp. nova (Chlorophyta), a common species of inland waters of Namibia. Algol Stud 91:1–10
Krienitz L, Hegewald E, Hepperle D et al (2003) The systematics of coccoid green algae: 18S rRNA gene sequence data versus morphology. Biologia 58:437–446
Krienitz L, Hegewald E, Hepperle D et al (2004) Phylogenetic relationship of Chlorella and Parachlorella gen. nov. (Chlorophyta, Trebouxiophyceae). Phycologia 43:529–542
Krienitz L, Takeda H, Hepperle D (1999) Ultrastructure, cell wall composition, and phylogenetic position of Pseudodictyosphaerium jurisii (Chlorococcales, Chlorophyta) including a comparison with other picoplanktonic green algae. Phycologia 38:100–107
Krienitz L, Ustinova I, Friedl T et al (2001) Traditional generic concepts versus 18S rRNA gene phylogeny in the green algal family Selenastraceae (Chlorophyceae, Chlorophyta). J Phycol 37:852–865
Lewis LA, McCourt RM (2004) Green algae and the origin of land plants. Am J Bot 91:1535–1556
Ling HU, Tyler PA (1976) Meiosis, polyploidy and taxonomy of the Pleurotaenium mamillatum complex (Desmidiaceae). Br phycol J 11:315–330
Logares RE (2006) Does the global microbiota consist of a few cosmopolitan species? Ecologia Austral 16:85–90
Luo W, Krienitz L, Pflugmacher S et al (2005) Genus and species concept in Chlorella and Micractinium (Chlorophyta, Chlorellaceae): genotype versus phenotypical variability under ecosystem conditions. Verh Int Ver Limnol 29:170–173
Marvan P, Komárek J, Comas A (1984) Weighting and scaling of features in numerical evaluation of coccal green algae (genera of the Selenastraceae). Algol Stud 37:363–399
Mayr E (1942) Systematics and the origin of species. Columbia Univ Press, New York
Okada Y (1949) Makinoella tosaensis, a new genus of the Oocystaceae. Jap J Bot 24:166–168
Peterfi LS (1973) Studies on Romanian Staurastra. I. Variability and taxonomy of Staurastrum spinosum (Brébisson) Ralfs. Nova Hedwigia 24:121–144
Potter D, Lajeunesse TC, Saunders GW et al (1997) Convergent evolution masks extensive biodiversity among marine coccoid picoplankton. Biodiv Conserv 6:99–107
Prescott GW, Bicudo CEM, Vinyard WC (1982) A Synopsis of North American Desmids. II, 4. Univ Nebraska Press, Lincoln, London
Rich F (1932) Contributions to our knowledge of the freshwater algae of Africa. Trans Roy Soc South Africa 20:149–188
Scott AM, Prescott GW (1961) Indonesian desmids. Hydrobiologia 17:1–13
Senousy HH, Beakes GW, Hack E. (2004) Phylogenetic placement of Botryococcus braunii (Trebouxiophaceae) and Botryococcus sudeticus isolate UTEX 2629 (Chlorophyceae). J Phycol 40:412–423
Slapeta J, López-García P, Moreira D (2006) Global dispersal and ancient cryptic species in the smallest marine eukaryotes. Mol Biol Evol 23:23–29
Thérézien Y, Couté A (1977) Algues d’eau douce des Iles Kerguelen et Crozet (à l’exclusion des Diatomées). CNFRA 43:1–91
Thomasson K (1960) Notes on the plankton of Lake Bangweulu. 2 Nova Acta Regiae Soc Sci Upsal 17:1–43
Trainor FR (1998) Biological aspects of Scenedesmus (Chlorophyceae) phenotypic plasticity. Nova Hedwigia, Beih 117:1–367
Turner WB (1892) The freshwater algae (principally Desmideae) of East India. Kongl Svensk Vetensk Acad Handl 25:1–187
Vanormelingen P, Hegewald E, Braband A et al (2007) The systematics of a small spineless Desmodesmus taxon, D. costato-granulatus (Sphaeropleales, Chlorophyceae), based on ITS2 rDNA sequence analyses and cell wall morphology. J Phycol 43:378–396
Wallich GC (1860) Desmidiaceae of Lower Bengal. Ann Mag Nat Hist 3:184–197, 273–285
Watanabe M, Ichimura T (1978) Biosystematic studies of the Closterium peracerosum-strigosum-littorale complex. II. Reproductive isolation and morphological variation among several populations from the northern Kanto area in Japan. Bot Mag Tokyo 91:1–10
Wille N (1924) Süsswasseralgen von der deutschen Südpolar-Expedition auf dem Schiff “Gauss”. 1.–2. Teil. Deutsche Südpolar-Expedition 8:373–445
Wolle F (1884) Desmids of the United States and list of American pediastrums. Moravian Publ Off, Bethlehem, P.A
Author information
Authors and Affiliations
Corresponding author
Additional information
Special Issue: Protist diversity and geographic distribution. Guest editor: W. Foissner.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License ( https://creativecommons.org/licenses/by-nc/2.0 ), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Coesel, P.F.M., Krienitz, L. Diversity and geographic distribution of desmids and other coccoid green algae. Biodivers Conserv 17, 381–392 (2008). https://doi.org/10.1007/s10531-007-9256-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-007-9256-5