
Abstract
The emergence and persistence of polymorphism within populations generally requires specific regimes of natural or sexual selection. Here, we develop a unified theoretical framework to explore how polymorphism at targeted loci can be generated and maintained by either disassortative mating choice or balancing selection due to, for example, heterozygote advantage. To this aim, we model the dynamics of alleles at a single locus A in a population of haploid individuals, where reproductive success depends on the combination of alleles carried by the parents at locus A. Our theoretical study of the model confirms that the conditions for the persistence of a given level of allelic polymorphism depend on the relative reproductive advantages among pairs of individuals. Interestingly, equilibria with unbalanced allelic frequencies were shown to emerge from successive introduction of mutants. We then investigate the role of the function linking allelic divergence to reproductive advantage on the evolutionary fate of alleles within the population. Our results highlight the significance of the shape of this function for both the number of alleles maintained and their level of genetic divergence. Large number of alleles are maintained with substantial replacement of alleles, when disassortative advantage slowly increases with allelic differentiation . In contrast, few highly differentiated alleles are predicted to be maintained when genetic differentiation has a strong effect on disassortative advantage. These opposite effects predicted by our model explain how disassortative mate choice may lead to various levels of allelic differentiation and polymorphism, and shed light on the effect of mate preferences on the persistence of balanced and unbalanced polymorphism in natural population.







Similar content being viewed by others

References
Arora J, Pierini F, McLaren PJ, Carrington M, Fellay J, Lenz TL (2020) HLA heterozygote advantage against HIV-1 is driven by quantitative and qualitative differences in HLA allele-specific peptide presentation. Mol Biol Evol 37(3):639–650
Billiard S, Castric V, Vekemans X (2007) A general model to explore complex dominance patterns in plant sporophytic self-incompatibility systems. Genetics 175(3):1351–1369
Billiard S, López-Villavicencio M, Devier B, Hood ME, Fairhead C, Giraud T (2011) Having sex, yes, but with whom? Inferences from fungi on the evolution of anisogamy and mating types. Biol Rev 86(2):421–442
Bloomfield G, Skelton J, Ivens A, Tanaka Y, Kay RR (2010) Sex determination in the social amoeba \(Dictyostelium discoideum\). Science 330(6010):1533–1536
Busch JW, Witthuhn T, Joseph M (2014) Fewer S-alleles are maintained in plant populations with sporophytic as opposed to gametophytic self-incompatibility. Plant Species Biol 29(1):34–46
Castric V, Bechsgaard J, Schierup MH, Vekemans X (2008) Repeated adaptive introgression at a gene under multiallelic balancing selection. PLoS Genet 4(8):e1000168
Castric V, Bechsgaard JS, Grenier S, Noureddine R, Schierup MH, Vekemans X (2010) Molecular evolution within and between self-incompatibility specificities. Mol Biol Evol 27(1):11–20
Chouteau M, Llaurens V, Piron-Prunier F, Joron M (2017) Polymorphism at a mimicry supergene maintained by opposing frequency-dependent selection pressures. Proc Natl Acad Sci 114(31):8325–8329
Coron C, Costa M, Leman H, Smadi C (2018) A stochastic model for speciation by mating preferences. J Math Biol 76(6):1421–1463
Durand E, Chantreau M, Le Veve A, Stetsenko R, Dubin M, Genete M, Llaurens V, Poux C, Roux C, Billiard S et al (2020) Evolution of self-incompatibility in the Brassicaceae: lessons from a textbook example of natural selection. Evolut Appl 13(6):1279–1297
Franklin-Tong VE (2008) Self-incompatibility in flowering plants. Evol Divers Mech 305. https://doi.org/10.1007/978-3-540-68486-2
Gervais CE, Castric V, Ressayre A, Billiard S (2011) Origin and diversification dynamics of self-incompatibility haplotypes. Genetics 188(3):625–636
Jay P, Chouteau M, Whibley A, Bastide H, Parrinello H, Llaurens V, Joron M (2021) Mutation load at a mimicry supergene sheds new light on the evolution of inversion polymorphisms. Nat Genet 53(3):288–293
Jiang Y, Bolnick DI, Kirkpatrick M (2013) Assortative mating in animals. Am Nat 181(6):E125–E138
Kimura M, Crow JF (1964) The number of alleles that can be maintained in a finite population. Genetics 49(4):725
Kimura M, Ohta T (2020) Chapter eight. Breeding structure of populations. Princeton University Press, Princeton, pp 117–140
Kronstad JW, Staben C (1997) Mating type in filamentous fungi. Annu Rev Genet 31(1):245–276
Krumbeck Y, Constable GWA, Rogers T (2020) Fitness differences suppress the number of mating types in evolving isogamous species. R Soc Open Sci 7(2):192126
LaSalle J (1960) Some extensions of Liapunov’s second method. IRE Trans Circuit Theory 7(4):520–527
Leducq JB, Llaurens V, Castric V, Saumitou-Laprade P, Hardy OJ, Vekemans X (2011) Effect of balancing selection on spatial genetic structure within populations: theoretical investigations on the self-incompatibility locus and empirical studies in Arabidopsis halleri. Heredity 106(2):319–329
Lenz TL (2011) Computational prediction of MHC II-antigen binding supports divergent allele advantage and explains trans-species polymorphism. Evol Int J Org Evol 65(8):2380–2390
Lewontin RC, Ginzburg LR, Tuljapurkar SD (1978) Heterosis as an explanation for large amounts of genic polymorphism. Genetics 88(1):149–169
Maisonneuve L, Beneteau T, Joron M, Smadi C, Llaurens V (2021) When do opposites attract? A model uncovering the evolution of disassortative mating. Am Nat 198(5):625–641
Metz JAJ, Geritz SAH, Meszéna G, Jacobs FJA, Van Heerwaarden JS (1996) Adaptive dynamics: a geometrical study of the consequences of nearly faithfull reproduction. In: van Strien SJ, Verduyn-Lunel SM (eds) Stochastic and spatial structures of dynamical systems. Elsevier, North Holland, pp 183–231
Meyer CD (2000) Matrix analysis and applied linear algebra, vol 71. SIAM, Philadelphia
Nieuwenhuis BPS, Billiard S, Vuilleumier S, Petit E, Hood ME, Giraud T (2013) Evolution of uni-and bifactorial sexual compatibility systems in fungi. Heredity 111(6):445–455
Phadke SS, Zufall RA (2009) Rapid diversification of mating systems in ciliates. Biol J Linn Soc 98(1):187–197
Phillips KP, Cable J, Mohammed RS, Herdegen-Radwan M, Raubic J, Przesmycka KJ, Van Oosterhout C, Radwan J (2018) Immunogenetic novelty confers a selective advantage in host-pathogen coevolution. Proc Natl Acad Sci 115(7):1552–1557
Potts WK, Manning CJ, Wakeland EK (1991) Mating patterns in seminatural populations of mice influenced by MHC genotype. Nature 352(6336):619–621
Schierup MH, Vekemans X, Charlesworth D (2000) The effect of subdivision on variation at multi-allelic loci under balancing selection. Genet Res 76(1):51–62
Smadi C, Leman H, Llaurens V (2018) Looking for the right mate in diploid species: how does genetic dominance affect the spatial differentiation of a sexual trait? J Theor Biol 447:154–170
Stefan T, Matthews L, Prada JM, Mair C, Reeve R, Stear MJ (2019) Divergent allele advantage provides a quantitative model for maintaining alleles with a wide range of intrinsic merits. Genetics 212(2):553–564
Sylvester JJ (1852) XIX. A demonstration of the theorem that every homogeneous quadratic polynomial is reducible by real orthogonal substitutions to the form of a sum of positive and negative squares. Lond Edinb Dublin Philos Mag J Sci 4(23):138–142
Tuttle EM, Bergland AO, Korody ML, Brewer MS, Newhouse DJ, Minx P, Stager M, Betuel A, Cheviron ZA, Warren WC et al (2016) Divergence and functional degradation of a sex chromosome-like supergene. Curr Biol 26(3):344–350
Tuttle EM (1993) Mate choice and stable polymorphism in the white-throated sparrow. PhD thesis, State University of New York at Albany
Vekemans X, Schierup MH, Christiansen FB (1998) Mate availability and fecundity selection in multi-allelic self-incompatibility systems in plants. Evolution 52(1):19–29
Wedekind C, Seebeck T, Bettens F, Paepke AJ (1995) MHC-dependent mate preferences in humans. Proc R Soc Lond Ser B Biol Sci 260(1359):245–249
Winternitz J, Abbate JL, Huchard E, Havlíček J, Garamszegi LZ (2017) Patterns of MHC-dependent mate selection in humans and nonhuman primates: a meta-analysis. Mol Ecol 26(2):668–688
Wright S (1939) The distribution of self-sterility alleles in populations. Genetics 24(4):538
Acknowledgements
This work has been supported by the Chair ”Modélisation Mathématique et Biodiversité” of Veolia Environnement-École Polytechnique-Muséum national d’Histoire naturelle-Fondation X. This work has been partially supported by the LabEx PERSYVAL-Lab (ANR-11-LABX-0025-01) funded by the French program Investissements d’avenir and by ANR project DEEV ANR- 20-CE40-0011-01.
We warmly thank Clément, Gabriel, Lucie and Agathe for enlarging our world during this work.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendices
Birth-and-death model and convergence to dynamical system
The dynamical system (1) governing the population dynamics can be obtained as a large population limit of a stochastic individual based model (similarly as in Coron et al. (2018), Smadi et al. (2018) for instance).
More precisely, let K be a large parameter that gives the order size of the population. The microscopic population is represented by the process \((Z^K_{i}(t))_{i\in \{1,\ldots ,k\}}\) where \(Z^K_{i}(t)\) gives the size, divided by K, of the population of type i individuals at time t. Each individual of type i reproduces with rate \(\beta _i\), that is, after a time distributed as an exponential random variable of parameter \(\beta _i\). As explained in the main text, the second parent is chosen uniformly at random among all individuals alive in the population. If the second parent is of type j, with probability \(p_{ij}\) this mating event leads to the birth of a new individual, being of type i with probability one half, and of type j with probability one half when there is no mutation (otherwise the mutation is applied to the genotype i or j with the same probability). All the indivivuals have the same death rate, which is given by
at time t. Under the assumption that the sequence of initial conditions \(\big ((Z^K_{i}(0))_{i\in \{1,\ldots ,k\}}\big )_{K\in \mathbb {N}}\) converges (in probability) when K goes to infinity, the sequence of stochastic functions \(\big ((Z^K_{i}(t))_{,i\in \{1,\ldots ,k\}}, t\in [0,T]\big )_{K\in \mathbb {N}}\) also converges (in probability for the uniform convergence) to the trajectory of the deterministic system (1).
Proof of the main results
This section gathers the theoretical results on the behavior of the solutions to system (2):
Recall the matrix \(M=(s_{ij})_{1\le i, j, \le k}\)
1.1 Existence of positive equilibria
Our first result states conditions for the existence of a positive equilibrium, meaning that each coordinate of the vector is strictly positive.
Proposition B.1
If \(\det (M)\ne 0\), the system admits a positive equilibrium if and only if \(M^{-1}\textbf{1}>0\). In this case, the equilibrium is unique and is given by
If \(\det (M)=0\), the system may have no equilibrium or an infinite number of equilibria.
Proof of Proposition B.1
\(\underline{\textbf{Case}\,\det (M)\ne 0:}\) Firstly, we assume that there exists a positive equilibrium \(Z^*=(z^*_i)_{i=1,..,k}\). From (2), the following equality holds for every \(i\in \{1,..,k\}\),
with \(z^*=\sum _{i=1}^k z^*_{i}\). Note that this equation implies that \(cz^*>b-d\), since \(s_{ij}>0\). Using a matrix formulation, we have the following linear system:
As \(\det (M)\ne 0\), the matrix M has an inverse and
Since \(\frac{{z^*}(cz^*-b+d)}{b} >0\), we thus deduce that \(M^{-1} \textbf{1}>0\).
Moreover by summing all coordinates, we find
and finally, the equilibrium is unique and defined by (6).
On the other hand, if we assume that \(M^{-1}\textbf{1}>0\), then we can define \(Z^*\) by (6) and, using the above computation, it is straightforward to verify that it is a positive equilibrium of (2). This ends the proof for the case \(\det (M)\ne 0\).
\(\underline{{\textbf {Case}}\,\det (M)=0:}\)
Let us verify whether there exists a vector X with positive coordinates such that \(MX=\textbf{1}\).
Indeed, if such a vector X does not exist, then (12) can never be satisfied and there is no equilibrium of (2).
On the contrary, if we can find such a vector X, we set
Then it is straightforward to verify that \(MZ^*=z^*\frac{cz^*+d-b}{b}\textbf{1}\), with \(z^*=\textbf{1}^TZ^*\). Hence, \(Z^*\) satisfies (12) and is a positive equilibrium of (2). However, this equilibrium is not unique. Indeed, \(\textbf{1}\) belongs to the image of M, and since M is a symmetric matrix, any vector Y in the kernel of M is orthogonal to \(\textbf{1}\), i.e. \(\textbf{1}^TY=0\). Then, for all vectors \(Y\in Ker(M)\), \(M(Z^*+Y)=z^*\frac{cz^*+d-b}{b}\textbf{1}\), with \(z^*= \textbf{1}^T Z^*=\textbf{1}^T(Z^*+Y)\), and \(Z^*+Y\) is also an equilibrium of (2).
We now construct examples for the two possibilities. Assume that M can be written as
\(\det (A)\ne 0\) and such that \(A^{-1}\textbf{1}>0\). Since A is a symmetric matrix, there exists an orthonormal basis \((V_i)_{i=1,..,k-1}\) of \(\mathbb {R}^{k-1}\) where \(V_i\) is an eigenvector of A associated to its non-null eigenvalue \(\lambda _i\) for all \(i\in \{1,..,k-1\}\). Let us decompose C in this basis \(C=\sum _{i=1}^{k-1}\alpha _i V_i\).
Our aim is to construct C, and thus to select \((\alpha _i)_{i=1,..,k-1}\), to find the examples. First, we have to select \((\alpha _i)_{i=1,..,k-1}\) such that \(C=\sum _{i=1}^{k-1}\alpha _iV_i>0\). Then using the Schur complement, we have that \(\det (M)=-(C^TA^{-1}C)\times \det (A)\) and thus \(\det (M)=0\) if and only if
This gives us a way to construct a matrix M with \(\det (M)=0\).
Finally, to ensure that there exists \(Y>0\) with \(MY=\textbf{1}\), let us consider vectors depending on a parameter \(\eta \in \mathbb {R}\) such that
Let us study the image by M of this vector
Thus if \(C^T A^{-1} \textbf{1}=\sum _{i=1}^{k-1}\frac{\alpha _i\omega _i}{\lambda _i}=1\) where \(\textbf{1}=\sum _{i=1}^{k-1}\omega _i V_i\), then we can find \(\eta >0\) sufficiently small, such that \(MY(\eta )=\textbf{1}\) with \(Y(\eta )>0\).
With this in mind, we can explicit an example for each possibility. Let
M and X have non-negative coordinates, \(\det (M)=0\) and \(MX=\textbf{1}\). Thus, in this example, the associated system of equations (2) has an infinite number of positive stationary states.
Now, let \(a>0\) and
In this example, \(\det (M)=0\), \(Ker(M)=Vect(X_0)=Im(M)^T\), however \(\textbf{1}^TX_0\ne 0\) hence \(\textbf{1}\) does not belong to the image of M. In other words, the associated system of equations (2) has no positive stationary state. \(\square \)
1.2 Convergence toward the positive equilibria
We now aim at describing the dynamics of the population described by system (2). The following proposition gives conditions for a positive equilibrium to be stable. Moreover, it states that conditions for local stability and global stability are identical, i.e. a locally stable equilibrium attracts all trajectories starting with positive initial conditions.
Proposition B.2
Assume that \(\det (M)\ne 0\) and \(M^{-1}\textbf{1}>0\), and denote the unique positive equilibrium of (2) by \(Z^*\), whose expression is given in (6).
-
i)
\(Z^*\) is locally stable if and only if M has 1 positive eigenvalue and \(k-1\) negative eigenvalues.
-
ii)
\(Z^*\) is locally stable if and only if it is globally stable on \((\mathbb {R}^*_+)^k\).
Our proof relies on a study of the Jacobian matrix of the system computed at the positive equilibria and on the construction of a Lyapunov function. Actually, we prove that the criterion on the eigenvalues of M is equivalent to the fact that the Jacobian matrix of the system admits only negative eigenvalues.
The second point of the proposition will be proved using the following function
which is a Lyapunov function for the dynamical system (2).
Proof
\(\underline{\mathrm{Proof\,of}\,i):}\) Let us first give the Jacobian matrix of the system (2) around equilibrium \(Z^*\). For all \(i\in \{1,..,k\}\), let
Recall that \(F_i(z^*)/z^*_i=0\). Then by differentiating the function \(F_i\) w.r.t \(z_i\) and \(z_j\) for \(j\ne i\), we find
Hence the jacobian Matrix of the system can be written
Then since for any \((A,B) \in \mathcal {M}_k(\mathbb {R})\), \(\det (AB)=\det (BA)\), we have
In other words, the eigenvalues of \(J^*\) are the same as the ones of
which is symmetric and has real eigenvalues. According to Proposition 2.1,
Hence, we can write the matrix B as
Let us prove that B has negative eigenvalues if M has only 1 positive eigenvalue. To this aim, notice that as M and DMD are congruent, according to Sylvester’s law of inertia (Sylvester 1852), they have the same numbers of positive/negative eigenvalues.
Let us first assume that M as well as DMD have only 1 positive eigenvalue. As DMD is symmetric, there exists an orthonormal basis \((V_i)_{i=1,..,k}\) of \(\mathbb {R}^k\) formed with eigenvectors of DMD, associated to its eigenvalues \((\lambda _i)_{i=1,..,k}\). In the following we denote by \(\lambda _1>0\) the positive eigenvalue. Using the definition of D in (15), we obtain that
Since by assumption \(\frac{1}{\textbf{1}^TM^{-1}\textbf{1}} >0\), we deduce that it is the unique positive eigenvalue of DMD and that \(D\textbf{1}\) is a positive eigenvector associated to the positive eigenvalue of DMD.
From (17), we have
Now, let us consider any \(Y\in \mathbb {R}^k\) and its decomposition in the eigenvectors’ basis: there exist \((\gamma _1,..,\gamma _k)\in \mathbb {R}^k\) such that \( Y=\sum _{i=1}^k\gamma _i V_i, \) and
since the eigenvalues \(\lambda _2,.., \lambda _k\) of DMD are negative by assumption. In other words, B is a negative definite matrix. Recalling that B and the Jacobian matrix \(J^*\) have the same eigenvalues, we deduce that the equilibrium is locally stable.
Conversely, if M has more than 1 positive eigenvalue, it is also the case of DMD. Then, assuming for example that \(\lambda _2>0\), we have, by an application of (18) with \(Y=V_2\),
Hence B has at least 1 positive eigenvalue. The stationary state is unstable.
\(\underline{\mathrm{Proof\,of}\,ii): }\) We are now ready to prove the second point of the proposition. We only have to prove the direct implication, as the other one is obvious. To this aim, let us assume that the equilibrium is locally stable and thus all eigenvalues of \(J^*\) are negative. Recall the definition of the function V in (14). By differentiating, we find
since \(\sum _\ell z_\ell /z=\sum _{\ell }z^*_\ell /z^*=1\). In addition with the fact that \(\sum _{j=1}^k s_{\ell j}\frac{z_{j}^*}{z^*}\) is a constant, we find
with
Now, we recall that the stationary state is assumed to be locally stable, i.e. all eigenvalues of \(J^*\) (or B) are negative. Since B and \((b-d-2cz^*)I+bM\) are congruent, these matrices are both negative definite. Furthermore, for all \(X\in \mathbb {R}^k\setminus \{0\}\), \(X^T((b-d-2cz^*)I+bM)X<0\). Since \(\textbf{1}^TY=0\), we find
as soon as \(Y\ne 0\), and \(\frac{d}{dt} V(z(t))=0\) if and only if \(Y=0\), i.e. \({z_i}/{z}={z_i^*}/{z^*}\) for all \(1 \le i \le k\). Moreover, we can prove that the set \(\{z\in \mathbb {R}^k, Y=0\}\) is a positive invariant set for the dynamical system (2). Indeed, for all \(i\in \{1,..,k\}\),
where we used (12). Theorem 1 of LaSalle (1960) then ensures that any bounded positive solution to (2) converges toward this invariant set. Finally, we prove that the solutions of (2) remain bounded by studying the dynamics of the total population size z, which satisfies
We easily obtain that
and thus classical results on the logistic equation ensure that the total population size remains bounded through time.
Lasalle’s Theorem then ensures that \(\frac{z_i(t)}{z(t)}\) converges to \(\frac{z_i^*}{z^*}\). To conclude the proof it remains to prove that z(t) converges to \(z^*\). Let us write \(\frac{z_i(t)}{z(t)}=\frac{z_i^*}{z^*}+\epsilon _i(t)\) and according to (12)
with
For any \(\varepsilon >0\), as soon as t is large enough, we deduce that
Using classical results on logistic equations, we deduce that z(t) converges to \(z^*\). This ends the proof of Proposition 2.1. \(\square \)
1.3 Complete study of the three allele case
Consider three alleles \(A_1\), \(A_2\), and \(A_3\) whose selection matrix is given by
Since the selective advantage parameters are positive, \(\det (M)>0\) and one obtains with a simple computation that the condition for existence of a three-allele positive equilibrium \(M^{-1}\textbf{1}>0\) can be written as (8). Moreover, the three-alleles equilibrium equals
where
In this case, the behavior of the dynamical system (2) can be fully characterized.
Proposition B.3
Let us consider a system of three alleles, whose interactions are characterized by matrix (19).
-
Assume (8), then the co-existence equilibrium given above is globally stable on \((\mathbb {R}^*_+)^3\).
-
otherwise, assume for example that \(s_{23}\ge s_{12}+s_{13}\), then the two-alleles equilibrium
$$\begin{aligned} \left( 0,\frac{b-d+b{s_{23}}/{2}}{2c},\frac{b-d+b{s_{23}}/{2}}{2c}\right) , \end{aligned}$$is globally stable on \((\mathbb {R}^*_+)^3\).
Proof
The first point easily derives from Proposition 2.1.
For the second point, we exhibit a Lyapunov function, which is similar to the one of the previous proof. Let us consider
Then, by a direct computation, we find
Recall that \(s_{23}\ge s_{12}+s_{13}\), and without loss of generality, we may assume that \(s_{12} \le s_{13}\) (otherwise, exchange the roles of \(z_2\) and \(z_3\) in the computations). Then
Therefore V is a Lyapunov function and \(\dot{V}({\textbf {z}})=0\) if and only if \(z_1=0\) and \(z_2=z_3\). We then conclude with Lasalle Theorem (Theorem 1 of LaSalle (1960)) and an argument similar to the one at the end of the previous proof. \(\square \)
1.4 Successive introductions of types
Assume that k types coexist. In view of previous sections, if \(\det (M)\ne 0\), there is coexistence of the k types if and only if \(M^{-1}\textbf{1}>0\) and the second eigenvalue of M is negative (or equivalently the eigenvalues of \(J^*\) are all negative).
If a mutant characterized by parameters \(S=(s_{k+1,i})_{i=1,..,k}\) and \(\sigma =s_{k+1,k+1}\) appears, this mutant may invade if the Jacobian matrix of (2) around equilibrium \((z^*,0)\) is unstable. This Jacobian matrix can be written
As previously written, the matrix \(J^*\) is the Jacobian matrix of the resident system of size k at equilibrium \(z^*\) and its eigenvalues are all negative, thus the equilibrium \((z^*,0)\) is unstable if the last eigenvalue is positive, i.e.
which gives (9).
In what follows, we will be interested in proving the following lemma which states that if the mutant may invade, the new community will be composed of \(k+1\) types if and only if the coexistence state with \(k+1\) types exists.
Proposition B.4
Let us consider a stable resident population with k types given by a matrix M, i.e M satisfies \(M^{-1}\textbf{1}>0\) and its second eigenvalue is negative.
Let us consider a mutant type arising in this resident population characterized by S and \(\sigma \). Denote the new fitness matrix by
If \(\bar{M}^{-1}\textbf{1}>0\), i.e. the equilibrium with \(k+1\) species exists, then it is globally asymptotically stable.
Proof
Let us first give conditions under which the second eigenvalue of \(\bar{M}\) is positive. As \(\bar{M}\) is a symmetric matrix and M a principal sub-matrix of \(\bar{M}\), Proposition D.1 implies that
where \((\lambda _i^M)_{i=1,\ldots ,k}\) denote the eigenvalues of M and \((\lambda _i^{\bar{M}})_{i=1,\ldots ,k+1}\) the ones of \(\bar{M}\). Since \(\lambda _2^M<0\), \(\bar{M}\) can have at most 2 positive eigenvalues. On the other hand, using Proposition D.2, we compute
Since \(\det (M)\) has the sign of \((-1)^{k-1}\), we deduce the following equivalences:
Let us now prove that this last condition holds as soon as \(\bar{M}^{-1}\textbf{1}>0\). For all \(i\in \{1,..,k+1\}\) we denote by \(M_i\) the matrix resulting from the suppresion of the i\(^{th}\) column and the i\(^{th}\) line of \(\bar{M}\), by \(S_i\) the i\(^{th}\) column of \(\bar{M}\) without the i\(^{th}\) coefficient, and by \(\sigma _i\) the coefficient \(\bar{M}_{ii}\). Notice that \(M_{k+1}=M\), \(S_{k+1}=S\) and \(\sigma _{k+1}=\sigma \). Using these notations and Proposition D.2, we notice that for all i
Let us now denote \(\bar{M}^{-1}\textbf{1}\) by \((u_i)_{i=1,..,k+1}\). Then using Cramer’s rule along with (23), we have for all \(i\in \{1,..,k+1\}\)
Hence, we deduce that if \(\bar{M}^{-1}\textbf{1}>0\), then \(u_{k+1}>0\), and since \(1-S^TM^{-1}\textbf{1}<0\), then \(\sigma -S^TM^{-1}S<0\). Therefore according to (22) and Proposition 2.1, the equilibrium with \(k+1\) species is globally stable. This ends the proof. \(\square \)
Now, let us study the case where the equilibrium with \(k+1\) species does not exist, i.e. there exists \(i\in \{1,..,k+1\}\) such that \(u_i<0\). We are not capable of deciphering which alleles will go extinct due to mutant invasion. Numerical simulations reveal that alleles \(1\le i\le k\) that verify \(u_i u_{k+1}>0\) might go extinct in the equilibrium. Notice that in some cases, all these alleles disappear, while in other cases, only a subset does.
Proofs concerning the genetic model
This section gathers theoretical results associated with the specific model in Sect. 3.1.
1.1 Globally stable equilibrium in a particular case
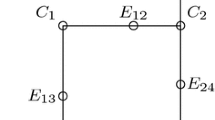
In this section we focus on the case where \(L=3\), such that there are 8 possible alleles (see Fig. 6 for a representation).
We first prove the convergence of the population state in a particular case for which there is no positive stable equilibrium. This example will be important for the proof of Proposition 3.1 below.
Assume that 4 alleles are present in the population:
up to permutation of the sites. Then, the selection matrix M equals
Our aim is to prove that the equilibrium
is globally positively stable in \((\mathbb {R}^+)^4\), i.e. all trajectories starting from \((\mathbb {R}^+)^4\) converge to this equilibrium.
To this aim, let us compute the function V defined in Eq. (14) with \(z^*\) defined by (25), i.e. \(V(z)=1-\frac{1}{2}\ln \left( \frac{z_3}{z}\right) -\frac{1}{2}\ln \left( \frac{z_4}{z}\right) \). We get
Finally, note that if \(\alpha \le 2\) then \(\alpha \le \frac{\ln (4)}{\ln (4/3)}\), and then the two dimensional function \((x,y)\mapsto x^2+3^{\alpha }y^2+2^{\alpha }xy\) is positive on \(\mathbb {R}^2\). So V is a Lyapunov function and its derivative is null only if \(z_1=z_2=0\) and \(z_3=z_4\). We then conclude by using an argument similar to the one of the end of proof of Proposition B.2. This finally proves the convergence of all positive trajectories to Equilibrium (25).
1.2 Proof of Proposition 3.1
Proposition 3.1 arises from two facts: we are able to exhibit stable sets of alleles (see Fig. 6) and we can characterize the impact of mutations on the number of coexisting alleles in the population.
In fact, we obtain a stochastic process which jumps across stable states of the population depending on the arrival of mutant alleles. This process is characterized by a transition matrix which explicits the possible transitions and is represented on Fig. 8.

Transition matrix for the allelic composition of the population. The red color corresponds to stable population states. Blue dashed arrows show the initial distribution after the first mutation
Proof
The set of possible alleles is \(\{0,1\}^3\). We first prove that the two following types of genetic compositions are stable: (Stable population 1) Only the two most-differentiated alleles, (Stable population 2) four alleles that are all at distance 2 from each other.
We already know that (stable population 1) is stable from the end of Sect. 3.1. For (stable population 2), we study the conditions for invasion of a mutant in a population composed of four alleles all at distance 2 from each other. The new mutant will necessarily be at distance 1 from 3 of the resident types, and at distance 3 from the forth one. Then following the notations of Eq. (9) in Sect. 2.3, we get that \(S=(1, 1, 1, 3^{\alpha })^T\). Therefore the new mutant can invade only if
In particular for \(\alpha \in [1,2]\), this gives that the mutant cannot invade, hence the population with four equidistant alleles is stable.
Now, we consider the different possible orders of mutations arising in the population to prove the proposition. Let us assume without loss of generality that the population starts with only individuals with allele (0, 0, 0). The second arising allele in the population can be at distance 1 (with probability 3/7), 2 (with probability (3/7), or 3 (with probability 1/7), from (0, 0, 0). In any case, Proposition B.3 implies that this mutant will invade and the population will then exhibit two different alleles at equilibrium.
-
If this first mutant is at distance 3 from the resident allele (0, 0, 0), then the population has reached the configuration (Stable population 1) and no new mutation will be able to invade.
-
If the first mutant is at distance 2 from the resident allele (0, 0, 0), then there are 3 possibilities for the second mutant (each arising with probability 1/3):
-
(i)
The second mutant can be at distance 1 from each of the two pre-existing alleles. Then Eq. (9) shows that it cannot invade, if \(\alpha >1\). The population thus stays with the two resident alleles, at a distance 2 from each other.
-
(ii)
it can be at distances 1 and 3 from the two first alleles, in which case only the mutant and its opposite type will remain in the population (according to Proposition B.3 and since \(3^\alpha >2^\alpha +1\)), which corresponds to a configuration (Stable population 1).
-
(iii)
The second mutant can be at distance 2 from the two pre-existing alleles. From Eq. (8), the population will then converge to an equilibrium with the three first alleles.
If the third mutant is in the fourth position at distance 2 from the three pre-existing alleles then it will also invade (see Sect. 2.3). This mutant arises with probability 1/5 and brings the population to (Stable population 2) (as illustrated in Fig. 6).
If the third mutant is in a position such that two most-differentiated alleles co-exist in the population then only these two alleles will remain (see Sect. C.1 for details). This occurs with probability 3/5.
Lastly, if (with probability 1/5) the third mutant is at distance 1 from the three first types then it cannot invade, according to Eq. (9).
-
(i)
-
Finally, if the first mutant is at distance 1 from (0, 0, 0), then there are two possibilities for the second mutant:
-
(i)
The second mutant is at distance 2 and 3 from the two resident alleles, respectively (with probability 1/3). Only the two most-differentiated alleles will then remain from Proposition B.3 and this population state is stable.
-
(ii)
The second mutant is at distances 1 and 2 (with probability 2/3) from the pre-existing alleles, respectively. Thus, one of the two pre-existing alleles will disappear and the population will exhibit only two alleles, at distance 2 from each other, as already proved by Proposition B.3.
-
(i)
Finally, the transition graph of the genetic composition of the population is given in Fig. 8. This gives that the probability that the population ends up with 4 equidistant genotypes is equal to
\(\square \)
Complements of linear algebra
Proposition D.1
(Eigenvalue Interlacing Theorem). Let \(A\in \mathbb {R}^{n\times n}\) be a symmetric matrix and B be a principal sub-matrix of size \(m<n\). If \(\lambda ^A_1\ge \lambda ^A_2 \ge \cdots \ge \lambda ^A_n\) the eigenvalues of A and \(\lambda ^B_1\ge \cdots \ge \lambda ^B_m\) the eigenvalues of B. Then for any \(1\le k\le m\)
In particular if \(m=n-1\) then
Proposition D.2
(Schur complement). Let M be a matrix defined by blocs as \(M=\begin{pmatrix} A&{}B\\ C&{}D\end{pmatrix}\) If D is invertible then the Schur complement of D is defined by \(A-BD^{-1}C\). Moreover
For proof of these two results, see Example 7.5.3 and Sect. 6.2 in Meyer (2000).
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Coron, C., Costa, M., Leman, H. et al. Origin and persistence of polymorphism in loci targeted by disassortative preference: a general model. J. Math. Biol. 86, 4 (2023). https://doi.org/10.1007/s00285-022-01832-1
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00285-022-01832-1