
Abstract
Recent simulation modeling has shown that species can coevolve toward clusters of coexisting consumers exploiting the same limiting resource or resources, with nearly identical ratios of coefficients related to growth and mortality. This paper provides a mathematical basis for such as situation; a full analysis of the global dynamics of a new model for such a class of n-dimensional consumer–resource system, in which a set of consumers with identical growth to mortality ratios compete for the same resource and in which each consumer is mutualistic with the resource. First, we study the system of one resource and two consumers. By theoretical analysis, we demonstrate the expected result that competitive exclusion of one of the consumers can occur when the growth to mortality ratios differ. However, when these ratios are identical, the outcomes are complex. Either equilibrium coexistence or mutual extinction can occur, depending on initial conditions. When there is coexistence, interaction outcomes between the consumers can transition between effective mutualism, parasitism, competition, amensalism and neutralism. We generalize to the global dynamics of a system of one resource and multiple consumers. Changes in one factor, either a parameter or initial density, can determine whether all of the consumers either coexist or go to extinction together. New results are presented showing that multiple competing consumers can coexist on a single resource when they have coevolved toward identical growth to mortality ratios. This coexistence can occur because of feedbacks created by all of the consumers providing a mutualistic service to the resource. This is biologically relevant to the persistence of pollination–mutualisms.





Similar content being viewed by others


References
Albrecht M, Padrn B, Bartomeus I, Traveset A (2014) Consequences of plant invasions on compartmentalization and species’ roles in plant–pollinator networks. Proc R Soc Lond B 281:20140773
Bascompte J, Jordano P, Melián CJ, Olesen JM (2003) The nested assembly of plant–animal mutualistic networks. Proc Natl Acad Sci 100:9383–987
Bronstein JL (1994) Conditional outcomes in mutualistic interactions. Trends Ecol Evol 9:214–217
Campbell C, Yang S, Albert R, Shea K (2015) Plant–pollinator community network response to species invasion depends on both invader and community characteristics. Oikos 124:406–413
Cantrell RS, Cosner C, Ruan S (2004) Intraspecific interference and consumer–resource dynamics. Discrete Cont Dyn Syst B 4:527–546
Fagan WF, Bewick S, Cantrell S, Cosner C, Varassin IG, Inouye DW (2014) Phenologically explicitmodelsforstudyingplant–pollinator interactions under climate change. Theor Ecol 7:289–297
Freedman HI, Waltman P (1984) Persistence in models of three interacting predator–prey populations. Math Bios 68:213–231
Gilbert LE (1980) Food web organization and conservation of neotropical diversity. In: Soule Michael, Wilcox Bruce A (eds) Conservation biology. An evolutionary-ecological perspective. Sinauer Associates, Sunderland, pp 11–34
Hale JK (1969) Ordinary differential equations. Wiley, New York
Hofbauer J, Sigmund K (1998) Evolutionary games and population dynamics. Cambridge University Press, Cambridge
Hubbell SP (2001) The unified neutral theory of biodiversity and biogeography. Princeton University Press
Jang SR-J (2002) Dynamics of herbivore–plant–pollinator models. J Math Biol 44:129–149
Li B, Smith HL (2007) Global dynamics of microbial competition for two resources with internal storage competition model. J Math Biol 55:481–511
Llibre J, Xiao D (2014) Global dynamics of a Lotka–Volterra model with two predators competing for one prey. SIAM J Appl Math 74:434–453
Lundberg S, Ingvarsson P (1998) Population dynamics of resource limited plants and their pollinators. Theor Popul Biol 54:44–49
Memmott J, Waser NM, Price MV (2004) Tolerance of pollination networks to species extinctions. Proc R Soc Lond B 271:2605–2611
Nguyen DH, Yin G (2017) Coexistence and exclusion of stochastic competitive Lotka–Volterra models. J Differ Equ 262:1192–1225
Oleson JM, Bascompte J, Dupont YL, Jordano P (2007) The modularity of pollination networks. Proc Natl Acad Sci 104:19891–19896
Perko L (2001) Differential equations and dynamical systems. Springer, New York
Revilla T (2015) Numerical responses in resource-based mutualisms: a time scale approach. J Theor Biol 378:39–46
Sakavara A, Tsirtsis G, Roelke DL, Nancy R, Spatharis S (2018) Lumpy species coexistence arises robustly in fluctuating resource environments. Proc Natl Acad Sci 115:738–743
Scheffer AM, van Nes EH (2006) Self-organized similarity, the evolutionary emergence of groups of similar species. Proc Natl Acad Sci 103:6230–6235
Scheffer M, van Nes EH, Vergnon R (2018) Toward a unifying theory of biodiversity. Proc Natl Acad Sci 115:639–641
Terry I (2001) Thrips and weevils as dual, specialist pollinators of the Australian cycad Macrozamia communis (Zamiaceae). Int J Plant Sci 162:1293–1305
Tilman D (1982) Resource competition and community structure. Princeton University Press, Princeton
Traveset A, Richardson DM (2006) Biological invasions as disrupters of plant reproductive mutualisms. Trends Ecol Evol 21:208–215
Vanbergen AJ (2013) Threats to an ecosystem service: pressures on pollinators. Front Ecol Environ 11:251–259
Vanbergen AJ, Woodcock BA, Heard MS, Chapman DS (2017) Network size, structure and mutualisms dependence affect the propensity for plant–pollinator extinction cascades. Funct Ecol. https://doi.org/10.1111/1365-2435.12823
Wang Y, DeAngelis DL (2016) Stability of an intraguild predation system with mutual predation. Commun Nonlinear Sci Numer Simul 33:141–159
Acknowledgements
We would like to thank the anonymous reviewers for their helpful comments on the manuscript. Yuanshi Wang and Hong Wu were supported by NSF of People’s Republic of China (11571382). D. L. DeAngelis acknowledges the support of the US Geological Survey’s Greater Everglades Priority Ecosystem Sciences program. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government.
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix A. Proof of Proposition 2.1
Proof
Let \(N(t)= (N_1(t), N_2(t), N_3(t))\) be a solution of (2.3) with a fixed initial value \(N(0) \ge 0\). It follows from the first equation of (2.3) that
Let \(K_1 = \bar{r}_1 / d_1\). The comparison principle (Hale 1969) implies \( \limsup _{t \rightarrow \infty }N_1(t) \le K_1. \) Then for \(\delta _0 >0\) small, there is \(T_1 >0\) such that if \(t > T_1\), then \(N_1(t) \le K_1 + \delta _0 \).
It follows from the second equation of (2.3) that if \(t> T_1\), then
Let \(K_2 = [ a_{21} (K_1 + \delta _0) -r_2] / r_2 \). If \(K_2 \le 0\), then \(\lim _{t \rightarrow \infty } N_2(t) =0\), which implies that there is \(T_{21}> T_1 >0\) such that when \(t>T_{21}\), we have \(N_2(t) < \delta _0\). If \(K_2 > 0\), then
which implies \(\limsup _{t \rightarrow \infty } N_2(t) \le K_2 \). Then there is \(T_{22}> T_1 >0\) such that when \(t>T_{22}\), we have \(N_2(t) < K_2 + \delta _0\). Thus, there exists \(T_2 >0\) such that if \(t>T_2\), then \(N_2(t) < |K_2| + \delta _0\).
Let \(K_3 = [ a_{31} (K_1 + \delta _0) -r_3] / r_3 \). By the third equation of (2.3) and a discussion similar to that for \(K_2\), we obtain that there is \(T_3 >T_2\) such that if \(t > T_3\), then \(N_3(t) < |K_3| + \delta _0\).
Let \(T = T_3\). If \(t > T\), then \(||N(t)|| = \sum _{i=1}^3 N_i(t) \le \sum _{i=1}^3 M_i\) with \(M_i= |K_i|+ \delta _0\). Thus solutions of (2.3) are bounded. \(\square \)
Appendix B. Proof of two cases in Theorem 3.3
Proof
(ii) The case of \(\lambda _1^{(2)} = 0\).
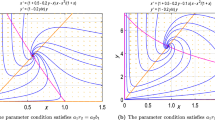
From \(\lambda _1^{(2)} = 0\), we have \(a_{21} = a_{21}^*\). If \(a_{21}^* < a_{21}^1\), there is no positive equilibrium in system (3.1) by Proposition 3.2. If \(a_{21}^* = a_{21}^1\), we have \(b_{12} = \bar{r}_1/2\) and \(a_{21} = a_{21}^0\), which implies that \(\Delta = 0\) and there is no positive equilibrium in system (3.1). Thus, \(E_1\) is globally asymptotically stable.
If \(a_{21} > a_{21}^1\), \(E_{12}^+\) is the unique positive equilibrium by Proposition 3.2. We apply the central manifold theorem to show that \(E_1\) has no stable manifold in int\(R_+^2\), which implies that \(E_{12}^+\) is globally asymptotically stable. The following transformation can change system (3.1) into a standard form near \(E_1\):
Then system (3.1) can be written as
which implies that the solution \(y_2 =0 \) is a stable manifold of equilibrium (0, 0) in system (6.1). Let \(y_1 = \phi (y_2) =a y_2^2 + o(y_2^2) \) be the central manifold of (6.1) at (0, 0). From \(a_{21} =a_{21}^* > a_{21}^1\), we have \(b_{12} = \hat{r}_1 - r_2 d_1 /a_{21} > \hat{r}_1 /2.\) A long but straightforward computation shows that
Thus equilibrium (0, 0) is a saddle-node point. On the central manifold \(y_1 = \phi (y_2) \), we have \(dy_2/dt >0\). Then equilibrium (0, 0) has no stable manifold in int\(R_+^2\). Thus, \(E_1\) has no stable manifold in int\(R_+^2\) and \(E_{12}^+\) is globally asymptotically stable.
(iii) The case of \(\lambda _1^{(2)} < 0, \Delta =0\) and \(a_{21} > a_{21}^1\).
From \(\lambda _1^{(2)} < 0\), we have \(a_{21} < a_{21}^*\) and equilibrium \(E_1\) is locally asymptotically stable.
Since \(\Delta = 0\) and \(a_{21} > a_{21}^1\), the two positive equilibria \(E_{12}^-\) and \(E_{12}^+\) coincide by Proposition 3.2. By (3.2), the Jacobian matrix of (3.1) at a positive equilibrium \((N_1,N_2)\) is
which implies tr\(J(E_{12}^\pm ) <0\). A direct computation shows that
which implies that there is a simple zero eigenvalue of \( J(E_{12}^-)\). We apply Sotomayor’s theorem Perko (2001) to show that saddle-node bifurcation occurs at \(E_{12}^-\) when \(\Delta =0\). For the simple zero eigenvalue, we have the left and right eigenvectors of \( J(E_{12}^-)\) by (6.2):
Let \(\mu = a_{21}\) be the parameter in Sotomayor’s theorem. Let \(F=(F_1,F_2)^T\) be the righthand side of (3.1). Then we have \( F_\mu = (0, N_1N_2g ) ^T.\) Thus \(w^T F_\mu = d_1N_1^2N_2g>0 \) at \(E_{12}^-\) and \(\mu =a_{21}^0\). Direct computations show that
A long but straightforward computation shows that
Thus, the Sotomayor’s theorem implies that saddle-node bifurcation occurs at \(E_{12}^-\) when \(a_{21} = a_{21}^0\): (1) If \(a_{21} > a_{21}^0\), then \( \det J (E_{12}^+) >0\) and \( \det J (E_{12}^-) <0\), which implies that \(E_{12}^+\) is a stable node and \(E_{12}^-\) is a saddle point. (2) If \(a_{21} = a_{21}^0\), \(E_{12}^-\) is a saddle-node point. (3) If \(a_{21} < a_{21}^0\), there is no positive equilibrium and \(E_1\) is globally asymptotically stable.
It follows from Proposition 3.1 that when \(E_{12}^-\) and \(E_{12}^+\) exist, the separatrices of \(E_{12}^-\) subdivide the interior of \((N_1,N_2)\)-plane into two regions: one is the basin of attraction of \(E_1\), while the other is that of \(E_{12}^+\). Thus, the result in the second case is proven. \(\square \)
Appendix C. Proof of Theorem 4.4
Proof
Since the proof for (ii) is similar to that for (ii) in Theorem 3.3, we omit the details.
-
(i)
Since \(\lambda _1^{(2)} > 0\), equilibrium \(E_1\) is a saddle point and has no stable manifold in int\(R_+^2\). From \(\lambda _1^{(2)} > 0\) we have \(a_{21} > a_{21}^*\) and \(a_{21} > a_{21}^0\). By Proposition 4.3, \(\hat{E}_{12}^+\) is the unique positive equilibrium. By Proposition 4.2, \(\hat{E}_{12}^+\) is globally asymptotically stable.
-
(ii)
It follows from \(\lambda _1^{(2)} < 0\) that the equilibrium \(E_1\) is asymptotically stable. From \(\lambda _1^{(2)}<0\) we obtain \(a_{21} < a_{21}^*\). When \(a_{21} \le a_{21}^1\), there is no positive equilibrium in system (4.1) by Proposition 4.3. Thus \(E_1\) is globally asymptotically stable.
Assume \(a_{21} > a_{21}^1\). If \(\Delta \ge 0\), then \(a_{21} \ge a_{21}^0\) and there are two positive equilibria \(\hat{E}_{12}^-\) and \(\hat{E}_{12}^+\) by Proposition 4.3. Let \(\hat{g}(N_2) = 1/(1 + N_2 + c N_2^s)\). By (4.2), the Jacobian matrix of (4.1) at a positive equilibrium \((N_1,N_2)\) is
which implies tr\(J(\hat{E}_{12}^\pm ) <0\). A direct computation shows that
Thus, if \(\Delta >0,\) then \(\hat{E}_{12}^-\) is a saddle point and \(\hat{E}_{12}^+\) is asymptotically stable.
When \( \Delta =0,\) equilibria \(\hat{E}_{12}^-\) and \(\hat{E}_{12}^+\) coincide and \( \det J(\hat{E}_{12}^-) =0\), which implies that there is a simple zero eigenvalue of \( J(\hat{E}_{12}^-)\). We apply Sotomayor’s theorem to show that saddle-node bifurcation occurs at \(\hat{E}_{12}^-\) when \(\Delta =0,\) i.e., \( a_{21} = a_{21}^0\). For the simple zero eigenvalue, we have the left and right eigenvectors of \( J(\hat{E}_{12}^-)\) by (6.4):
Let \(\mu = a_{21}\) be the parameter in Sotomayor’s theorem. Let \(\hat{F}=(\hat{F}_1,\hat{F}_2)^T\) be the righthand side of (4.1). Then \(\hat{F}_\mu = (0, N_1N_2\hat{g} ) ^T,\) which implies that \(w^T \hat{F}_\mu = d_1N_1^2N_2\hat{g}>0 \) at \(\hat{E}_{12}^-\) and \(\mu =a_{21}^0\). Direct computations show that
A long but straightforward computation shows that
Thus, the Sotomayor’s theorem implies that saddle-node bifurcation occurs at \(\hat{E}_{12}^-\) when \(\Delta =0\). By a discussion similar to the proof for Theorem 3.3(iii), the result in (iii) is proven. \(\square \)
Rights and permissions
About this article

Cite this article
Wang, Y., Wu, H. & DeAngelis, D.L. Global dynamics of a mutualism–competition model with one resource and multiple consumers. J. Math. Biol. 78, 683–710 (2019). https://doi.org/10.1007/s00285-018-1288-9
Received:
Revised:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00285-018-1288-9