
Abstract
The plants of the genus Salacia L. are the storehouse of several bioactive compounds, and are involved in treating human diseases and disorders. Hitherto, a number of reports have been published on in vitro biotechnology as well as microbial involvement in the improvement of Salacia spp. The present review provides comprehensive insights into biotechnological interventions such as tissue culture for plant propagation, in vitro cultures, and endophytic microbes for up-scaling the secondary metabolites and biological potential of Salacia spp. Other biotechnological interventions such as molecular markers and bio-nanomaterials for up-grading the prospective of Salacia spp. are also considered. The in vitro biotechnology of Salacia spp. is largely focused on plant regeneration, callus culture, cell suspension culture, somatic embryogenesis, and subsequent ex vitro establishment of the in vitro–raised plantlets. The compiled information on tissue cultural strategies, involvement of endophytes, molecular markers, and nanomaterials will assist the advanced research related to in vitro manipulation, domestication, and commercial cultivation of elite clones of Salacia spp. Moreover, the genetic diversity and other molecular-marker based assessments will aid in designing conservation policies as well as support upgrading and breeding initiatives for Salacia spp.
Key points:
• Salacia spp. plays a multifaceted role in human health and disease management.
• Critical and updated assessment of tissue culture, endophytic microbes, metabolites, molecular markers, and bio-nanomaterials of Salacia spp.
• Key shortcomings and future research directions for Salacia biotechnology.
Similar content being viewed by others


Introduction
The medicinal and aromatic plants have immense value for the well-being of humans. These plants are not only valued for food but also serve as major natural resources for bioactive molecules and remedies against several diseases. Moreover, the natural extracts, surfactants, and viscous juice of aromatic and medicinal plants possess enormous catalytic influence during synthesizing pharmaceutically important chemicals (Sapkal et al. 2023). Genus Salacia L. (Celastraceae-Salacioideae) possesses medicinally important plants with worldwide distribution in their native range in the tropics and subtropics. Plants of the genus Salacia are climbing shrubs, small trees, or lianas. They consist of over 200 species, primarily distributed in the Indian Subcontinent, African countries, and torrid-zone areas such as Brazil (Nandikar 2021). The maximum diversity of the species is found in South Africa (covering over half of the total count), and the remaining species occur in the neotropics and tropical Asia. The distribution of the plants of the genus Salacia is presented in Fig. 1. Many species are routinely used in numerous traditional systems of medicine due to the occurrence of numerous bioactive compounds belonging to xanthones, terpenes, phenolics, glycosides, thiosugars, etc. (Chavan et al. 2015a; Morikawa et al. 2021; Chavan and Santa‑Catarina 2023). In recent years, dietary supplements and health foods formulated from Salacia plants have been attaining more admiration in Japan and other countries (Dubey et al. 2011). The morphological similarities between Salacia plants, particularly in the medicinally used parts (roots and stems), become a serious challenge in the well-regulated usage of Salacia health foods. However, recent advancements in plant biotechnology, especially molecular markers, assisted in identifying the adulteration of other plant species and authenticating Salacia-based health foods (Zhu et al. 2021).

Worldwide distribution of Salacia spp
Inadequate cultivation practices and indiscriminate collection of many Salacia species for supplementing universal requirements of herbal medicine becomes a hurdle for the survival of the natural populations of many Salacia spp. However, plant tissue culture and recent developments in plant-based endophyte provide insights into the involvement of these tools in modulating the biosynthesis of numerous industrially important phytochemicals, which helped to reduce pressure on natural resources. Interventions of in vitro biotechnology and endophyte-mediated secondary metabolite production open new avenues for bioprospecting the Salacia spp.
Morphological attributes of Salacia spp.
Salacia species are scandent or sarmentosa shrub or small trees or climbing or strangling shrub. Stem color grayish to greenish, acute branching, and strangling stem. Leaves: elliptic or oblong, leaf blade shape narrowly ovate/round, leaf apex shape acute to acuminate, leaf base shape obtuse/wedge, or round or entire to crenulated leaf edges jagged/serrated, margins dentate, leaf blade shape elliptic/narrowly ovate. Flowers: inflorescence verticil, 3–6 flowers on axillary fascicles, petal 0.3 cm, pedicle 0.4–0.6 cm long, flower color yellowish green, stellate corolla, petals broadly ovate, sepal shape oblong or obovate or elliptic, sepal length up to 0.1 cm, anther thecae color orange, calyx lobe triangular, minute ovate to entire. Fruits: immature fruit color green, red when ripening, fruit 1–5 cm in diameter, fruit shape round/ovate, apex shape obtuse, fruit surface smooth/tuberculate/rugose, pericarp thin, flesh thickness thin, mesocarp pale yellow/light orange. Seeds: one-two seeded, shape round/oblong/ellipsoid color brown/light orange/gray. The phenotypic attributes of a few Salacia spp. are presented in Fig. 2.

Phenotypic attributes of Salacia spp. A Habit of S. chinensis; B inflorescence of S. chinensis; C flower of S. chinensis; D fruits of S. chinensis; E root of S. macrosperma; F stem of S. macrosperma
Traditional implications, phytochemistry, and biological properties
Many Salacia spp. have proven their role as traditional remedies for different diseases and disorders in humans. The published reviews and original articles provide an account of the phytochemical composition and biological properties of Salacia spp. (Paarakh et al. 2008; Arunakumara and Subasinghe 2010; Stohs and Ray 2015; Kushwaha et al. 2016; Morikawa et al. 2021). These reports highlight the possible role of Salacia spp. in the traditional medicinal system, as these plants are practiced as acrid, bitter, thermogenic, for urinary infections, astringent, anodyne, anti-inflammatory, depurative, emmenagogue, vulnerary, liver tonic, and stomachic and many more. The crude extracts and the bioactive constituents isolated possess pharmacological properties including antioxidant, anti-obese, anticancer, antiproliferative, anti-HIV etc. and as remedies for Alzheimer’s and Parkinson’s diseases (Chavan et al. 2013; Feng et al. 2019). Moreover, different plant organs accumulate several phytochemicals that are routinely used in clinical practices (Ghadage et al. 2017; Chavan and Santa‑Catarina 2023). The key phytochemicals reported from various Salacia spp. are presented in Fig. 3.

Noteworthy bioactive phytoconstituents of Salacia spp
Conventional propagation strategies
Conventional propagation using vegetative organs such as roots, shoots, leaves, and seeds is among the safe and effective routes for preserving genetic authenticity and producing metabolite-rich clones of medicinally important plants. The published articles confirm root and stem cuttings as common conventional strategies for propagation of Salacia spp. Available literature confirms that scientific knowledge on vegetative propagation for the genus Salacia was initiated by Sasidharan et al. (2010) in S. oblonga. This study reported the requirement of 6000 ppm of indole-3-butyric acid (IBA) application on the stem cuttings of S. oblonga, which resulted in 80% rooting and a 60% plant survival rate. On the other hand, Rathnayaka and Subasinghe (2010) successfully employed a pruning strategy for enhanced growth of stems and leaves of S. reticulata following different treatments with biofertilizers. The results suggested that pruning at 50 cm height produces a significantly higher number of branches, and even more branches are developed when plants are fertigated with T200.
Nayana et al. (2015) assessed the efficacy of the maturity stage of stem cuttings and potting media on vegetative propagation of S. reticulata. Soft wood, semi-hard wood, and hard wood stem cuttings of S. reticulata were transferred to a potting mixture of top soil, and compost (1:1) to assess the effect of maturity stage on the rooting of stem cuttings. Moreover, different types and concentrations of potting substrate such as sand, top soil, coir dust, and compost were tested either alone or in their combinations. The findings confirmed that the mixture of top soil and compost (1:1) emerged as the optimal potting substrate for planting semi-hard stem cuttings of S. reticulata. The root and stem cuttings of S. oblonga were treated with diverse applications of IBA (0–500 ppm), wherein the stem with leaves showed the maximum shooting response at 300 ppm, and the roots showed the maximum response at 200 ppm (Deepak et al. 2015a). Muhammad Anaz et al. (2017) developed a stem cutting–mediated vegetative propagation technique for five Salacia spp. including S. brunoniana, S. malabarica, S. oblonga, S. gambleana, and S. fruticosa. Among the different IBA concentrations, 4000 ppm (S. brunoniana), 7000 ppm (S. malabarica), 7000 ppm (S. oblonga), 8000 ppm (S. gambleana), and 7000 ppm (S. fruticosa) served optimum for the induction of roots and subsequent plantlet establishment. Recently, Gunaga and Vasudeva (2019) successfully employed the air-layering technique for the propagation of the threatened and potential anti-diabetic plant, Salacia macrosperma.
Tissue culture and in vitro propagation of Salacia spp.
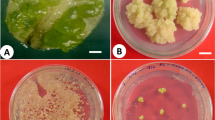
Plant tissue culture is a crucial biotechnological approach for large-scale propagation and subsequent conservation of endemic, endangered, aromatic, medicinal, and wild relatives of crop plants (Chavan et al. 2018a; Babar et al. 2020; Kaur et al., 2021; Chavan and Dey 2023). Micropropagation, somatic embryogenesis, callus culture, cell suspension culture, thin cell layer, synthetic seeds, and in vitro organ cultures are among the routinely employed tissue-culture strategies for propagation, conservation, secondary metabolite production, and improvement of the agronomical traits of several medicinal plants. These in vitro strategies are reliable, rapid, cost-effective, and able to produce a paramount number of saplings in a limited time duration. Nowadays, the trend has shifted toward large-scale production and obtaining medicinal plants with elite attributes of commercial importance using tissue culture-based propagation strategies (Kshirsagar et al. 2021; Sanyal et al. 2023a). Salacia spp. are propagated using various in vitro propagation systems for different purposes. Table 1 represents the various in vitro regeneration approaches for Salacia spp. Moreover, the in vitro regeneration stages of S. chinensis are presented in Fig. 4.

The selection of explants and their surface disinfection have prime importance in plant tissue culture techniques . An attempt at in vitro culture establishment in Salacia spp. was made using nodal segments, especially in the cultures that were initiated for shoot multiplication and propagation purposes (Sasidharan et al. 2010; Dhanasri et al. 2013; Chavan et al. 2015b; Deepak et al. 2015b; Majid et al. 2017, 2018; Laxmi et al. 2018; Kamat et al. 2021; Mahendra et al. 2020a). However, diverse surface sterilization procedures have been employed for different Salacia spp. Most of the nodal explants cleaned undergoing tap water wash followed by immersion in different types of detergents. Mercuric chloride is a monotonous surface disinfectant used for sterilizing the nodal explants of S. chinensis (Majid et al. 2014; Chavan et al. 2015b; Laxmi et al. 2018; Kamat et al. 2021), S. macrosperma (Mahendra et al. 2020a), and S. oblonga (Deepak et al. 2015b), wherein sodium hypochloride has been used to disinfect nodal segments of S. reticulata (Dhanasri et al. 2013) and S. chinensis (Majid et al. 2016, 2017). Shoot tip explants have also been tested for their efficiency during establishing in vitro cultures of S. chinensis (Majid et al. 2016, 2017), whereas in both attempts, the shoot tips were surface sterilized with sodium hypochloride. Kamat et al. (2021) established the in vitro cultures of S. chinensis through sodium hypochloride–treated seed explants. Callus cultures of most of the Salacia spp. have been initiated using leaf explants (Chavan et al. 2015b, 2021, 2022; Majid et al. 2017; Mahendra et al. 2020a, b; Kamat et al. 2021), with a few exceptions such as node and shoot tip explants (Majid et al. 2017; Mahendra et al. 2020a).
Murashige and Skoog’s (MS) medium have been unanimously used for shoot induction and multiplication for almost all studied Salacia spp.; however, the requirements of distinct class and concentrations of plant growth regulators (PGRs) are found to be species specific. The first attempt at shoot formation has been reported for S. reticulata by Dhanasri et al. (2013). Among the distinct types and concentrations of PGRs tested, MS medium supplemented with a mixture of benzyl adenine (BA) (3.5 mg/l) and indole-3-acetic acid (IAA) (0.5 mg/l) was found to produce 10.64 ± 0.96 shoots per explant. For S. chinensis, nodes cultured on MS medium with 2.0 mg/l of BAP, 0.8 mg/l of α-naphthalene acetic acid (NAA), and 100 mg/l of ascorbic acid (AA) were found suitable for the production of shoots (6.7 ± 1.0) in 75% cultures (Chavan et al. 2015b). Majid et al. (2016) were able to produce 5.37 ± 0.02 shoots per explant in 87.81 ± 3.22% cultures when nodal buds were transferred to MS medium augmented with BAP (1.0 mg/l) and NAA (0.5 mg/l). In another attempt, the same research group tested elevated concentrations of BAP and NAA for shoot multiplication in S. chinensis (Majid et al. 2017), wherein maximum shoot induction frequency (93.33 ± 2.02%) with 5.12 ± 0.09 was accomplished when nodal buds were cultured on BAP (1.5 mg/l) and NAA (1.0 mg/l) enriched MS medium. In further studies, Majid et al. (2018) employed a similar protocol for the production of shoots in S. chinensis and assessed them for their anticancer and anti-diabetic potential. Further research on S. chinensis also confirmed the requirement of MS medium alongside BAP (2.0 mg/l) and NAA (0.8 mg/l) for shoot proliferation (Laxmi et al. 2018). Kamat et al. (2021) noted the highest shoot multiplication frequency for S. chinensis in MS medium supplemented with MS medium with BAP (3.5 mg/l) and IBA (1 mg/l). However, shoot multiplication from seed explants of S. chinensis was dependent on the addition of 2iP (2.0 mg/l) and BAP (3.0 mg/l) in MS medium (Kamat et al. 2021). A mixture of BAP (1.0 mg/l), NAA (0.5 mg/l), and TDZ (0.5 mg/l) was found suitable for maximum shoot production in S. macrosperma (Mahendra et al. 2020a), wherein an average of 13.00 ± 0.57 shoots per explant have been produced. MS medium mixed with BAP (3.5 mg/l) and IBA (1 mg/l) was found suitable for shoot multiplication in S. oblonga (Deepak et al. 2015b).
Callus cultures provide a platform for indirect plant regeneration as well as enhanced secondary metabolite accumulation. For the genus Salacia, the first attempt at callus culture has been reported for S. chinensis (Chavan et al. 2015b). The results confirmed the addition of 2,4-D (2.0 mg/l) and BAP (1.5 mg/l) in MS medium for leaf-derived calli induction and proliferation calli (92% of callus induction frequency). In subsequent studies, Majid et al. (2017) achieved over 93.43 ± 2.75% callus induction frequency in S. chinensis when explants inoculated on MS medium supplemented with NAA (1.0 mg/l) and BAP (2.0 mg/l). Chavan et al. (2021) employed MS medium alongside BAP (2.0 mg/l), NAA (0.8 mg/l), and JA (75 μM) for calli biomass production from the leaves of S. chinensis. The calli biomass was further assessed for its phytochemical composition and antioxidant potential. The same procedure has been practiced by Chavan et al. (2022) for calli production to assess their antibacterial properties. In contrast, MS medium with exclusive supplementation of TDZ (0.5 mg/l) was found suitable for leaf segment–derived callus induction in 100% cultures of S. chinensis (Kamat et al. 2021). The calli were further practiced for their indirect shoot regeneration efficiency, where 12.33 ± 0.33 shoots per calli explant were recorded in 85% cultures (Kamat et al. 2021). Mahendra et al. (2020b) achieved callus production in 98.33% of culture vessels when leaf segments of S. macrosperma were transferred to MS medium with 2,4-D (2.5 mg/l) and BAP (1.5 mg/l). The same observation was recorded in further studies of S. macrosperma (Mahendra et al. 2020a). The pro-embryos were developed in 60% liquid cultures from leaf-derived calli.
In general, rooting of in vitro–generated shoots largely depends on media type and strength, PGR concentrations and combinations, plant genotype, incubation conditions, etc. For the genus Salacia, different concentrations and combinations of auxins and cytokinins have been tested for in vitro rooting; however, it was observed that MS medium fortified with either IBA or IAA served better for root induction in Salacia spp. It was also noted that there was no induction of roots in a PGR-free medium. Dhanasri et al. (2013) reported the best root induction frequency when in vitro–raised shoots of S. reticulata were transferred to half-strength MS medium supplemented with IBA (2.0 mg/l). In this media composition, 3.05 ± 1.55 roots have been developed. Likewise, the shoots of S. chinensis rooted best in IBA (1.5 mg/l) enriched half-strength MS medium, wherein rooting was observed in 88% of cultures and 5.3 ± 0.2 roots were produced per shoot (Chavan et al. 2015b). In subsequent studies on S. chinensis, the half-strength MS medium with IBA (2.0 mg/l) served better for in vitro root induction (Majid et al. 2016, 2017, 2018). In contrast, Kamat et al. (2021) confirmed the utility of IAA (0.5 mg/l) instead of IBA for successful in vitro rooting in S. chinensis. In vitro rooting in microshoots of S. oblonga was achieved by the incorporation of IBA (0.5 mg/l) in MS medium (Deepak et al. 2015b). Moreover, microshoots of S. macrosperma also rooted best (4.33 ± 0.88 roots/shoot) in MS medium supplemented with IAA at 1.0 mg/l of concentration (Mahendra et al. 2020a).
The efficiency of different planting substrates has been assessed during the ex vitro establishment of in vitro–raised plantlets of different Salacia spp. In vitro–raised plantlets of S. reticulata with well-developed shoots and roots were transferred to a mixture of soil and vermicompost (2:1) for hardening and acclimation (Dhanasri et al. 2013). Garden soil, river sand, and coco peat (1:1:1) was served optimum during the ex vitro establishment of S. chinensis plantlets, wherein over 80% survival rate of plantlets has been recorded (Chavan et al. 2015b). However, Majid et al. (2016) transplanted the in vitro–derived plantlets of S. chinensis into plastic pots with sterile compost consisting of organic fertilizer, sand, and peat (2:2:1) and maintained them at 18–28 °C with relative humidity ranging from 75 to 90%. Over 87% of survivability in field conditions was recorded. In subsequent studies from the same research group, a similar composition of planting substrate served better during the hardening of S. chinensis plantlets (Majid et al. 2017). On the other hand, sand, soil, and vermiculite (1:1:1) supported the ex vitro growth of S. chinensis plantlets (Kamat et al. 2021); however, a vermiculite and perlite mixture (1:1) has been required during the hardening of S. macrosperma plantlets with 80% survivability (Mahendra et al. 2020a). A mixture of soil and vermiculate (1:1) supported the acclimatization of in vitro–raised plantlets of S. oblonga (Deepak et al. 2015b).
In general, the tissue culture and in vitro regeneration of different Salacia spp. require MS medium during different regeneration stages. The shoot formation was significantly influenced by PGRs; however, BAP and 2,4-D play crucial roles during shoot multiplication. In vitro rooting has been totally dependent on the incorporation of either IBA or IAA in half-strength MS medium, wherein IBA showed superiority over IAA. Sand and soil are the common planting substrates required during the ex vitro establishment of tissue culture–raised plantlets of different Salacia spp.
In vitro cultures and endophytic microbes in elevating secondary metabolite production and bioactivities
Involvement of diverse types of in vitro culture systems is among the regular exercises for augmenting secondary metabolite yield in several medicinally important plants. The shoots, roots, callus, and a mixture of roots and shoots obtained in vitro, and roots from micropropagated plants and roots from field-grown plants of S. chinensis have been assessed for mangiferin content (Chavan et al. 2015b). The results confirmed the higher accumulation of mangiferin (593.87 ppm) in the callus cultures of S. chinensis on MS medium supplied with 2,4-D (2.0 mg/l) and BAP (1.5 mg/l). Similarly, different in vitro regeneration stages significantly alter the metabolite profile of numerous medicinal and aromatic plants (Chavan et al. 2018b). Majid et al. (2018) evaluated the micropropagated plantlets of S. chinensis for their antioxidant and anti-diabetic properties using DPPH, FRAP, α-amylase, and α-glucosidase assays. The experimental outcome confirmed the superiority of micropropagated plantlets over field-grown plantlets with respect to their antioxidant and anti-diabetic potential.
A recent study on elicitation confirmed the augmentation of calli biomass, polyphenolics, mangiferin yield, and antioxidant properties of S. chinensis callus cultures (Chavan et al. 2021). The calli established on MS medium supplemented with 2,4-D (2.0 mg/l), BAP (1.5 mg/l), and treated with jasmonic acid (75 µM) served best for achieving the highest calli biomass (8.10 ± 0.6 g of fresh weight), the content of total phenolics (68.49 ± 0.90 mg of gallic acid equivalent per grams of dry weight), flavonoids (26.18 ± 0.35 mg of quercetin equivalent per gram of dry weight), and mangiferin content (8.493 ± 0.193 mg per gram of dry weight). The same elicitor treatment also augmented the antioxidant potential of calli when analyzed using DPPH, FRAP, and metal chelating assays. In another study, Chavan et al. (2022) reported that the same media composition and elicitor treatment enhanced the antibacterial potential of calli against Proteus vulgaris. In contrast, the antimicrobial potential has been reported to be higher in the leaves of field-grown plants than the in vitro–derived calli of S. macrosperma (Mahendra et al. 2020b). The leaf extract is known to possess strong antibacterial activity against E. coli (15.50 mm), Salmonella typhi (17.33 mm), Staphylococcus aureus (15.50 mm), Bacillus subtilis (17.16 mm), and Pseudomonas aeruginosa (16.66 mm). Kamat et al. (2021) evaluated the in vitro–raised leaves and calli of S. chinensis for their phytochemical composition. Gas chromatography–mass spectrophotometric analysis detected 16 and 11 phytochemicals in calli and leaves, respectively. Moreover, calli also possessed an almost fivefold higher amount of mangiferin (26 mg/ml) as compared to leaves (4 mg/ml).
Endophytes are the microbes that reside at least a part of their life cycle in plants and animals without causing any harmful symptoms. Several medicinal and aromatic plant species have been used as complementary forms of medicine since ancient times. Current research on the bioprospecting of endophytes for secondary metabolite production confirms the highest priority in the scientific world (Sanyal et al. 2023b). The work on entophyte isolation, characterization, and assessment of their pharmacological potential for Salacia spp. has begun since the last decade (Table 2). Bhagya et al. (2011) isolated a fungal endophyte, Colletotrichum gloeosporioides Penz, from the stem and leaves of S. chinensis, which showed sensitivity toward Amphotericin B and Nystatin above a concentration of 100 μg/ml. In subsequent studies, Sheik et al. (2020) confirmed the role of C. gloeosporioides as an anticancer agent. Moreover, C. gloeosporioides is significantly involved in decolorizing carcinogenic dyes such as methylene blue and Congo red. Roopa et al. (2017) performed fungal endophyte diversity assessment on different species of Salacia (S. chinensis, S. oblonga, S. fruticosa, and S. macrosperma) and reported the Salacia spp. with rich endophytic microbes. The results confirmed the occurrence of 15 fungal endophytes such as Alternaria alternata, Aspergillus niger, Cladosporium herbarum, Colletotrichum spp., Curvularia spp., Diaporthe perjuncta, Drechslera spp., Phoma spp., Penicillium notatum, Myrothecium verrucaria, Gliocladium roseum, Fusarium oxysporium, Sterile spp., Trichophyton mentagrophytes, and Xylaria spp. from the stem of S. chinensis. This study also confirmed the occurrence of 12 fungal endophytes such as Botryosphaeria rhodina, Cladosporium herbarum, Trichoderma longibrachiatum, Aspergillus terreus, A. niger, Fusarium oxysporium, Lasiodiplodia theobromae, Penicillium notatum, Phoma spp., Sterile spp., Coriolopsis caperata, and Pestalotiopsis from the stem of S. oblonga. Ten different fungal endophytes, viz., Drechslera spp., Curvularia spp., Cladosporium herbarum, Pestalotiopsis spp., Trichoderma longibrachiatum, Aspergillus terreus, Colletotrichum spp., Diaporthe perjuncta, Myrothecium verrucaria, and Xylaria spp. were isolated from the stem of S. fruticosa (Roopa et al. 2017). This study also identified nine fungal endophyes such as Aspergillus terreus, A. niger, Phoma spp., Cladosporium herbarum, Colletotrichum spp., Penicillium notatum, Trichophyton mentagrophytes, Alternaria alternata, and Fusarium oxysporium from the stem of S. macrosperma.
Webster et al. (2020) reported four bacterial endophytes such as Klebsiella spp., K. variicola, Stenotrophomonas spp., and Enterobacter spp. from S. chinensis; furthermore, this study also identified a fungal endophyte, Bacillus spp., in S. macrosperma. The fungal endophytes Penicillium capsulatum and Aspergillus fumigatus isolated from the stem and roots of S. chinensis and roots of S. oblonga significantly modulate the mangiferin biosynthesis (Kaur et al. 2022). Mangiferin is a naturally occurring multipotent xanthone that possesses numerous pharmacological properties and is involved in treating numerous human diseases and disorders (Chavan et al. 2015a; Feng et al. 2019). Eight fungal endophytes, i.e., Botryosphaeria rhodina, Trichoderma longibrachiatum, Lasiodiplodia theobromae, Aspergillus niger, A. terreus, Coriolopsis caperata, Phomopsis spp., and Fusarium solani, have been isolated from the stem and roots of S. oblonga, where Alternaria, Fusarium, and Aspergillus niger significantly modulate taxol biosynthesis (Roopa et al. 2015). Taxus brevifolia and their endophytic microbes are the original sources of taxol, a widely used diterpenoid as a chemotherapy drug to treat lung, breast, and other types of cancer (Stierle et al. 1995). The stem and roots of S. reticulata also host four bacterial endophytes, such as B. licheniformis, B. subtilis, B. zanthoxyli, B. aryabhattai, and a fungal endophyte, i.e., Aspergillus spp. (Bhosale et al. 2021). Recently, Roopa et al. (2022) investigated different plant parts of four Salacia spp. (S. chinensis, S. oblonga, S. fruticosa, and S. macrosperma) for assessing endophyte diversity and their potential role in augmenting biological properties as well as the accumulation of secondary metabolites. The results confirmed that F. oxysporum, Penicillium notatum, Pestalotiopsis spp., Phoma spp., and Colletotrichum spp. have been found predominantly in Salacia spp. Moreover, these endophytes have shown potential anti-diabetic activity through alpha-amylase inhibitor activity and accumulate steroids, tannins, sugars, proteins, flavonoids, saponins, terpenoids, and glycosides at different levels.
The literature on the isolation and characterization of endophytes from Salacia spp. confirms the occurrence of fungal as well as bacterial microbes, which accumulate numerous secondary metabolites and significantly modulate the biosynthesis of marker compounds. These findings strongly suggest that the crude extracts and the phytochemicals isolated from the endophytic microbes of Salacia spp. can be industrialized as potent drug molecules in view of their pharmaceutical significance.
Molecular markers for assessment and improvement of Salacia spp.
The advancement in DNA-based molecular markers has become a milestone in plant science research. These molecular markers are commonly practiced for the improvement of numerous plants, especially through plant breeding, genetic diversity analysis, population genetics, phylogenetics, mapping of mutations, assessment of genetic uniformity among tissue culture–raised plantlets, identifying adulteration in health-based foods, taxonomy and evolution, confirming the identity of individuals, etc. Literature on the utility of molecular markers confirmed their extensive role in genetic diversity assessment, phylogenetic studies, genetic integrity analysis of micropropagated plantlets, and authentication of health foods from Salacia spp (Table 3).
The first attempt at the integration of molecular markers in the genus Salacia was by Savolainen et al. (1994). The phylogenetic analysis of genera falling under order Celastrales through rbcL confirmed the familiarity of genus Salacia with genera Euonymus and Hippocratea. In subsequent studies on phylogenetics, 26S nrDNA, phytochrome B, rbcL, and atpB nested four Salacia spp. (S. impressifolia, S. nitida, S. pallescens, and S. undulata) with close genera such as Catha, Maytenus, and Pristimera (Simmons et al. 2001). The Indian clade of Salacia has been characterized through the nuclear ribosomal DNA (nrDNA) internal transcribed spacer (ITS 1 and ITS 2) region for establishing the relationship among species (Devipriya et al. 2015). The investigation of a total of 24 species of the genus Salacia (8 onsite collection and 16 through GenBank accession), 6 allied genera, and 2 outgroups showed divergence among species, and it was also confirmed that the morphological species demarcation did not corroborate with the ITS phylogeny. In further studies, Dev et al. (2015) confirmed the species discrimination through ITS2, trnH-psbA, matK, and rbcL across one variety and eight species of genus Salacia, including S. beddomei, S. chinensis, S. fruticosa, S. macrosperma, S. malabarica, S. oblonga var. oblonga, S. oblonga var. kakkayamana, S. vellaniana, and S. agasthiamalana with 100% accuracy. Alongside the ITS marker, RAPD and ISSR also significantly contributed toward assessing genetic diversity and establishing possible relationships among Salacia spp., viz., S. chinensis, S. macrosperma, S. fruticosa, and S. oblonga (Bajpe et al. 2018). The molecular fingerprints confirmed the polymorphism revealed by RAPD (41.45 ± 10%), ISSR (33.58 ± 6.52%), and ITS (25.50 ± 17.25%), which revealed significant variance within and among the Salacia spp. Recently, ITS-2 secondary structure-based molecular characterization has been done for construction phylogeny of eight Salacia spp. (Anaz et al. 2021). The phylogenetic tree based on the sequence and structural features of ITS 2 confirmed the two evolutionary lines among Salacia spp. One line leads to the present-day S. chinensis, and the other line further diversifies and leads to the rest of the Salacia spp., i.e., S. agasthiamalana, S. brunoniana, S. fruticosa, S. gambleana, S. oblonga, S. malabarica, and S. wayanadica.
RAPD and ISSR markers contributed significantly to detect genetic divergence within and among the Salacia spp. The RAPD marker was found suitable for the assessment of genetic variation among different populations of S. chinensis (Mulye et al. 2013). Further investigations on RAPD analysis reported the greatest genetic polymorphism among different samples of S. oblonga (Maheswari et al. 2013). Different accessions of S. reticulata were also analyzed using RAPD markers, and the results also supported the previous findings with the extent of polymorphism among the accessions (Dhanasri et al. 2015). In subsequent studies, Priya et al. (2019) also detected genetic variability within and among the eight Salacia spp. using RAPD markers. The results also confirmed the distinctiveness of S. chinensis from other species such as S. beddomei, S. fruticosa, S. gambleana, S. macrosperma, S. malabarica, and S. oblonga. In another study, an average of 29.21 ± 7.89% of polymorphism among four Salacia spp. such as S. chinensis, S. fruticosa, S. macrosperma, and S. oblonga was detected by ISSR markers (Badiger et al. 2021).
DNA barcoding is the most effective and emerging method of molecular identification of flowering plants as well as plant-based health foods (Kshirsagar et al. 2017; Anaz et al. 2021). A literature survey confirmed that DNA barcodes have been developed for authentication and accurate identification of several Salacia spp. Badiger et al. (2021) developed barcodes using rbcL, matK, and ITS markers for accurate identification of four Salacia species, viz., S. chinensis, S. fruticosa, S. macrosperma, and S. oblonga. In another study, Zhu et al. (2021) employed nrDNA ITS sequences and RFLP markers for genetic identification and confirmation of Salacia-based health food authentication. The markers found useful to authenticate Salacia spp., and the results also confirmed that S. chinensis and S. reticulata are the chief sources of commercially existing Salacia-derived food products.
Confirmation of genetic integrity among tissue culture–raised clones of ornamental, medicinally important, and agricultural crop plants is the key stage before their large-scale propagation and commercialization. Molecular markers including RAPD, RFLP, AFLP, FRAPD, SSR, ISSR, SSAP, and SCoT have been regularly utilized to confirm the genetic integrity of tissue culture–regenerated plantlets of numerous flowering plants (Chavan et al. 2015b; Bhattacharyya et al., 2013; Majid et al. 2018). In Salacia, few reports focused on the genetic stability analysis of tissue-culture plantlets. Chavan et al. (2015b) confirmed the genetic uniformity of in vitro propagated plantlets of S. chinensis using ISSR and RAPD markers. Combined, 3871 bands were produced by 17 ISSR and 12 RAPD primers across a mother plant and 20 micropropagated plantlets, wherein 3858 (99.66%) bands showed similarity among tested plantlets and the rest of the bands showed divergence. The high level of similarity confirmed the genetic homogeneity of the micropropagated clones of S. chinensis (Fig. 5). Another report on S. chinensis detected 5% variation among donor and in vitro–propagated plantlets as assessed by FRAPD (Majid et al. 2018).

Genetic fidelity assessment of in vitro–raised plantlets of S. chinensis by ISSR analysis (UBC: 801)
Complete plastome sequencing is a vital intervention for knowing the complete genetic make-up of organisms which supports policy design, especially conservation and improvement of plants. Lin et al. (2019) performed the complete plastome sequencing for S. amplifolia, which is the only report available for the genus Salacia. The analysis confirmed the size of the complete plastome (163,255 bp in length) which contains 113 genes, of which 79 are unique protein-coding genes, 30 are unique tRNA genes, and 4 are unique rRNA genes. The utility of molecular markers in microbial endophyte improvement is well established in flowering plants. Only a report by Roopa et al. (2015) addressed the ITS-based genome mining approach for the discovery of novel taxol-producing endophytic fungi from S. oblonga.
RAPD, ISSR, and FRAPD are routinely practiced for the genetic diversity analysis and confirmation of genetic integrity among tissue culture–raised plantlets of Salacia spp. However, ITS, phytochrome B, rbcL, atpB, trnH-psbA, and matK are among the commonly used markers for establishing phylogeny and developing barcodes for the precise authentication of species and identifying Salacia-based health food adulterations.
Salacia-based nanomaterials in enhancing biological properties
Plant-based nanomaterials are routinely used to elevate secondary metabolites as well as the biological properties of several plant species. Such biogenic nanomaterials have advantages over other nanomaterials because they are biodegradable, biocompatible, and generally recognized as safe (Patil and Chandrasekaran 2020). Different types of nanomaterials have been fabricated from various organs of Salacia spp. The first attempt at the synthesis, characterization, and analysis of biological potential was by Jadhav et al. (2015) for S. chinensis. The results confirmed the antibacterial potential of silver nanoparticles synthesized from bark, which were found to be effective against Staphylococcus aureus and Pseudomonas aeruginosa. The aqueous leaf extract of S. chinensis have been used for synthesis of silver and copper nanomaterials (Chavan and Ghadage 2018). Both the nanoparticles have potential antibacterial properties against Bacillus subtilis, Escherichia coli, S. aureus, and P. aeruginosa that validated the action of these nanoparticles against these infectious microbes. In further studies in S. chinensis, the silver nanoparticles have the potential to enhance antioxidant and antimicrobial potential (Abhijit et al. 2019). The synthesized nanoparticles were assessed for their antioxidant potential using a DPPH assay, and it was found that nanoparticles have potent scavenging activity over crude extract. Moreover, the antimicrobial potential of the particles was tested against pathogenic fungi and bacteria, wherein nanomaterials showed moderate growth inhibition on E. coli, Candida albicans, and C. tropicalis. Recently, Nagesh et al. (2022) synthesized silver nanomaterials from the roots of S. chinensis, which is able to modulate biological properties such as antibacterial, antifungal, and antiproliferative effects. The leaf extract–derived silver nanoparticles from S. mulbarica proved to be a better antibacterial agent against E. coli and B. subtilis and also possess the capability of ct-DNA damage via releasing of reactive oxygen species (Espenti et al. 2020). In recent times, environmentally benign silver bio-nanomaterials have been synthesized from the root extracts of S. oblonga and served as potent antioxidant, antibacterial, and anti-diabetic agents (Dugganaboyana et al. 2023). Recently, Sabeena et al. (2023) performed a comparative study on chemical and green approaches for synthesis of nanocomposites from leaf extracts of S. reticulata and subsequently characterized for in vivo and in vitro biological properties. The green synthesized nanocomposites, i.e., iron oxide, cerium oxide, titanium dioxide, silica gel, and chitosan, of S. reticulata possesses strong anti-inflammatory, antibacterial, and anti-diabetic properties, as well as considerable suppression of high activation in in vivo zebrafish embryo toxicity (Sabeena et al. 2023).
Among the metal nanoparticles, gold nanoparticles are recognized as the most potent, biocompatible, and environment friendly (Khan et al. 2019). The first attempt at the synthesis and characterization of nanoparticles (gold) in the genus Salacia was by Jadhav et al. (2018). The gold nanoparticles synthesized from the bark of S. chinensis were evaluated for their osteoinductive potential for application in implant dentistry. The results confirmed that the stable, biocompatible, and eco-friendly gold nanoparticles can be used as an effective bone inductive agent during dental implant therapy. The gold nanoparticles have been synthesized using leaf extracts of S. fruticosa, which have antibacterial activity against the bacterial pathogens such as S. aureus and P. aeruginosa (Keshavamurthy and Ravishankar Rai 2021). The results on green nanomaterial synthesis using different plant organs of Salacia spp. are mainly through silver, copper, gold, iron, cerium, titanium, silica, and chitosan nanoparticles. These nanomaterials proved their role in elevating the biological potential of Salacia spp. Moreover, these green synthesized nanomaterials are more affordable, environmentally friendly, and biocompatible than chemically synthesized ones.
Conclusion and future perspectives
In conclusion, the in vitro biotechnology of Salacia spp. is mainly focused on in vitro regeneration and secondary metabolite production. Nodal segments are among the commonly used explants for initiating the cultures, while leaves served better for initiating callus cultures. MS medium is the exclusive requirement during various in vitro stages alongside diverse types and concentrations of PGRs, especially BAP and either IBA or IAA for shoot initiation and multiplication in most of the Salacia spp. IBA supported in vitro rooting in almost all Salacia spp., while a combination of BAP and 2,4-D served better for induction of callus. Different in vitro regeneration stages and the elicitation of various cultures significantly enhance the accumulation of secondary metabolites. Moreover, microbial endophytes and nanomaterials (silver, copper, and gold) also up-scale the biochemical marker compounds and bio-efficacy of different Salacia spp. Molecular markers found suitable for assessing genetic diversity among and between the species (ITS, 26S nrDNA, Phytochrome B, rbcL, atpB, trnH-psbA, matK, RAPD, ISSR) confirmed the genetic stability of tissue culture–raised plantlets (RAPD, FRAPD, and ISSR), as well as authentication and identification of Salacia spp. (ISSR, rbcL, matK, ITS). Moreover, it serves as an authentic tool for confirmation of adulteration in Salacia-based health foods (nrDNA ITS sequences, RFLP). There are several untouched areas of research on in vitro biotechnology of Salacia spp., such as synthetic seed production, cryopreservation, agrobacterium-mediated genetic transformation, and omics, that deserve special attention to advance the biotechnology of Salacia spp.
Data availability
All data generated or analyzed during this study are included in the submitted manuscript.
References
Abhijith P, Bhavya G, Ashika P, Geetha N (2019) Characterization of silver nanoparticles from the endangered medicinal plant Salacia chinensis and analysis of its bioactive potentials. Life Sci Edge 5(1):43–50
Anaz KM, Sasidharan N, Dilsha MV (2017) Standardization of propagation through stem cuttings of five Salacia species, important antidiabetic medicinal plants of Western Ghats. Int J Adv Res 5(9):500–505
Anaz KM, Sasidharan N, Remakanthan A, Dilsha MV (2021) ITS 2 and RNA secondary structure-based analysis reveals a clear picture on phylogeny of South Indian Salacia spp. Comput Biol Chem 91:107438
Arunakumara KKIU, Subasinghe S (2010) Salacia reticulata Wight: a review of botany, phytochemistry and pharmacology. Trop Agric Res Ext 13(2):2010
Babar PS, Deshmukh AV, Salunkhe SS, Chavan JJ (2020) Micropropagation, polyphenol content and biological properties of Sweet Flag (Acorus calamus): a potent medicinal and aromatic herb. Vegetos 33:296–303
Badiger AS, Rudrappa MK, Bajpe SN (2021) Assessing genetic diversity and DNA barcodes efficiency of Salacia and related species from Western Ghats. Karnataka J Appl Biol Biotechnol 9(2):67–74
Bajpe SN, Bharathi TR, Marulasiddaswamy KM, Kumara KKS, Prakash HS, Kini RK (2018) Efficiency of RAPD, ISSR and ITS markers in detecting genetic variability among Salacia species sampled from the Western Ghats of Karnataka. Mol Biol Rep 45:931–941
Bhagya N, Sheik S, Sharma MS, Chandrashekar KR (2011) Isolation of endophytic Colletotrichum gloeosporioides Penz from Salacia chinensis and its antifungal sensitivity. J Phytol 3(6):20–22
Bhattacharyya P, Kumaria S, Kumar S, Tandon P (2013) Start Codon Targeted (SCoT) marker reveals genetic diversity of Dendrobium nobile Lindl., an endangered medicinal orchid species. Gene 529(1):21–26
Bhosale R, Sakkan S, Kolhe S, Yewale S, Rudraiah SM, Padmanabhan S (2021) Characterization of the microbes isolated from Salacia reticulata raw material and its extract: a simple method to reduce the microbial load in herbal products. Acta Sci Microbiol 4(12):119–129
Chavan JJ, Dey A (2023) Zingiber zerumbet (L.) Roscoe ex Sm.: biotechnological advancements and perspectives. Appl Microbiol Biotechnol 107:5613–5625
Chavan JJ, Ghadage DM (2018) Biosynthesis, characterization and antibacterial capability of silver and copper nanoparticles using aqueous leaf extract of Salacia chinensis L. J Nanomed Nanotechnol 9:1
Chavan JJ, Santa-Catarina C (2023) Multidirectional insights into nutritional, phytochemical, antioxidant capability and multivariate analysis of underutilized edible berry plant (Salacia macrosperma Wight) - a novel source for food and pharmaceutical industry. Food Chem Adv 2:100284
Chavan JJ, Jagtap UB, Gaikwad NB, Dixit GB, Bapat VA (2013) Total phenolics, flavonoids and antioxidant activity of Saptarangi (Salacia chinensis L.) fruit pulp. J Plant Biochem Biotechnol 22(4):409–413
Chavan JJ, Ghadage DM, Bhoite AS, Umdale SD (2015a) Micropropagation, molecular profiling and RP-HPLC determination of mangiferin across various regeneration stages of Saptarangi (Salacia chinensis L.). Ind Crops Prod 76:1123–1132
Chavan JJ, Ghadage DM, Kshirsagar PR, Kudale SS (2015b) Optimization of extraction techniques and RP-HPLC analysis of antidiabetic and anticancer drug mangiferin from roots of ‘Saptarangi’ (Salacia chinensis L.). J Liq Chromatogr Relat Technol 38:963–969
Chavan JJ, Gaikwad NB, Dixit GB, Yadav SR, Bapat VA (2018a) Biotechnological interventions for propagation, conservation and improvement of ‘Lantern Flowers’ (Ceropegia spp.). S Afr J Bot 114:192–216
Chavan JJ, Kshirsagar PR, Pai SR, Pawar NV (2018b) Micropropagation, metabolite profiling, antioxidant activities and chromatographic determination of bioactive molecules across in vitro conditions and subsequent field cultivation stages of ‘Shampoo Ginger’ (Zingiber zerumbet L. Roscoe ex Sm). Biocatalysis Agricult Biotechnol 16:79–89
Chavan JJ, Kshirsagar PR, Jadhav SG, Nalavade VM, Gurme ST, Pai SR (2021) Elicitor-mediated enhancement of biomass, polyphenols, mangiferin production and antioxidant activities in callus cultures of Salacia chinensis L. 3 Biotech 11(6):285
Chavan JJ, Ghorpade VM, Bhamare MR, Kakulte VR (2022) Antibacterial potential of elicitor-treated callus cultures of Saptarangi (Salacia chinensis L.). J Inf Comput Sci 12(8):114–125
Deepak KGK, Suneetha G, Surekha Ch (2015a) A simple and effective method for vegetative propagation of an endangered medicinal plant Salacia oblonga Wall. J Nat Med 70:115–119
Deepak KGK, Suneetha G, Surekha Ch (2015b) In vitro clonal propagation of Salacia oblonga Wall. An endangered medicinal plant. Ann Phytomed 4(2):67–70
Dev SA, Anoop BS, Anoja K, Udayan PS, Muralidharan EM (2015) Species discrimination through DNA barcoding in the genus Salacia of the Western Ghats in India. Nord J Bot 33:722–728
Devipriya MS, Devipriya V, Udayan PS, Yohannan R (2015) Molecular systematics of Indian Salacia based on ITS sequences of nrDNA. Int J Adv Res 3(7):1513–1521
Dhanasri G, Srikanth Reddy M, Naresh B, Cherku PD (2013) Micropropagation of Salacia reticulata - an endangered medicinal plant. Plant Tissue Cult Biotechnol 23(2):221–229
Dhanasri G, Srikanth Reddy M, Naresh B, Devi P (2015) Molecular genetic analysis of Salacia reticulata, a threatened medicinal plant for the study of genetic diversity. Int J Pure App Biosci 3(1):92–99
Dubey GP, Agarwal A, Vyas N, Rajamanickam VG (2011) Herbal formulation for the prevention and management of diabetes mellitus and diabetic micro-vascular complications. https://patents.google.com/patent/US20090214678A1/en.
Dugganaboyana GK, Kumar Mukunda C, Jain A, Kantharaju RM, Nithya RR, Ninganna D, Ahalliya RM, Shati AA, Alfaifi MY, Elbehairi SEI, Silina E, Stupin V, Velliyur Kanniappan G, Achar RR, Shivamallu C, Kollur SP (2023) Environmentally benign silver bionanomaterials as potent antioxidant, antibacterial, and antidiabetic agents: green synthesis using Salacia oblonga root extract. Front Chem 11:1114109
Espenti CS, Rao KSVK, Ramesh P, Sekhar AC, Rao KM (2020) Salacia mulbarica leaf extract mediated synthesis of silver nanoparticles for antibacterial and ct-DNA damage via releasing of reactive oxygen species. IET Nanobiotechnol 14(6):485–490
Feng ST, Wang ZZ, Yuan YH, Sun HM, Chen NH, Zhang Y (2019) Mangiferin: a multipotent natural product preventing neurodegeneration in Alzheimer’s and Parkinson’s disease models. Pharmacol Res 146:104336
Ghadage DM, Kshirsagar PR, Pai SR, Chavan JJ (2017) Extraction efficiency, phytochemical profiles and antioxidative properties of different parts of Saptarangi (Salacia chinensis L.) - an important underutilized plant. Biochem Biophys Rep 12:79–90
Gunaga SV, Vasudeva R (2019) Vegetative propagation through air-layering in Salacia macrosperma Wight: a threatened anti-diabetic plant of the Western Ghats. J Non-Timber for Prod 26(1):27–29
Jadhav K, Dhamecha D, Dalvi B, Patil M (2015) Green synthesis of silver nanoparticles using Salacia chinensis: characterization and its antibacterial activity. Part Sci Technol 33(5):445–455
Jadhav K, Rajeshwari HR, Deshpande S, Jagwani S, Dhamecha D, Jalalpure S, Subburayan K, Baheti D (2018) Phytosynthesis of gold nanoparticles: characterization, biocompatibility, and evaluation of its osteoinductive potential for application in implant dentistry. Mater Sci Eng C 93:664–670
Kamat SG, Vasudeva R, Patil CG (2021) Rapid mass propagation of endangered valuable medicinal plant Salacia chinensis L. and GCMS/LC-MS analysis of active compounds produced in callus and leaf extracts. Srilankan J Biol 6(2):1–13
Kaur P, Pandey DK, Gupta RC, Kumar V, Dwivedi P, Sanyal R, Dey A (2021) Biotechnological interventions and genetic diversity assessment in Swertia sp.: a myriad source of valuable secondary metabolites. Appl Microbiol Biotechnol 105:4427–4451
Kaur J, Verma M, Panda BP, Aeri V (2022) Enhancement of mangiferin by fungal endophytes isolated from Salacia chinensis L. and Salacia oblonga Wall. Indian J Exp Biol 60:781–788
Keshavamurthy M, Ravishankar Rai V (2021) Green synthesis, characterization and screening for antibacterial activity of gold nanoparticles produced by Salacia fruticosa leaf extract. Biosc Biotech Res Comm 14(2):879–885
Khan T, Ullah N, Khan MA, Mashwani ZR, Nadhman A (2019) Plant-based gold nanoparticles; a comprehensive review of the decade-long research on synthesis, mechanistic aspects and diverse applications. Adv Colloid Interface Sci 272:102017
Kshirsagar P, Umdale S, Chavan J, Gaikwad N (2017) Molecular authentication of medicinal plant, Swertia chirayita and its adulterant species. Proc Natl Acad Sci India Sect B Biol Sci 87(1):101–107
Kshirsagar PR, Mohite A, Suryawanshi S, Chavan JJ, Gaikwad NB, Bapat VA (2021) Plant regeneration through direct and indirect organogenesis, phyto-molecular profiles, antioxidant properties and swertiamarin production in elicitated cell suspension cultures of Swertia minor (Griseb.) Knobl. Plant Cell Tiss Org Cult 144:383–396
Kushwaha PS, Singh AK, Keshari AK, Maity S, Saha S (2016) An updated review on the phytochemistry, pharmacology, and clinical trials of Salacia oblonga. Pharmacogn Rev 10(20):109–114
Laxmi M, Raviraja Shetty G, Souravi K, Rajasekharan PE (2018) In-vitro conservation studies in Salacia chinensis L. a threatened medicinal plant. J Pharmacogn Phytochem SP3:78–81
Lin RY, Zhao KK, Wang HX, Zhu ZX, Wang HF (2019) Complete plastome sequence of Salacia amplifolia (Celastraceae): an endemic shrub in Hainan, China. Mitochondr DNA Part B 4(1):1977–1978
Mahendra C, Manasa G, Murali M, Sudarshana MS (2020a) Multiple shoot regeneration, establishment of callus and cell suspension cultures in Salacia macrosperma Wight. Biotechnol Res Innov 4(1):e2019053
Mahendra C, Murali M, Manasa G, Sudarshana MS (2020b) Biopotentiality of leaf and leaf derived callus extracts of Salacia macrosperma Wight - an endangered medicinal plant of Western Ghats. Ind Crops Prod 143:111921
Maheswari P, Chitra M, Usha Rani S (2013) Modification of protocol for isolation and purification of genomic DNA. Int J Eng Sci Math 2(2):235–247
Majid BN, Roopa G, Sampath KKK, Kini RK, Prakash HS, Abbagani S, Kiani M, Geetha N (2014) Establishment of an efficient explant surface sterilization protocol for in vitro micropropagation of Salacia chinensis L., an endangered anti-diabetic medicinal plant. World J Pharm Pharm Sci 3(12):1–10
Majid BN, Sampath KKK, Prakash HS, Geetha N (2016) Rapid mass propagation of Salacia chinensis L., an endangered valuable medicinal plant through direct organogenesis. Indian J Sci Technol 9(4):1–8
Majid B, Mehdi S, Prakash HS, Geetha N (2017) Establishment of an improved, efficient and ecofriendly micropropagation system in Salacia chinensis L. an endangered anti-diabetic medicinal plant. Agric For 63(3):167–176
Majid B, Mahesh MG, Saidi M, Kini KR, Prakash HS, Geetha N (2018) Evaluation of genetic stability using FRAPD markers as novel method along with antioxidant and anti-diabetic properties of micropropagated Salacia chinensis L. Acta Physiol Plant 40:128
Morikawa T, Ninomiya K, Tanabe G, Matsuda H, Yoshikawa M, Muraoka O (2021) A review of antidiabetic active thiosugar sulfoniums, salacinol and neokotalanol, from plants of the genus Salacia. J Nat Med 75:449–466
Mulye CS, Shirolkar AR, Dhar B, Murthy SN, Pawar SD (2013) RAPD-PCR analysis of Bixa orellana L. and Salacia chinensis L. to study genetic diversity. Int J Adv Biotechnol Res 4(3):380–383
Nagesh MR, Kumar N, Khan JM, Ahmed MZ, Kavitha R, Kim SJ, Vijayakumar N (2022) Green synthesis and pharmacological applications of silver nanoparticles using ethanolic extract of Salacia chinensis L. J King Saud Univ Sci 34:102284
Nandikar MD (2021) The genus Salacia (Celastraceae: Salaciodeae) in peninsular India. Webbia 76(2):167–193
Nayana EKE, Subasinghe S, Amarasinghe MKTK, Arunakumara KKIU, Kumarasinghe HKMS (2015) Effect of maturity and potting media on vegetative propagation of Salacia reticulata (Kothalahimbatu) through stem cuttings. Int J Min Fruit Med Aromat Plant 1(1):47–54
Paarakh PM, Patil LJ, Thanga SA (2008) Genus Salacia: a comprehensive review. J Nat Remedies 8(2):116–131
Patil S, Chandrasekaran R (2020) Biogenic nanoparticles: a comprehensive perspective in synthesis, characterization, application and its challenges. J Genet Eng Biotechnol 18:67
Priya G, Gopalakrishnan M, Rajesh E, Sekar T (2019) Assessment of species variability in Salacia using RAPD Markers. Int J Sci Res Rev 8(2):30–40
Rathnayaka RMSMB, Subasinghe S (2010) Development of bushy type Kothala Himbutu (Salacia reticulata White) plantation for sustainable leaf/stem harvesting. Proc 8th Acad Sessions Univ Ruhuna 8:206
Roopa G, Madhusudhan MC, Sunil KCR, Lisa N, Calvin R, Poornima R, Zeinab N, Kini KR, Prakash HS, Geetha N (2015) Identification of taxol-producing endophytic fungi isolated from Salacia oblonga through genomic mining approach. J Genet Eng Biotechnol 13(2):119–127
Roopa G, Madhusudhan MC, Majid BN, Sampath Kumara KK, Prakash HS, Geetha N (2017) Diversity of fungal endophytes from Salacia species in Western Ghats of Karnataka, India. Res J Pharm Biol Chem Sci 8(2):549–555
Roopa G, Madhusudhan M, Nischita R, Geetha N (2022) In vitro studies on anti-diabetic, anti-bacterial and phytochemical activities of endophytic fungal extracts from Salacia species. J Med Plant 10:31–37
Sabeena G, Vainath Praveen S, Pushpalakshmi E, Annadurai G (2023) Comparative synthesis and characterization of nanocomposites using chemical and green approaches including a comparison study on in vivo and in vitro biological properties. Nanoscale Adv 5:767–785
Sanyal R, Manokari M, Pandey S, Nandi S, Biswas P, Dewanjee S, Gopalakrishnan AV, Jha NK, Jha SK, Joshee N, Pandey DK, Dey A, Shekhawat MS (2023a) Biotechnological interventions and production of galanthamine in Crinum spp. Appl Microbiol Biotechnol 107:2155–2167
Sanyal R, Pandey S, Nandi S, Mondal R, Samanta D, Mandal S, Manokari M, Mishra T, Dhama K, Pandey DK, Shekhawat MS, Dey A (2023b) Biotechnology of Passiflora edulis: role of agrobacterium and endophytic microbes. Appl Microbiol Biotechnol 107:5651–5668
Sapkal A, Attar S, Chavan J, Kumbhar A, Kamble S (2023) Zingiber zerumbet: a green and ecofriendly natural surfactant for the synthesis of bis(indolyl)methane, tris-indoline and spirooxindole derivatives. Sustain Chem Pharm 35:101228
Sasidharan N, Muralidharan EM, Chacko KC (2010) Production of quality planting material of rare medicinal plants of commercial importance. https://docs.kfri.res.in/KFRI-RR/KFRI-RR380.pdf.
Savolainen V, Manen JF, Douzery E, Spichiger R (1994) Molecular phylogeny of families related to Celastrales based on rbcl 5ʹ flanking sequences. Mol Phylogenet Evol 3(1):27–37
Sheik S, Bhagya N, Chandrashekar KR (2020) Cytotoxic and decolorizing potential of Colletotrichum gloeosporioides Penz., isolated from Salacia chinensis Linn. S Afr J Bot 134:146–150
Simmons MP, Savolainen V, Clevinger CC, Archer RH, Davis JI (2001) Phylogeny of the Celastraceae inferred from 26S nuclear ribosomal DNA, phytochrome B, rbcL, atpB, and morphology. Mol Phylogenet Evol 19(3):353–366
Stierle A, Strobel G, Stierle D, Grothaus P, Bignami G (1995) The search for a taxol-producing microorganism among the endophytic fungi of the Pacific yew. Taxus Brevifolia J Nat Prod 58(9):1315–1324
Stohs SJ, Ray S (2015) Anti-diabetic and anti-hyperlipidemic effects and safety of Salacia reticulata and related species. Phytother Res 29(7):986–995
Webster G, Mullins AJ, Cunningham-Oakes E, Renganathan A, Aswathanarayan JB, Mahenthiralingam E, Vittal RR (2020) Culturable diversity of bacterial endophytes associated with medicinal plants of the Western Ghats, India. FEMS Microbiol Ecol 96(9):fiaa147
Zhu S, Liu Q, He J, Nakajima N, Samarakoon SP, Swe S, Zaw K, Komatsu K (2021) Genetic identification of medicinally used Salacia species by nrDNA ITS sequences and a PCR-RFLP assay for authentication of Salacia-related health foods. J Ethnopharmacol 274:113909
Funding
The first author received financial support by a research grant from the Department of Science and Technology (DST), Govt. of India, New Delhi, under the Young Scientist Fellowship Scheme (Registration No. SB/FT/LS-259/2012 dated 2 May 2013).
Author information
Authors and Affiliations
Contributions
JC and CS conceived and designed the research. PP, AP, AD, AM, PL, and PY conducted literature survey and data collection. MB, AK, DN, BP, and AJ contributed photographs and analytical tools. JC, MS, and CS analyzed data. JC wrote the manuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chavan, J., Patil, P., Patil, A. et al. Salacia spp.: recent insights on biotechnological interventions and future perspectives. Appl Microbiol Biotechnol 108, 200 (2024). https://doi.org/10.1007/s00253-023-12998-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00253-023-12998-z