
Abstract
n-Butanol represents a key commodity chemical and holds significant potential as a biofuel. It can be produced naturally by Clostridia species via the ABE pathway. However, butanol production via such systems can be associated with significant drawbacks. Therefore, substantial efforts have been made toward engineering a suitable industrial host for butanol production. For instance, we previously generated a metabolically engineered Saccharomyces cerevisiae strain that produces ~300 mg/L butanol from combined endogenous and exogenous pathways. In this current study, the endogenous and exogenous pathways of butanol production were further characterised, and their relative contribution to the overall butanol titre was assessed. Deletion of any single component of the exogenous ABE pathway was sufficient to significantly reduce butanol production. Further evidence for a major contribution from the ABE pathway came with the discovery that specific yeast deletion mutants only affected butanol production from this pathway and had a significant impact on butanol levels. In previous studies, the threonine-based ketoacid (TBK) pathway has been proposed to explain endogenous butanol synthesis in ADH1 mutants. However, we find that key mutants in this pathway have little impact on endogenous butanol production; hence, this pathway does not explain endogenous butanol production in our strains. Instead, endogenous butanol production appears to rely on glycine metabolism via an α-ketovalerate intermediate. Indeed, yeast cells can utilise α-ketovalerate as a supplement to generate high butanol titres (> 2 g/L). The future characterisation and optimisation of the enzymatic activities required for this pathway provides an exciting area in the generation of robust butanol production strategies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
n-Butanol (butanol) represents a key intermediate in the chemical industry with uses in the production of paints, adhesives, cosmetics, solvents and artificial flavouring. It also has wide potential as an alternative fuel to gasoline (Ndaba et al. 2015; Peralta-Yahya et al. 2012). For instance, in comparison with bioethanol, n-butanol is less hygroscopic, it is less corrosive, it has a higher energy content and it can be used at any ratio with gasoline, including directly as a fuel without engine or infrastructure modification (Cascone 2008).
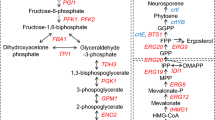
Traditionally, n-butanol has been produced from various Clostridia species via acetone–butanol-ethanol (ABE) fermentation (Lee et al. 2008). In the acidogenic exponential growth phase, acetate and butyrate are produced; then cells arrest growth and convert these acids to ethanol, butanol and acetone during a solventogenic phase (Patakova et al. 2013). The precise pathway for butanol production involves the condensation of two acetyl-CoA molecules to acetoacetyl-CoA followed by sequential reduction to n-butanol (Fig. 1). However, bacterial production can be associated with a range of difficulties, including butanol toxicity, bacteriophage contamination, the complexity of the two phase fermentation (acidogenesis and solventogenesis), sporulation during solventogenesis, high levels of by-products and very low titre at scale (Zheng et al. 2009; Hong and Nielsen 2012; Xue et al. 2013; Huffer et al. 2012). Therefore, a number of studies have focussed on producing butanol from alternative hosts, which may overcome some of the problems associated with the use of Clostridia species.

Endogenous and exogenous metabolic pathways for butanol production in S. cerevisiae. A schematic diagram of the exogenous acetone-butanol-ethanol (ABE) pathway enzymes, and the endogenous threonine-based ketoacid (TBK) and glycine-butanol (GB) pathways. Gene cassettes expressing components of the ABE fermentation pathway and upstream enzymes that have been inserted into the yeast genome are depicted: Hbd (3-hydroxybutyryl-CoA dehydrogenase), Crt (3-hydroxybutyryl-CoA dehydratase), Bcd (crotonyl-CoA reductase) and Adhe2 (alcohol dehydrogenase) from Clostridium beijerinckii, and Erg10p (thiolase), Ald6p (aldehyde dehydrogenase) and Acs2p (acetyl-CoA synthetase) from S. cerevisiae. The deletion of the yeast ADH1 gene is also depicted as a red cross, and relevant components of the endogenous pathways are highlighted, Leu1p (Isopropylmalate isomerase), Leu2p (β-isopropylmalate dehydrogenase) and Leu4p/Leu9p (α-isopropylmalate synthase)
A body of work has demonstrated that various strategies can be used to engineer yeast such that they produce n-butanol. The introduction of the Clostridial ABE pathway into yeast results in a low titre of butanol (Steen et al. 2008; Swidah et al. 2015), but additional modifications can improve this titre. For instance, levels of butanol can be increased using strategies to increase the level of cytosolic acetyl-CoA. For instance, overexpression of the machinery involved in converting ethanol to acetyl-CoA (the ALD6, ACS2 and ADH2 genes), or deletion of the glyoxylate cycle components that normally deplete cytosolic acetyl-CoA (CIT2 and MLS1) significantly improve butanol production (Krivoruchko et al. 2013; Swidah et al. 2015). Another strategy to improve butanol titre involves the restriction of competing pathways and restoration of redox balance caused by the introduction of the ABE pathway. Hence, deletion of the alcohol and glycerol dehydrogenase genes involved in ethanol and glycerol biosynthesis respectively has been shown to improve butanol titre (Lian et al. 2014; Schadeweg and Boles 2016; Swidah et al. 2015).
While studying butanol production from exogenous Clostridial pathways in yeast, it has become clear that some of the modifications designed to improve titre, actually facilitated butanol production in the absence of an exogenous pathway. For instance, deletion of ADH1, a gene encoding the major alcohol dehydrogenase for ethanol production activates an endogenous butanol production pathway (Si et al. 2014; Swidah et al. 2015). It has been proposed that this pathway stems from threonine catabolism in mitochondria and borrows enzymes from the leucine biosynthetic pathway (Si et al. 2014); hence, it has been termed the threonine-based ketoacid (TBK) pathway (Shi et al. 2016) (Fig. 1). Indeed, overexpression of appropriate pathway genes and elimination of competing pathways further improved the butanol titre (Si et al. 2014; Shi et al. 2016). Intriguingly, the existence of an endogenous butanol pathway connected with hydrophobic amino acid metabolism was suggested as early as the 1960s (Ingraham et al. 1961). The combination of endogenous and exogenous pathways within butanol producer strains likely represents a fundamental strategy for the further enhancement of butanol titre (Shi et al. 2016; Swidah et al. 2015).
Another pathway that has been studied in yeast with a view to butanol production is a glycine-butanol (GB) pathway. Here, butanol is produced when glycine is used as a substrate in the fermentation media (Branduardi et al. 2013). Once again, a combination of exogenous and endogenous enzymes improves butanol production, as introduction of goxB from B. subtilis enhances the conversion of glycine into glyoxylate, which is subsequently converted to butanol in a series of reactions (Fig. 1).
In previous work, we have engineered a strain of S. cerevisiae for n-butanol production. We showed that the butanol is synthesised in parallel from an endogenous butanol synthetic pathway activated by ADH1 deletion and a heterologous ABE synthetic pathway. Combining these pathways with a strategy to channel the carbon flux toward acetyl-CoA production by overexpressing ALD6 ACS2 further enhanced titre (Swidah et al. 2015). In this current study, we have characterised the relative contribution of the endogenous and exogenous pathways involved in n-butanol production in these yeast strains. We present various experiments suggesting that the exogenous pathway is responsible for much of the butanol production in our strains. Endogenous butanol production that is activated by ADH1 deletion does not rely upon leucine biosynthetic enzymes and so the threonine-based ketoacid (TBK) pathway is unlikely to play a role. In attempts to characterise potential routes for endogenous butanol production, different substrates were added to the fermentation media. Butanol titres were increased when glycine was used as a supplement, and exceeded 2400 mg/L when α-ketovalerate was supplied. Therefore, yeast harbour the enzymatic machinery required for butanol production, and the characterisation and subsequent optimisation of this machinery provides intriguing avenues for further research into yeast as a cellular biofuel factory.
Methods
Yeast growth and engineering
All the engineered yeast used in this work were derived from S. cerevisiae W303-1A (ATCC: MYA-2181). Yeast were grown at 30 °C on standard yeast extract/peptone/dextrose (2% w/v) media (YPD) (Guthrie and Fink 1991). In the supplementation experiments, α-ketovalerate (Sigma-Aldrich) and glycine were added to YPD at 0.2%. For α-ketovalerate, the media was buffered at pH 5.0 using a citrate/phosphate buffer. The MLS1 and CIT2 genes were deleted using standard yeast PCR-based gene disruption methods and the loxp-natNT2-loxp marker from the pZC2 vector (Carter and Delneri 2010). Subsequent removal of the natNT2 marker was achieved using a cre recombinase strategy, which allowed construction of the double mls1Δcit2Δ mutants. The LEU2 prototroph was generated by transformation with an SspI 1.5-kb fragment LEU2 fragment isolated from the pRS vector. The LEU1 and LEU4 genes were deleted using the loxp-natNT2-loxp cassette using standard yeast PCR-based gene disruption methods. Genotype alterations were verified by PCR analysis on genomic DNA and assessment of known phenotypic consequences.
Measurements of butanol and ethanol
Yeast were grown as semi-anaerobic batch cultures from a starting OD600 of 0.1 in liquid YPD media using 50-ml vials over a 21-day period. Media samples were taken, passed through a 0.22-μ filter into gas chromatography (GC) vials and analysed by GC-FID using an Agilent 6850A GC system with an Agilent 4513A automatic injector, sampler and controller. A DB-WAX capillary column (30 m × 0.25 mm, 0.25 μM) was used for separation. Samples were collected every 2 days and quantified relative to standards of ethanol and butanol. Where specific values are shown, levels on days 11 or 13 were used.
Western blot analysis of flag-tagged proteins
Yeast cultures (5 ml) were grown to an OD600 of 0.7 in YPD, pelleted; then protein samples were prepared and processed for electrophoresis and immunoblot analysis as described previously (Ashe et al. 2000) using an Anti-Flag antibody (Sigma-Aldrich).
Results
The ABE pathway explains a major part of the butanol production in engineered S. cerevisiae
Previously, we have described an engineered version of W303-1A (termed A6A2adh1Δ + 5g) that produces substantial levels of butanol. In this strain, five butanol production enzymes (four from Clostridia—Hbd, Crt, Bcd and Adhe2—plus one from S. cerevisiae—Erg10p) and the enzymes for cytosolic acetyl-CoA synthesis (Ald6p and Acs2p) are overexpressed, while the major alcohol dehydrogenase ADH1 is deleted (Fig. 1). Intriguingly, the ADH1 deletion by itself, without other strain alterations, leads to low level butanol production (Swidah et al. 2015). This finding is consistent with what has been shown by other investigators and is suggestive that endogenous pathways for butanol production exist in yeast (Si et al. 2014). The deletion of ADH1 also causes a range of physiological alterations including significant reductions in ethanol production, the accumulation of acetaldehyde and acetate and a decrease in cellular growth rate (de Smidt et al. 2012; Swidah et al. 2015). The presence of the butanol production enzymes in an adh1Δ strain led to minor restorative effects for some of these phenotypes: for instance, the growth rate was slightly improved (Swidah et al. 2015). This means it is formally possible that the enzymes of the heterologous butanol production pathway are not responsible for butanol production per se. Instead, the endogenous pathway that produces low levels of butanol in the adh1 deletion strain is somehow enhanced by the presence of these enzymes: perhaps simply by improving the growth of the strain. Therefore, a key question is whether the heterologous ABE pathway that has been introduced into S. cerevisiae is active and how much it contributes to butanol levels relative to potential endogenous butanol production pathways. To address this, a genetic strategy was taken where the effect of having four of the five butanol production pathway genes was assessed in strains carrying the ADH1 deletion and overexpressing ALD6 and ACS2.
Evaluation of the expression levels of the various enzymes in these strains confirmed that all of butanol synthetic enzymes as well as Ald6p and Acs2p enzymes were highly expressed in the parent strain (A6A2adh1Δ+5g) (Fig. 2a). In contrast, expression of one of the enzymes (Crt, Bcd, Erg10p, Hbd or Adhe2) was deficient in the appropriate deletion strain (Fig. 2a). As described previously for the A6A2adh1Δ+5g production strain, ~300–400 mg/L butanol was produced over the course of a fermentation, whereas the adh1Δ strain produced < 100 mg/L butanol. In all of the strains carrying four of the five butanol production pathway enzymes, the levels of butanol produced were similar or less than that produced by the adh1Δ strain (Fig. 2c). Interestingly, across the fermentations for the constructed mutants, the levels of ethanol are higher compared to the parental A6A2adh1Δ+5g strain or the adh1Δ strain (Fig. 2b). It is possible that in the absence of the butanol production pathway, any acetate, acetaldehyde or acetyl-CoA that accumulate due to the deletion of ADH1 and overexpression of ALD6/ACS2 gets converted to ethanol over the course of the fermentation, whereas the presence of an active butanol production pathway provides a driving force minimising back flux to ethanol.

The exogenous ABE pathway is a major factor defining butanol levels in the butanol production strain. a Western blot using an anti-Flag antibody to detect protein expression from protein extracts prepared from the indicated strains. The butanol production strain—A6A2adh1∆ + 5 g (carrying the five enzymes of the ABE pathway, overexpressing Ald6/Acs2p and deleted for ADH1), the wild type parent (WT) and the A6A2adh1∆ strain (overexpressing Ald6/Acs2p and deleted for ADH1) are used as controls. They are compared to five strains derived from the production strain, but each lacking one of the proteins from the ABE pathway (-crt, -bcd, -ERG10, -hbd1 and -adhe2). Proteins derived from the five integrated cassettes of the ABE pathway and the integrated cassette overexpressing Acs2p and Ald6p are labelled on the right. Bullets represent the position where the expected protein is absent from the blot. b, c Graphs depicting the level of ethanol or butanol from the constructed strains on day 14 using semi-anaerobic fermentation conditions. Error bars are ± SD from 5 biological repeats
Overall, these data suggest that in the parent strain producing high levels of butanol, the heterologous butanol production pathway is active, all five components of the pathway are required for activity and the pathway explains a large proportion of the butanol synthesised by the strain. Residual butanol production is apparent and this presumably stems from an endogenous butanol production pathway.
Mutations to the glyoxylate shunt pathway inhibit the heterologous butanol production pathway without affecting the endogenous pathway
During the course of studies aimed at optimising butanol synthesis from the butanol production strain A6A2adh1Δ+5g, further evidence was obtained supporting the existence of both heterologous and endogenous butanol production pathways. Initially, we reasoned that increasing cytosolic acetyl-CoA might lead to higher butanol levels from our production strain. Previous studies have shown deleting the glyoxylate shunt genes, MLS1 and CIT2, encoding acetyl-CoA depleting components (Fig. 3a), leads to greater butanol production in the context of strains overexpressing a number of other yeast and Clostridial genes (Krivoruchko et al. 2013). Therefore, MLS1 and CIT2 were deleted singly or in combination in both our butanol production strain and the adh1Δ strain. However, in the context of our butanol production strain A6A2adh1Δ+5g, no improvement in butanol titre was evident in the deletions. In fact, the deletions caused a large reduction in butanol titre (Fig. 3b). Notably though, in the adh1Δ strain where the source of butanol is an endogenous pathway, the deletions in CIS2 or MLS1 had no impact on butanol levels (Fig. 3c). Equally, the deletions reduce butanol production in the A6A2adh1Δ+5g strain to the level observed in the adh1Δ strain, suggesting that the endogenous pathway is still active in this strain. Therefore, these mutant data are suggestive that there are two distinct butanol pathways, the heterologous butanol production pathway that is sensitive to alterations to the glyoxylate shunt pathway, and the endogenous pathway that is insensitive to these gene deletions.

Acetyl-CoA consuming reactions from the Glyoxylate cycle impact on the exogenous but not the endogenous butanol pathway. a Schematic diagram of the glyoxylate cycle in yeast focussing on acetyl-CoA and reactions that impact upon its levels in the cytosol and peroxisome (Hiltunen et al. 2003). The reactions targeted in the deleted strategy to increase the pool of acetyl-CoA are marked with a red cross b, c Bar charts show butanol levels for single and double deletions of MLS1 and CIT2 in either the butanol production strain (A6A2adh1∆ + 5g) or the adh1∆ strain from semi-anaerobic fermentations
The question as to why deletion of glyoxylate shunt components in our A6A2adh1Δ+5g producer strain reduces butanol production, specifically from the heterologous pathway, while in other strain contexts it activates butanol production (Krivoruchko et al. 2013), likely reflects the distinct context in the relative strains. In our strain, flux toward ethanol has been inhibited by deletion of ADH1, which likely leads to the accumulation of toxic intermediates such as acetaldehyde and acetate. Deletion of MLS1 and CIT2 might further exacerbate this effect by stemming depletion of cytosolic acetyl-CoA, which may reduce both growth and butanol production. Indeed, the accumulation of biomass was somewhat delayed in the MLS1/CIT2 deletion strains relative to the parent (data not shown). In contrast, in studies where deletion of MLS1 and CIT2 improved butanol titre (Krivoruchko et al. 2013), ADH2 was overexpressed to drive production of acetaldehyde from ethanol. It is unlikely that such overexpression in the context of an active ADH1 gene (which catalyses the reverse reaction) would lead to the same accumulation of acetaldehyde and acetate. Therefore, in this strain, the MLS1 and CIT2 deletions may have the desired effect of leading to accumulation of cytosolic acetyl-CoA: the substrate for the butanol pathway.
A previously characterised endogenous butanol pathway does not explain the butanol accumulation
S. cerevisiae can accumulate up to 120 mg/L n-butanol after deletion of the ADH1 gene (Si et al. 2014; Swidah et al. 2015). Previous studies have suggested that an endogenous threonine-based ketoacid (TBK) pathway is involved in this butanol production, where threonine catabolism to 2-ethylmalate in the mitochondria precedes cytosolic conversion to n-butanol through the action of various components of the leucine biosynthetic pathway (Si et al. 2014; Shi et al. 2016). The leucine biosynthetic genes Leu1p, Leu2p, Leu4p and Leu9p form part of this native pathway (Fig. 1).
Previously, we found that an adh1Δ mutant strain produces n-butanol in the absence of other alterations, even though the strain is a leucine auxotroph carrying the leu2-3,112 mutant allele (Swidah et al. 2015). Leu2p is a β-isopropylmalate dehydrogenase that is also capable of converting 3-ethylmalate to α-ketovalerate as part of a threonine-based ketoacid pathway (Shen and Liao 2008; Si et al. 2014; Branduardi et al. 2013; Andreadis et al. 1984). In order to assess whether this reaction was important for the endogenous pathway in our strains and if so, to improve the efficiency of butanol production, we undertook a strategy to rescue the LEU2 gene in the adh1Δ strain background. In order to generate this LEU2 prototrophic strain, a LEU2 rescue fragment was isolated, transformed into the adh1Δ strain, screened on selective media and validated for integration at the endogenous locus. Intriguingly, rescuing LEU2 in the adh1Δ strain had no impact on n-butanol or ethanol production (Fig. 4a, b). However, it seemed plausible that in our adh1Δ strain another enzyme could be catalysing this reaction in the absence of Leu2p. Interestingly, previous authors have also speculated that residual activity in a LEU2 deletion strain could be due to other enzymes (Branduardi et al. 2013).

Elements of the Leu pathway are not involved in the endogenous pathway of butanol production. A. & C. Graphs depicting the level of ethanol and butanol after rescuing the LEU2 gene in adh1∆ strain over 18 day semi-anaerobic fermentations. D. & E. Graphs depicting the level of ethanol and butanol after deleting LEU1 in adh1∆ strain and butanol production strain (A6A2adh1∆ + 5 g) over a 18 days using semi-anaerobic fermentation conditions. Error bars are ±SD from 5 biological repeats
A prediction of this would be that deletion of other leucine biosynthetic genes in the threonine-based ketoacid pathway, such as LEU1, would block butanol production from the endogenous pathway. This would be expected to decrease the n-butanol titre considerably for the adh1Δ strain and only slightly for A6A2adh1Δ+5g production strain. Leu1p is a cytosolic FeS dehydratase that can catalyse the conversion of 2-ethylmalate into 3-ethylmalate, as well as the second step in the leucine biosynthetic pathway (Skala et al. 1991). Although, the mitochondrial aconitase and homoaconitase enzymes of the TCA cycle and lysine biosynthetic pathway, respectively, are from the same family as Leu1p they have very distinct substrate characteristics and subcellular localization (Lill et al. 1999). Therefore, the LEU1 gene was deleted in the adh1Δ, A6A2adh1Δ+5g and parent strains, and the resulting strains were confirmed as leucine auxotrophs. Surprisingly, deletion of LEU1 did not decrease ethanol or n-butanol levels for the leu1Δadh1Δ strain compared to the parent adh1Δ strain (Fig. 4c). Equally, LEU1 deletion had little effect on titre from the A6A2adh1Δ+5g strain (Fig. 4d). Similar observations were made when the LEU4 gene was deleted (data not shown). Leu4p is the major isozyme responsible for the first step in the leucine biosynthesis pathway and can convert α-ketobutyrate into 2-ethylmalate in the mitochondria (Drain and Schimmel 1988; Baichwal et al. 1983; Beltzer et al. 1988). Our results suggest that specific enzymes of the leucine biosynthetic pathway are not required for butanol production from an endogenous pathway that is activated after deletion of ADH1.
α-Ketovalerate is efficiently converted to n-butanol
Two butanol production pathways have been described where much or the vast majority of the pathway relies upon endogenous enzymes, the TBK pathway described above and the glycine-butanol pathway (Si et al. 2014; Shi et al. 2016; Branduardi et al. 2013). A common element to both of these pathways is the conversion of α-ketovalerate to n-butanol. This conversion would require an α-keto acid decarboxylase (KDC) to generate butyraldehyde and an alcohol dehydrogenase to give n-butanol (Romagnoli et al. 2012). In order to assess whether in our strains enzymes that would enable such a conversion are active, we added α-ketovalerate to the fermentation media. While all of the strains tested produced expected levels of ethanol in rich media (YPD) and rich media with α-ketovalerate (Fig. 5a, b), the addition of α-ketovalerate not only improves butanol production for the strain lacking ADH1 (up to 140 mg/L for adh1Δ and 600 mg/L for A6A2adh1Δ+5g production strain), but also leads to a marked improvement for the strains harbouring functional ADH1: the WT produces (1.4 g/L), whereas the strain bearing 5 butanol synthetic genes produces (2.4 g/L). It is entirely possible that the variation in butanol levels for these strains is relevant to the presence of the ABE pathway in the butanol production strain. For instance, strains carrying the ABE pathway contain the Adhe2 enzyme, which could participate in boosting the butanol titre compared to the parent strain.

Using α-ketovalerate or glycine in the growth media significantly improves butanol production. Bar charts depict the results of fermentation experiments in YPD medium with and without 0.2% α-ketovalerate (a, b) or 0.2% glycine (c, d) as indicated. The level of ethanol (A and C) or butanol (b, d) was quantified on day 13 of a standard semi-anaerobic fermentation. Error bars are ± SD from 5 biological repeats. For selected comparisons * represents a difference where p < 0.05
These results not only highlight that the enzymes responsible for converting α-ketovalerate to n-butanol are present in yeast, but that there is potential for increasing butanol titre significantly by targeting endogenous pathways of butanol production, particularly in combination with exogenous pathways.
Glycine is a likely source of endogenous n-butanol
Previous studies have shown that S. cerevisiae strains are able to accumulate n-butanol from glycine as a nitrogen source (Branduardi et al. 2013). To enable slightly higher levels of production, a Bacillus subtilis glycine oxidase (goxB) gene was introduced into yeast to enhance the conversion of glycine to glyoxylate, which could then be converted to n-butanol via α-ketovalerate (Branduardi et al. 2013) (Fig. 1). In yeast, generally, glycine is synthesised from serine, which stems from glycolytic intermediates (Hahn 1993). Given the potential for a glycine-butanol (GB) pathway, we decided to explore the impact of using glycine as substrate on n-butanol production in our system. Interestingly, for the mutants bearing only the native butanol production pathway (adh1Δ and adh1ΔLEU2), we found that the addition of glycine improves n-butanol production significantly (40 mg/L to 120 mg/L). The levels of n-butanol were also slightly increased for the A6A2adh1Δ+5g production strain bearing both exogenous and endogenous pathways, although this increase was not statistically significant (Fig. 5c, d).
Discussion
Previously, we have engineered the S. cerevisiae W303-1A strain with a view to the production of the key commodity chemical and biofuel, n-butanol (Swidah et al. 2015). Here, we show that two parallel pathways contribute to this production: an endogenous pathway activated after deletion of the major alcohol dehydrogenase, ADH1, and an exogenous ABE-based pathway, which together have the capacity to produce 300–380 mg/L butanol.
Using genetic analysis, we show that the heterologous ABE pathway is producing the majority of the butanol in the production strain. Firstly and most significantly, deletion of any single gene from the ABE pathway is entirely sufficient to reduce butanol levels to the base level produced when ADH1 alone is deleted. Second mutations in the glyoxylate pathway impact upon production via the ABE pathway enzymes without effecting production from an ADH1 deletion strain. These data combined strongly suggest that two parallel butanol production pathways are present and active within our strains. This agrees with other recent studies highlighting the value in terms of butanol titre in combining different strategic approaches for butanol production within a single strain (Shi et al. 2016).
The presence of an endogenous pathway within our adh1Δ strain, however, presented a conundrum based upon other studies with adh1Δ mutant strains (Si et al. 2014). As seemingly, the most likely route for butanol production is via the threonine-based ketoacid pathway involving a number of leucine biosynthetic enzymes including Leu2p (Si et al. 2014; Ingraham et al. 1961), whereas our adh1Δ strain carries a non-reverting auxotrophic allele of the LEU2 gene (Hinnen et al. 1978). We considered that perhaps another activity might be compensating for the Leu2p enzyme in our yeast strains. With this in mind, we replaced the auxotrophic LEU2 allele with the wild type expecting an improvement in butanol levels. However, no such improvement was evident. Equally, deletion of either the LEU1 or LEU4 genes higher in the pathway than LEU2 did not reduce butanol production in any way. On this basis, we conclude that the TBK pathway is not involved in butanol production in our adh1Δ strain.
Another study has described butanol production from glycine (Branduardi et al. 2013). In this study, the hypothesis was put forward that glycine could be converted to butanol via conversion to glyoxylate, followed by glyoxylate condensation with butyryl-CoA to 3-ethylmalate and 3-ethylmalate conversion to α-ketovalerate then on to n-butanol. Interestingly, using α-ketovalerate as a precursor for butanol production in the fermentation media not only leads to improved butanol titre in all our metabolically engineered strains but also a non-engineered parental strain produced up to 2 g/L. This result shows that S. cerevisiae actively express enzymes capable decarboxylating and reducing α-ketovalerate to n-butanol. Previous work has suggested that a range of 2-ketoacid decarboxylase enzymes including the pyruvate decarboxylase enzymes (Pdc1p, Pdc5p or Pdc6p) would be capable of undertaking the decarboxylation reaction and a number of alcohol dehydrogenases could convert butyraldehyde to n-butanol (Romagnoli et al. 2012; Larroy et al. 2002, b). Further evidence that a pathway from glycine might be the source of n-butanol in our strain stems from the supplementation of the yeast fermentation media with glycine. Here, a 3-fold increase in the level of butanol was observed. However, the interpretation that a glycine-butanol pathway is responsible for butanol production in the adh1Δ strain is difficult to reconcile with a number of other observations. For instance, the Leu2p enzyme is once again thought to play a key role, catalysing the conversion of 3-ethylmalate to α-ketovalerate. Yet our strain is auxotrophic for the Leu2p enzyme, and when we complement the LEU2 auxotrophic mutations, no effect on butanol levels was observed. However, in other studies, following deletion of the LEU2 gene, the resulting yeast strains still harboured at least some capacity to carry out this enzymatic reaction (Branduardi et al. 2013). Therefore, it might be possible that a redundant enzyme could catalyse this reaction. Another issue with this pathway is that we have deleted the malate synthase gene, MLS1, and found that the adh1Δ strain still produces similar levels of butanol. Mls1p catalyses the condensation of butyryl-CoA with glyoxylate to give 3-ethylmalate downstream of glycine. Once again, redundancy may represent an answer, as Dal7p also catalyses the same reaction and might compensate for the lack of Mls1p. A final concern is that while several studies suggest the existence of a yeast enzyme that is capable of converting glycine into glyoxylate (Branduardi et al. 2013; Villas-Boas et al. 2005), it has not yet been identified. In previous studies, a glyoxylate oxidase from Bacillus subtilis was used to bolster this reaction, but without knowing the precise gene involved it is difficult to prove a role for the reaction.
In conclusion, we provide evidence that in our engineered yeast butanol production strains, two parallel pathways are important for butanol synthesis. Firstly, the introduced heterologous ABE pathway is responsible for a large part of the butanol produced. Secondly, an endogenous pathway activated by deletion of the ADH1 gene is also responsible for butanol generation. In the interest of potentially optimising this endogenous pathway further, we sought to characterise it. Of the two previously described pathways for butanol production, our studies favour a route that stems from glycine and goes via an α-ketovalerate intermediate. Future optimisation of systems where different pathways converge on the same end product holds significant promise in the era of synthetic biotechnology.
References
Andreadis A, Hsu YP, Hermodson M, Kohlhaw GB, Schimmel P (1984) Yeast LEU2. Repression of mRNA levels by leucine and primary structure of the gene product. J Biol Chem 259:8059–8062
Ashe MP, De Long SK, Sachs AB (2000) Glucose depletion rapidly inhibits translation initiation in yeast. Mol Biol Cell 11:833–848
Baichwal VR, Cunningham TS, Gatzek PR, Kohlhaw GB (1983) Leucine biosynthesis in yeast: identification of two genes (LEU4, LEU5) that affect alpha-isopropylmalate synthase activity and evidence that LEU1 and LEU2 gene expression is controlled by alpha-Isopropylmalate and the product of a regulatory gene. Curr Genet 7:369–377
Beltzer JP, Morris SR, Kohlhaw GB (1988) Yeast LEU4 encodes mitochondrial and nonmitochondrial forms of alpha-isopropylmalate synthase. J Biol Chem 263:368–374
Branduardi P, Longo V, Berterame NM, Rossi G, Porro D (2013) A novel pathway to produce butanol and isobutanol in Saccharomyces cerevisiae. Biotechnol Biofuels 6:68
Carter Z, Delneri D (2010) New generation of loxP-mutated deletion cassettes for the genetic manipulation of yeast natural isolates. Yeast 27:765–775
Cascone R (2008) Biobutanol - a replacement for bioethanol? Chem Eng Prog 104:S4–S9
de Smidt O, du Preez JC, Albertyn J (2012) Molecular and physiological aspects of alcohol dehydrogenases in the ethanol metabolism of Saccharomyces cerevisiae. FEMS Yeast Res 12:33–47
Drain P, Schimmel P (1988) Multiple new genes that determine activity for the first step of leucine biosynthesis in Saccharomyces cerevisiae. Genetics 119:13–20
Guthrie C, Fink GR (1991) Guide to yeast genetics and molecular biology (Academic Press: San Diego, California)
Hahn RG (1993) Dose-dependent half-life of glycine. Urol Res 21:289–291
Hiltunen JK, Mursula AM, Rottensteiner H, Wierenga RK, Kastaniotis AJ, Gurvitz A (2003) The biochemistry of peroxisomal beta-oxidation in the yeast Saccharomyces cerevisiae. FEMS Microbiol Rev 27:35–64
Hinnen A, Hicks JB, Fink GR (1978) Transformation of yeast. Proc Natl Acad Sci U S A 75:1929–1933
Hong KK, Nielsen J (2012) Metabolic engineering of Saccharomyces cerevisiae: a key cell factory platform for future biorefineries. Cell Mol Life Sci 69:2671–2690
Huffer S, Roche CM, Blanch HW, Clark DS (2012) Escherichia coli for biofuel production: bridging the gap from promise to practice. Trends Biotechnol 30:538–545
Ingraham JL, Crowell EA, Guymon JF (1961) Pathway of formation of N-butyl and N-amyl alcohols by a mutant strain of Saccharomyces cerevisiae. Arch Biochem Biophys 95:169–175
Krivoruchko A, Serrano-Amatriain C, Chen Y, Siewers V, Nielsen J (2013) Improving biobutanol production in engineered Saccharomyces cerevisiae by manipulation of acetyl-CoA metabolism. J Ind Microbiol Biotechnol 40:1051–1056
Larroy C, Fernandez MR, Gonzalez E, Pares X, Biosca JA (2002) Characterization of the Saccharomyces cerevisiae YMR318C (ADH6) gene product as a broad specificity NADPH-dependent alcohol dehydrogenase: relevance in aldehyde reduction. Biochem J 361:163–172
Larroy C, Pares X, Biosca JA (2002) Characterization of a Saccharomyces cerevisiae NADP (H)-dependent alcohol dehydrogenase (ADHVII), a member of the cinnamyl alcohol dehydrogenase family. Eur J Biochem 269:5738–5745
Lee SY, Park JH, Jang SH, Nielsen LK, Kim J, Jung KS (2008) Fermentative butanol production by Clostridia. Biotechnol Bioeng 101:209–228
Lian J, Si T, Nair NU, Zhao H (2014) Design and construction of acetyl-CoA overproducing Saccharomyces cerevisiae strains. Metab Eng 24:139–149
Lill R, Diekert K, Kaut A, Lange H, Pelzer W, Prohl C, Kispal G (1999) The essential role of mitochondria in the biogenesis of cellular iron-sulfur proteins. Biol Chem 380:1157–1166
Ndaba B, Chiyanzu I, Marx S (2015) N-butanol derived from biochemical and chemical routes: a review. Biotechnol Rep (Amst) 8:1–9
Patakova P, Linhova M, Rychtera M, Paulova L, Melzoch K (2013) Novel and neglected issues of acetone-butanol-ethanol (ABE) fermentation by clostridia: Clostridium metabolic diversity, tools for process mapping and continuous fermentation systems. Biotechnol Adv 31:58–67
Peralta-Yahya PP, Zhang F, del Cardayre SB, Keasling JD (2012) Microbial engineering for the production of advanced biofuels. Nature 488:320–328
Romagnoli G, Luttik MA, Kotter P, Pronk JT, Daran JM (2012) Substrate specificity of thiamine pyrophosphate-dependent 2-oxo-acid decarboxylases in Saccharomyces cerevisiae. Appl Environ Microbiol 78:7538–7548
Schadeweg V, Boles E (2016) n-Butanol production in Saccharomyces cerevisiae is limited by the availability of coenzyme A and cytosolic acetyl-CoA. Biotechnol Biofuels 9:44
Shen CR, Liao JC (2008) Metabolic engineering of Escherichia coli for 1-butanol and 1-propanol production via the keto-acid pathways. Metab Eng 10:312–320
Shi S, Si T, Liu Z, Zhang H, Ang EL, Zhao H (2016) Metabolic engineering of a synergistic pathway for n-butanol production in Saccharomyces cerevisiae. Sci Rep 6:25675
Si T, Luo Y, Xiao H, Zhao H (2014) Utilizing an endogenous pathway for 1-butanol production in Saccharomyces cerevisiae. Metab Eng 22:60–68
Skala J, Capieaux E, Balzi E, Chen WN, Goffeau A (1991) Complete sequence of the Saccharomyces cerevisiae LEU1 gene encoding isopropylmalate isomerase. Yeast 7:281–285
Steen EJ, Chan R, Prasad N, Myers S, Petzold CJ, Redding A, Ouellet M, Keasling JD (2008) Metabolic engineering of Saccharomyces cerevisiae for the production of n-butanol. Microb Cell Factories 7:36
Swidah R, Wang H, Reid PJ, Ahmed HZ, Pisanelli AM, Persaud KC, Grant CM, Ashe MP (2015) Butanol production in S. cerevisiae via a synthetic ABE pathway is enhanced by specific metabolic engineering and butanol resistance. Biotechnol Biofuels 8:97
Villas-Boas SG, Kesson M, Nielsen J (2005) Biosynthesis of glyoxylate from glycine in Saccharomyces cerevisiae. FEMS Yeast Res 5:703–709
Xue C, Zhao XQ, Liu CG, Chen LJ, Bai FW (2013) Prospective and development of butanol as an advanced biofuel. Biotechnol Adv 31:1575–1584
Zheng YN, Li LZ, Xian M, Ma YJ, Yang JM, Xu X, He DZ (2009) Problems with the microbial production of butanol. J Ind Microbiol Biotechnol 36:1127–1138
Funding
This work was largely funded by a Biotechnology and Biological Sciences Research Council (BBSRC) project grant (BB/K002767/1). OO was supported by the Tertiary Education Trust Fund Nigeria.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Swidah, R., Ogunlabi, O., Grant, C.M. et al. n-Butanol production in S. cerevisiae: co-ordinate use of endogenous and exogenous pathways. Appl Microbiol Biotechnol 102, 9857–9866 (2018). https://doi.org/10.1007/s00253-018-9305-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-018-9305-x