
Abstract
In bacteria, adaptation to changing environmental conditions is often mediated by two-component signal transduction systems. In the prototypical case, a specific stimulus is sensed by a membrane-bound histidine kinase and triggers autophosphorylation of a histidine residue. Subsequently, the phosphoryl group is transferred to an aspartate residue of the cognate response regulator, which then becomes active and mediates a specific response, usually by activating and/or repressing a set of target genes. In this review, we summarize the current knowledge on two-component signal transduction in Corynebacterium glutamicum. This Gram-positive soil bacterium is used for the large-scale biotechnological production of amino acids and can also be applied for the synthesis of a wide variety of other products, such as organic acids, biofuels, or proteins. Therefore, C. glutamicum has become an important model organism in industrial biotechnology and in systems biology. The type strain ATCC 13032 possesses 13 two-component systems and the role of five has been elucidated in recent years. They are involved in citrate utilization (CitAB), osmoregulation and cell wall homeostasis (MtrAB), adaptation to phosphate starvation (PhoSR), adaptation to copper stress (CopSR), and heme homeostasis (HrrSA). As C. glutamicum does not only face changing conditions in its natural environment, but also during cultivation in industrial bioreactors of up to 500 m3 volume, adaptability can also be crucial for good performance in biotechnological production processes. Detailed knowledge on two-component signal transduction and regulatory networks therefore will contribute to both the application and the systemic understanding of C. glutamicum and related species.
Similar content being viewed by others


Introduction
Corynebacterium glutamicum is a Gram-positive, facultative anaerobic, nonpathogenic soil bacterium which is used for the large-scale industrial production of the flavor enhancer l-glutamate (2.2 million tons in 2009) and the food additive l-lysine (1.5 million tons in 2011). Recent metabolic engineering studies have shown that C. glutamicum is also capable of producing a variety of other commercially interesting compounds, e.g. other l-amino acids (Wendisch et al. 2006a), d-amino acids (Stäbler et al. 2011), organic acids such as succinate (Okino et al. 2008; Litsanov et al. 2012a, b, c), diamines such as cadaverine (Mimitsuka et al. 2007) or putrescine (Schneider and Wendisch 2010), biofuels such as ethanol or isobutanol (Inui et al. 2004; Smith et al. 2010; Blombach et al. 2011), or proteins (Meissner et al. 2007). An overview of the product spectrum of C. glutamicum can be found in a recent review (Becker and Wittmann 2011). Due to its function as microbial cell factory, C. glutamicum has become a prominent model organism in industrial biotechnology and simultaneously for systems biology (Eggeling and Bott 2005; Wendisch et al. 2006b; Burkovski 2008). Another important aspect fostering research on C. glutamicum is its close phylogenetic relationship to important pathogens, in particular Mycobacterium tuberculosis (Ciccarelli et al. 2006). Both Corynebacteriaceae and Mycobacteriaceae belong to the suborder Corynebacterineae within the Actinomycetales (Stackebrandt et al. 1997).
Despite the industrial usage of C. glutamicum since the 1960s, studies on regulatory processes at the transcriptional and posttranscriptional level started only four decades later. LysG, the activator of the lysine exporter gene lysE, was the first transcriptional regulator to be reported (Bellmann et al. 2001). With the availability of genome sequences (Ikeda and Nakagawa 2003; Kalinowski et al. 2003; Yukawa et al. 2007), DNA microarrays for genome-wide expression analysis (Muffler et al. 2002; Hüser et al. 2003; Wendisch 2003), and proteomics (Hermann et al. 1998; Schaffer et al. 2001), research on regulation was boosted, particularly at the transcriptional level (for a review, see Schröder and Tauch 2010), but also at the posttranscriptional level (Bendt et al. 2003).
Protein phosphorylation is a key mechanism for the regulation of cellular activities at different levels. In C. glutamicum, four serine/threonine protein kinases (PknA, PknB, PknG, and PknL) and a single phosphoserine/phosphothreonine protein phosphatase have been annotated and were studied experimentally (Niebisch et al. 2006; Fiuza et al. 2008b; Schultz et al. 2009). A few target proteins of the kinases including phosphorylation sites have been identified, the first one being OdhI, a 15-kDa protein with a forkhead-associated domain. In its unphosphorylated state, OdhI inhibits the activity of the 2-oxoglutarate dehydrogenase complex by binding to the OdhA subunit, and this inhibition can be relieved by the PknG-catalyzed phosphorylation of OdhI at threonine-14 (Niebisch et al. 2006; Barthe et al. 2009; Krawczyk et al. 2010). This inhibition was shown to be crucial for glutamate production (Schultz et al. 2007). In addition, MurC (Fiuza et al. 2008a), FtsZ (Schultz et al. 2009), and RsmP (Fiuza et al. 2010), which are involved in peptidoglycan biosynthesis, cell division, and cell morphology, respectively, were identified as targets of serine/threonine protein kinases. Although phosphorylation of serine and threonine residues can also be involved in transcriptional regulation (Sharma et al. 2006a, b), histidine and aspartate phosphorylation by two-component signal transduction systems is the much more important and prevalent manner in bacteria.
Basics of two-component signal transduction
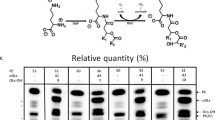
Two-component systems (TCS) consist of a usually membrane-bound sensor kinase or histidine kinase (HK) and a response regulator (RR), which in most cases functions as transcriptional regulator. Both HKs and RRs are modular proteins (Fig. 1). Typical HKs are composed of a sensor domain, which is highly variable among different HKs, and a conserved cytoplasmic kinase core consisting of two distinct domains: a dimerization and histidine-phosphotransfer domain, designated HisKA domain in PFAM, and a C-terminal catalytic and ATP-binding (CA) domain, termed HATPase_c domain in the PFAM database (Punta et al. 2012). The HATPase domain binds ATP and catalyzes the transfer of the γ-phosphoryl group from ATP to the histidine residue, which is located within the HisKA domain. Several sequence motifs of the HATPase domain involved in ATP binding (G1, F, G2) are highly conserved. In many cases, additional domains such as HAMP domains are located between the N-terminal sensor domain and the C-terminal kinase core. Typical RRs are composed of a conserved N-terminal receiver domain (response_reg domain in PFAM), which contains the phosphorylatable aspartate residue, and a variable C-terminal effector or output domain. The HK responds to a certain stimulus by autophosphorylation of the conserved histidine residue in the HisKA domain and the phosphoryl group is subsequently transferred to the aspartate residue in the receiver domain of the RR in a reaction catalyzed by the RR. Phosphorylation activates (or in exceptional cases inhibits) the RR which then elicits a stimulus-specific response, usually the activation or repression of target genes (for reviews, see Stock et al. 1989, 2000; Bourret et al. 1991; Parkinson and Kofoid 1992; Mascher et al. 2006; Gao and Stock 2009). In this review, we summarize the experimental knowledge currently available for the TCS of the C. glutamicum type strain ATCC 13032 and we present an in silico analysis of TCS in Corynebacterium species for which complete genome sequences are available.

Schematic overview on the histidine kinases and their cognate response regulators of C. glutamicum ATCC 13032. The putative transmembrane helices were predicted by TopPred-II (von Heijne 1992; Claros and von Heijne 1994). Only candidates with a score above 1.2 were considered. The conserved domains and their location are indicated as predicted by PFAM (Punta et al. 2012)
Two-component systems in C. glutamicum ATCC 13032
The first Corynebacterium genome that was completely sequenced and analyzed with respect to TCS was the one of C. glutamicum ATCC 13032 (Ikeda and Nakagawa 2003; Kalinowski et al. 2003). Genes for 13 HKs and 13 RRs were identified, all forming HK–RR or RR–HK pairs (Kocan et al. 2006). No orphan HKs or RRs were found as in many other bacteria like Escherichia coli and Bacillus subtilis (Mizuno 1997; Fabret et al. 1999). Bioinformatic analyses revealed that the HKs as well as the RRs of C. glutamicum can be classified into three different classes, as shown in Tables 1 and 2 which also list the locus tags given in the different genome annotations. According to the classification scheme of Grebe and Stock, seven HKs belong to Histidine Protein Kinase subfamily HPK1, one to HPK5, and the remaining five to HPK7 (Grebe and Stock 1999). Classification of the RRs according to their output domains revealed a comparable distribution to the one of the sensor kinases: seven RRs belong to the OmpR family, one to the CitB family, and five to the LuxR family of RRs. Interestingly, all sensor kinases of HPK1 are paired with an OmpR-type response regulator, the HPK5-type sensor kinase is paired with the CitB-type RR, and all sensor kinases of HPK7 are paired with a LuxR-type RR (Fig. 1). All of the output domains of the C. glutamicum RRs contain DNA-binding motifs, suggesting that all of them function as transcriptional regulators (Kocan et al. 2006).
In order to test for the essentiality of the C. glutamicum ATCC 13032 TCS, a deletion study was performed which revealed that all TCS genes except for regX3 (= cgtR4, cg0484) could be deleted (Kocan et al. 2006). Thus, of the 13 TCS only the SenX3-RegX3 system appears to be essential for growth.
The CitA-CitB system: control of citrate utilization
The HK CitA and its cognate RR CitB of C. glutamicum belong to a family of TCS controlling the uptake and metabolism of citrate and dicarboxylates, the founding member being the citrate utilization (CitAB) TCS of Klebsiella pneumoniae (Bott et al. 1995). Citrate is a ubiquitous natural compound which can be utilized as a carbon and energy source by many bacterial species. Whereas anaerobic catabolism of citrate, which occurs for example in enteric bacteria (Bott 1997) and lactic acid bacteria (Bekal et al. 1998), requires a number of specific enzymes, in particular citrate lyase (Bott and Dimroth 1994), aerobic bacteria possessing a complete tricarboxylic acid cycle usually only require a citrate uptake system in order to be able to metabolize citrate.
C. glutamicum is able to grow aerobically in minimal medium with citrate as sole carbon and energy source (Polen et al. 2007; Brocker et al. 2009). When glucose is present in addition to citrate, both substrates are consumed simultaneously (Brocker et al. 2009), a feature typical for C. glutamicum. Global gene expression studies using DNA microarrays revealed that two putative citrate transport systems showed strongly increased expression in the presence of citrate, i.e. CitH (previously also named CitM or CitP) and TctABC (Polen et al. 2007). The former is a member of the citrate-Mg2+/H+/citrate-Ca2+/H+ symporter family (CitMHS), the latter belongs to the tripartite tricarboxylate transporter family (TTT). Expression of either citH or the tctCBA operon in E. coli enabled citrate utilization, confirming that both CitH and TctABC are functional citrate transporters (Brocker et al. 2009). Growth studies suggested that CitH is active with Ca2+ or Sr2+, but not with Mg2+, whereas TctABC is active with Mg2+ or Ca2+ but not with Sr2+. Evidence was obtained that 2 mM Ca2+ is sufficient to achieve maximal growth rates of C. glutamicum on citrate, whereas Mg2+ is required at 50-fold higher concentrations (Brocker et al. 2009). Either CitH alone or TctABC alone are sufficient for growth on citrate.
The genes encoding the CitAB TCS of C. glutamicum are located immediately upstream of citH and in opposite direction. The HK CitA (58.6 kDa) contains two putative transmembrane helices that border an extracytoplasmic domain extending from residues 48–188. The RR CitB (23.4 kDa) is composed of the receiver domain and a DNA-binding domain belonging to the CitB family. A C. glutamicum mutant lacking the citAB genes was unable to grow with citrate as the sole carbon and energy source, but grew like wild type on glucose or pyruvate. The Cit− phenotype could be abolished by transformation of the ΔcitAB mutant with a citAB expression plasmid, confirming that the CitAB TCS is required for citrate utilization (Brocker et al. 2009). DNA microarray and primer extension experiments revealed that the citrate-inducible expression of both citH and tctCAB is strictly dependent on the CitAB TCS. Furthermore, the purified RR CitB was shown to bind to the promoter regions of citH and tctCBA (Brocker et al. 2009). The exact DNA-binding motif has not yet been identified. According to these results, CitA serves as a sensor for extracellular citrate and triggers the phosphorylation of CitB, which then activates the expression of the citrate transport genes citH and tctCBA (Fig. 2).

Control of citrate transport by the CitAB two-component system of C. glutamicum. The target genes shown in green are activated in the presence of extracellular citrate and encode two distinct citrate transport systems
The question how citrate is sensed by CitA has not yet been answered experimentally for the protein of C. glutamicum, but for the HKs CitA from K. pneumoniae and E. coli. For K. pneumoniae it was shown that the isolated periplasmic domain CitAP binds citrate, presumably the dianionic form H-citrate2−, in a 1:1 stoichiometry with a K d of 5 μM at pH 7 (Kaspar et al. 1999). In the case of E. coli CitA, the periplasmic domain bound citrate with a K d of about 0.3 μM at pH 7 (Kaspar and Bott 2002). The crystal structure of K. pneumoniae CitAP in complex with citrate was the first one solved for a periplasmic domain of a HK and revealed a PAS-fold, a versatile ligand-binding structural motif. The groups responsible for citrate binding were identified as Thr-58, Arg-66, His-69, Ser-101, Leu-102, Lys-109, Ser124, and Arg-107. Four of these (R66, H69, R107, and K109) had been identified before as important for citrate binding by showing that their exchange to alanine increased the K d 38- to >300-fold (Gerharz et al. 2003). In a subsequent study, structures of K. pneumoniae CitAP in the citrate-free and citrate-bound states were solved and their comparison showed that ligand binding causes a considerable contraction of the sensor domain (Sevvana et al. 2008). This contraction may represent the molecular switch that activates transmembrane signaling in the receptor, causing a piston-like movement of the second transmembrane helix towards the periplasm. In C. glutamicum CitA, all of the citrate-binding residues of K. pneumoniae CitA except Thr-58 and Ser-101 are conserved, suggesting that also the corynebacterial CitA protein directly senses the presence of citrate via its extracytoplasmic domain.
The MtrB-MtrA system: osmoregulation and control of cell wall metabolism
The MtrAB TCS, which was the first one studied in C. glutamicum, is highly conserved in sequence and genomic organization in actinobacteria (Hoskisson and Hutchings 2006) and the RR MtrA was shown to be essential in M. tuberculosis (Zahrt and Deretic 2000). The RR MtrA of C. glutamicum is a 24.9 kDa protein with an OmpR-type DNA-binding output domain (Fig. 1). In contrast to M. tuberculosis, deletion of the mtrA gene alone and together with mtrB was possible in C. glutamicum. ΔmtrAB mutant cells exhibited a pleiotropic phenotype. The cells were elongated, segmented, and some showed irregular septum formation. In addition, they were more sensitive to penicillin and vancomycin, inhibitors of transpeptidases in cell wall synthesis, but more resistant to ethambutol, which interferes with the synthesis of the arabinogalactan moiety of the cell wall in Corynebacterineae (Belanger et al. 1996). These facts implied that the MtrAB TCS is somehow involved in cell wall homeostasis (Möker et al. 2004).
DNA microarray analysis comparing the ΔmtrA or the ΔmtrAB mutant with the wild type combined with different types of MtrA-DNA-interaction studies (ChIP-chip analyses, DNA affinity chromatography, and electrophoretic mobility shift assays) revealed 22 MtrA target genes/operons, some being transcriptionally activated and some being transcriptionally repressed by MtrA (Fig. 3; Brocker and Bott 2006; Brocker et al. 2011). These contrary functions of activation and repression correlate with the position of the MtrA binding site in the promoter region of the corresponding target gene(s): it is located in the vicinity of the −10 region when MtrA acts as a repressor, whereas it is located upstream of the −35 region, when MtrA acts as an activator. In vitro phosphorylation of MtrA by phosphoramidate caused dimerization of the response regulator and enhanced its DNA-binding affinity, indicating that MtrA is activated by phosphorylation. The DNA-binding site of MtrA as determined by experimental work and bioinformatics was found to be a loosely conserved 8-bp direct repeat separated by a 3-bp linker (consensus sequence (A/G)TAACAATttn(A/G)TAACAAT), whose length is important for MtrA binding (Brocker et al. 2011). Direct repeats are often found as binding sites of OmpR-type regulators (Gupta et al. 2006; Hickey et al. 2011 and references therein). Based on structural studies with PhoB of E. coli it was proposed that upon phosphorylation OmpR-type RRs get activated by a mechanism in which the receiver domains form a twofold symmetric dimer while the DNA-binding domains bind to DNA with tandem symmetry (Bachhawat et al. 2005). The results obtained for MtrA of C. glutamicum are in accordance with such a mechanism.

Regulon of the MtrAB two-component system of C. glutamicum showing its involvement in osmoregulation and cell wall metabolism. Genes shown in red are repressed and genes shown in green are activated by MtrA. Target proteins with known function are indicated. For genes highlighted in yellow, a binding of the response regulator MtrA upstream of these genes was observed, but the mRNA level of the genes was not altered in mutants lacking mtrAB or mtrA
Most of the MtrA target genes code for proteins of unknown function. Among the characterized target genes, betP and proP code for uptake carriers for the compatible solutes glycine betaine and proline or ectoine, respectively, which are involved in the response to hyperosmotic stress (Peter et al. 1998; Morbach and Krämer 2005). The genes mepA, mepB, and nlpC code for putative cell wall peptidases, csbD for a homolog of a protein belonging to the σB-dependent general stress regulon in B. subtilis (Pragai and Harwood 2002), and rpf2 for a resuscitation promoting factor (Hartmann et al. 2004). The phenotype of the ΔmtrAB mutant together with the function of the proteins encoded by the MtrA target genes indicated that the MtrAB system is involved in cell wall metabolism and the osmostress response. This conclusion is in agreement with experimental results on MtrA proteins of mycobacteria, which show high sequence identity to MtrA of C. glutamicum (e.g. 74 % to MtrA of M. tuberculosis [MtrA M.t. ]). In M. smegmatis, downregulation of mtrA expression using an antisense mRNA technique resulted in elongated cells and an increased sensitivity to the antituberculosis drugs isoniazid and streptomycin (Li et al. 2010). In mycobacteria, several MtrA target genes have been described, such as the promoter regions of dnaA and fbpB coding for the initiator protein of DNA replication and the major secreted immunodominant antigen Ag85B, respectively, and the chromosomal origin of replication oriC (Fol et al. 2006; Li et al. 2010; Rajagopalan et al. 2010). MtrA M.t. is constitutively expressed throughout growth in human macrophages (Haydel and Clark-Curtiss 2004) and MtrA can be detected in sera from TB patients (Singh et al. 2001) indicating that the MtrAB TCS is involved in pathogenesis in M. tuberculosis.
MtrB (54.7 kDa), the cognate HK of MtrA, contains two putative transmembrane helices which border an extracytoplasmatic domain of ~151 aa. To search for the stimulus sensed by MtrB, Strep-tagged MtrB was purified, reconstituted into proteoliposomes and its activities, autophosphorylation and phosphoryl group transfer to MtrA, were compared in the absence and presence of different stimuli (Möker et al. 2007a). Potassium ions were shown to stimulate MtrB activity, but this effect was also observed for DcuS of E. coli, a sensor kinase involved in recognition of C4 dicarboxylates. Therefore, K+ seems to have a general stimulation effect on HKs rather than being the stimulus of MtrB to sense hyperosmotic stress (Möker et al. 2007a). In subsequent studies, membrane shrinkage was excluded as being the specific stimulus for MtrB. Various compounds such as amino acids, sugars, and polyethylene glycols were shown to activate MtrB, presumably not by binding to a specific binding site, but by changing the hydration state of MtrB. As this activation was independent of the periplasmic loop and the HAMP domain (Fig. 1), the kinase domain was proposed to sense hypertonicity (Möker et al. 2007b).
Immediately downstream of mtrA–mtrB, the gene lpqB is located which encodes a lipoprotein of unknown function. In M. tuberculosis, it has been shown that the LpqB protein interacts with the extracellular domain of MtrB, thereby influencing MtrA phosphorylation and expression of the MtrA target gene dnaA (Nguyen et al. 2010). Hence, MtrAB together with LpqB seem to form a three-component system, as already suggested previously (Hoskisson and Hutchings 2006).
The PhoS-PhoR system: coping with phosphate starvation
Phosphorus is one of the macroelements of all cells and makes up 1.5–2.1 % of the cell dry weight of C. glutamicum (Liebl 2005). Inorganic phosphate (Pi) is the preferred phosphorus source of C. glutamicum and half-maximal growth rates are obtained at a Pi concentration of about 0.1 mM (Monod constant; Ishige et al. 2003). Besides Pi, various other inorganic and organic phosphates can serve as phosphorus sources for C. glutamicum (Wendisch and Bott 2005; Wendisch and Bott 2008). When Pi is abundant, C. glutamicum accumulates up to 600 mM polyphosphate (Pallerla et al. 2005; Klauth et al. 2006) and various enzymes involved polyphosphate synthesis and degradation have been characterized (Lindner et al. 2007, 2009, 2010a, b).
When Pi becomes scarce, a genetic program is started and expression of a group of genes, called phosphate starvation inducible (psi) genes, increases (Ishige et al. 2003). From the kinetics of the transcriptional response, the following strategy to cope with Pi limitation was deduced. The first response after sensing Pi limitation is the increased expression of the pstSCAB operon, which encodes an ABC transporter for high affinity Pi uptake whose activity allows uptake of low residual Pi from the environment. Secondly, activated expression of ugpAEBC encoding an ABC transporter for glycerol 3-phosphate and glpQ1 coding for a glyceryophosphoryl diester phosphodiesterase confers the ability to liberate glycerol 3-phosphate from lipids and to import this organophosphate into the cell. Simultaneously, increased expression of the pctABCD operon coding for an ABC transporter with yet unknown substrate specificity may allow the uptake of phosphorus-containing compounds. In a next phase, expression of a number of genes coding for secreted enzymes is increased allowing the mobilization of phosphate from nontransportable organophosphates. This group includes the nucH, ushA, and phoC. NucH is presumably a secreted nuclease for hydrolysis of extracellular DNA and RNA to desoxynucleotides and nucleotides. UshA was shown to be a secreted enzyme with UDP-sugar hydrolase and 5-nucleotidase activity, allowing access to phosphate in nucleotides (Rittmann et al. 2005). PhoC is proposed to function as a cell wall-associated phosphatase with unknown substrate specificity.
As TCS are obvious candidates for being involved in the regulation of the Pi starvation response, the set of 12 non-essential C. glutamicum TCS deletion mutants was screened for growth under Pi limitation. One of the mutants, strain ΔphoRS (originally named ΔcgtRS3), had a growth defect under Pi limitation, but not under Pi excess (Kocan et al. 2006). Interestingly, the phoRS genes were the only two-component genes whose expression was rapidly (within 10 min) and transiently induced after a shift from Pi excess to Pi starvation (Ishige et al. 2003). Both results suggested that the PhoRS TCS was involved in the adaptation to Pi limitation. Transcriptome comparisons and primer extension studies of the ΔphoRS mutant and the wild type demonstrated that none of the psi genes except pstSCAB was induced in the mutant within 60 min after a shift from Pi excess to Pi limitation. Activation of the pstSCAB genes was weaker in the ΔphoRS mutant than in the wild type (Kocan et al. 2006).
The HK PhoS (52.4 kDa) contains two transmembrane helices delimiting an extracytoplasmic domain of about 120 amino acids and in the cytoplasm a HAMP domain followed by the HisKA and HATPase domains. The RR PhoR (26.4 kDa) is composed of an N-terminal receiver domain and a C-terminal output domain of the OmpR family (Fig. 1). In vitro studies revealed that the cytoplasmic part of PhoS showed constitutive autokinase activity and allowed rapid phosphorylation of PhoR. PhoR ~ P bound with different affinity to eight promoters of psi genes/operons, i.e. pstSCAB, phoRS, phoC, ushA, ugpAEBC, nucH, phoH1, and glpQ1 (Fig. 4). In addition, PhoR ~ P also bound to the promoter of the porin gene porB, whose expression was reduced in the ΔphoRS mutant, and to the promoter of the pitA gene encoding a low-affinity secondary phosphate transporter (Schaaf and Bott 2007). Expression of pitA was reduced after a shift to Pi limitation and PhoR ~ P might act as a repressor of pitA. The affinity of unphosphorylated PhoR was about fivefold lower than that of PhoR ~ P, indicating that the latter is the active form of the protein (Schaaf and Bott 2007). The PhoR binding sites in the pstSCAB promoter and in the phoRS promoter were defined as 19-bp motifs composed of a loosely conserved 8-bp tandem repeat separated by a 3-bp linker, whose length is important for binding. The highest affinity was found for an artificial motif containing two perfect 8-bp tandem repeats: CCTGTGAAaatCCTGTGAA (Schaaf and Bott 2007). Again, this type of binding motif is in accord with that proposed for OmpR-type regulators (Bachhawat et al. 2005). The position of the binding motifs in the two promoters suggested different mechanisms of interactions with the RNA polymerase (Schaaf and Bott 2007).

Role of the PhoRS two-component system of C. glutamicum in phosphate starvation. Genes shown in green are activated by PhoR, the pitA gene shown in red is repressed by PhoR. The function of some target proteins is indicated
The positive autoregulation of the phoRS genes, which were supported by reporter gene fusions, could be responsible for the successive expression of the psi genes. Whereas uninduced levels of PhoR ~ P could be sufficient to induce the high-affinity pstSCAB and phoRS promoters, elevated levels of PhoR ~ P might be required for induction of the lower affinity target promoters. Consequently, the PhoRS system may function as a rheostat rather than a simple switch.
The mechanism by which the PhoRS TCS senses phosphate limitation is still an open issue. Experiments in our laboratory with purified PhoS reconstituted into proteoliposomes argued against the idea that the environmental Pi concentration is directly sensed by PhoS. For the PhoR-PhoB TCS of E. coli, which performs a similar function in the phosphate starvation response as PhoRS in C. glutamicum, a model was proposed in which the phosphate ABC transporter PstSCAB is responsible for sensing the periplasmic Pi concentration and transfers this information to the sensor kinase PhoR, whereby also the PhoU protein plays a role (Hsieh and Wanner 2010). As a PhoU homolog is also present in C. glutamicum (encoded by cg2842), a similar mechanism as suggested for E. coli might be involved in Pi sensing by C. glutamicum.
The CopS-CopR system: handling copper stress
Due to its ability to change between Cu(II) and Cu(I), copper serves as a redox cofactor for many enzymes, such as cytochrome c oxidases (Ridge et al. 2008). However, free copper ions can trigger the formation of reactive oxygen species and lead to sulfhydryl depletion. Therefore, high copper concentrations are toxic for cells. Most organisms have the ability to adapt to elevated copper concentrations by preventing the accumulation of free intracellular copper ions. Suitable strategies are for example the induction of copper exporters, of copper chaperons, or of multicopper oxidases to get rid of toxic intracellular copper levels, to sequester free copper ions, or to oxidize Cu(I) to the less toxic Cu(II), respectively (Osman and Cavet 2008).
Except for two putative multicopper oxidases, the only copper-dependent enzyme currently known in C. glutamicum is cytochrome aa 3 oxidase, which forms a supercomplex with the cytochrome bc 1 complex (Niebisch and Bott 2001, 2003; Bott and Niebisch 2003). As this supercomplex is critical for aerobic respiration and oxidative phosphorylation, copper is also required by C. glutamicum. On the other hand, copper concentrations of ≥50 μM inhibited growth of C. glutamicum (Schelder et al. 2011). DNA microarrays studies revealed that the genes encoding the HK CopS (cg3284) and the RR CopR (cg3285) as well as the up- and downstream genes (cg3286-cg3289 and cg3283-cg3281) showed strongly increased expression when cells were cultivated in the presence of 21 μM copper rather than at the routinely used 1 μM, indicating that this gene region is important for the adaptation to copper stress (Schelder et al. 2011).
The relevance of the CopSR TCS for copper homeostasis was confirmed by the finding that a ΔcopSR mutant showed an increased susceptibility to copper ions, but not to nickel, manganese, zinc, silver, cobalt, lead, or cadmium ions. This phenotype was reversed by plasmid-borne copRS expression in the ΔcopSR mutant (Schelder et al. 2011). The HK CopS (43.0 kDa) presumably contains two transmembrane helices bordering a small extracytoplasmic region of about 30 amino acids, a HAMP domain and the characteristic HisKA and HATPase domains (Table 1 and Fig. 1). The RR CopR (26.7 kDa) is composed of a receiver domain and a DNA-binding domain of the OmpR family (Table 2 and Fig. 1). DNA microarray studies revealed no differences in gene expression between the ΔcopSR mutant and the wild type when the strains were cultivated in glucose minimal medium with 1 μM copper, whereas 43 genes displayed a more than threefold altered mRNA level when the strains were grown in the presence of 21 μM copper. In particular, the genes cg3286-cg3289, which are located upstream of copS in reverse orientation showed 50- to 100-fold lower expression in the ΔcopSR mutant, whereas expression of the three genes downstream of copS, cg3283-cg3281 was reduced only by a factor of about two. Binding studies with purified CopR uncovered a single binding site located in the intergenic region between copR and cg3286, which represents a 9-bp tandem repeat separated by 2-bp (TGAAGATTTnnTGAAGATTT). Phosphorylation by acetyl phosphate was shown to enhance the binding affinity of CopR to its DNA target about sixfold. Reporter gene assays indicated that CopR activates both the cg3286 and the copR promoter. According to these data, the CopSR system is activated by elevated copper levels and phosphorylated CopR activates expression of the cg3286-cg3287-cg3288-cg3289 genes and of the copR-copS-cg3283-cg3282-cg3281 genes (Fig. 5).

Response to copper stress by the CopRS two-component system of C. glutamicum. All genes displayed are activated by CopR in response to elevated copper concentrations
Some proteins encoded by these CopR target genes can obviously be linked to copper homeostasis: CopB (encoded by cg3281) is a putative copper export ATPase which exports copper out of the cytoplasm into the extracytoplasmatic space, where a putative multicopper oxidase (encoded by cg3287) can possibly oxidase Cu(I) to the less toxic and less membrane-permeable Cu(II). The function of the putative secreted copper-binding protein Cg3282 in copper homeostasis could be sequestration of excess copper ions or delivery of copper ions from CopB to the multicopper oxidase. The function of the other encoded proteins is still unclear.
Although there is clear evidence that CopS senses elevated copper concentrations, the mechanism of sensing has not yet been clarified. In principle, either CopS alone could be responsible for detection, or one or more additional proteins could be involved, such as a copper-binding protein or a copper transporter. For the copper-responsive HK CinS from Pseudomonas putida, a putative copper binding site has been identified which includes two histidine residues in the periplasmic loop (Quaranta et al. 2009). The periplasmic loop of CopS (LFHDHMLMTGREDPSLELFHAEQAYRDAN) also contains three histidine residues as well as two methionine residues, which might also be involved in direct copper binding as shown for other copper-binding proteins (Davis and O'Halloran 2008).
The HrrS-HrrA system: control of heme homeostasis
Heme is a cofactor of various enzymes, in particular complexes of the respiratory chain, but can also serve as a source of iron. In C. glutamicum, prominent heme-containing enzymes are succinate dehydrogenase, also called succinate:menaquinone oxidoreductase (Kurokawa and Sakamoto 2005), the cytochrome bc 1-aa 3 supercomplex (Niebisch and Bott 2001, 2003; Sone et al. 2001), cytochrome bd oxidase (Kusumoto et al. 2000), respiratory nitrate reductase (Bott and Niebisch 2003), or catalase. Thus, both aerobic and anaerobic respiration of C. glutamicum are strictly dependent on heme (Bott and Niebisch 2003; Nishimura et al. 2007; Takeno et al. 2007) as well as the detoxification of reactive oxygen species generated by respiration. Heme biosynthesis in C. glutamicum occurs via the C5 pathway that uses glutamate as the substrate for the synthesis of δ-aminolevulinic acid (Bott and Niebisch 2003).
As heme is an iron-containing porphyrin, heme metabolism is intimately connected with iron metabolism. Similar to copper, iron is both essential as protein cofactor but also dangerous as ferreous iron catalyzes the formation of reactive oxygen species. Therefore, organisms have evolved sophisticated strategies to ensure sufficient iron supply, but to avoid high, toxic intracellular iron concentrations (Andrews et al. 2003). In C. glutamicum, DtxR has been identified as the master regulator of iron homeostasis (Brune et al. 2006; Wennerhold and Bott 2006; Frunzke and Bott 2008). Among the target genes repressed by DtxR under iron excess were several ones related to heme metabolism, i.e. a putative operon predicted to encode a secreted heme transport-associated protein (htaA, cg0466) and an ABC transporter for heme uptake (hmuTUV, cg0467-cg0468-cg0469), a putative operon predicted to encode to further secreted heme transport-associated proteins (htaB-htaC, cg0470-cg0471), another putative secreted heme transport-associated protein (htaD, cg3156), and the hmuO gene (cg2445) encoding heme oxygenase (Wennerhold and Bott 2006). The presence of genes related to heme import and degradation which are derepressed under iron limitation suggested that the non-pathogenic C. glutamicum can use heme as an iron source. In fact, non-toxic concentrations of hemin (2.5 μM) as sole iron source allowed comparable growth rates and cell yields as the same concentration of FeSO4 (Frunzke et al. 2011). Mutants lacking either the hmu operon (htaA-hmuTUV) or the htaBC operon showed a slight, but significant growth defect when hemin was supplied as iron source, whereas growth of a ΔhmuO mutant lacking heme oxygenase was strongly impaired. These phenotypes support the involvement of the corresponding proteins in heme uptake and degradation. Transcriptome studies revealed that all of the aforementioned genes related to heme metabolism and a few additional ones showed increased expression levels when heme was used as sole iron source (Frunzke et al. 2011).
The set of target genes repressed by DtxR under iron-sufficient conditions in C. glutamicum also includes genes for transcriptional regulators, in particular those for the AraC-type regulator RipA (Wennerhold et al. 2005) and for the RR CgtR11. The gene cgtR11 (hrrA) is located downstream of cgtS11 (hrrS) encoding the cognate HK, which however is not repressed by DtxR (Wennerhold and Bott 2006). Due to the high sequence identity of the CgtSR11 system of C. glutamicum to the HrrSA TCS of C. diphtheriae (Bibb et al. 2007), it was renamed accordingly. The C. diphtheriae HrrSA system was shown to be involved in the heme-dependent activation of hmuO and repression of hemA, encoding the heme biosynthesis enzyme glutamyl-tRNA reductase (Bibb et al. 2007). A ΔhrrA mutant of C. glutamicum showed a strong growth defect on agar plates containing hemin as sole iron source, suggesting that also the C. glutamicum HrrSA system plays a role in heme metabolism. Transcriptome comparisons of the ΔhrrA mutant and the wild type and in vitro studies with purified HrrS and HrrA protein led to the identification of six promoter regions to which HrrA binds and to the definition of the HrrA regulon (Frunzke et al. 2011). The DNA binding motif of HrrA, which is composed of a receiver domain and a LuxR-type DNA binding domain (Fig. 1), has not yet been determined.
Like many other RR, HrrA functions both as an activator and as a repressor. The genes activated by HrrA ~ P code for heme oxygenase (hmuO), for subunit III of cytochrome aa 3 oxidase and the three subunits of the cytochrome bc 1 complex (ctaE-qcrCAB operon), and for subunit I of cytochrome aa 3 oxidase (ctaD). The genes repressed by HtrA ~ P code for 11 proteins involved in heme biosynthesis and cytochrome c maturation (hemE-hemY-hemL-cg0519-ccsX-ccdA-resB-resC; hemA-hemC; hemH). Thus, when heme is available, HrrSA stimulates heme degradation and the synthesis of the heme-containing cytochrome bc 1-aa 3 supercomplex and at the same time reduces heme biosynthesis. It thus plays a key role in heme homeostasis, together with the master regulator DtxR (Fig. 6).

Control of heme homeostasis by the HrrSA two-component system of C. glutamicum. The genes shown in green are activated by HrrA, the genes encircled in red are repressed by HrrA. The genes shown in black, which encode an ABC transporter for heme uptake, are not regulated by HrrA. Rather, they are repressed under iron excess by DtxR, similar to some of the HrrA target genes. For genes highlighted in yellow, a binding of the response regulator HrrA upstream of these genes was observed. It is still unclear if binding of HrrA to this region leads to activation or repression of one or both of the divergently orientated operons. The CgtSR8 two-component system possibly activates expression of the hrtBA genes, which encode a putative heme exporter, in response to elevated heme concentrations. Note that alternative names exist for some of the genes involved in heme biosynthesis: hemY/hemG, cg0520/ccsX/resA, ccsA/ccdA, cg0523/ccsB/resB, ccsB/ccsA/resC
The HrrSA TCS is assumed to be activated by heme, but biochemical studies confirming this assumption are not yet available. The HK HrrS (Fig. 1) is predicted to contain three transmembrane helices that border two extracytoplasmic regions of about 54 and 38 amino acid residues and the conserved HisKA and HATPase domains (Kocan et al. 2006). Heme recognition could occur either at the periplasmic side or within the cytoplasmic membrane.
The CgtS8-CgtR8 system: a second TCS possibly involved in heme homeostasis
Besides HrrSA, a second TCS of C. glutamicum, CgtSR8, might also be involved in the regulation of heme homeostasis. The CgtSR8 system shows high sequence identity to the ChrSA system of C. diphtheriae, which activates hmuO expression and represses hemA expression in a heme-dependent manner (Schmitt 1999; Bibb et al. 2005, 2007). More recently, also expression of the hrtBA genes of C. diphtheriae, which code for an ABC transporter conferring resistance to heme toxicity, was reported to be activated by ChrSA in this bacterium (Bibb and Schmitt 2010). Genes homologous to hrtBA (cg2202, cg2204) are also present in the C. glutamicum genome immediately upstream of cgtSR8 in opposite orientation and expression of both genes was activated by heme (Frunzke et al. 2011). Furthermore, the RR HrrA was shown to bind to the cgtS8-hrtB intergenic region and expression of cgtSR8 and hrtBA was increased two- and ten-fold, respectively, in the ΔhrrA mutant during growth with heme as sole iron source. These data suggest that CgtSR8 might have a similar function as ChrSA in C. diphtheriae and that HrrSA and CgtSR8 of C. glutamicum, which show sequence similarity to each other, are interrelated (Frunzke et al. 2011).
For ChrS of C. diphtheriae, experimental evidence was provided that it functions as heme sensor. The autophosphorylation of purified ChrS reconstituted into proteoliposomes was shown to be stimulated by 1 μM hemin, but not by other metalloporphyrins and iron. In addition, UV-spectra supported a direct interaction between ChrS and hemin (Ito et al. 2009). The N-terminal region of ChrS (and CgtS8) is predicted to contain five transmembrane helices and the deduced topology was supported by PhoA and LacZ fusions (Bibb and Schmitt 2010). Distinct amino acid substitutions (R34A, Y61F, R70A, D75N, F114N) in the N-terminal region of ChrS were found to inhibit or prevent heme-dependent activation of the hrtB promoter or to cause heme-independent, constitutive promoter activation (H21L). These data support a role of the N-terminal ChrS region in heme sensing and signal transfer to the kinase domain (Bibb and Schmitt 2010).
Conservation of the C. glutamicum two-component systems in other species of corynebacteria
Except for the TCS of C. glutamicum ATCC 13032 described above and the HrrSA and ChrSA systems of C. diphtheriae, no other TCS of corynebacteria have been experimentally studied to our knowledge. However, a variety of genome sequences of Corynebacterium species were determined in recent years, which are of interest because of their pathogenicity, their role in cheese ripening, or in amino acid production. We performed an in silico analysis of these genomes to determine the conservation of the C. glutamicum TCS. The results of this analysis are summarized in Table 3 and Table S1, which lists the GI numbers of the orthologous proteins.
In C. glutamicum strain R, genes for 13 sensor kinases and 14 response regulators are found (Yukawa et al. 2007). Homologs of CgtSR6 and of CgtS7 are absent, while two TCS not present in strain ATCC 13032 are found in strain R (cgR_2292, cgR_2299, cgR_0540, cgR_0541). Thus, even in strains of the same species the presence of TCS can vary.
Considering their conservation in Corynebacterium strains with known genome sequence, the TCS can be divided into three groups. Group A contains three highly conserved TCS, namely MtrAB, PhoSR and CgtSR4 (SenX3/RegX3). They are present in all (MtrAB) or all except for one (PhoRS) or two (SenX3/RegX3) species analyzed here. It can be assumed that these TCS play important roles in the physiology of corynebacteria, which is supported in the case of MtrAB and PhoRS by the large regulons and in the case of CgtR4 by its essentiality in C. glutamicum ATCC 13032. Group B involves four TCS that are present in 12–18 of the 22 species analyzed here: CopSR, HrrSA, CgtSR2, and CgtSR7. The function of the latter two is not yet known. Finally group C contains nine TCS that are found in one to nine species. Of these, only the function of the CitAB and the ChrSA (CgtSR8) system is currently known.
Concluding remarks
The work summarized above illustrates that significant progress was achieved in understanding the role of TCS in C. glutamicum since the genome sequence was published in 2003 and allowed the in silico identification of these signal transduction systems. Cellular functions were elucidated for five of the 13 TCS present in the type strain ATCC 13032 by using the following approaches: (1) comparison with TCS of known functions from other bacteria; (2) inspection of the genomic environment of the TCS genes; (3) search for phenotypes of deletion mutants lacking a particular TCS; and (4) identification of the target genes of the RRs. The five characterized systems include CitAB (citrate uptake), MtrAB (osmoregulation and cell wall homoeostasis), PhoSR (phosphate limitation), CopSR (copper stress), and HrrSA (heme homeostasis). The stimuli sensed by the corresponding HK are predicted to be citrate (CitA), phosphate limitation (PhoS), copper ions (CopS), and heme (HrrS); however, direct biochemical evidence for these predictions is not yet available. Further studies along the lines outlined above should allow to elucidate the roles of all 13 TCS of C. glutamicum. The resulting knowledge will contribute to a systemic understanding of this species and can be used for optimization of strains or process conditions used for industrial purposes.
References
Andrews SC, Robinson AK, Rodriguez-Quinones F (2003) Bacterial iron homeostasis. FEMS Microbiol Rev 27:215–237
Bachhawat P, Swapna GV, Montelione GT, Stock AM (2005) Mechanism of activation for transcription factor PhoB suggested by different modes of dimerization in the inactive and active states. Structure 13:1353–1363
Barthe P, Roumestand C, Canova MJ, Kremer L, Hurard C, Molle V, Cohen-Gonsaud M (2009) Dynamic and structural characterization of a bacterial FHA protein reveals a new autoinhibition mechanism. Structure 17:568–578
Becker J, Wittmann C (2011) Bio-based production of chemicals, materials and fuels—Corynebacterium glutamicum as versatile cell factory. Curr Opin Biotechnol 23:1–10
Bekal S, Van Beeumen J, Samyn B, Garmyn D, Henini S, Divies C, Prevost H (1998) Purification of Leuconostoc mesenteroides citrate lyase and cloning and characterization of the citCDEFG gene cluster. J Bacteriol 180:647–654
Belanger AE, Besra GS, Ford ME, Mikusova K, Belisle JT, Brennan PJ, Inamine JM (1996) The embAB genes of Mycobacterium avium encode an arabinosyl transferase involved in cell wall arabinan biosynthesis that is the target for the antimycobacterial drug ethambutol. Proc Natl Acad Sci USA 93:11919–11924
Bellmann A, Vrljic M, Patek M, Sahm H, Krämer R, Eggeling L (2001) Expression control and specificity of the basic amino acid exporter LysE of Corynebacterium glutamicum. Microbiology 147:1765–1774
Bendt AK, Burkovski A, Schaffer S, Bott M, Farwick M, Hermann T (2003) Towards a phosphoproteome map of Corynebacterium glutamicum. Proteomics 3:1637–1646
Bibb LA, Schmitt MP (2010) The ABC transporter HrtAB confers resistance to hemin toxicity and is regulated in a hemin-dependent manner by the ChrAS two-component system in Corynebacterium diphtheriae. J Bacteriol 192:4606–4617
Bibb LA, King ND, Kunkle CA, Schmitt MP (2005) Analysis of a heme-dependent signal transduction system in Corynebacterium diphtheriae: deletion of the chrAS genes results in heme sensitivity and diminished heme-dependent activation of the hmuO promoter. Infect Immun 73:7406–74012
Bibb LA, Kunkle CA, Schmitt MP (2007) The ChrA-ChrS and HrrA-HrrS signal transduction systems are required for activation of the hmuO promoter and repression of the hemA promoter in Corynebacterium diphtheriae. Infect Immun 75:2421–2431
Blombach B, Riester T, Wieschalka S, Ziert C, Youn JW, Wendisch VF, Eikmanns BJ (2011) Corynebacterium glutamicum tailored for efficient isobutanol production. Appl Environ Microbiol 77:3300–3310
Bott M (1997) Anaerobic citrate metabolism and its regulation in enterobacteria. Arch Microbiol 167:78–88
Bott M, Dimroth P (1994) Klebsiella pneumoniae genes for citrate lyase and citrate lyase ligase: localization, sequencing, and expression. Mol Microbiol 14:347–356
Bott M, Niebisch A (2003) The respiratory chain of Corynebacterium glutamicum. J Biotechnol 104:129–153
Bott M, Meyer M, Dimroth P (1995) Regulation of anaerobic citrate metabolism in Klebsiella pneumoniae. Mol Microbiol 18:533–546
Bourret RB, Borkovich KA, Simon MI (1991) Signal transduction pathways involving protein phosphorylation in prokaryotes. Annu Rev Biochem 60:401–441
Brocker M, Bott M (2006) Evidence for activator and repressor functions of the response regulator MtrA from Corynebacterium glutamicum. FEMS Microbiol Lett 264:205–212
Brocker M, Schaffer S, Mack C, Bott M (2009) Citrate utilization by Corynebacterium glutamicum is controlled by the CitAB two-component system through positive regulation of the citrate transport genes citH and tctCBA. J Bacteriol 191:3869–3880
Brocker M, Mack C, Bott M (2011) Target genes, consensus binding site, and role of phosphorylation for the response regulator MtrA of Corynebacterium glutamicum. J Bacteriol 193:1237–1249
Brune I, Werner H, Hüser AT, Kalinowski J, Pühler A, Tauch A (2006) The DtxR protein acting as dual transcriptional regulator directs a global regulatory network involved in iron metabolism of Corynebacterium glutamicum. BMC Genomics 7:21
Burkovski A (ed) (2008) Corynebacteria: genomics and molecular biology. Caister Academic Press, Norfolk, UK
Cerdeno-Tarraga AM, Efstratiou A, Dover LG, Holden MTG, Pallen M, Bentley SD, Besra GS, Churcher C, James KD, De Zoysa A, Chillingworth T, Cronin A, Dowd L, Feltwell T, Hamlin N, Holroyd S, Jagels K, Moule S, Quail MA, Rabbinowitsch E, Rutherford KM, Thomson NR, Unwin L, Whitehead S, Barrell BG, Parkhill J (2003) The complete genome sequence and analysis of Corynebacterium diphtheriae NCTC13129. Nucleic Acids Res 31:6516–6523
Ciccarelli FD, Doerks T, von Mering C, Creevey CJ, Snel B, Bork P (2006) Toward automatic reconstruction of a highly resolved tree of life. Science 311:1283–1287
Claros MG, von Heijne G (1994) TopPred II: an improved software for membrane protein structure predictions. Comput Appl Biosci 10:685–686
Davis AV, O'Halloran TV (2008) A place for thioether chemistry in cellular copper ion recognition and trafficking. Nat Chem Biol 4:148–151
Eggeling L, Bott M (eds) (2005) Handbook of Corynebacterium glutamicum. CRC Press, Boca Raton, FL, USA
Fabret C, Feher VA, Hoch JA (1999) Two-component signal transduction in Bacillus subtilis: how one organism sees its world. J Bacteriol 181:1975–1983
Fiuza M, Canova MJ, Patin D, Letek M, Zanella-Cleon I, Becchi M, Mateos LM, Mengin-Lecreulx D, Molle V, Gil JA (2008a) The MurC ligase essential for peptidoglycan biosynthesis is regulated by the serine/threonine protein kinase PknA in Corynebacterium glutamicum. J Biol Chem 283:36553–36563
Fiuza M, Canova MJ, Zanella-Cleon I, Becchi M, Cozzone AJ, Mateos LM, Kremer L, Gil JA, Molle V (2008b) From the characterization of the four serine/threonine protein kinases (PknA/B/G/L) of Corynebacterium glutamicum toward the role of PknA and PknB in cell division. J Biol Chem 283:18099–18112
Fiuza M, Letek M, Leiba J, Villadangos AF, Vaquera J, Zanella-Cleon I, Mateos LM, Molle V, Gil JA (2010) Phosphorylation of a novel cytoskeletal protein (RsmP) regulates rod-shaped morphology in Corynebacterium glutamicum. J Biol Chem 285:29387–29397
Fol M, Chauhan A, Nair NK, Maloney E, Moomey M, Jagannath C, Madiraju MV, Rajagopalan M (2006) Modulation of Mycobacterium tuberculosis proliferation by MtrA, an essential two-component response regulator. Mol Microbiol 60:643–657
Frunzke J, Bott M (2008) Regulation of iron homeostasis in Corynebacterium glutamicum. In: Burkovski A (ed) Corynebacteria: genomics and molecular biology. Caister Academic Press, Norfolk, UK, pp 241–266
Frunzke J, Gätgens C, Brocker M, Bott M (2011) Control of heme homeostasis in Corynebacterium glutamicum by the two-component system HrrSA. J Bacteriol 193:1212–1221
Gao R, Stock AM (2009) Biological insights from structures of two-component proteins. Annu Rev Microbiol 63:133–154
Gerharz T, Reinelt S, Kaspar S, Scapozza L, Bott M (2003) Identification of basic amino acid residues important for citrate binding by the periplasmic receptor domain of the sensor kinase CitA. Biochemistry 42:5917–5924
Grebe TW, Stock JB (1999) The histidine protein kinase superfamily. Adv Microb Physiol 41:139–227
Gupta S, Sinha A, Sarkar D (2006) Transcriptional autoregulation by Mycobacterium tuberculosis PhoP involves recognition of novel direct repeat sequences in the regulatory region of the promoter. FEBS Lett 580:5328–5338
Hartmann M, Barsch A, Niehaus K, Pühler A, Tauch A, Kalinowski J (2004) The glycosylated cell surface protein Rpf2, containing a resuscitation-promoting factor motif, is involved in intercellular communication of Corynebacterium glutamicum. Arch Microbiol 182:299–312
Haydel SE, Clark-Curtiss JE (2004) Global expression analysis of two-component system regulator genes during Mycobacterium tuberculosis growth in human macrophages. FEMS Microbiol Lett 236:341–347
Hermann T, Wersch G, Uhlemann EM, Schmid R, Burkovski A (1998) Mapping and identification of Corynebacterium glutamicum proteins by two-dimensional gel electrophoresis and microsequencing. Electrophoresis 19:3217–3221
Hickey JM, Weldon L, Hefty PS (2011) The atypical OmpR/PhoB response regulator ChxR from Chlamydia trachomatis forms homodimers in vivo and binds a direct repeat of nucleotide sequences. J Bacteriol 193:389–398
Hoskisson PA, Hutchings MI (2006) MtrAB-LpqB: a conserved three-component system in actinobacteria? Trends Microbiol 14:444–449
Hsieh YJ, Wanner BL (2010) Global regulation by the seven-component Pi signaling system. Curr Opin Microbiol 13:198–203
Hüser AT, Becker A, Brune I, Dondrup M, Kalinowski J, Plassmeier J, Pühler A, Wiegrabe I, Tauch A (2003) Development of a Corynebacterium glutamicum DNA microarray and validation by genome-wide expression profiling during growth with propionate as carbon source. J Biotechnol 106:269–286
Ikeda M, Nakagawa S (2003) The Corynebacterium glutamicum genome: features and impacts on biotechnological processes. Appl Microbiol Biotechnol 62:99–109
Inui M, Kawaguchi H, Murakami S, Vertes AA, Yukawa H (2004) Metabolic engineering of Corynebacterium glutamicum for fuel ethanol production under oxygen-deprivation conditions. J Mol Microbiol Biotechnol 8:243–254
Ishige T, Krause M, Bott M, Wendisch VF, Sahm H (2003) The phosphate starvation stimulon of Corynebacterium glutamicum determined by DNA microarray analyses. J Bacteriol 185:4519–4529
Ito Y, Nakagawa S, Komagata A, Ikeda-Saito M, Shiro Y, Nakamura H (2009) Heme-dependent autophosphorylation of a heme sensor kinase, ChrS, from Corynebacterium diphtheriae reconstituted in proteoliposomes. FEBS Lett 583:2244–2428
Kalinowski J, Bathe B, Bartels D, Bischoff N, Bott M, Burkovski A, Dusch N, Eggeling L, Eikmanns BJ, Gaigalat L, Goesmann A, Hartmann M, Huthmacher K, Krämer R, Linke B, McHardy AC, Meyer F, Möckel B, Pfefferle W, Pühler A, Rey DA, Rückert C, Rupp O, Sahm H, Wendisch VF, Wiegrabe I, Tauch A (2003) The complete Corynebacterium glutamicum ATCC 13032 genome sequence and its impact on the production of l-aspartate-derived amino acids and vitamins. J Biotechnol 104:5–25
Kaspar S, Bott M (2002) The sensor kinase CitA (DpiB) of Escherichia coli functions as a high-affinity citrate receptor. Arch Microbiol 177:313–321
Kaspar S, Perozzo R, Reinelt S, Meyer M, Pfister K, Scapozza L, Bott M (1999) The periplasmic domain of the histidine autokinase CitA functions as a highly specific citrate receptor. Mol Microbiol 33:858–872
Klauth P, Pallerla SR, Vidaurre D, Ralfs C, Wendisch VF, Schoberth SM (2006) Determination of soluble and granular inorganic polyphosphate in Corynebacterium glutamicum. Appl Microbiol Biotechnol 72:1099–1106
Kocan M, Schaffer S, Ishige T, Sorger-Herrmann U, Wendisch VF, Bott M (2006) Two-component systems of Corynebacterium glutamicum: deletion analysis and involvement of the PhoS-PhoR system in the phosphate starvation response. J Bacteriol 188:724–732
Krawczyk S, Raasch K, Schultz C, Hoffelder M, Eggeling L, Bott M (2010) The FHA domain of OdhI interacts with the carboxyterminal 2-oxoglutarate dehydrogenase domain of OdhA in Corynebacterium glutamicum. FEBS Lett 584:1463–1468
Kurokawa T, Sakamoto J (2005) Purification and characterization of succinate:menaquinone oxidoreductase from Corynebacterium glutamicum. Arch Microbiol 183:317–324
Kusumoto K, Sakiyama M, Sakamoto J, Noguchi S, Sone N (2000) Menaquinol oxidase activity and primary structure of cytochrome bd from the amino-acid fermenting bacterium Corynebacterium glutamicum. Arch Microbiol 173:390–397
Li Y, Zeng J, Zhang H, He ZG (2010) The characterization of conserved binding motifs and potential target genes for M. tuberculosis MtrAB reveals a link between the two-component system and the drug resistance of M. smegmatis. BMC Microbiol 10:242
Liebl W (2005) In: Eggeling L, Bott M (eds) Handbook of Corynebacterium glutamicum. CRC Press, Boca Raton, FL, USA, pp 9–34
Lindner SN, Vidaurre D, Willbold S, Schoberth SM, Wendisch VF (2007) NCgl2620 encodes a class II polyphosphate kinase in Corynebacterium glutamicum. Appl Environm Microbiol 73:5026–5033
Lindner SN, Knebel S, Wesseling H, Schoberth SM, Wendisch VF (2009) Exopolyphosphatases Ppx1 and Ppx2 from Corynebacterium glutamicum. Appl Environm Microbiol 75:3161–3170
Lindner SN, Knebel S, Pallerla SR, Schoberth SM, Wendisch VF (2010a) Cg2091 encodes a polyphosphate/ATP-dependent glucokinase of Corynebacterium glutamicum. Appl Microbiol Biotechnol 87:703–713
Lindner SN, Niederholtmeyer H, Schmitz K, Schoberth SM, Wendisch VF (2010b) Polyphosphate/ATP-dependent NAD kinase of Corynebacterium glutamicum: biochemical properties and impact of ppnK overexpression on lysine production. Appl Microbiol Biotechnol 87:583–593
Litsanov B, Kabus A, Brocker M, Bott M (2012a) Efficient aerobic succinate production from glucose in minimal medium with Corynebacterium glutamicum. Microb Biotechnol 5:116–128
Litsanov B, Brocker M, Bott M (2012b) Towards homosuccinate fermentation: metabolic engineering of Corynebacterium glutamicum for anaerobic succinate production from glucose and formate. Appl Environ Microbiol 78(9):3325–3337
Litsanov B, Brocker M, Bott M (2012c) Glycerol as a substrate for aerobic succinate production in minimal medium with Corynebacterium glutamicum. Microb Biotechnol. doi:10.1111/j.1751-7915.2012.00347.x
Mascher T, Helmann JD, Unden G (2006) Stimulus perception in bacterial signal-transducing histidine kinases. Microbiol Mol Biol Rev 70:910–938
Meissner D, Vollstedt A, van Dijl JM, Freudl R (2007) Comparative analysis of twin-arginine (Tat)-dependent protein secretion of a heterologous model protein (GFP) in three different Gram-positive bacteria. Appl Microbiol Biotechnol 76:633–642
Mimitsuka T, Sawai H, Hatsu M, Yamada K (2007) Metabolic engineering of Corynebacterium glutamicum for cadaverine fermentation. Biosci Biotechnol Biochem 71:2130–2135
Mizuno T (1997) Compilation of all genes encoding two-component phosphotransfer signal transducers in the genome of Escherichia coli. DNA Res 4:161–168
Möker N, Brocker M, Schaffer S, Krämer R, Morbach S, Bott M (2004) Deletion of the genes encoding the MtrA-MtrB two-component system of Corynebacterium glutamicum has a strong influence on cell morphology, antibiotics susceptibility and expression of genes involved in osmoprotection. Mol Microbiol 54:420–438
Möker N, Krämer J, Unden G, Krämer R, Morbach S (2007a) In vitro analysis of the two-component system MtrB-MtrA from Corynebacterium glutamicum. J Bacteriol 189:3645–3649
Möker N, Reihlen P, Krämer R, Morbach S (2007b) Osmosensing properties of the histidine protein kinase MtrB from Corynebacterium glutamicum. J Biol Chem 282:27666–27677
Morbach S, Krämer R (2005) Osmoregulation. In: Eggeling L, Bott M (eds) Handbook of Corynebacterium glutamicum. CRC Press, Boca Raton, pp 417–435
Muffler A, Bettermann S, Haushalter M, Horlein A, Neveling U, Schramm M, Sorgenfrei O (2002) Genome-wide transcription profiling of Corynebacterium glutamicum after heat shock and during growth on acetate and glucose. J Biotechnol 98:255–268
Nguyen HT, Wolff KA, Cartabuke RH, Ogwang S, Nguyen L (2010) A lipoprotein modulates activity of the MtrAB two-component system to provide intrinsic multidrug resistance, cytokinetic control and cell wall homeostasis in Mycobacterium. Mol Microbiol 76:348–364
Niebisch A, Bott M (2001) Molecular analysis of the cytochrome bc 1-aa 3 branch of the Corynebacterium glutamicum respiratory chain containing an unusual diheme cytochrome c 1. Arch Microbiol 175:282–294
Niebisch A, Bott M (2003) Purification of a cytochrome bc 1-aa 3 supercomplex with quinol oxidase activity from Corynebacterium glutamicum—identification of a fourth subunit of cytochrome aa 3 oxidase and mutational analysis of diheme cytochrome c 1. J Biol Chem 278:4339–4346
Niebisch A, Kabus A, Schultz C, Weil B, Bott M (2006) Corynebacterial protein kinase G controls 2-oxoglutarate dehydrogenase activity via the phosphorylation status of the OdhI protein. J Biol Chem 281:12300–12307
Nishimura T, Vertes AA, Shinoda Y, Inui M, Yukawa H (2007) Anaerobic growth of Corynebacterium glutamicum using nitrate as a terminal electron acceptor. Appl Microbiol Biotechnol 75:889–897
Nishio Y, Nakamura Y, Kawarabayasi Y, Usuda Y, Kimura E, Sugimoto S, Matsui K, Yamagishi A, Kikuchi H, Ikeo K, Gojobori T (2003) Comparative complete genome sequence analysis of the amino acid replacements responsible for the thermostability of Corynebacterium efficiens. Genome Res 13:1572–1579
Okino S, Noburyu R, Suda M, Jojima T, Inui M, Yukawa H (2008) An efficient succinic acid production process in a metabolically engineered Corynebacterium glutamicum strain. Appl Microbiol Biotechnol 81:459–464
Osman D, Cavet JS (2008) Copper homeostasis in bacteria. Adv Appl Microbiol 65:217–247
Pallerla SR, Knebel S, Polen T, Klauth P, Hollender J, Wendisch VF, Schoberth SM (2005) Formation of volutin granules in Corynebacterium glutamicum. FEMS Microbiol Lett 243:133–140
Parkinson JS, Kofoid EC (1992) Communication modules in bacterial signaling proteins. Annu Rev Genet 26:71–112
Peter H, Weil B, Burkovski A, Krämer R, Morbach S (1998) Corynebacterium glutamicum is equipped with four secondary carriers for compatible solutes: identification, sequencing, and characterization of the proline/ectoine uptake system, ProP, and the ectoine/proline/glycine betaine carrier, EctP. J Bacteriol 180:6005–6012
Polen T, Schluesener D, Poetsch A, Bott M, Wendisch VF (2007) Characterization of citrate utilization in Corynebacterium glutamicum by transcriptome and proteome analysis. FEMS Microbiol Lett 273:109–119
Pragai Z, Harwood CR (2002) Regulatory interactions between the Pho and σB-dependent general stress regulons of Bacillus subtilis. Microbiology 148:1593–1602
Punta M, Coggill PC, Eberhardt RY, Mistry J, Tate J, Boursnell C, Pang N, Forslund K, Ceric G, Clements J, Heger A, Holm L, Sonnhammer EL, Eddy SR, Bateman A, Finn RD (2012) The Pfam protein families database. Nucleic Acids Res 40:D290–D301
Quaranta D, McEvoy MM, Rensing C (2009) Site-directed mutagenesis identifies a molecular switch involved in copper sensing by the histidine kinase CinS in Pseudomonas putida KT2440. J Bacteriol 191:5304–5311
Rajagopalan M, Dziedzic R, Al Zayer M, Stankowska D, Ouimet MC, Bastedo DP, Marczynski GT, Madiraju MV (2010) Mycobacterium tuberculosis origin of replication and the promoter for immunodominant secreted antigen 85B are the targets of MtrA, the essential response regulator. J Biol Chem 285:15816–15827
Ridge PG, Zhang Y, Gladyshev VN (2008) Comparative genomic analyses of copper transporters and cuproproteomes reveal evolutionary dynamics of copper utilization and its link to oxygen. PLoS One 3:e1378
Rittmann D, Sorger-Herrmann U, Wendisch VF (2005) Phosphate starvation-inducible gene ushA encodes a 5'-nucleotidase required for growth of Corynebacterium glutamicum on media with nucleotides as the phosphorus source. Appl Environ Microbiol 71:4339–4344
Schaaf S, Bott M (2007) Target genes and DNA-binding sites of the response regulator PhoR from Corynebacterium glutamicum. J Bacteriol 189:5002–5011
Schaffer S, Weil B, Nguyen VD, Dongmann G, Günther K, Nickolaus M, Hermann T, Bott M (2001) A high-resolution reference map for cytoplasmic and membrane-associated proteins of Corynebacterium glutamicum. Electrophoresis 22:4404–4422
Schelder S, Zaade D, Litsanov B, Bott M, Brocker M (2011) The two-component signal transduction system CopRS of Corynebacterium glutamicum is required for adaptation to copper-excess stress. PLoS One 6:e22143
Schmitt MP (1999) Identification of a two-component signal transduction system from Corynebacterium diphtheriae that activates gene expression in response to the presence of heme and hemoglobin. J Bacteriol 181:5330–5340
Schneider J, Wendisch VF (2010) Putrescine production by engineered Corynebacterium glutamicum. Appl Microbiol Biotechnol 88:859–868
Schröder J, Tauch A (2010) Transcriptional regulation of gene expression in Corynebacterium glutamicum: the role of global, master and local regulators in the modular and hierarchical gene regulatory network. FEMS Microbiol Rev 34:685–737
Schröder J, Maus I, Trost E, Tauch A (2011) Complete genome sequence of Corynebacterium variabile DSM 44702 isolated from the surface of smear-ripened cheeses and insights into cheese ripening and flavor generation. BMC Genomics 12:545
Schultz C, Niebisch A, Gebel L, Bott M (2007) Glutamate production by Corynebacterium glutamicum: dependence on the oxoglutarate dehydrogenase inhibitor protein OdhI and protein kinase PknG. Appl Microbiol Biotechnol 76:691–700
Schultz C, Niebisch A, Schwaiger A, Viets U, Metzger S, Bramkamp M, Bott M (2009) Genetic and biochemical analysis of the serine/threonine protein kinases PknA, PknB, PknG and PknL of Corynebacterium glutamicum: evidence for non-essentiality and for phosphorylation of OdhI and FtsZ by multiple kinases. Mol Microbiol 74:724–741
Sevvana M, Vijayan V, Zweckstetter M, Reinelt S, Madden DR, Herbst-Irmer R, Sheldrick GM, Bott M, Griesinger C, Becker S (2008) A ligand-induced switch in the periplasmic domain of sensor histidine kinase CitA. J Mol Biol 377:512–523
Sharma K, Gupta M, Krupa A, Srinivasan N, Singh Y (2006a) EmbR, a regulatory protein with ATPase activity, is a substrate of multiple serine/threonine kinases and phosphatase in Mycobacterium tuberculosis. FEBS J 273:2711–2721
Sharma K, Gupta M, Pathak M, Gupta N, Koul A, Sarangi S, Baweja R, Singh Y (2006b) Transcriptional control of the mycobacterial embCAB operon by PknH through a regulatory protein, EmbR, in vivo. J Bacteriol 188:2936–2944
Singh KK, Zhang X, Patibandla AS, Chien P Jr, Laal S (2001) Antigens of Mycobacterium tuberculosis expressed during preclinical tuberculosis: serological immunodominance of proteins with repetitive amino acid sequences. Infect Immun 69:4185–4191
Smith KM, Cho KM, Liao JC (2010) Engineering Corynebacterium glutamicum for isobutanol production. Appl Microbiol Biotechnol 87:1045–1055
Sone N, Nagata K, Kojima H, Tajima J, Kodera Y, Kanamaru T, Noguchi S, Sakamoto J (2001) A novel hydrophobic diheme c-type cytochrome. Purification from Corynebacterium glutamicum and analysis of the qcrCBA operon encoding three subunit proteins of a putative cytochrome reductase complex. Biochim Biophys Acta 1503:279–290
Stäbler N, Oikawa T, Bott M, Eggeling L (2011) Corynebacterium glutamicum as a host for synthesis and export of d-amino acids. J Bacteriol 193:1702–1709
Stackebrandt E, Rainey FA, WardRainey NL (1997) Proposal for a new hierarchic classification system, Actinobacteria classis nov. Int J Syst Bacteriol 47:479–491
Stock JB, Ninfa AJ, Stock AM (1989) Protein phosphorylation and regulation of adaptive responses in bacteria. Microbiol Rev 53:450–490
Stock AM, Robinson VL, Goudreau PN (2000) Two-component signal transduction. Annu Rev Biochem 69:183–215
Takeno S, Ohnishi J, Komatsu T, Masaki T, Sen K, Ikeda M (2007) Anaerobic growth and potential for amino acid production by nitrate respiration in Corynebacterium glutamicum. Appl Microbiol Biotechnol 75:1173–1182
Tauch A, Kaiser O, Hain T, Goesmann A, Weisshaar B, Albersmeier A, Bekel T, Bischoff N, Brune I, Chakraborty T, Kalinowski J, Meyer F, Rupp O, Schneiker S, Viehoever P, Pühler A (2005) Complete genome sequence and analysis of the multiresistant nosocomial pathogen Corynebacterium jeikeium K411, a lipid-requiring bacterium of the human skin flora. J Bacteriol 187:4671–4682
Tauch A, Schneider J, Szczepanowski R, Tilker A, Viehoever P, Gartemann KH, Arnold W, Blom J, Brinkrolf K, Brune I, Gotker S, Weisshaar B, Goesmann A, Droge M, Pühler A (2008a) Ultrafast pyrosequencing of Corynebacterium kroppenstedtii DSM44385 revealed insights into the physiology of a lipophilic corynebacterium that lacks mycolic acids. J Biotechnol 136:22–30
Tauch A, Trost E, Tilker A, Ludewig U, Schneiker S, Goesmann A, Arnold W, Bekel T, Brinkrolf K, Brune I, Gotker S, Kalinowski J, Kamp PB, Lobo FP, Viehoever P, Weisshaar B, Soriano F, Droge M, Puhler A (2008b) The lifestyle of Corynebacterium urealyticum derived from its complete genome sequence established by pyrosequencing. J Biotechnol 136:11–21
Trost E, Gotker S, Schneider J, Schneiker-Bekel S, Szczepanowski R, Tilker A, Viehoever P, Arnold W, Bekel T, Blom J, Gartemann KH, Linke B, Goesmann A, Pühler A, Shukla SK, Tauch A (2010a) Complete genome sequence and lifestyle of black-pigmented Corynebacterium aurimucosum ATCC 700975 (formerly C. nigricans CN-1) isolated from a vaginal swab of a woman with spontaneous abortion. BMC Genomics 11:91
Trost E, Ott L, Schneider J, Schroder J, Jaenicke S, Goesmann A, Husemann P, Stoye J, Dorella FA, Rocha FS, Soares Sde C, D'Afonseca V, Miyoshi A, Ruiz J, Silva A, Azevedo V, Burkovski A, Guiso N, Join-Lambert OF, Kayal S, Tauch A (2010b) The complete genome sequence of Corynebacterium pseudotuberculosis FRC41 isolated from a 12-year-old girl with necrotizing lymphadenitis reveals insights into gene-regulatory networks contributing to virulence. BMC Genomics 11:728
Trost E, Al-Dilaimi A, Papavasiliou P, Schneider J, Viehoever P, Burkovski A, Soares SC, Almeida SS, Dorella FA, Miyoshi A, Azevedo V, Schneider MP, Silva A, Santos CS, Santos LS, Sabbadini P, Dias AA, Hirata R Jr, Mattos-Guaraldi AL, Tauch A (2011) Comparative analysis of two complete Corynebacterium ulcerans genomes and detection of candidate virulence factors. BMC Genomics 12:383
von Heijne G (1992) Membrane protein structure prediction. Hydrophobicity analysis and the positive-inside rule. J Mol Biol 225:487–494
Wendisch VF (2003) Genome-wide expression analysis in Corynebacterium glutamicum using DNA microarrays. J Biotechnology 104:273–285
Wendisch VF, Bott M (2005) Phosphorus metabolism. In: Eggeling L, Bott M (eds) Handbook of Corynebacterium glutamicum. CRC Press, Boca Raton, FL, USA, pp 377–396
Wendisch VF, Bott M (2008) Phosphorus metabolism and its regulation. In: Burkovski A (ed) Corynebacteria: genomics and molecular biology. Caister Academic Press, Norfolk, pp 203–216
Wendisch VF, Bott M, Eikmanns BJ (2006a) Metabolic engineering of Escherichia coli and Corynebacterium glutamicum for biotechnological production of organic acids and amino acids. Curr Opin Microbiol 9:268–274
Wendisch VF, Bott M, Kalinowski J, Oldiges M, Wiechert W (2006b) Emerging Corynebacterium glutamicum systems biology. J Biotechnol 124:74–92
Wennerhold J, Bott M (2006) The DtxR regulon of Corynebacterium glutamicum. J Bacteriol 188:2907–2918
Wennerhold J, Krug A, Bott M (2005) The AraC-type regulator RipA represses aconitase and other iron proteins from Corynebacterium under iron limitation and is itself repressed by DtxR. J Biol Chem 280:40500–40508
Yukawa H, Omumasaba CA, Nonaka H, Kos P, Okai N, Suzuki N, Suda M, Tsuge Y, Watanabe J, Ikeda Y, Vertes AA, Inui M (2007) Comparative analysis of the Corynebacterium glutamicum group and complete genome sequence of strain R. Microbiology 153:1042–1058
Zahrt TC, Deretic V (2000) An essential two-component signal transduction system in Mycobacterium tuberculosis. J Bacteriol 182:3832–3838
Acknowledgments
We would like to thank Cornelia Gätgens, Martina Kočan, Boris Litsanov, Christina Mack, Sarah Schaaf, Steffen Schaffer, Stephanie Schelder, Brita Weil, Han-Min Woo, and Daniela Zaade for their contributions to two-component signal transduction research in our lab. Financial support by the German Federal Ministry of Education and Research (BMBF) and by Evonik Industries AG (Division Health and Nutrition) is gratefully acknowledged.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Corresponding authors
Electronic supplementary materials
Below is the link to the electronic supplementary material.
ESM 1
PDF 18.8 kb
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Bott, M., Brocker, M. Two-component signal transduction in Corynebacterium glutamicum and other corynebacteria: on the way towards stimuli and targets. Appl Microbiol Biotechnol 94, 1131–1150 (2012). https://doi.org/10.1007/s00253-012-4060-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-012-4060-x