
Abstract
Aims/hypothesis
An association between elevated fasting plasma glucose and the common rs560887 G allele in the G6PC2/ABCB11 locus has been reported. In Danes we aimed to examine rs560887 in relation to plasma glucose and serum insulin responses following oral and i.v. glucose loads and in relation to hepatic glucose production during a hyperinsulinaemic–euglycaemic clamp. Furthermore, we examined rs560887 for association with impaired fasting glycaemia (IFG), impaired glucose tolerance (IGT), type 2 diabetes and components of the metabolic syndrome.
Methods
rs560887 was genotyped in the Inter99 cohort (n = 5,899), in 366 young, healthy Danes, in non-diabetic relatives of type 2 diabetic patients (n = 196), and in young and elderly twins (n = 159). Participants underwent an OGTT, an IVGTT or a 2 h hyperinsulinaemic–euglycaemic clamp.
Results
The rs560887 G allele associated with elevated fasting plasma glucose (p = 2 × 10−14) but not with plasma glucose levels at 30 min (p = 0.9) or 120 min (p = 0.9) during an OGTT. G allele carriers had elevated levels of serum insulin at 30 min during an OGTT (p = 1 × 10−4) and relatives of type 2 diabetes patients carrying the G allele had an increased acute insulin response (p = 4 × 10−4) during an IVGTT. Among elderly twins, G allele carriers had higher basal hepatic glucose production (p = 0.04). Finally, the G allele associated with the risk of having IFG (OR 1.26, 95% CI 1.08–1.47, p = 0.002), but not with IGT (OR 0.94, 95% CI 0.82–1.08, p = 0.4) or type 2 diabetes (OR 0.93, 95% CI 0.84–1.04, p = 0.2).
Conclusions/interpretation
The common rs560887 G allele in the G6PC2/ABCB11 locus is associated with increased fasting glycaemia and increased risk of IFG, associations that may be partly related to an increased basal hepatic glucose production rate.
Similar content being viewed by others


Introduction
In humans the concentration of fasting plasma glucose is subject to tight homeostatic control: a mechanism directed by both neural and humoral factors. A few genetic variants have been reported to influence plasma glucose levels in the general population including GCK rs1799884 [1, 2], GCKR rs780094 [3] and variants in the MTNR1B locus [4–6]; however, these variants explain only a minor part of the total inter-individual variation in plasma glucose levels.
In a meta-analysis of two genome-wide association studies totalling 5,088 non-diabetic individuals a significant association between the rs563694 A allele in the G6PC2/ABCB11 locus and elevated fasting plasma glucose was reported (p = 4 × 10−7). This finding was substantiated in a large-scale replication study involving 23,524 non-diabetic white individuals (p = 7 × 10−26) [7] showing an effect size of 0.065 mmol/l per A allele. A total of 5,734 individuals from the Danish population-based Inter99 cohort genotyped for rs563694 [7–9] was included in the study. Additional analyses of the locus harbouring rs563694 revealed an even stronger association of rs560887 (p = 3 × 10−10) and rs853789 (p = 1 × 10−9) with fasting plasma glucose levels in the initial sample of 5,088 non-diabetic individuals from Finland and Sardinia [7]. Another study including 9,353 French individuals demonstrated that rs560887 was strongly associated with fasting plasma glucose with an effect size of 0.06 mmol/l per G allele (p = 4 × 10−23) [10].
rs560887 is located within the third intron of the glucose-6-phosphatase catalytic subunit 2 gene (G6PC2) and is in strong linkage disequilibrium (LD) (r 2 > 0.8) with rs563694 located 10.9 kb away from rs560887 downstream of G6PC2 [7], and with rs853789 located in intron 19 of the ATP-binding cassette, sub-family B, member 11 gene (ABCB11) 38.3 kb from rs560887 and 27.4 kb from rs563694 (r 2 > 0.8) [7].
Although the association of the G6PC2/ABCB11 variants with fasting plasma glucose has been demonstrated, more detailed physiological studies are lacking. The aim of the present study was to evaluate the impact of rs560887 on plasma glucose and serum insulin levels following oral or i.v. glucose loads and to measure hepatic glucose production both in the basal state and during hyperinsulinaemia. Furthermore, we examined the effect of the variants on the risk of having impaired fasting glycaemia (IFG), impaired glucose tolerance (IGT) and type 2 diabetes. In addition, potential associations with components of the metabolic syndrome were examined.
Methods
Participants
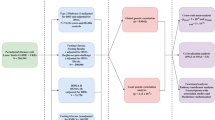
Details of the study populations are given in Electronic supplementary material (ESM) Table 1. Participants from the population-based Inter99 cohort (ClinicalTrials.gov ID no: NCT00289237) [8, 9] involving 4,407 glucose-tolerant individuals who were characterised by an OGTT were investigated for association between the rs560887 genotype and quantitative metabolic traits. Studies of insulin release after an i.v. glucose load (0.3 g glucose/kg body weight) were performed in 366 individuals with normal fasting glucose levels from a population-based sample of young, healthy Danes recruited at the Research Centre for Prevention and Health, Glostrup, Denmark [11] and in a sample of 196 glucose-tolerant relatives of Danish type 2 diabetic patients recruited at the Steno Diabetes Center, Gentofte, Denmark (SDC) [12]. Additionally, rs560887 was examined in a sample of 53 elderly and 106 young twins with normal glucose tolerance subjected to a 2 h hyperinsulinaemic–euglycaemic clamp (40 mU m−2 min−1) with measures of hepatic glucose production during the fasting state and upon insulin infusion [13].
The case–control studies of impaired glucose regulation involving rs560887 included unrelated individuals with isolated IFG recruited from the Inter99 cohort (n = 486) and at SDC (n = 22), isolated IGT (Inter99 n = 466, SDC n = 64), both IFG and IGT (Inter99 n = 199, SDC n = 16), type 2 diabetes (Inter99 n = 323, SDC n = 1,640), and glucose-tolerant control individuals (Inter99 n = 4,407, SDC n = 506). All control participants had normal fasting glycaemia and were glucose-tolerant following an OGTT. Diabetes, IFG and IGT were defined in accordance with World Health Organization 1999 criteria [14].
All participants from the Inter99 cohort (including patients with treated type 2 diabetes) involving a total of 5,899 middle-aged individuals were evaluated in a case–control study examining the association between the rs560887 genotype and the WHO-defined metabolic syndrome (n = 1,326) [14]. In the latter study, those having no components of the metabolic syndrome were considered as control individuals (n = 1,702). More detailed description of participants, biochemical assays and anthropometrical measurements are given in the ESM.
All participants were of Danish nationality and informed written consent was obtained from all individuals before participation. The studies were approved by the Ethical Committee of Copenhagen and were in accordance with the principles of the Helsinki Declaration II.
Biochemical and anthropometric measures
The plasma glucose and serum insulin concentrations were measured as described in the ESM.
Genotyping
Genotyping of rs560887 was performed using TaqMan allelic discrimination (KBioscience, Hoddesdon, UK). Genotype data were obtained in more than 97% of the DNA samples, and among 459 duplicate pairs no discrepancies were found. All genotype groups obeyed Hardy–Weinberg equilibrium.
Statistical analysis
Logistic regression analysis and Fisher’s exact test were applied to test for differences in genotype distribution or allele frequencies. A general linear model was applied to test quantitative variables for differences between genotype groups assuming an additive model. Assuming an allele frequency of the rs560887 A allele of 30% our study of quantitative traits in the Inter99 sample (n = 4,407) has a power of 58%, 93% and 99% to detect a genotype effect difference of 5%, 8% and 10% of an SD, respectively, considering an α of 5%. In the cohort of young, healthy Danes (n = 366) we have a power of 9%, 17% and 22% to detect a genotype effect difference of 5%, 8% and 10% of an SD. Power for the quantitative traits was estimated using simulations assuming an additive genetic model both for the simulation of the data and for testing the data using a linear model. The empirical variance of the observed traits was used to simulate phenotypes from a normal distribution so that variance across genotypes was drawn from the estimated variance. The statistical analyses were performed using RGui version 2.6.1 (available at www.r-project.org, accessed 28 May 2009). Analyses in the sample of non-diabetic relatives of Danish type 2 diabetic patients were performed applying a mixed model including the family relationship (coded as a family number) as a random factor, the genotype and sex as fixed factors, and age and BMI as covariates using the Statistical Package for Social Sciences (SPSS, Chicago, IL, USA) version 14.0. Analyses of hepatic glucose production in the sample of young and elderly twins were calculated using the SAS system version 8.2 (Cary, NC, USA) applying a mixed model including a random-effect term for twin pair membership and a fixed-effects term for zygosity. p < 0.05 was considered significant.
Results
The impact of rs560887 on OGTT-derived diabetes-related quantitative traits was evaluated in a population-based sample of glucose-tolerant individuals from the Inter99 cohort (n = 4,407) (Table 1). The rs560887 G allele was associated with elevated fasting plasma glucose (p = 2 × 10−14), as well as elevated levels of serum insulin (p = 1 × 10−4), 30 min after an oral glucose load and with increased AUC for serum insulin (p = 0.003) during an OGTT, applying an additive genetic model adjusting for age, sex and BMI. Furthermore, the rs560887 G allele was associated with an increased glucose-stimulated insulin release as assessed by the insulinogenic index (p = 3 × 10−6). The same genotype was not associated with 30 or 120 min plasma glucose levels following an OGTT. The insulin concentrations at each time point during the OGTT (0, 30 and 120 min) were additionally analysed adjusting for age, sex, BMI and the simultaneous or fasting plasma glucose concentration; however, the results remained the same when compared with the insulin data adjusted for age, sex and BMI alone (data not shown).
In a subsequent analysis of rs560887 in a population-based sample of 366 young, healthy glucose-tolerant Danes based on normal fasting glucose according to the WHO criteria [14], we found a tendency towards increased acute serum insulin response during an i.v. glucose load (p = 0.09) for carriers of the G allele (Table 2). These findings were substantiated in a study of 196 glucose-tolerant relatives of Danish type 2 diabetic patients. Here we observed a significant association of rs560887 G allele carriers with increased acute serum insulin response following an i.v. glucose load (p = 4 × 10−4; Table 2). The results remained unchanged when adjusting the acute serum insulin response for fasting plasma glucose.
To further investigate the effect of rs560887 on hepatic glucose production we examined a group of 53 elderly and 106 young glucose-tolerant twins all subjected to a 2 h hyperinsulinaemic–euglycaemic clamp. Among the elderly twins, G allele carriers had a higher basal hepatic glucose production (p = 0.04), whereas no effect of genotype was seen on hepatic glucose production (p = 0.5) or insulin-stimulated glucose disposal rate (p = 1.0) during the clamp (Table 3). In the group of young twins (Table 3), and when combining data from young and elderly twins, we found no genotype effect on hepatic glucose production.
The risk of having IFG was evaluated in a case–control study of 508 individuals with IFG and 4,913 glucose-tolerant control individuals. With an OR of 1.26 (95% CI 1.09–1.47) the rs560887 G allele associates with an increased risk of having isolated IFG (p = 0.002) (Table 4). On the contrary, the rs560887 G allele was not associated with the risk of having isolated IGT (OR 0.94, 95% CI 0.82–1.08, p = 0.4) or type 2 diabetes (OR 0.93, 95% CI 0.84–1.04, p = 0.2). However, the rs560887 G allele associates with an increased risk of having combined IFG and IGT (OR 1.40, 95% CI 1.11–1.76, p = 0.005) (Table 4).
Potential associations of the rs560887 G allele with components of the WHO-defined metabolic syndrome were evaluated in a case–control study involving all participants from the Inter99 cohort (n = 5,881). We found no association of the rs560887 G allele with the risk of having either the metabolic syndrome (OR 1.01, 95% CI 0.89–1.15, p = 0.9) or any components of the metabolic syndrome (ESM Table 2).
Discussion
In the present study we show that even though the G allele of rs560887 associates with increased fasting plasma glucose, the G allele is not implicated in altered plasma glucose levels during an OGTT. Furthermore, the G allele associates with increased insulin release after both an oral glucose load and an i.v. glucose injection. Interestingly, the G allele shows nominal association with elevated basal hepatic glucose production in elderly twins. Also, we show that the variant associates with IFG, but not with IGT, type 2 diabetes, or any component of the 1999 WHO-defined metabolic syndrome.
The human G6PC2 encodes the islet-specific glucose-6-phosphatase-related peptide (IGRP). IGRP is known to be specifically expressed in pancreatic beta cells in humans [15], and although no catalytic activity has been reported for IGRP [15–19] it may be assumed that IGRP dephosphorylates glucose 6-phosphate to form glucose, opposing the action of glucokinase, the glucose sensor of the beta cell. Therefore, the effect of variants in the G6PC2/ABCB11 locus on fasting plasma glucose may, as suggested by previous reports [7, 10], imply that the rs560887 G allele results in an increased expression or higher functional activity of IGRP, hereby decreasing the glucose flux into the glycolytic pathway directing the pancreatic beta cell to a higher glucostatic set-point and thus upregulating the impact of the concentration of plasma glucose on insulin secretion in the fasting state [7, 10]. These findings and interpretations are in accord with observations in the G6pc2 knockout mice, where lack of G6pc2 results in a decreased fasting blood glucose concentration [20]. Moreover, a French functional study reported that the G allele of rs573225, a variant in high LD with rs560887, associates with higher promoter activity, suggesting a higher expression of G6PC2 and thereby IGRP [21]. If the rs560887 G allele or another variant in high LD increases the IGRP activity in the beta cell it would most likely lead to elevated plasma glucose levels during an OGTT as a result of a relatively decreased glucose-stimulated serum insulin release. However, we found normal plasma glucose levels and elevated serum insulin levels 30 min after oral and i.v. glucose loads. Thus, our data does not support a sole IGRP-mediated effect of rs560887 on plasma glucose levels.
rs560887 is in high LD with rs853787 and rs853789 located within ABCB11. ABCB11 encodes the bile salt export pump (BSEP), which is evolutionarily highly conserved and solely expressed in the liver. BSEP is involved in the ATP-dependent secretion of bile salts in the hepatocytes and the protein has been linked to inherited forms of cholestasis, resulting in diverse hepato-pathological phenotypes [22]. Variation in ABCB11 impairs bile acid secretion [23–25], and studies involving bile acid sequestrants have shown association between interruption of bile acid re-absorption and decreased levels of plasma glucose and improved insulin sensitivity, most probably as a result of lowered serum triacyglycerol levels [26]. Interestingly, although exploratory because of the low number of individuals in the cohort, it seems from the present study of elderly twins that the increase in fasting plasma glucose in carriers of the rs560887 G allele may be explained by an increase in glucose production from the liver. This finding could, however, not be replicated in the group of young twins and may in theory be explained by an age-dependency in the expression of this distinct phenotypic trait. If the elevated basal hepatic glucose production in G allele carriers is a true finding, it might imply that the raised insulin levels after both oral and i.v. glucose challenges are a compensatory beta cell response primed by the increased fasting plasma glucose concentrations. An interesting approach to further evaluate the ABCB11 locus would be to examine rs560887 in relation to concentrations of bile acid; however, these data are not available in the present study. Given the complex pattern of association we cannot exclude the possibility that two independent signals of association exist, which influence both the function of G6PC2 in the beta cell and ABCB11 in the liver. Functional studies and/or further fine mapping of the locus will be necessary to identify the actual causal variant and the exact mechanisms leading to elevated fasting plasma glucose levels.
Interestingly, even though the rs560887 G allele associates strongly with fasting plasma glucose and IFG it is not associated with IGT or overt type 2 diabetes in the present study. A recent large-scale study of loci affecting fasting plasma glucose levels reported a significant protective effect of the rs560887 G allele on type 2 diabetes (OR 0.93, 5% CI 0.89–0.97, p = 0.0017) [6] with a similar effect size to that observed in the present study. Combined, these studies strongly indicate that the rs560887 G allele associates with elevated fasting plasma glucose levels, and a decreased risk of type 2 diabetes, probably via a hyper-response of the beta cell to postprandial elevation in circulating glucose levels.
A progressive relationship between elevated plasma glucose levels and increased cardiovascular risk, which extends below the threshold chosen for diabetes, has been demonstrated [27]. We have previously shown that the GCK rs1799884 A allele associates with both fasting and post-OGTT plasma glucose levels and with different components of the metabolic syndrome [1]. However, we find no association of the rs560887 G allele with any components of the metabolic syndrome. This might be because of the lack of association with elevated glucose levels after an oral glucose load or the low number of individuals in the sub-analyses.
In conclusion, the present study reports evidence that the observed association of the common rs560887 G allele in the G6PC2/ABCB11 locus with increased fasting plasma glucose and IFG may be related to increased hepatic glucose production rather than a pancreatic beta cell dysfunction. Furthermore, even though the rs560887 G allele is strongly associated with elevated fasting plasma glucose and glucose-stimulated serum insulin release, the variant is not involved in the inter-individual variation of glucose levels after an oral glucose load or the risk of having the metabolic syndrome among middle-aged Danes.
Abbreviations
- IFG:
-
Impaired fasting glycaemia
- IGRP:
-
Islet-specific glucose-6-phosphatase-related peptide
- IGT:
-
Impaired glucose tolerance
- LD:
-
Linkage disequilibrium
- SDC:
-
Steno Diabetes Center
References
Rose CS, Ek J, Urhammer SA et al (2005) A -30G>A polymorphism of the beta-cell-specific glucokinase promoter associates with hyperglycemia in the general population of whites. Diabetes 54:3026–3031
Weedon MN, Clark VJ, Qian Y et al (2006) A common haplotype of the glucokinase gene alters fasting glucose and birth weight: association in six studies and population-genetics analyses. Am J Hum Genet 79:991–1001
Sparsø T, Andersen G, Nielsen T et al (2008) The GCKR rs780094 polymorphism is associated with elevated fasting serum triacylglycerol, reduced fasting and OGTT-related insulinaemia, and reduced risk of type 2 diabetes. Diabetologia 51:70–75
Lyssenko V, Nagorny CL, Erdos MR et al (2009) Common variant in MTNR1B associated with increased risk of type 2 diabetes and impaired early insulin secretion. Nat Genet 41:82–88
Bouatia-Naji N, Bonnefond A, Cavalcanti-Proença C et al (2009) A variant near MTNR1B is associated with increased fasting plasma glucose levels and type 2 diabetes risk. Nat Genet 41:89–94
Prokopenko I, Langenberg C, Florez JC et al (2009) Variants in MTNR1B influence fasting glucose levels. Nat Genet 41:77–81
Chen WM, Erdos MR, Jackson AU et al (2008) Variations in the G6PC2/ABCB11 genomic region are associated with fasting glucose levels. J Clin Invest 118:2620–2628
Jørgensen T, Borch-Johnsen K, Thomsen TF, Ibsen H, Glümer C, Pisinger C (2003) A randomized non-pharmacological intervention study for prevention of ischaemic heart disease: baseline results Inter99. Eur J Cardiovasc Prev Rehabil 10:377–386
Glümer C, Jørgensen T, Borch-Johnsen K, Inter99 study (2003) Prevalences of diabetes and impaired glucose regulation in a Danish population: the Inter99 study. Diabetes Care 26:2335–2340
Bouatia-Naji N, Rocheleau G, Van-Lommel L et al (2008) A polymorphism within the G6PC2 gene is associated with fasting plasma glucose levels. Science 320:1085–1088
Clausen JO, Borch-Johnsen K, Ibsen H et al (1996) Insulin sensitivity index, acute insulin response, and glucose effectiveness in a population-based sample of 380 young healthy Caucasians. Analysis of the impact of gender, body fat, physical fitness, and life-style factors. J Clin Invest 98:1195–1209
Hansen T, Ambye L, Grarup N et al (2001) Genetic variability of the SUR1 promoter in relation to beta-cell function and Type II diabetes mellitus. Diabetologia 44:1330–1334
Poulsen P, Levin K, Petersen I, Christensen K, Beck-Nielsen H, Vaag A (2005) Heritability of insulin secretion, peripheral and hepatic insulin action, and intracellular glucose partitioning in young and old Danish twins. Diabetes 54:275–283
World Health Organization Study Group (1999) Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: Diagnosis and classification of diabetes mellitus. World Health Organization, Geneva
Martin CC, Bischof LJ, Bergman B et al (2001) Cloning and characterization of the human and rat islet-specific glucose-6-phosphatase catalytic subunit-related protein (IGRP) genes. J Biol Chem 276:25197–25207
Petrolonis AJ, Yang Q, Tummino PJ et al (2004) Enzymatic characterization of the pancreatic islet-specific glucose-6-phosphatase-related protein (IGRP). J Biol Chem 279:13976–13983
Shieh JJ, Pan CJ, Mansfield BC, Chou JY (2005) In islet-specific glucose-6-phosphatase-related protein, the beta cell antigenic sequence that is targeted in diabetes is not responsible for the loss of phosphohydrolase activity. Diabetologia 48:1851–1859
Arden SD, Zahn T, Steegers S et al (1999) Molecular cloning of a pancreatic islet-specific glucose-6-phosphatase catalytic subunit-related protein. Diabetes 48:531–542
Shieh JJ, Pan CJ, Mansfield BC, Chou JY (2004) The islet-specific glucose-6-phosphatase-related protein, implicated in diabetes, is a glycoprotein embedded in the endoplasmic reticulum membrane. FEBS Lett 562:160–164
Wang Y, Martin CC, Oeser JK et al (2007) Deletion of the gene encoding the islet-specific glucose-6-phosphatase catalytic subunit-related protein autoantigen results in a mild metabolic phenotype. Diabetologia 50:774–778
Dos-Santos C, Bougnéres P, Fradin D (2009) A single-nucleotide polymorphism in a methylatable Foxa2 binding site of the G6PC2 promoter is associated with insulin secretion in vivo and increased promoter activity in vitro. Diabetes 58:489–492
Lam P, Pearson CL, Soroka CJ, Xu S, Mennone A, Boyer JL (2007) Levels of plasma membrane expression in progressive and benign mutations of the bile salt export pump (Bsep/Abcb11) correlate with severity of cholestatic diseases. Am J Physiol Cell Physiol 293:C1709–C1716
Strautnieks SS, Bull LN, Knisely AS et al (1998) A gene encoding a liver-specific ABC transporter is mutated in progressive familial intrahepatic cholestasis. Nat Genet 20:233–238
Gerloff T, Stieger B, Hagenbuch B et al (1998) The sister of P-glycoprotein represents the canalicular bile salt export pump of mammalian liver. J Biol Chem 273:10046–10050
Jansen PL, Strautnieks SS, Jacquemin E et al (1999) Hepatocanalicular bile salt export pump deficiency in patients with progressive familial intrahepatic cholestasis. Gastroenterology 117:1370–1379
Staels B, Kuipers F (2007) Bile acid sequestrants and the treatment of type 2 diabetes mellitus. Drugs 67:1383–1392
Coutinho M, Gerstein HC, Wang Y, Yusuf S (1999) The relationship between glucose and incident cardiovascular events. A metaregression analysis of published data from 20 studies of 95, 783 individuals followed for 12.4 years. Diabetes Care 22:233–240
Acknowledgements
The authors wish to thank A. Forman, I.-L. Wantzin, T. Lorentzen and M. Stendal for technical assistance and G. Lademann for secretarial support. The study was supported by grants from the Lundbeck Foundation Centre for Applied Medical Genomics in Personalized Disease Prediction, Prevention and Care (LUCAMP), the Danish Diabetes Association, the European Union (EUGENE2, grant no. LSHM-CT-2004-512013), the FOOD Study Group/the Danish Ministry of Food, Agriculture and Fisheries & Ministry of Family and Consumer Affairs (grant no. 2101-05-0044), the Danish Clinical Intervention Research Academy and the Danish Medical Research Council.
Duality of interest
K. Borch-Johnsen, O. Pedersen and T. Hansen hold stock in Novo Nordisk and have received lecture fees from pharmaceutical companies. All other authors declare that there is no duality of interest associated with this manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM Table 1
Clinical characteristics of examined Danish participants (PDF 14 kb)
ESM Table 2
Genotype frequency of rs560887 according to components of the 1999 WHO-defined metabolic syndrome among middle-aged individuals from the Inter99 cohort (PDF 23 kb)
ESM 1
(PDF 23 kb)
Rights and permissions
About this article
Cite this article
Rose, C.S., Grarup, N., Krarup, N.T. et al. A variant in the G6PC2/ABCB11 locus is associated with increased fasting plasma glucose, increased basal hepatic glucose production and increased insulin release after oral and intravenous glucose loads. Diabetologia 52, 2122–2129 (2009). https://doi.org/10.1007/s00125-009-1463-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-009-1463-z