
Abstract
Avian astroviruses comprise a diverse group of viruses affecting many avian species and causing enteritis, hepatitis, and nephritis. To date, six different astroviruses have been identified in avian species based on the species of origin and viral genome characteristics: two turkey-origin astroviruses [Turkey Astrovirus type 1 (TAstV-1) and type 2 (TAstV-2)]; two chicken-origin astroviruses [Avian Nephritis Virus (ANV) and Chicken Astrovirus (CAstV)]; and two duck-origin astrovirus [Duck Astrovirus type 1 (DAstV-1) and type 2 (DAstV-2)]. ANV has also been detected in turkeys, ducklings, pigeons, and guinea fowl; and TAstrovirus-2-like viruses have also been found in guinea fowl. Astroviruses are commonly associated with enteric disease syndromes in poultry including runting-stunting syndrome of broilers (RSS), poult enteritis complex or syndrome (PEC or PES), poult enteritis mortality syndrome (PEMS), and enteritis in guinea fowl. The molecular characterization of the different avian astroviruses shows great genetic variability among each type, and this variability influences the ability to detect these viruses by molecular and serological techniques. In this chapter, we review the different aspects related to avian astroviruses, including molecular biology, pathogenesis, diagnosis, and control.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Introduction
Avian astroviruses comprise a diverse group of viruses affecting many avian species and causing enteritis, hepatitis, and nephritis. Reports of astroviruses as cause of illness in birds predate that of mammals. In 1965, a fatal hepatitis was described in ducklings [2] and was eventually shown to be caused by an astrovirus [22, 23]. Another astrovirus, avian nephritis virus (ANV), was first isolated in 1976 and was associated with growth depression and kidney lesions in young chickens. Initially classified as an Enterovirus, it was later reclassified as an astrovirus following the complete sequencing of the viral genome [30]. To date, six different astroviruses have been identified in avian species based on the species of origin and viral genome characteristics: two turkey-origin astroviruses [Turkey Astrovirus type 1 (TAstV-1) and type 2 (TAstV-2)]; two chicken-origin astroviruses [ANV and Chicken Astrovirus (CAstV)]; and two duck-origin astrovirus [Duck Astrovirus type1 (DAstV-1) and type 2 (DAstV-2)]. ANV has also been detected in turkeys [18, 70], ducklings [6], pigeons [45, 116], and guinea fowl [8]; and TAstV-2-like viruses also have been found in guinea fowl [9, 10].
As in mammals, astroviruses are commonly associated with enteritis in avian species. Enteric diseases cause substantial economic losses in commercial poultry, and many enteric disease syndromes have been described including runting-stunting syndrome of broilers (RSS), poult enteritis complex or syndrome (PEC or PES), and poult enteritis mortality syndrome (PEMS). Astroviruses are among the most common viruses found in cases of PEC, PES, and PEMS in turkeys [2, 3, 34–37, 42, 68, 69, 75, 76, 79, 81, 99]. Astroviruses have also been isolated from cases of RSS in broilers and have been previously associated with poor weight gain, enteric disease, and kidney disease in chickens [4, 15, 27, 83, 85, 86, 105], as well as in guinea fowl suffering from enteritis [10].
Astroviruses were found in turkeys in the UK in 1980 by McNulty et al. [56], and later in the 1980s, in the USA (TAstV-1) [75, 79, 80]. A second TAstV, antigenically and genetically distinct from the previously described isolates, was later characterized [44] and this and similar astroviruses have been designated TAstV-2 [70, 71, 98]. Beyond this classification, data from genetic analyses of field viruses detected in turkeys indicate the circulation of multiple lineages of TAstV-2 [9, 18, 33, 70, 94].
Avian nephritis virus has also been shown to be widely distributed in chicken flocks [30, 53, 70, 103]. Under field conditions, clinical signs associated with infection with this virus broiler chickens have varied from subclinical to outbreaks of RSS and baby chick nephropathy [26]. A second type of astrovirus affecting chickens was shown to be antigenically and genetically distinct from ANV and was named chicken astrovirus (CastV, 4). CAstVs that share high levels of nucleotide sequence identity with this first characterized CAstV have been detected in the USA in broiler chickens affected with RSS [67]. Recently, molecular and antigenic characterization of enterovirus-like viruses (ELVs) demonstrated that they were CAstV, some of them closely related to the first characterized CAstV, and some of them more distantly related [102].
The molecular characterization of the different avian astroviruses shows great genetic variability among each type, and this variability influences the ability to detect these viruses by molecular and serological techniques. It is known that ANV and TAstV-2 each comprise at least two serotypes but many more are suspected. Some studies have shown that TAstV-2, ANV, and CAstV display substantial levels of capsid protein sequence variability, possibly indicating that there are numerous serotypes of each virus [33, 71, 72, 88, 94, 104]. Concomitant infections with different strains have been reported for avian astroviruses and this can result in recombination between strains [71, 72, 94, 104]. Recombination is likely to provide an important mechanism by which the sequence diversity of avian astroviruses is increased. In this chapter, we will review the different aspects related to avian astroviruses, including molecular biology, pathogenesis, diagnosis, and control.
Virus Aspects
Genomic Organization of Avian Astroviruses
The complete genomic sequence of 22 avian astrovirus isolates have been reported, including the genomes of TAstV-1,TAstV-2, ANV, DAstV, and CAstV [20, 30, 38, 39, 44, 94, 113, 115]. Like all other members of the Astroviridae family, avian astroviruses or avastroviruses are composed of a positive sense RNA ranging from 6,623 to 7,722 nt in length, excluding the poly-A tail (Fig. 9.1). While the specific lengths of individual regions vary among isolates, in general each comprises a short 5′ untranslated region (UTR), a large open reading frame (ORF) predicted to encode several nonstructural proteins (ORF1a), a second reading frame predicted to encode the RNA dependent RNA polymerase (ORF1b), a third reading frame which encodes the capsid protein (ORF2), and a 3′UTR, followed by a poly-A tail (Fig. 9.1).

Genomic organization and lengths of regions for each fully characterized avastrovirus genome. Diagrams represent the three basic organizations of the avian astrovirus genomes, and list the specific length of each feature of the fully characterized avian astrovirus isolates. The numbers presented for ORF1a and ORF2 are the nucleotide positions of the start and stop codons. The numbers presented for ORF1a are the nucleotides from the beginning of the “slippery sequence” (AAAAAAC) to the ORF1a stop codon
Interestingly, while these basic features are well conserved across all of Astroviridae, examination of each of the reported genomes demonstrates essentially three basic genome configurations within the avian astroviruses. The ANV-like genome in which ORF1a and ORF2 are in the same frame and ORF1b is in the +1 reading frame; the TAstV-like genome in which ORF1b and ORF2 are both in the +1 reading frame; and the DAstV-like genome in which each ORF is in a distinct reading frame (Fig. 9.1). In addition to these organizational differences, the TAstV-2-like viruses are the only members of Avastrovirus missing the conserved s2m RNA stem-loop motif [44, 94]. While not unprecedented, as the bat, rat, and MBL1 astroviruses are missing this motif [11, 12], its absence suggests this feature is not essential. The prevalence of the s2m motif among Astroviridae as well as other viruses led Monceyron et al. to propose that it was introduced into an astrovirus progenitor via a recombination event between non-related viruses [62]. The biological significance of the loss of s2m or the differences in the organization of the ORFs is unclear, but may reflect differences in evolutionary pressures.
Phylogenetic comparison of the Avastrovirus genomes demonstrates that individual isolates cluster based primarily on the host they have been described to infect. One exception is that of TAstV-1, which is found more closely associated with chicken astroviruses than the other turkey astroviruses (Fig. 9.2). This discrepancy, however, is not unexpected as TAstV-1 has been classified as a member genogroup I (see Taxonomy) with the ANV-like viruses based on phylogenetic comparisons of the capsid genes [6]. The one non-ANV-like CAstV isolate included in this alignment, however, was found to group between genogroups I and II. At present, it is unclear if this represents a distinct genogroup or an evolutionary transition as it has some elements more consistent with genogroup I member TAstV-1 (genomic organization, Fig. 9.1) and others more consistent with genogroup II members TAstV-2 and DAstV (overall length and 5′ UTR, Figs. 9.1 and 9.3).

Phylogenetic relationship of avastroviruses. The full-length genomes of each of the 22 avastroviruses were aligned using ClustalW2, and the phylogeny estimated neighbor using the neighbor joining method and 1,000 bootstrap replicates were calculated. The viruses that have been assigned to one of the two genogroups have been highlighted (genogroup I = brown, genogroup II = blue)

Comparison of the avastrovirus 5′ UTRs and putative “promoter element.” (a) The 5′ UTR sequence from representative ANV (NC003790), CAstV (JF414802), TAstV-1 (NC002470), DAstV (NC012437), and TAstV-2 (NC005790) genomes were aligned using ClustalW. Bolded nucleotides represent positions conserved across a majority of isolates. Underlined positions are predicted to complex to form hairpin. (b) The 5′ UTR sequences were analyzed for secondary structure using mFold. The start codon for ORF1a is underlined. (c) The putative promoter sequence upstream of ORF2 was aligned based on the previously reported astrovirus consensus sequence (REF) which is reported above the alignment (Astro). Positions which match the astrovirus consensus sequence are shown in bold. The boxed positions highlight the ORF2 start codon. The consensus sequence of the avastrovirus isolates (Avastro) is shown at the bottom of the alignment. Lower case letters denote consensus in the variable region. The CCGAA motif found at the very beginning of the 5′ UTR and in the putative promoter region are denoted by asterisk
RNA Functional Elements
Comparison of the 5′ UTR of the different avastrovirus isolates demonstrates a conserved motif in spite of the differences in the lengths between the different viruses (Figs. 9.1 and 9.3). The conserved motif is even more prominent when viewed in the context of its predicted secondary structure (Fig. 9.3b). The conserved stem-loop structure has been proposed to function as a promoter element for RNA transcription [38, 107]. This idea is supported by the fact that a similar highly conserved sequence can be found immediately upstream of ORF2 in the area where RNA transcription of the subgenomic message would begin (Fig. 9.3c). The function of these elements has not been demonstrated experimentally in any astrovirus isolate; however, given their conservation within the genome as well as across Astroviridae this suggests a functional role.
The formation of another RNA structural element, the retrovirus-like frame shift stem-loop, has been a defining characteristic of Astroviridae [32]. The presence or absence of this structure, along with the other basic genomic organization, has been one of the items used in the classification of novel viruses to the family [20, 30, 39, 44]. The stem-loop formed has been described to play a key role in the translation of the ORF1b as part of the ORF1a polyprotein in human astrovirus infected cells [48–50, 55]. Experimental evidence demonstrated that the stem-loop formed allowed for a frame shift to occur upstream of the ORF1a stop-codon, and that brought ORF1b into frame with ORF1a resulting in the translation of one polyprotein [21, 49]. Based on this observation, it has been assumed that the presence of this structure in Avastrovirus genomes meant a similar mechanism allowed for the translation of the avian astrovirus RNA-dependent RNA polymerase (RdRp). Recently, studies by Kang et al. have suggested that the avian astroviruses may not use this strategy [39]. Comparison of the stem-loop predicted for human astrovirus to that of the avian viruses demonstrates differences in their ability to induce the needed frame shift (Fig. 9.4). In human astrovirus, the loop structure is between the “slippery sequence” (AAAAAAC) and the ORF1a stop codon (Fig. 9.4). Conversely, in the avian astroviruses, the ORF1a stop codon is immediately downstream of the “slippery sequence,” and upstream of the stem loop structure (Fig. 9.4). Therefore, any frame shift induced may not allow for translation of ORF1a to read through to ORF1b. Kang et al. proposed that translation of ORF1b in the CAstV could initiate via a start codon just upstream of the “slippery sequence” [39]; however, this is the only avian virus to have a start site immediately upstream of the “slippery sequence” and would require the stem-loop structure not to be formed. Examination of these viruses for other alternative start sites revealed the first ATG in ORF1b was between 68 and 257 nt downstream from the ORF1a stop codon (Fig. 9.4). It is possible that a more complex stem-loop and frame shift structure is produced in these viruses that allows for the translation of ORF1a–1b or, as proposed by Kang et al., perhaps the avian astroviruses use a different strategy for the translation of their RdRp [39].


Avastroviruses have conserved frame shift structure, but may have alternative translational strategy. The ORF1a-ORF1b region reported to form a frame shift structure for the translation of ORF1b was analyzed by mFold for ANV (NC003790), ChAstV (JF414802), TAstV-1 (NC002470), DAstV (NC012437), and TAstV-2 (NC005790) genomes, and compared with HAstV-1 (AY720892). For each genome, the predicted structure formed from a 65 nt region starting 10 nt upstream of the slippery sequence (AAAAAAC, highlighted by dashed box). The ORF1a stop codon is highlighted by a solid box, and potential start codons in the ORF1b frame are underlined
Viral Proteins
ORF1a and ORF1b are thought to encode six or more mature non-structural proteins; however, much of what is known about these proteins is based on sequence comparisons with human astroviruses. Analyses of the Avastrovirus genomes have found evidence of transmembrane proteins, a nuclear localization signal, and the serine protease in ORF1a and the RdRp in ORF1b [20, 30, 38, 44]. While additional studies are needed to further characterize the nonstructural proteins encoded by astroviruses in general, and avastroviruses specifically, recent studies by Nighot et al. and Meyerhoff et al. describe the use of an antibody directed against one the non-structural proteins encoded by TAstV-2 [59, 66]. The development of these reagents and others will facilitate our ability to identify these proteins and understand their role in the virus life cycle.
The best characterized astrovirus protein is the capsid protein; however, even here much of our understanding of the avian astroviruses is based on studies of the human astroviruses. Across the different avian astroviruses, ORF2 encodes a protein ranging in size from 670 to 744 amino acids in length resulting in a precursor protein of 73–81 kDa. It is unclear how this capsid precursor protein is processed to result in mature capsid subunit proteins, or how the subunits then assemble into intact virions, but it is largely assumed that similar to the human astroviruses, the precursor is cleaved by trypsin-like proteases to produce 2–3 capsid subunits ranging in size from 25 to 34 kDa [7]. This assumption is supported by the reports of Tang et al. who identified structural proteins of 31 and 34.5 kDa [98].
While detailed structural and functional studies of the avian astrovirus capsids are lacking, this region of the virus has been the most intensely examined at the genetic level and much of what we know about these viruses and their relationship to each other has come from these bioinformatic analyses. Comparisons among the capsid sequences of avastroviruses and mamastroviruses, as well as within avastroviruses, indicate that all of the astrovirus capsid sequences have essentially three main domains (Fig. 9.5). The N-terminal end of the capsid protein is rich in basic amino acids; 23–35 of the first 100 residues are basic, primarily lysine or arginine. The numbers of basic residues is higher among TAstV-2, DAstV, and CAstV isolates, which have 32–35 positive residues, and contain a conserved SRSRSRSRSRSR motif. The TAstV-1 and ANV isolates have 23–25 positive residues but do not have the SR repeat region. Highly basic N-terminal regions have been demonstrated to play a key role in the encapsidation of viral RNAs for several viruses, and therefore it is assumed that these residues play a similar role in astroviruses [46].

Avastrovirus capsid domains. The relative position of the three functional domains of the astrovirus capsid protein, as described by Krishna (2005), are represented by the blue bar and is superimposed on percent similarity plot demonstrating the level of amino acid homology across 34 avastrovirus capsid sequences (see Fig. 9.5). The similarity plot was constructed using Vector NTI 10.0.1 and using a 25 amino acid window
Analysis of the rest of the avian astrovirus capsid protein demonstrates, as previously described for human astroviruses, the N-terminal half of the protein is far more conserved than the C-terminal half (Fig. 9.5). Analysis of the human astroviruses indicate that the more conserved region of the capsid is important in virion assembly while the more variable region is found on the outer surface of the virion and is involved in interacting with cellular receptors and the immune system [46]. This suggests that these different regions of the capsid protein are under different evolutionary pressures. The first approximately 400 residues are important for encapsidation of the viral RNA and formation of the virion. As such, there are strong structural constraints to the sequence and therefore potential selection against amino acid substitutions. Conversely, the last approximately 300 residues would be exposed to antibodies produced by the host as well as binding to potential new host cells. Especially in the context of interspecies transmission, which appears to occur with some regularity among the avian astroviruses (see “Epidemiology” and “Pathogenesis” sections), one could imagine considerable evolutionary pressure driving amino acid changes in this region. Studies by Strain et al. support this idea [94]. Strain et al. identified 12 codons within the TAstV-2 capsid with evidence of positive selection. Among these 12 codons, 10 were located in the C-terminal half. Additional studies are required to better understand the roles each of these regions play in the life cycle of avian astroviruses and specifically how these different roles affect their evolution.
Taxonomy
Currently, avastrovirus species are based on the hosts from which they were isolated [7]; however, at present the International Committee on Taxonomy of Viruses (ICTV) only recognizes three species within the genus: Chicken astroviruses (ANV-1, ANV-2, ANV-3), Duck astrovirus (DAstV-C-NGB), and Turkey astrovirus (TAstV-1). As such, the species do not correspond to genetic phylogenies (Figs. 9.2 and 9.6), and all the other related viruses are considered members of the genus but have not been approved as species. To address these issues, the classification of avastrovirus species is currently being redefined based on the complete amino acid sequence of the capsid protein. Based on this analysis, the avian astroviruses can be divided into two main genogroups: genogroup I and genogroup II (Fig. 9.6).

Phylogenetic relationship of avastroviruses. The full-length amino acid sequences of the capsid proteins from 34 avastroviruses were aligned using ClustalW2, and the phylogeny estimated using the neighbor joining method and 1,000 bootstrap replicates were calculated using MEGA5. The viruses that have been assigned to one of the two genogroups have been highlighted (genogroup I = brown, genogroup II = blue). Isolates in bold denote those with full genomic sequence represented in Fig. 9.2. The mean amino acid genetic distance (p-dist) ± standard deviation for each of the major clades was calculated using MEGA5
Comparison of the predicted phylogeny of avian astroviruses based on the full-length capsid amino acid sequence (Fig. 9.6) with that based on the available full-length genomic sequence (Fig. 9.1) shows two similar evolutionary relationships. In general, all of the isolates are from a particular type of cluster in distinct clades, and these clades largely correspond with the species the viruses were isolated from. The one major exception is the TAstV-1 isolate, which is found more similar to the ANV viruses (Figs. 9.2 and 9.6). However, more careful inspection of the branch order of the TAstV-2 isolates between Figs. 9.2 and 9.6 demonstrates potentially important differences. Most notably, in Fig. 9.6 the CO/2001 and MI/2001 isolates are on the same branch; however, in Fig. 9.2 they were found to be more divergent. This discrepancy is due to a potential recombination event which occurred between these two viruses at the beginning of ORF2 [94]. This results in two isolates which appear to be identical based on the capsid analysis, but, in fact, are more divergent, and suggests analysis of the capsid region in most cases may be sufficient to assign an individual isolate to a particular group; understanding how that isolate may be related to others within that group requires more extensive comparisons.
As we identify new types of avian astroviruses, such as the CAstVs, it may be necessary to adopt new criteria for determining which genogroup it should be assigned to. Based on the full-genome phylogenetic analysis (Fig. 9.2) CAstVs appear to be closer to the genogroup II (DAstV and TAstV-2) viruses than the genogroup I viruses (ANV and TAstV-1). This observation is supported by the alignment of the capsid sequences (Fig. 9.6); however, there appears to be considerably more capsid diversity among the CAstV isolates than other avian astroviruses. In fact, based on the capsid there appears to be two distinct groups (Fig. 9.6). Comparisons of these groups based on limited regions of ORF1b demonstrate far more homology among these viruses [90], providing more evidence that understanding how individual isolates are related to other astroviruses may require comparisons at the whole genome level.
Epidemiology
Information about the range of avian species infected with astroviruses and the prevalence of avian astrovirus infections has been accumulated from a relatively small number of serological studies and an increasing number of recent studies based on the use of virus-specific RT-PCR tests. Using indirect immunofluorescence (IF) performed with the reference ANV-1 virus (G-4260), investigations in the 1980s and 1990s have shown that ANV infections are highly prevalent in commercial chickens in Japan and in several European countries [13, 31]. Antibody to ANV was also detected in turkey flocks and in SPF flocks [13, 58]. More recently, application of two indirect IF tests developed to separately detect antibodies to two antigenically different CAstVs demonstrated that infections with both CAstV antigenic groups were very common in UK broilers and in broiler parent flocks. Antibodies to both were also detected in parent flocks from many European countries, and were also detected in some UK turkeys [105]. Using a recently developed serological test for DAstV-1, it was found that 63.2% and 51% sera samples from two commercial flocks of ducks that survived DAstV-1 were positive for DAstV [108]. Interestingly, DAstV-specific antibodies were also detected in 28.6% of 42 sera samples from a healthy flock, indicating a wide distribution of subclinical infections caused by DAstV [108]. To our knowledge, seroprevalence studies have not been reported for the TAstVs.
The majority of RT-PCR tests developed for detecting TAstV, ANV, CAstV, and DAstV have used primers that are located in the comparatively well-conserved RNA polymerase (ORF 1b) gene. Based on ORF 1b RT-PCR testing, ANVs have not only been detected in commercial chickens in the USA, UK, and Hungary but have also been detected in turkeys [14], pigeons [116], ducks [6], and guinea fowl [8]. Although the ANVs detected in these species appear to have closely related ORF 1b sequences, it remains to be determined whether the ANVs detected in other bird types are the same as those detected in chickens in terms of their capsid protein sequences. Irrespective of the capsid sequence findings, ANV appears to have a relatively wide host range including non-poultry species [45]. Their ability to infect more than one avian species is likely to add to the diversity exhibited by this virus, and through RNA recombination, allow new variants to emerge.
Other avian astroviruses also appear to be capable of infecting more than one avian species. The detection of TAstV-2 ORF1b fragments by RT-PCR in samples from guinea fowl supports the view that TAstV-2 may have a wider host range [9]. However, knowledge of capsid protein sequences may be required to determine whether the astrovirus from guinea fowl can be classified as a TAstV-2. Full-genome sequence analysis indicated that the DA93 DAstV possessed capsid and RNA polymerase protein sequences that shared ∼70% amino acid identities with that of TAstV-2. It remains to be determined whether ducks and turkeys are infected with the same variants. Although CAstV-like RNA has not yet been detected in other avian species, the detection by indirect IF of CAstV antibodies in turkeys suggests these birds could be infected with CAstV or antigenically related viruses [105]. The host range of TAstV-2 and CAstV appears to extend beyond turkeys and chickens, respectively, and, like ANV, may further extend to non-poultry species.
Although infections with ANV and CAstV are highly prevalent in commercially reared chickens, and infections with TAstVs are also highly prevalent in commercially reared turkeys, there is little or no information about the prevalence of these viruses in other avian species or about the prevalence of DAstVs in ducks. RT-PCR detection rates of 96% and 100% were obtained for CAstV and ANV when sample sets (n = ∼50) from UK broiler flocks affected by enteritis and growth retardation were investigated [88, 103]. Although large numbers of samples from “normal” chickens have not yet been tested, RT-PCR findings to date suggest ANV and CAstV infections are endemic in chickens. TAstVs have a wide geographic distribution and are considered one of the most prevalent virus infections in 1–5-week-old poults suffering from diarrhea [34, 67, 70, 76, 77, 79, 94]. The prevalence of TAstV varies from 40% to 80% in symptomatic poults [9, 18, 34, 76] to less than 50% in healthy flocks [34, 76]. Field observations indicate that TAstV-2 is currently more prevalent in turkeys than TAstV-1 [9, 70] and its occurrence is often associated with other enteric viruses [34, 67, 69, 76].
The application of RT-PCR tests to longitudinal survey samples has shown that ANV and CAstV can be detected in the intestinal contents from chickens at all ages from 4 days of age to slaughter [89, 90, 103]. Given that as high as 108 virus RNA copies/extracted sampled are detected at some time points, it is likely that high levels of virus will be excreted, and, since astroviruses are relatively resistant to chemical and physical inactivation, environmental contamination will be high [89]. Another longitudinal survey conducted to detect enteric viruses in commercial turkey operations demonstrated the presence of TAstV-2, ANV, and TAstV-1 from 2 to 12 weeks of age [69].
With astroviruses being primarily considered as causing enteric infections, spread via the fecal–oral route is likely to be the major means of horizontal transmission. However, horizontal transmission by respiratory or reproductive (male to female via semen) routes cannot be ruled out. Young broiler or turkey poult flocks can become horizontally infected if the housing and equipment within is contaminated with virus due to inadequate cleaning and disinfection between flocks or if the virus is introduced into the house due to poor on-farm biosecurity. Young flocks can also become infected with virus that is acquired vertically. The detection of ANV infections in chickens maintained under quarantine conditions provided indirect evidence of vertical transmission [13], but recently, the detections of low levels of ANV in late-dead duck embryos [6] and chick embryos (Todd et al., unpublished) have provided direct evidence. CAstV has also been detected by RT-PCR in similar dead-in-shell embryo samples (Todd et al., unpublished). Astrovirus excreted from vertically infected chicks will spread horizontally within the flocks.
Pathogenesis
Diseases Caused by Avian Astroviruses
Avian astroviruses, as in mammalian species, have been not only associated with enteritis, but can also cause extra-intestinal diseases including hepatitis and nephritis. Direct correlation of astroviruses identified in feces with enteric disease in birds is complicated by the presence of other enteric pathogens. When separated from other viruses, experimental infection with avian astroviruses can cause illness but usually attenuated when compared to what is seen clinically. The difficulty in obtaining pure astrovirus isolates has hindered pathogenesis studies on some of these viruses.
Duck Astrovirus
Duck astrovirus (DAstV) is an important duck pathogen and causes a highly contagious and fatal hepatitis in ducklings [109]. To date, two different astroviruses causing hepatitis have been recognized in ducks, both of which were initially classified as picornaviruses and named duck hepatitis viruses 2 and 3 (DHV-2 and DHV-3). Duck hepatitis virus 2 (DHV-2), isolated originally by Asplin (1965) [1], was morphologically characterized as an astrovirus in 1984 and renamed duck astrovirus 1 (DAstV-1) [22, 23]. The virus was later definitively identified as an astrovirus [102] and completely sequenced [20]. Duck hepatitis virus 3 (DHV-3), first reported by Toth in 1969 [106] and characterized by Haider and Calnek in 1979 [24], was recently identified by sequencing as a second astrovirus in ducks (DAstV-2) [102].
DAstV-1 (DHV-2) can cause a severe acute disease in young ducks which can result in up to 50% mortality [109]. Infection occurs through oral and cloacal routes. Deaths occur within 1–4 days, usually within 1–2 h after the appearance of clinical signs, which include polydypsia with loose droppings, excessive urate excretion, and sometimes convulsions and acute opisthotonos. Affected ducks usually die in good condition and both the mean death time and the mortality rate (10–50%) depend on the age of the ducks [22]. Survivors excrete virus for at least 1 week after infection and rear normally, with little evidence of retarded growth. The target organs for DAstV-1 appear to be liver and kidneys. The liver is usually pale pink with hemorrhages, and the kidneys are often swollen. Microscopic changes are characterized by extensive necrosis of hepatocytes and bile duct hyperplasia [22]. DAstV-2 has a low pathogenicity for ducklings experimentally infected, and only ducklings appear to be infected by the virus. Mortality rarely exceeds 30% [109] and mature ducks are refractory to the disease [22].
In a recent study, very low mortality rates (4.67% and 0.63%) were observed in two flocks affected by DAstV-1 [108], in sharp contrast with the high mortality rate (up to 50%) in a recent Chinese outbreak and in the 1983 UK outbreak [20, 22]. This may suggest differences in pathogenicity among DAstVs. Co-infections of DAstV and other viruses may also play a role in high mortality observed in some outbreaks.
Turkey Astrovirus
Turkey astroviruses are commonly detected in commercial turkeys experiencing viral enteritis. TAstV-2 has been associated with many enteric disease syndromes in turkeys including poult enteritis complex or syndrome (PEC or PES), and poult enteritis mortality syndrome (PEMS) [8, 18, 34, 44, 70, 75, 112]. Clinical signs of the disease usually develop between 1 and 3 weeks of age and generally last up to 2 weeks [78]. Severity is usually mild to moderate and is characterized by diarrhea, listlessness, litter eating, and nervousness. Although mortality rate is low, morbidity occurring as decreased growth is of great economic concern [2, 78]. The exact role of TAstV-2 as the etiological agent of enteritis in turkeys is difficult to estimate since it can also be detected in apparently healthy turkeys [69, 76]. TAstV-1 has also been detected in healthy turkeys and turkeys with enteric disease, but is less prevalent than TAstV-2.
Experimentally, young turkeys infected with TAstV-2 develop a diarrhea by 2 days post infection which continues for several days [41, 63, 68, 92]. TAstV-2 infection results in overall growth depression of infected birds. Common lesions observed at necropsy are dehydration, distended intestines filled with watery contents and undigested feed, and dilated ceca with foamy contents. Microscopic lesions present in the intestines consist of mild crypt hyperplasia, mild villous atrophy, and mild lymphocytic infiltration, and were most common in the jejunum [5, 41, 68]. Presence of the viruses has been demonstrated by immunohistochemistry (IHC) and by in situ hybridization (ISH) in both villi and crypt enterocytes in the duodenum, jejunum, ileum, and ceca (Fig. 9.7) [5, 41, 59, 66, 68]. Mild lymphocytic infiltration was also observed in other organs including pancreas, liver, spleen, and kidneys. Viremia occurred after infection with TAstV-2 [41, 68]; however, only intestinal cells appear to support viral replication. The possibility exists that low levels of viral replication may occur in other tissues and may not be detected with the tests used which would explain the extra-intestinal lesions observed.

Photomicrographs of jejunum sections from a turkey infected with TAstV-2, 3 days after inoculation; immunohistochemistry (IHC) and in situ hybridization (ISH) staining for detection of the virus. (a) Viral antigen staining present in the cytoplasm of the enterocytes at the middle section of the villi (dark brown); immunoperoxidase labeling, hematoxylin counterstain. (b) Viral RNA staining present in the cytoplasm of the enterocytes (red); digoxygenin labeling, BCIP-NBT staining. Magnification 200×. Pathogenesis of type 2 turkey astroviruses with variant capsid genes in 2-day-old specific pathogen free poults. M. Pantin-Jackwood, E. Spackman and J.M. Day. Avian Pathology 37, 193-201. Reprinted with permission of the publisher (Taylor & Francis Ltd, http://tandf.co.uk/journals)
Mild-to-moderate atrophy of the bursa of Fabricius and thymus was found in TAstV-2-infected poults [5, 41, 63, 68, 74, 81, 92, 99], and was associated with the depletion of lymphoid areas. The cause of these changes in the primary immune organs is unclear and could result in the immune suppression that some have reported with TAstV-2 infections (see section “Immunity”); however, the lymphoid depletion could also be a consequence of maldigestion and malabsorption leading to nutritional deficiencies in these rapidly growing poults.
Early mortality has been reported in poults experimentally inoculated with TAstV-2 viruses [68, 111]. The mortality in these cases most likely occurred because of the high viral challenge the poults received at a very young age, which induced a moderately severe lethargy and feed and water withdrawal that resulted in death. In an earlier study, the pathogenicity of two turkey astrovirus isolates was compared in older turkeys (14–28 days old). Both viruses caused enteritis and growth depression but no mortality [99].
Based on RT-PCR, TAstV-2-like viruses have also been detected in guinea fowl presenting enteric disease but these viruses require further genetic characterization to confirm their identity as TAstV-2 [10].
Avian Nephritis Virus
Avian nephritis virus (ANV) was first isolated from rectal contents of apparently healthy broiler chicks [110] and experimental infections revealed that it mainly results in a sub-clinical disease, although mild growth depression and mortality have been reported for infections with ANV strains including the serotype 1G-4260 strain (ANV-1) [29, 52, 110]. ANV typically causes histological changes in the kidneys although the viral antigen can be detected in other organs including liver, spleen, pancreas, jejunum, and rectum [27, 29, 84, 86]. Young chicks are the most susceptible and become resistant to disease after the first month of life [28]. ANV infections in chicks can vary from subclinical to outbreaks of runting stunting syndrome (RSS) and nephropathy [19, 31, 86, 95, 96, 110]. However, the role of ANV infection on RSS is not clear. Although ANV affected body gain in SPF chicks [85], an experimental study on the effect of ANV and several unclassified enterovirus-like viruses (ELVs) in commercial chicks demonstrated that only one ELV, later also characterized as an ANV, analyzed among the broad panel produced clinical signs in inoculated chicks [57]. Similarly, infection of 1-day-of-age chicks with ELVs, which were later identified as the FP3 and 612 isolates of CAstV and an ANV, showed that the ELVs are able to induce more severe lesions in the intestine, the kidneys, and the pancreas than the G-4260 strain of ANV [89]. Passive and active surveys indicate that ANV is widely distributed worldwide and that serological evidence can be found in turkeys as well [13, 16, 58, 65, 95]. Although an ANV genetically related to ANV-1 was detected in the feces of turkeys [70, 72], to date the infection has never been associated with the disease in turkeys. An attempt to infect poults with the chicken-derived ANV-1 strain was unsuccessful [61].
In chickens experimentally infected with ANV, the virus was first detected in feces 2 days after inoculation, with maximum virus shedding at 4–5 days after inoculation. The virus was widely distributed in tissues, with high titers in the kidney and jejunum and lower titers in the bursa of Fabricius, spleen, and liver [26]. Transient diarrhea and weight gain depression are common signs among 1-day-old infected chickens. At necropsy, mild-to-severe discoloration and swelling in the kidneys are observed, and in dead chicks visceral urate deposits are found [26]. The microscopic lesions described in infected chickens include necrosis and degeneration of epithelial cells of the proximal convoluted tubules of the kidney, interstitial lymphocyte infiltration, and moderate fibrosis. ANV virus particles and antigen were demonstrated in the degenerating kidney epithelium by electron microscopy and IF respectively [26].
Recently, novel ANVs have been isolated from chickens and turkeys suffering from runting and stunting-like and/or locomotory disease [15]. One of these isolates, isolate 19, induced diarrhea and runting and stunting in 3-week-old SPF layer chickens; however, the clinical tenosynovitis, which was seen regularly in the broilers and poults from the field, was not seen in the experimentally infected layers [15]. The isolation of these new avian astrovirus strains from chickens and turkeys with acute tenosynovitis and arthritis without the isolation of any other detectable pathogen suggests that they also might play a role in leg disorders. The pathogenicity of this virus was further demonstrated by the mortality and histopathological findings and seems to be broader than is reported for ANV-1 [15]. Many studies have shown that the pathogenicity of ANV-1 is limited to the kidney of chickens, in contrast to the damage observed in the kidney, pancreas, duodenum, thymus, bursa, and liver seen after infection with this new ANV [15]. Other ANVs have been shown to vary in pathogenicity and tropism. A Belgian PV2 isolate, originally recognized as an ELV, was shown to cause substantial growth retardation, and severe and long-lasting intestinal lesions following experimental infection [17]. Lesions were detected in the pancreas and proventriculus and were absent in the kidneys from chickens experimentally infected with PV2, suggesting that some ANV isolates such as PV2 may have different tissue tropisms [17].
The presence of ANV has also been demonstrated by RT-PCR in organ samples collected from 30-day-old ducklings and from dead-in-shell ducklings [6]. Histological examination of many tissues indicated that lesions were confined to the kidney and intestine. The finding of ANV in ducklings dead in shell indicates naturally occurring vertical infection and it is possible that ANV may play a role in the death of duckling embryos. ANVs were also detected by RT-PCR in 89% of fecal samples from young pigeons suffering from diarrhea, whereas samples from healthy pigeons were negative [115]. In another study, a very high prevalence of ANV was found among feral pigeons and wood pigeons [45]. Astrovirus was detected in almost all juvenile birds, while the detection rate among the adults was 62%. In this case, most birds appeared healthy, and it was not possible to correlate the presence of a virus to any illness in the birds. ANV-like viruses have also been detected in guinea fowl but the role they play in disease is not clear [8].
Chicken Astrovirus
The common detection of CAstV by RT-PCR in cases of broiler chickens affected by growth retardation suggests that infections with this avian astrovirus are contributing to poor growth performance [89]. High levels of CAstV RNA and CAstV antigen have also been detected in kidney tissue from chickens affected by nephritis and visceral gout (Todd, unpublished results).
Experimental infections of 1-day-of-age and 5-day-of-age SPF layer chickens with CAstV caused mild diarrhea and, except for some areas of limited damage at the base of the villi of the small intestine in some birds, no histological visible damage in the liver, kidney, spleen, thymus, or bursa was observed [4]. Used ELV-4 isolate 612, later typed as a CAstV [102], to infect commercial 1-day-old broilers with maternal antibodies against the virus and found only a growth depression of 11.3%, 20.4%, and 9.6% in the three trials that were performed [56]. Experimental infections of 1-day-old SPF layer chickens with CAstV strain 612 caused only very mild histological changes in the intestine and liver, and mild-to-moderate lesions in the kidney [87]. An experimental infection with the FP3 CAstV strain, which was originally known as ELV-3 and which is genetically and antigenically distinct from the 612 strain isolate, caused no clinical signs in 1-day-old broilers [57]. However, experimental infections of 1-day-old SPF layer chickens with the FP3 isolate caused mild-to-moderate histological changes in the kidney, pancreas, and intestine, with virus-specific antigen being detected in these tissues [87].
Mechanisms of Viral Pathogenesis
Our understanding of how astroviruses cause disease trails behind our knowledge of its molecular and epidemiologic properties and is confined mainly to work done with TAstV. Studies by Thouvenelle et al. and Reynolds et al. suggested that astrovirus infection induces changes in the expression of constituent enzymes of the brush border, specifically maltase, with a significant decrease in intestinal d-xylose absorption, resulting in disaccharide maldigestion, malabsorption, and subsequently osmotic diarrhea, even though it causes only mild microscopic lesions [75, 101]. The cellular mechanisms responsible for these changes are still unknown.
Most of the studies done with TAstV have demonstrated that astrovirus infection induces diarrhea in the absence of significant changes in the intestinal morphology. Furthermore, there is little evidence of inflammation and little or no increase in cell death following astrovirus infection [5, 41, 101], further suggesting astrovirus clinical signs result from submicroscopic changes in the intestine. Initial studies to identify physiological changes in infected poults demonstrated changes to ultrastructure of the epithelial cells including rearrangement of F-actin. These changes were associated with decreased apical expression of the sodium/hydrogen exchanger 3 (NHE3) with a concomitant decrease in sodium absorption [66]. Collectively, these data suggest that astrovirus infection induces a malabsorption, osmotic diarrhea, possibly through redistribution of specific brush border enzymes and transporters resulting from dysregulation of the actin cytoskeleton [64, 66, 75, 101].
Immunity
The immune response to astroviruses has not been well characterized in most species. In humans, it has been postulated that anti-astrovirus antibodies developed following natural infections are key mediators in protecting normal healthy adults [47, 60]. In birds; however, this is far less clear and may differ among host species and/or types of avian astroviruses.
Studies with ANV, CAstV, and DAstV suggest that recovery from natural infection and/or vaccination leads to an immune response that protects the host from subsequent infection [1, 22, 28, 83, 109]. Multiple serotypes have been reported for each of these astrovirus types [4, 86, 104]. There are reports of at least two serotypes of TAstV-2; however, curiously there is no experimental evidence of an anamnestic response [41, 100]. This lack of an adaptive immune response has led several laboratories to investigate its effects on the immune system. These studies suggest TAstV-2 may induce immune dysregulation and/or localized inhibition [40, 41, 73, 74].
Studies by Qureshi et al. examined the response of lymphocytes from TAstV-2 or mock-infected poults to the mitogen concanavalin A [74]. These studies demonstrated a decrease in the ex vivo proliferative response to ConA by peripheral blood lymphocytes from TAstV-2 infected poults as compared with controls. Additionally, thymocytes and splenocytes that were exposed to the virus in vitro also demonstrated a reduced proliferative response [74]. In subsequent studies, Qureshi et al. reported TAstV-2 infections also resulted in impaired migration, phagocytic activity, and bacterial killing activity of innate immune cells, as compared with controls [73]. Functional examination of the innate immune cells (macrophages and heterophils) from TAstV-2 infected poults demonstrated reduced expression of the pro-inflammatory cytokines IL-1 and IL-6 [73]. Interestingly, studies by Koci et al. demonstrated significant increases in the levels of active TGF-β in the serum of TAstV-2 infected poults [41]. Collectively, these findings suggest that TAstV-2 infection leads to some sort of immune dysfunction, the specific mechanism of which is still not understood.
In spite of the immune inhibitory effects reported with TAstV-2 infection, the clinical disease is largely resolved within 12–14 days [41], suggesting that some anti-viral host factors are produced. Studies by Qureshi et al. and Koci et al. have both demonstrated that macrophages and adherent splenocytes from TAstV-2-infected poults produce more nitric oxide (NO) following ex vivo stimulation with lipopolysccharide than do cells from uninfected controls [40, 73]. NO has long been recognized as a potent anti-viral effector molecule; however, histological examination of TAstV-2 infected poults does not demonstrate large influxes of macrophages or other innate immune cells. To determine if NO could play a role in the regulation of TAstV-2 replication, the virus was exposed to NO donor compounds in vitro prior to inoculating embryos, and found to inactivate the virus [40]. Conversely, when embryos were pretreated with an inhibitor of the inducible nitric oxide synthase (iNOS) enzyme virus replication increased significantly [40, 51]. Finally, to determine if TAstV-2-infected intestines demonstrated an increase in the expression of iNOS, tissues from TAstV-2-infected turkey poults were stained for the presence of TAstV-2 antigen and turkey iNOS. These experiments demonstrated an increase in iNOS expression following TAstV-2 infection, with the majority of the staining localized to the intestinal epithelium suggesting that in the absence of a response from either innate or adaptive immune cells the intestine is capable of mounting its own innate defense [59].
Diagnosis
Astrovirus infections of commercially reared chickens and turkeys are endemic and this may also be the case with commercially reared ducks. Although infections are common, many are likely to be subclinical, with the incidence of clinical disease that can be directly attributed to a single avian astrovirus being relatively small. Therefore, with regard to diagnosis, it is important to differentiate whether astrovirus-related diseases are being diagnosed or whether astrovirus infections are being diagnosed.
The detection of clinical signs, gross pathological findings, and histological lesions is required before disease can be diagnosed. However, in the case of astroviruses, these in themselves may not be sufficient to identify the causal virus. For example, hepatitis in ducklings may be caused by one of the two different astroviruses: DAstV-1 and DAstV-2, originally known as duck hepatitis virus types 2 and 3, or the picornavirus, originally known as duck hepatitis virus type 1 [109]. Similarly, histological lesions consistent with nephritis in chickens can be caused by ANV, infectious bronchitis virus (IBV), and CAstV (Todd et al., unpublished results). In cases of hepatitis and nephritis, the presence of a specific astrovirus needs to be demonstrated in the affected tissue, preferably at the site of the histological lesion. This can be achieved using immunohistochemistry (IHC), undertaken with paraffin-embedded formalin-fixed tissue, or immunofluorescence (IF) performed with frozen tissue or tissue impression smears. However, due to the limited availability of avian astrovirus-specific antibodies, to date these methods have received limited application. IF is used to diagnose hepatitis in ducklings caused by DAstV-2 [24]. Very recently, a prototype immunoperoxidase-based IHC method has been applied to demonstrate the presence of CAstV antigen in nephritis-associated histological lesions (Todd et al., unpublished results; Fig. 9.8). This work suggested that CAstV alone was capable of causing nephritis in chickens and highlighted the need to identify the virus responsible in cases of nephritis and visceral gout. Imada et al. has reported that IF is a useful diagnostic procedure for ANV, detecting the group antigens and staining infected kidneys during the early acute phase of the disease [26]. In situ hybridization (ISH) can also be used to detect the presence of viral nucleic acid at the site of the histological lesion. Because probes generated from cloned virus-specific cDNA fragments may be more easily produced than virus-specific antibodies, ISH may prove to be an easier approach to develop. ISH has been used to demonstrate the presence of TAstV-2 RNA within the enterocytes present on the sides of the villi from virus-infected turkeys, but its use to detect other avian astroviruses has not yet been reported [5, 41, 68].

Photomicrograph of a kidney section from a broiler chicken naturally infected with CAstV; immunohistochemistry (IHC) staining for detection of the virus. Viral antigen staining present in kidney epithelium (dark brown); immunoperoxidase labeling, hematoxylin counterstain. Magnification 200×
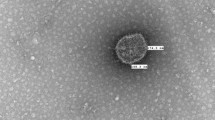
Diagnosing astrovirus infections is considerably easier than diagnosing astrovirus-associated disease. The method first used was negative contrast electron microscopy (EM), which depended on the visualization of the characteristic five- and six-pointed star morphology that characterizes astroviruses. EM performed with enteric samples has been used for the diagnosis of TAstVs and for the diagnosis of DAstV-1 in liver samples from ducks with hepatitis [23, 56]. However, astrovirus diagnosis by EM has a number of drawbacks, the most important being that some avian astroviruses fail to display the characteristic star morphology. This appears to be the case for ANV, DAstV-2, and CAstV, which are detected as small spherical viruses without obvious surface structure, and, on this basis, these were originally considered to be picornaviruses (ANV) or enterovirus-like viruses (CAstV, DAstV-2) [57]. Additional drawbacks of EM include its relative lack of sensitivity, and its unsuitability for the testing of large samples due to the time, labor, and equipment required.
Virus isolation in embryonated eggs or cell culture has been successful for most of the avian astroviruses including ANV, CAstV, TAstV, and DAstV. TAstV and DAstV can be propagated in turkey and duck embryos, respectively, and sometimes in chick embryos as well [98, 109]. Although the propagation of DAstV-1 (formerly DHV-2) in cell culture has been reported [4], the propagation of TAstV in cell cultures has not. The growth of high yields of TAstV isolates in turkey embryos has facilitated the production of purified virus, with which virus-specific antibodies have been prepared [97]. The astroviruses from chickens can be propagated in chicken embryos and in cell culture. CAstVs can be propagated in SPF chick embryos following yolk sac inoculation to produce variable effects including early embryo death, dwarfing, hemorrhages, edema, and feather changes [91]. ANVs can be propagated in SPF embryos following egg inoculation using a variety of routes. Isolation of ANV in cell cultures has been reported in a number of studies [19, 25, 110], and induces a cell rounding CPE in primary chicken kidney cells typically within 72 h post infection [26] . Experience from some laboratories suggests that some ANVs may be non-cultivable and the inconsistent growth of some ANVs in primary cell culture and embryos has prompted speculation that embryos containing ANV antibody may inhibit in vitro propagation (Todd et al. unpublished results). Baxendale and Mebatsion have reported the isolation and propagation of CAstVs in primary chick embryo liver cells and LMH cells, in which plaques similar to those produced by DAstV (DHV-2) were obtained [4]. The CAstV isolates have been used in virus neutralization tests, with which antibody to CAstV was detected [4]. There is also evidence that CAstVs can non-productively infect when inoculated into primary chick embryo liver cells [105]. In these cases, the inoculated CAstV undergoes partial replication, producing virus proteins that can be detected by indirect IF without producing infectious virus and CPE [105]. The effects produced by particular avian astroviruses in cell culture or in experimentally inoculated embryos are relatively nonspecific and cannot easily be distinguished from the effects caused by other avian astroviruses or other viruses. As such, the cultured virus needs to be identified; this can be accomplished through the detection of virus-specific antigens or nucleic acid. Although cultivable virus isolates are essential for biological characterization and pathogenesis studies, virus isolation is not routinely used to detect avian astroviruses, the principal reason being that astrovirus-containing samples are frequently contaminated with viruses such as adenoviruses or reoviruses, which replicate faster than the astroviruses in cell culture and/or in embryos. In addition, virus isolation and identification is time-consuming and labor-intensive, making it unsuitable for testing large samples.
A microtiter plate-based ELISA has been developed for the detection of TAstV-2 antigen in feces or intestinal content samples [97]. This test depended on the use of rabbit polyclonal antibodies that were raised by immunization with two different TAstV-2, which had been purified following replication in turkey embryos. Results compared favorably with those obtained by EM in terms of the numbers of positive samples recorded. Despite their convenience and low cost, the absence of virus-specific antibodies for other avian astroviruses has restricted the development of antigen-detecting ELISAs.
Attention has focused increasingly on the use of molecular diagnostic tests for detecting avian astroviruses. Since the first report describing an RT-PCR test for detecting TAstV [43], conventional and/or real-time tests have been described for ANV [14, 54, 103], CAstV [70]; [89], and DAstV [6] as well as TAstV-1 and TAstV-2 [14, 41, 93, 97]. The primers for these tests are usually located in the genomic regions encoding nonstructural proteins such as the RNA polymerase (ORF 1b), which are more conserved than the capsid protein encoding region (ORF 2). However, the high levels of sequence diversity displayed by avian astroviruses species make it difficult to select primers that will detect all variants within one type. Therefore, primer design should be based on comparing sequences from as large a number of virus variants as possible and the need to incorporate degenerate primers recognized. The importance of primer selection was highlighted when it was shown that the RT-PCR test using primers located in the 3′ untranslated region (3′ UTR) was more successful in detecting ANV-positive samples than tests based on primers located in the nonstructural protease or RNA polymerase [89, 103]. Similarly, CAstV from all genetic groupings was detected by the conventional and real-time RT-PCR tests described by Smyth et al. [88, 89], whereas one CAstV grouping was not detected by another published test [14]. Applications of these molecular tests have demonstrated that TAstV, ANV, and CAstV are highly prevalent in samples from their respective hosts, and there is a strong likelihood that the vast majority of commercial chicken and turkey flocks will become infected at some point in their lifetimes [18, 69, 70, 89]. Therefore, since all enteric samples are likely to be positive using these highly sensitive RT-PCR methods, knowing that an enteric sample is astrovirus positive is of questionable importance. However, with regard to the impact that the infection may be having on the flock or bird, there may be considerably more value in knowing the amounts of virus present, and in knowing whether internal tissues such as the kidney or liver are virus positive.
In comparison with other avian viruses, the serological diagnoses of avian astroviruses have received relatively little attention. A major reason for this is the limited number of avian astrovirus strains that have been adapted to cell culture. Although a number of ANV isolates can be propagated in cell culture, with very few exceptions cell culture isolations of CAstVs, TAstVs, and DAstVs have not been described. The lack of cell culture methods limits the development of indirect IF assays and virus neutralization tests to analyze chicken, turkey, and duck sera for antibodies. Additionally, the development of other serological assays such as ELISAs requires the ability to produce ample quantities of virus and virus antigen. With limited cell culture propagation methods, only a few laboratories have developed these assays for understanding astrovirus serology. An ELISA for detecting ANV antibody was described by Decaesstecker and Meulemans [16]. Sellers et al. have reported the use of an ELISA for detecting antibodies to CAstV using recombinantly expressed capsid protein [83]. Wang et al. [108] and Zhao et al. [114] also developed similar ELISA tests using recombinant capsid proteins for detecting DAstV and ANV-specific antibodies in duck and chicken serum samples.
The antigenic diversities displayed by avian astrovirus species need to be taken into consideration when using serological tests. For example, Shirai et al. [85] have demonstrated that ANV strains representing serotype 1 (ANV-1) and serotype 2 (ANV-2) exhibit very low levels of cross-reactive antibodies using indirect IF. Todd et al. [104] have recently reported the common occurrence of ANV variants, which displayed very different capsid protein sequences from those of ANV-1 and ANV-2, and which, on this basis, are likely to be antigenically different. This view is supported by a recent work by de Wit et al. [16], who showed that serum samples containing antibodies to a novel ANV isolate (isolate 19) were not detected by an ANV-1 specific indirect IF assay. It is recognized that the CAstVs can be subdivided into two major antigenic groups, designated A and B, which display relatively low levels of cross-reactivity by indirect IF [90, 105]. For this reason, a recent CAstV seroprevalence study involved the application of two separate indirect IF tests, one based on isolate 612 for detecting antibodies to the CAstVs from Group A, and the second based on isolate 11672 for detecting antibodies to the Group B CAstVs [105]. Therefore, the antigenic diversities exhibited by ANV and CAstV need to be considered in relation to their serological diagnosis. Serological tests based on the use of single ANV or CAstV strains will not detect antibodies to all ANV or CAstV infections. This is of particular importance when testing SPF flocks to demonstrate freedom from infection.
Control
Although it has been possible to produce and maintain specific pathogen free (SPF) flocks that are free from ANV infection as determined by serological testing, the common and widespread occurrence of avian astroviruses such as ANV, CAstV, and TAstVs in commercial poultry, and, in the case of ANV, their occurrence in wild birds [45], when combined with their potential for vertical transmission, strongly suggests that their eradication from commercial poultry would not be feasible. In addition, astroviruses are very stable in the environment and resistant to inactivation by most routinely used disinfectants, which complicates its elimination from infected premises [82]. Strict biosecurity, increased down time between flocks, and effective disinfection of the premises would help reduce the likelihood of contracting avian astrovirus infections.
Convalescent antisera have been used to control outbreaks of DAstV-1 and DAstV-2 [109]. Although experimental live attenuated vaccines were also developed for these astroviruses, these were not commercially produced [109]. Similarly, commercial vaccines have not been developed for ANV, CAstV, and TAstVs. In the case of ANV and CAstV, which are known to be vertically transmitted, vaccine protocols focused on the parent flocks during rear should be considered as a method to prevent or limit transmission to progeny in ovo. In addition, breeder vaccination should ensure uniformly higher levels of maternal antibody, which would better equip the young birds to withstand early viral challenges. The development of live attenuated or inactivated avian astrovirus vaccines will both depend on the availability of cell or embryo culture methods. Alternatively, recombinant expression systems, such as recombinant baculovirus system, might be used to produce astrovirus capsid subunit vaccines. Sellers et al. [83] recently described the production of a prototype recombinant baculovirus-derived CAstV vaccine, which, following multiple administrations to breeders, offered some protection to progeny chicks against experimentally induced RSS.
Conclusions
In comparison with other avian viruses, such as fowl adenoviruses, avian paramyxoviruses, and avian coronaviruses, relatively little is known about avian astroviruses and their impact on birds. A major reason for this has been the absence in most cases of cell culture growth systems, with which viruses can be isolated and biologically characterized; however, the level of genetic diversity, species affected, and permutations of clinical diseases caused by the various members of Astroviridae complicate our ability to develop a cohesive narrative as other avian viruses. In spite of these challenges, within the last decade, considerable advances have been made. The different avian astroviruses are second only to the human astroviruses in terms of their level of characterization, and in many cases, such a pathogenic mechanism and immunity are the best characterized. Regardless, considerably more research is needed.
Most notably, a standardized process for naming and classifying both existing as well as new members of the genera needs to be developed. As avian virology laboratories continue to develop new detection methods, and identify and characterize novel avian astroviruses in commercial poultry as well as wild bird species, a more consistent method of comparing (molecularly or antigenically) would aid in our ability to understand the relationship of viruses within the genera as well as identifying potential virulence determinants. As part of this effort to better understand avian astroviruses, the avian virology community should be encouraged to more fully characterize representative members of each genotype, serotype, and subgroup. This would include not only sequencing, but also the development of antibodies and other reagents. Together these efforts would all be in support of increasing our understanding of the cellular, physiological, and immunological changes astroviruses induce in their various host species so that we can develop more effective prevention, control, and therapeutic strategies.
References
Asplin FD. Duck hepatitis: vaccination against two serological types. Vet Rec. 1965;77:1529–30.
Barnes GL, Guy JS, Vaillancourt JP. Poult enteritis complex. Rev Sci Tech. 2000;19:565–88.
Barnes GL, Guy JS. Poult enteritis-mortality syndrome. In: Saif YM, Barnes HJ, Glisson JR, Faddly AM, MacDougald LR, Swayne DE, editors. Diseases of poultry. 11th ed. Ames IA: Blackwell Publishing; 2003.
Baxendale W, Mebatsion T. The isolation and characterisation of astroviruses from chickens. Avian Pathol. 2004;33:364–70.
Behling-Kelly E, Schultz-Cherry S, Koci M, Kelley L, Larsen D, Brown C. Localization of astrovirus in experimentally infected turkeys as determined by in situ hybridization. Vet Pathol. 2002;39:595–8.
Bidin M, Lojkic I, Bidin Z, Tisljar M, Majnaric D, Mikec M. Detection and characterization of avian nephritis virus in ducklings. Avian Pathol. 2011;40:173–7.
Bosch A, Guix S, Krishna NK, Méndez E, Monroe SS, Pantin-Jackwood M, et al. Family Astroviridae. In: King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ, editors. Virus taxonomy: ninth report of the international committee on taxonomy of viruses. London: Elsevier Inc; 2012.
Canelli E, Cordioli P, Barbieri I, Catella A, Pennelli D, Ceruti R, et al. Astroviruses as causative agents of poultry enteritis: genetic characterization and longitudinal studies on field conditions. Avian Dis. 2012;56(1):173–82.
Cattoli G, De Battisti C, Toffan A, Salviato A, Lavazza A, Cerioli M, et al. Co-circulation of distinct genetic lineages of astroviruses in turkeys and guinea fowl. Arch Virol. 2007;152:595–602.
Cattoli G, Toffan A, De Battisti C, Salviato A, Terregino C, Capua I. Astroviruses found in the intestinal contents of guinea fowl suffering from enteritis. Vet Rec. 2005;156:220.
Chu DK, Chin AW, Smith GJ, Chan KH, Guan Y, Peiris JS, et al. Detection of novel astroviruses in urban brown rats and previously known astroviruses in humans. J Gen Virol. 2010;91:2457–62.
Chu DK, Poon LL, Guan Y, Peiris JS. Novel astroviruses in insectivorous bats. J Virol. 2008;82:9107–14.
Connor TJ, McNeilly F, McFerran JB, McNulty MS. A survey of avian sera from Northern Ireland for antibody to avian nephritis virus. Avian Pathol. 1987;16:15–20.
Day JM, Spackman E, Pantin-Jackwood M. A multiplex RT-PCR test for the differential identification of turkey astrovirus type 1, turkey astrovirus type 2, chicken astrovirus, avian nephritis virus, and avian rotavirus. Avian Dis. 2007;51:681–4.
de Wit JJ, Dam GB, de Laar JM, Biermann Y, Verstegen I, Edens F, et al. Detection and characterization of a new astrovirus in chicken and turkeys with enteric and locomotion disorders. Avian Pathol. 2011;40:453–61.
Decaesstecker M, Meulemans. An ELISA for the detection of antibodies to avian nephritis virus and related entero-like viruses. Avian Pathol. 1991;20:523–30.
Decaesstecker M, Charlier G, Peeters J, Meulemans G. Pathogenicity of fowl enteroviruses. Avian Pathol. 1989;18:697–713.
Domanska-Blicharz K, Seroka A, Minta Z. One-year molecular survey of astrovirus infection in turkeys in Poland. Arch Virol. 2011;156:1065–72.
Frazier JA, Howes K, Reece RL, Kidd AW, Cavanagh D. Isolation of noncytopathic viruses implicated in the aetiology of nephritis and baby chick nephropathy and serologically related to avian nephritis virus. Avian Pathol. 1990;19:139–60.
Fu Y, Pan M, Wang X, Xu Y, Xie X, Knowles NJ, et al. Complete sequence of a duck astrovirus associated with fatal hepatitis in ducklings. J Gen Virol. 2009;90:1104–8.
Gibson CA, Chen J, Monroe SA, Denison MR. Expression and processing of nonstructural proteins of the human astroviruses. Adv Exp Med Biol. 1998;440:387–91.
Gough RE, Borland ED, Keymer IF, Stuart JC. An outbreak of duck hepatitis type II in commercial ducks. Avian Pathol. 1985;14:227–36.
Gough RE, Collins MS, Borland E, Keymer LF. Astrovirus-like particles associated with hepatitis in ducklings. Vet Rec. 1984;114:279.
Haider SA, Calnek BW. In-vitro isolation, propagation and characterisation of duck hepatitis virus type III. Avian Dis. 1979;23:715–29.
Hewson KA, O’Rourke D, Noormohammadi AH. Detection of avian nephritis virus in Australian chicken flocks. Avian Dis. 2010;54:990–3.
Imada T. Avian nephritis. In: Saif YM, Faddly AM, Glisson JR, MacDougald LR, Nolan LK, Swayne DE, editors. Diseases of poultry. Ames, IA: Blackwell Publishing; 2008.
Imada T, Maeda M, Furuta K, Yamaguchi S, Kawamura H. Pathogenicity and distribution of avian nephritis virus (G-4260 strain) in inoculated laying hens. Natl Inst Anim Health Q (Tokyo). 1983;23:43–8.
Imada T, Taniguchi T, Yamaguchi S, Minetoma T, Maeda M, Kawamura H. Susceptibility of chickens to avian nephritis virus at various inoculation routes and ages. Avian Dis. 1981;25:294–302.
Imada T, Yamaguchi S, Kawamura H. Pathogenicity for baby chicks of the G-4260 strain of the picornavirus “avian nephritis virus”. Avian Dis. 1979;23:582–8.
Imada T, Yamaguchi S, Mase M, Tsukamoto K, Kubo M, Morooka A. Avian nephritis virus (ANV) as a new member of the family Astroviridae and construction of infectious ANV cDNA. J Virol. 2000;74:8487–93.
Imada T, Yamaguchi S, Miura N, Kawamura H. Antibody survey against avian nephritis virus among chickens in Japan. Natl Inst Anim Health Q (Tokyo). 1980;20:79–80.
Jiang B, Monroe SS, Koonin EV, Stine SE, Glass RI. RNA sequence of astrovirus: distinctive genomic organization and a putative retrovirus-like ribosomal frameshifting signal that directs the viral replicase synthesis. Proc Natl Acad Sci USA. 1993;90:10539–43.
Jindal N, Patnayak DP, Chander Y, Ziegler AF, Goyal SM. Comparison of capsid gene sequences of turkey astrovirus-2 from poult-enteritis-syndrome-affected and apparently healthy turkeys. Arch Virol. 2011;156:969–77.
Jindal N, Patnayak DP, Chander Y, Ziegler AF, Goyal SM. Detection and molecular characterization of enteric viruses from poult enteritis syndrome in turkeys. Poult Sci. 2010;89:217–26.
Jindal N, Patnayak DP, Ziegler AF, Lago A, Goyal SM. Duration of growth depression and pathogen shedding in experimentally reproduced poult enteritis syndrome. Avian Dis. 2009;53:517–22.
Jindal N, Patnayak DP, Ziegler AF, Lago A, Goyal SM. Experimental reproduction of poult enteritis syndrome: clinical findings, growth response, and microbiology. Poult Sci. 2009;88:949–58.
Jindal N, Patnayak DP, Ziegler AF, Lago A, Goyal SM. A retrospective study on poult enteritis syndrome in Minnesota. Avian Dis. 2009;53:268–75.
Jonassen CM, Jonassen TT, Sveen TM, Grinde B. Complete genomic sequences of astroviruses from sheep and turkey: comparison with related viruses. Virus Res. 2003;91:195–201.
Kang KI, Icard AH, Linnemann E, Sellers HS, Mundt E. Determination of the full length sequence of a chicken astrovirus suggests a different replication mechanism. Virus Genes. 2012;44(1):45–50.
Koci MD, Kelley LA, Larsen D, Schultz-Cherry S. Astrovirus-induced synthesis of nitric oxide contributes to virus control during infection. J Virol. 2004;78:1564–74.
Koci MD, Moser LA, Kelley LA, Larsen D, Brown CC, Schultz-Cherry S. Astrovirus induces diarrhea in the absence of inflammation and cell death. J Virol. 2003;77:11798–808.
Koci MD, Schultz-Cherry S. Avian astroviruses. Avian Pathol. 2002;31:213–27.
Koci MD, Seal BS, Schultz-Cherry S. Development of an RT-PCR diagnostic test for an avian astrovirus. J Virol Methods. 2000;90:79–83.
Koci MD, Seal BS, Schultz-Cherry S. Molecular characterization of an avian astrovirus. J Virol. 2000;74:6173–7.
Kofstad T, Jonassen CM. Screening of feral and wood pigeons for viruses harbouring a conserved mobile viral element: characterization of novel astroviruses and picornaviruses. PLoS One. 2011;6:e25964.
Krishna NK. Identification of structural domains involved in astrovirus capsid biology. Viral Immunol. 2005;18:17–26.
Kurtz J, Lee T. Astrovirus gastroenteritis age distribution of antibody. Med Microbiol Immunol. 1978;166:227–30.
Lewis TL, Matsui SM. An astrovirus frameshift signal induces ribosomal frameshifting in vitro. Arch Virol. 1995;140:1127–35.
Lewis TL, Matsui SM. Astrovirus ribosomal frameshifting in an infection-transfection transient expression system. J Virol. 1996;70:2869–75.
Lewis TL, Matsui SM. Studies of the astrovirus signal that induces (-1) ribosomal frameshifting. Adv Exp Med Biol. 1997;412:323–30.
MacMicking J, Xie QW, Nathan C. Nitric oxide and macrophage function. Annu Rev Immunol. 1997;15:323–50.
Maeda M, Imada T, Taniguchi T, Horiuchi T. Pathological changes in chicks inoculated with the picornavirus “avian nephritis virus”. Avian Dis. 1979;23:589–96.
Mandoki M, Bakonyi T, Ivanics E, Nemes C, Dobos-Kovacs M, Rusvai M. Phylogenetic diversity of avian nephritis virus in Hungarian chicken flocks. Avian Pathol. 2006;35:224–9.
Mandoki M, Dobos-Kovacs M, Bakonyi T, Rusvai M. Molecular diagnosis of avian nephritis: preliminary report. Acta Vet Hung. 2006;54:51–60.
Marczinke B, Bloys AJ, Brown TD, Willcocks MM, Carter MJ, Brierley I. The human astrovirus RNA-dependent RNA polymerase coding region is expressed by ribosomal frameshifting. J Virol. 1994;68:5588–95.
McNeilly F, Connor TJ, Calvert VM, Smyth JA, Curran WL, Morley AJ, Thompson D, Singh S, McFerran JB, Adair BM, McNulty MS. Avian Pathol. 1994 Jun;23(2):313-27.
McNulty MS, Curran WL, McFerran JB. Detection of astroviruses in turkey faeces by direct electron microscopy. Vet Rec. 1980;106:561.
McNulty MS, Connor TJ, McNeilly F, McFerran JB. Biological characterization of avian enteroviruses and enterovirus-like viruses. Avian Pathol. 1990;19:75–87.
McNulty MS, Connor TJ, McNeilly F. A survey of specific pathogen-free chicken flocks for antibodies to chicken anemia agent, avian nephritis virus and group A rotavirus. Avian Pathol. 1989;18:215–20.
Meyerhoff RR, Nighot PK, Ali RA, Blikslager AT, Koci MD. Characterization of turkey inducible nitric oxide synthase and identification of its expression in the intestinal epithelium following astrovirus infection. Comp Immunol Microbiol Infect Dis. 2012;35(1):63–9.
Mitchell DK. Astrovirus gastroenteritis. Pediatr Infect Dis J. 2002;21:1067–9.
Mockett AP, Huggins MB, Woods M, Orbell S. A compariosn of three serological methods to detect chicken and turkey antibodies to avian nephritis virus and the use of virus-specific monoclonal antibodies. Avian Pathol. 1993;22(4):751–70.
Monceyron C, Grinde B, Jonassen TO. Molecular characterisation of the 3′-end of the astrovirus genome. Arch Virol. 1997;142:699–706.
Mor SK, Abin M, Costa G, Durrani A, Jindal N, Goyal SM, et al. The role of type-2 turkey astrovirus in poult enteritis syndrome. Poult Sci. 2011;90:2747–52.
Moser LA, Carter M, Schultz-Cherry S. Astrovirus increases epithelial barrier permeability independently of viral replication. J Virol. 2007;81:11937–45.
Nicholas RAJ, Goddard RD, Luff PR. Prevalence of avian nephritis virus in England. Vet Rec. 1988;123:398.
Nighot PK, Moeser A, Ali RA, Blikslager AT, Koci MD. Astrovirus infection induces sodium malabsorption and redistributes sodium hydrogen exchanger expression. Virology. 2010;401:146–54.
Pantin-Jackwood MJ, Day JM, Jackwood MW, Spackman E. Enteric viruses detected by molecular methods in commercial chicken and turkey flocks in the United States between 2005 and 2006. Avian Dis. 2008;52:235–44.
Pantin-Jackwood MJ, Spackman E, Day JM. Pathogenesis of type 2 turkey astroviruses with variant capsid genes in 2-day-old specific pathogen free poults. Avian Pathol. 2008;37:193–201.
Pantin-Jackwood MJ, Spackman E, Day JM, Rives D. Periodic monitoring of commercial turkeys for enteric viruses indicates continuous presence of astrovirus and rotavirus on the farms. Avian Dis. 2007;51:674–80.
Pantin-Jackwood MJ, Spackman E, Woolcock PR. Molecular characterization and typing of chicken and turkey astroviruses circulating in the United States: implications for diagnostics. Avian Dis. 2006;50:397–404.
Pantin-Jackwood MJ, Spackman E, Woolcock PR. Phylogenetic analysis of turkey astroviruses reveals evidence of recombination. Virus Genes. 2006;32:187–92.
Pantin-Jackwood MJ, Strother KO, Mundt E, Zsak L, Day JM, Spackman E. Molecular characterization of avian astroviruses. Arch Virol. 2011;156:235–44.
Qureshi MA, Saif YM, Heggen-Peay CL, Edens FW, Havenstein GB. Induction of functional defects in macrophages by a poult enteritis and mortality syndrome-associated turkey astrovirus. Avian Dis. 2001;45:853–61.
Qureshi MA, Yu M, Saif YM. A novel “small round virus” inducing poult enteritis and mortality syndrome and associated immune alterations. Avian Dis. 2000;44:275–83.
Reynolds DL, Saif YM. Astrovirus: a cause of an enteric disease in turkey poults. Avian Dis. 1986;30:728–35.
Reynolds DL, Saif YM, Theil KW. Enteric viral infections of turkey poults: incidence of infection. Avian Dis. 1987;31:272–6.
Reynolds DL, Saif YM, Theil KW. A survey of enteric viruses of turkey poults. Avian Dis. 1986;31:89–98.
Reynolds DL, Schultz-Cherry SL. Astrovirus infections. In: Saif YM, Faddly AM, Glisson JR, MacDougald LR, Nolan LK, Swayne DE, editors. Diseases of poultry. Ames, IA: Blackwell Publishing; 2008.
Saif LJ, Saif YM, Theil KW. Enteric viruses in diarrheic turkey poults. Avian Dis. 1985;29:798–811.
Saif YM, Saif LJ, Hofacre CL, Hayhow C, Swayne DE, Dearth RN. A small round virus associated with enteritis in turkey poults. Avian Dis. 1990;34:762–4.
Schultz-Cherry S, Kapczynski DR, Simmons VM, Koci MD, Brown C, Barnes HJ. Identifying agent(s) associated with poult enteritis and mortality syndrome: importance of the thymus. Avian Dis. 2000;44:256–65.
Schultz-Cherry S, King DJ, Koci MD. Inactivation of an astrovirus associated with poult enteritis mortality syndrome. Avian Dis. 2001;45:76–82.
Sellers H, Linneman E, Icard AH, Mundt E. A purified recombinant baculovirus expressed capsid protein of a new astrovirus provides partial protection to runting-stunting syndrome in chickens. Vaccine. 2010;28:1253–63.
Shirai J, Nakamura K, Narita M, Furuta K, Hihara H, Kawamura H. Visceral urate deposits in chicks inoculated with avian nephritis virus. Vet Rec. 1989;124:658–61.
Shirai J, Nakamura K, Shinohara K, Kawamura H. Pathogenicity and antigenicity of avian nephritis isolates. Avian Dis. 1991;35:49–54.
Shirai J, Tanimura N, Uramoto K, Narita M, Nakamura K, Kawamura H. Pathologically and serologically different avian nephritis virus isolates implicated in etiology of baby chick nephropathy. Avian Dis. 1992;36:369–77.
Smyth JA, Connor TJ, McNeilly F, Moffet DA, Calvert VM, McNulty MS. Studies on the pathogenicity of enterovirus-like viruses in chickens. Avian Pathol. 2007;36:119–26.
Smyth VJ, Jewhurst HL, Adair BM, Todd D. Detection of chicken astrovirus by reverse transcriptase-polymerase chain reaction. Avian Pathol. 2009;38:293–9.
Smyth VJ, Jewhurst HL, Wilkinson DS, Adair BM, Gordon AW, Todd D. Development and evaluation of real-time TaqMan(R) RT-PCR assays for the detection of avian nephritis virus and chicken astrovirus in chickens. Avian Pathol. 2010;39:467–74.
Smyth VJ, Todd D, Trudgett J, Lee A, Welsh MD. Capsid protein sequence diversity of chicken astrovirus. Avian Pathol. 2012;41(2):151–9.
Spackman D, Gough RE, Collins MS, Lanning D. Isolation of an enterovirus-like agent from the meconium of dead-in-shell chicken embryos. Vet Rec. 1984;114:216–8.
Spackman E, Day JM, Pantin-Jackwood MJ. Astrovirus, reovirus, and rotavirus concomitant infection causes decreased weight gain in broad-breasted white poults. Avian Dis. 2010;54:16–21.
Spackman E, Kapczynski D, Sellers H. Multiplex real-time reverse transcription-polymerase chain reaction for the detection of three viruses associated with poult enteritis complex: turkey astrovirus, turkey coronavirus, and turkey reovirus. Avian Dis. 2005;49:86–91.
Strain E, Kelley LA, Schultz-Cherry S, Muse SV, Koci MD. Genomic analysis of closely related astroviruses. J Virol. 2008;82:5099–103.
Takase K, Shinohara K, Tsuneyoshi M, Yamamoto M, Yamada S. Isolation and characterization of cytopathic avian enteroviruses from broiler chickens. Avian Pathol. 1989;18:631–42.
Takase K, Uchimura T, Yamamoto M, Yamada S. Susceptibility of embryos and chicks, derived form immunized breeding hens, to avian nephritis virus. Avian Pathol. 1994;23:117–25.
Tang Y, Ismail MM, Saif YM. Development of antigen-capture enzyme-linked immunosorbent assay and RT-PCR for detection of turkey astroviruses. Avian Dis. 2005;49:182–8.
Tang Y, Murgia AM, Saif YM. Molecular characterization of the capsid gene of two serotypes of turkey astroviruses. Avian Dis. 2005;49:514–9.
Tang Y, Murgia MV, Ward L, Saif YM. Pathogenicity of turkey astroviruses in turkey embryos and poults. Avian Dis. 2006;50:526–31.
Tang Y, Saif YM. Antigenicity of two turkey astrovirus isolates. Avian Dis. 2004;48:896–901.
Thouvenelle ML, Haynes JS, Reynolds DL. Astrovirus infection in hatchling turkeys: histologic, morphometric, and ultrastructural findings. Avian Dis. 1995;39:328–36.
Todd D, Smyth VJ, Ball NW, Donnelly BM, Wylie M, Knowles NJ, et al. Identification of chicken enterovirus-like viruses, duck hepatitis virus type 2 and duck hepatitis virus type 3 as astroviruses. Avian Pathol. 2009;38:21–30.
Todd D, Trudgett J, McNeilly F, McBride N, Donnelly B, Smyth VJ, et al. Development and application of an RT-PCR test for detecting avian nephritis virus. Avian Pathol. 2010;39:207–13.
Todd D, Trudgett J, Smyth VJ, Donnelly B, McBride N, Welsh MD. Capsid protein sequence diversity of avian nephritis virus. Avian Pathol. 2011;40:249–59.
Todd D, Wilkinson DS, Jewhurst HL, Wylie M, Gordon AW, Adair BM. A seroprevalence investigation of chicken astrovirus infections. Avian Pathol. 2009;38:301–9.
Toth TE. Studies of an agent causing mortality among ducklings immune to duck virus hepatitis. Avian Dis. 1969;13:834–46.
Walter JE, Briggs J, Guerrero ML, Matson DO, Pickering LK, Ruiz-Palacios G, et al. Molecular characterization of a novel recombinant strain of human astrovirus associated with gastroenteritis in children. Arch Virol. 2001;146:2357–67.
Wang X, Wang Y, Xie X, Zhang B, Zhang D. Expression of the C-terminal ORF2 protein of duck astrovirus for application in a serological test. J Virol Methods. 2011;171:8–12.
Woolcock PR. Duck hepatitis. In: Saif YM, Faddly AM, Glisson JR, MacDougald LR, Nolan LK, Swayne DE, editors. Diseases of poultry. Ames, IA: Blackwell Publishing; 2008.
Yamaguchi S, Imada T, Kawamura H. Characterisation of a picornavirus isolated from broiler chicks. Avian Dis. 1979;23:571–81.
Yu M, Ismail MM, Qureshi MA, Dearth RN, Barnes HJ, Saif YM. Viral agents associated with poult enteritis and mortality syndrome: the role of a small round virus and a turkey coronavirus. Avian Dis. 2000;44:297–304.
Yu M, Tang Y, Guo M, Zhang Q, Saif YM. Characterization of a small round virus associated with the poult enteritis and mortality syndrome. Avian Dis. 2000;44:600–10.
Zhao W, Hua XG, Yuan L, Cui L, Shan TL, Dai XQ, et al. Sequence analyses of the representative Chinese-prevalent strain of avian nephritis virus in healthy chicken flocks. Avian Dis. 2011;55:65–9.
Zhao W, Zhu AL, Yu Y, Yuan CL, Zhu CX, Lan DL, et al. Segmentation expression of capsid protein as an antigen for the detection of avian nephritis virus infection in chicken flocks. J Virol Methods. 2012;179(1):57–61.
Zhao W, Zhu AL, Yu Y, Yuan CL, Zhu CX, Yang ZB, et al. Complete sequence and genetic characterization of pigeon avian nephritis virus, a member of the family Astroviridae. Arch Virol. 2011;156:1559–65.
Zhao W, Zhu AL, Yuan CL, Yu Y, Zhu CX, Lan DL, et al. Detection of astrovirus infection in pigeons (Columbia livia) during an outbreak of diarrhoea. Avian Pathol. 2011;40:361–5.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Science+Business Media New York
About this chapter
Cite this chapter
Pantin-Jackwood, M., Todd, D., Koci, M.D. (2012). Avian Astroviruses. In: Schultz-Cherry, S. (eds) Astrovirus Research. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-4735-1_9
Download citation
DOI: https://doi.org/10.1007/978-1-4614-4735-1_9
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-4734-4
Online ISBN: 978-1-4614-4735-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)