
Role of Decorin in the Lens and Ocular Diseases


1
Department of Ophthalmology, Kanazawa Medical University, Kanazawa 920-0293, Ishikawa, Japan
2
Department of Ophthalmology, University of Nebraska Medical Center, Omaha, NE 68198, USA
*
Author to whom correspondence should be addressed.
Cells 2023, 12(1), 74; https://doi.org/10.3390/cells12010074
Submission received: 27 October 2022
/
Revised: 15 December 2022
/
Accepted: 19 December 2022
/
Published: 24 December 2022
(This article belongs to the Special Issue New Advances in Lens Biology and Pathology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Decorin is an archetypal member of the small leucine-rich proteoglycan gene family and is involved in various biological functions and many signaling networks, interacting with extra-cellular matrix (ECM) components, growth factors, and receptor tyrosine kinases. Decorin also modulates the growth factors, cell proliferation, migration, and angiogenesis. It has been reported to be involved in many ischemic and fibrotic eye diseases, such as congenital stromal dystrophy of the cornea, anterior subcapsular fibrosis of the lens, proliferative vitreoretinopathy, et al. Furthermore, recent evidence supports its role in secondary posterior capsule opacification (PCO) after cataract surgery. The expression of decorin mRNA in lens epithelial cells in vitro was found to decrease upon transforming growth factor (TGF)-β-2 addition and increase upon fibroblast growth factor (FGF)-2 addition. Wound healing of the injured lens in mice transgenic for lens-specific human decorin was promoted by inhibiting myofibroblastic changes. Decorin may be associated with epithelial–mesenchymal transition and PCO development in the lens. Gene therapy and decorin administration have the potential to serve as excellent therapeutic approaches for modifying impaired wound healing, PCO, and other eye diseases related to fibrosis and angiogenesis. In this review, we present findings regarding the roles of decorin in the lens and ocular diseases.
1. Introduction
Decorin (DCN) belongs to a growing family of class I small leucine-rich proteoglycan proteins (SLRPs) that consist of a single glycosaminoglycan side chain (GAG), including a core protein containing leucine-rich repeats (LRR) with a molecular weight of approxi-mately 42 kDa [1]. Decorin is mainly synthesized by fibroblasts, stressed vascular endo-thelial cells, and smooth muscle cells. It is widely distributed in the stromal tissues and is expressed and secreted in various connective tissues, including the skin, bone, cartilage, ligaments, breast and vessel walls, and eyeballs, and in body fluids such as the plasma, aqueous humor, and vitreous humor [1,2,3,4,5,6,7,8,9,10,11,12,13,14]. Decorin is the main component of the extracellular matrix (ECM), together with collagen fibrils, and acts as a bridging molecule between type I and type VI collagen [15,16,17]. Decorin also regulates various cellular processes, including the ECM remodeling, fibrosis, cell proliferation, migration, differentiation, wound healing, autophagy, and angiogenesis. It also has essential inhibitory roles in inflammation, fibrotic disorders, and tumors [3,6,18,19,20,21,22,23,24,25,26]. Decorin interacts with and modulates multiple growth factors and signaling pathways, such as the transforming growth factor (TGF)-β pathway, fibroblast growth factor (FGF), platelet-derived growth factor (PDGF), and cellular communication network factor 2/connective tissue growth factor (CCN2/CTGF). Moreover, it directly blocks various receptor tyrosine kinase (RTK) family proteins, such as the epidermal growth factor receptor (EGFR), Erb4, and its other family members, insulin-like growth factor-I receptor (IGF-IR), vascular endothelial growth factor receptor 2 (VEGFR2) and the hepatocyte growth factor receptor 1 (HGFR1), and Toll-like receptors (TLRs) binding to multiple binds multiple cell surface receptors [4,5,26,27,28]. Decorin also acts as a reservoir for several different growth factors, sequestering signaling molecules in the ECM [29]. Thus, it is well known that decorin evokes potent tumor repression and wound-healing properties.
Here, we review the current knowledge of the role of decorin in the lens and ocular diseases. We hypothesize that decorin plays an important regulatory role in inhibiting ocular inflammatory and fibroblastic responses.
2. Structure and Function of Decorin
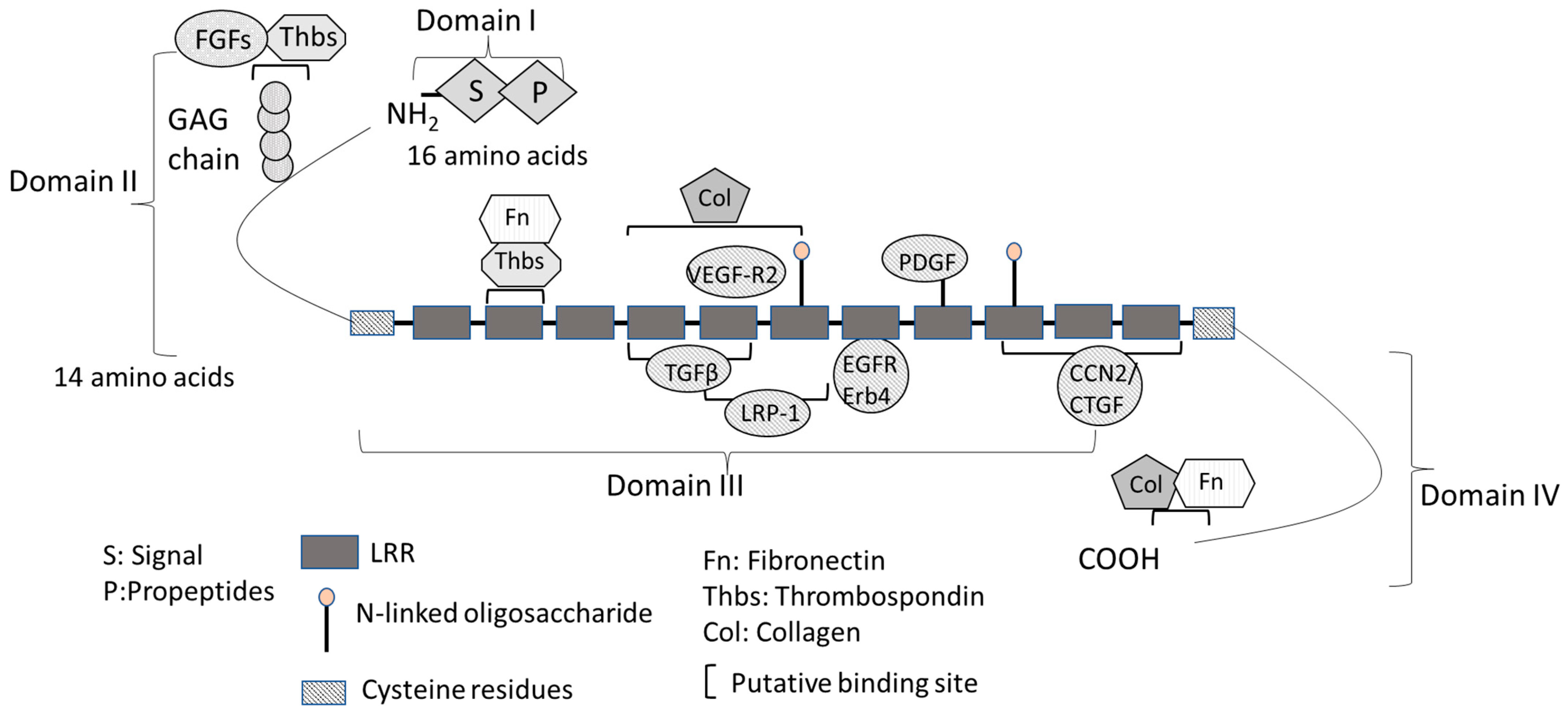
The structural and putative functional features of decorin are shown in Figure 1. The mature decorin molecule consists of a globular core protein and a covalently- linked GAG chain [30]. The primary translation product of human decorin is 359 amino acids long and can be divided into four functional domains (Figure 1) [30]. Domain I contains the initial 16-amino-acid long signal peptide directing the nascent protein to the rough endo plasmic reticulum, which is absent in the mature decorin protein [31]. Domain II contains 14 amino acid propeptide residues, regulating the attachment of GAGs composed of either dermatan or chondroitin sulfate [31]. GAG interacts with thrombospondins (Thbs), other GAGs, cytokines, and growth factors, such as TNF (tumor necrosis factor)-α and FGF1, 2, 7, and 8, regulating cell growth, wound healing, axon regeneration, and neural stem cell proliferation [32,33]. Decorin GAG may also inhibit blood coagulation by interacting with heparin cofactor II [34].
Domain III is the most important domain of decorin and has a major regulatory role by interacting with different signaling molecules [30]. Domain III consists of twelve tan-dem LRRs flanked by conserved cysteine (Cys)-rich domains and is important for its biological function. Decorin has a high-affinity binding site for collagen at LRRs 4–6 and a low-affinity site at the C-terminus for interacting with multiple collagen molecules simultaneously [31]. Decorin is also involved in the regulation of collagen fibrillogenesis. Mutant mice with a targeted disruption of the decorin gene (Dcn−/−) have fragile skin with abnormal collagen architecture in the dermis and reduced tensile strength, and showed a delay in epidermal and dermal wound healing [6,35,36]. Decorin has two binding sites for TGF-β isoforms at LRR4 and LRR5 with varying affinity in its core protein domain III, thus regulating TGF-β/ECM interactions (Figure 1) [37]. Decorin binds with high affinity to all isoforms of the TGF-β family and neutralizes their interaction with pro-fibrotic receptors [20]. LDL receptor-related protein 1 (LRP-1), an endocytic receptor for decorin involved in mediating TGF-β-dependent signaling, binds LRR5 and LRR6 [38,39,40]. CCN2/CTGF is an matricellular protein which that bridges the functional divide division between structural macromolecules and growth factors, which regulates regulating many cellular functions including cell adhesion, matrix production, structural remodeling, an-giogenesis, and cell proliferation, fibrosis, and differentiation [41]. CCN2/CTGF binds and induces growth factors (e.g., TGF-β and VEGF, thereby regulating their functions) and ECM proteins [41]. CCN2/CTGF also binds to LRR10-12, stimulating decorin and repressing the activity of important cytokines [42,43]. The LRR7 domain binds to EGFR and ErB4 and activates the mitogen-activated protein kinase (MAPK) pathway [44]. The LRR5 domain binds directly to VEGFR2 at a site that partially overlaps with the canonical binding site for VEGF-A [45]. Domain IV, the carboxyl terminus domain, contains binding sites for collagen fibrils and fibronectin (Fn) [30].
Although they are do not act as binding ligands for decorin, proteases play an im-portant role in the decorin interaction network. These proteases primarily include matrix metalloproteinases (MMP) 2, 3, and 7 [46,47,48]. MMP2, MMP3, and MMP7 induce the deg-radation of decorin, which is digested into fragments via interactions with its protein core [49]. This digestion of decorin allows the release of TGF-β1 [49]. MMP2 and MMP9 also activate TGF-β2 [50,51]. Thus, a single decorin molecule could simultaneously sequester multiple important mediators of cell growth and antagonize several signaling pathways crucial for fibrotic diseases.
3. Interaction between Decorin and TGF-β
TGF-β is considered a potent and ubiquitous multifunctional cytokine involved in wound repair, proliferation, migration, apoptosis, differentiation, and fibrosis, including a pleiotropic effect in immune response [52,53]. The expressions of TGF-β is increased in almost all fibrotic diseases involving all organ systems, including ocular diseases [54,55,56,57,58,59,60]. Three isoforms of TGF-β exist: TGFβ1, TGFβ2, and TGFβ3. They are secreted by many types of cells and are involved in a wide range of actions, including the promotion of cell differentiation, promotion or inhibition of cell proliferation, induction of ECM production, immunosuppression, and increase in mitochondrial reactive oxygen species (ROS) production [59]. TGF-β is also considered a potent immunosuppressive cytokine and is considered a target for immunotherapy to suppresses tissue rejection and prolong corneal allograft survival after penetrating keratoplasty [53,61]. The intracellular signaling of the TGF-β family is regulated by Smads. TGF-β signaling through the canonical Smad pathway has been well-described for its regulation of multiple target genes, including fibrosis inducible genes [60]. Smad2 and Smad3 are the two major downstream regulators, while Smad7 serves as a negative feedback regulator of the TGF-β/Smad pathway [62]. The canonical TGF-β signaling pathway involves ligand-receptor complex formation, the activation of kinase domains within the cell membrane receptors and its subsequent phosphorylation cascade, and the activation of intracellular signaling downstream of Smad-2/3 [63], which are transcriptional regulators that accumulate in the nucleus. Thus, Smad-2/3 both transmit signals into the nucleus and regulate gene transcription by binding to Smad-binding elements present in the promoters of their target genes [63,64]. This phenomenon has been demonstrated in several cell types in vitro and in vivo [40,48,49]. Besides Smad pathways, other non-canonical pathways of TGF-β signaling, including the MAPK, phosphatidylinositol-3-kinase (PI3K), and Rho-like GTPase pathways, are also crucial for the effect of TGF-β on different pathways [65,66,67]. These non-canonical path-ways of TGF-β signaling indirectly regulate gene transcription in the nucleus as activators of Smad-dependent pathways [52]. Moreover, TGF-β upregulates epithelial–mesenchymal transition (EMT) related genes, such as fibronectin, α-smooth muscle actin (αSMA), and tropomyo-sin (Tpm) [7,68,69,70]. Decorin regulates the Smad pathway by interacting with LRP1, an endocytic receptor for decorin [44]. In addition, decorin can bind to TGF-β and inhibit the binding of TGF-β ligands to their receptors. The bound complex formation with decorin prevents the binding of TGF-β to its receptor, repressing its activity [30].
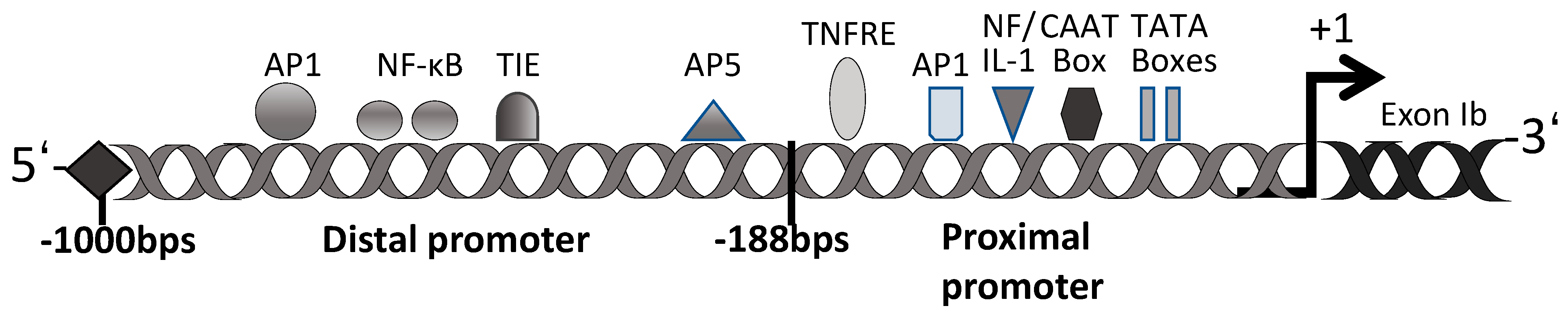
Many previous studies have confirmed the regulatory role of decorin for TGF-β activ-ity [71,72]. Our and other previous studies have demonstrated that TGF-β regulates gene transcription, and some genes, including the decorin gene, contain putative TGF-β-negative cis-acting elements (TIE) [73,74,75,76,77,78]. The promoter of decorin is divided into two main regions: a proximal promoter at 188 bp and a distal promoter at ~800 bp (Figure 2). The proximal promoter contains two functional TATA and CAAT boxes [79]. This region also contains two TNF-α-responsive elements (TNFRE) that mediate the TNF-α-induced transcriptional repression of the decorin gene [80]. The proximal promoter region also contains an activator protein 1 (AP1) binding site, a dual regulator of decorin gene expression repression by TNF-α and induction by interleukin-1 (NF/IL-1) [81]. The distal promoter of decorin has many homologous cis-acting factors, including AP1, AP5, and nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB), several direct repeat factors, and a TIE [79]. It has been reported that the TIE at position −685 of the promoter region of the decorin gene exon Ib is a transcriptional silencer [76,77,78]. This TIE has been found in various protease gene sequences that are downregulated by TGF-β and could inhibit decorin gene transcription [78]. Supporting this regulatory mechanism, we reported that decorin expression was downregulated in lens epithelial cells (LECs) and rabbit articular chondrocytes after the addition of TGF-β2 [7,82]. On the other hand, the expression of decorin was reported to be upregulated in mesangial cells and rat liver cells, and after treatment with TGF-β [83]. These results show that the molecular mechanisms for decorin induction may differ among cell types and that many aspects of the underlying regulatory mechanisms are still currently unknown.
The ocular tissues in the eyes are susceptible to fibrotic diseases, such as primary an-terior subcapsular fibrosis (ASF) in cataracts, secondary posterior capsule opacification (PCO) after cataract surgery, scarring in the cornea and conjunctiva , primary open-angle glaucoma (POAG), age-related macular degeneration (AMD), proliferative diabetic reti-nopathy (PDR), and proliferative vitreoretinopathy (PVR) [2,57,84,85,86,87,88,89]. TGF-β signaling is involved in the pathogenesis of fibrotic eye diseases, impairing the quality of vision and intra- and extra-ocular homeostasis [2,57,84,85,86,87,88,89]. Physiologically, TGF-β is mainly pro-duced in the ciliary epithelium, LECs, and retinal pigment epithelial cells (RPEs) as a la-tent, inactive form of mature TGF-β [90,91,92,93]. TGF-β2 was also detected in human tears [94]. TGF-β2 levels change in the aqueous humor during the clinical course of various ophthalmic diseases. For example, changes in TGF-β2 levels in the eyes of patients with PVR could cause post-retinal detachment and retinal fibrosis. In addition, TGF-β2 concentrations in the vitreous fluid increase as retinal fibrosis progresses [95]. Levels of total and active TGF-β2 are higher in cataract patients with diabetic retinopathy, neovascular glaucoma, and POAG than in cataract patients without these ocular diseases [96,97,98,99]. Altered decorin expression is also associated with several fibrotic eye diseases, including congenital stromal dystrophy of the cornea [100,101], corneal wound healing [102], PCO [2,7], glaucoma [10,88], and PVR [103]. Thus, interactions between decorin and TGF-β may be involved in the pathogenesis of ocular diseases that involve fibrotic changes.
4. Distribution of Decorin in the Eye
Decorin is mainly localized to the connective tissue and extracellular matrix and primarily involves collagen fiber formation. In the eyes, decorin is expressed in all ocular tissues such as the corneal stroma, conjunctival stroma, trabecular meshwork, lens, and in all layers of the retina, optic nerve, and sclera. It is secreted in the tears, aqueous humor, and vitreous humor [1,2,7,9,13,14,88,103,104]. Decorin is typically synthesized and se-creted by fibroblasts, maintaining the balance of ECM homeostasis [105]. However, epi-thelial and endothelial cells can also synthesize decorin [105]. Recent work has shown that decorin expression is conisderably upregulated in LECs during PCO and pseudoexfoliation glaucoma (PXG), in the aqueous humor of subjects in PDR [2,7,9,106], and in the vitreous humor of patients with rhegmatogenous retinal detachment and PVR [103,107]. On the other hand, decorin protein levels were also markedly decreased in the aqueous and vitreous humor in patients with POAG [88]. We previously reported that the protein level of decorin was not altered in the aqueous humor and LECs with aging when we compared the grade and type of lens opacity using human anterior lens capsule samples obtained during cataract surgery [7]. The expression of decorin is modulated by various cytokines and can reduce inflammation and fibrosis. Therefore, the altered expression levels of decorin in eye diseases suggest that it may play a role in modulating the ECM and cellular and microvascular integrity.
5. The Role of Decorin in the Lens
No drug has been discovered to treat or prevent cataracts; thus, the only current treatment for cataracts is surgery. In modern cataract surgery, a lens capsular bag is produced after the aspiration and removal of the lens contents, consisting of a portion of the anterior and the entire posterior and intraocular lens (IOL) inserted into the lens capsule. However, after several years post-operation, patients may develop opacity in the lens posterior capsule. Due to PCO, visual function is impaired; this is referred to as “after-cataracts” [86]. Months to years after cataract surgery, fibroblast-like differentiation, EMT, and the formation of a regenerating lens (called Elschnig pearls) may develop, leading to clouding and shrinkage of the lens capsule [86,108]. EMT results in an increased cell motility and ECM accumulation in LECs and has also been reported to be implicated in cell invasion, wound healing, and fibrosis [57,109]. After cataract surgery, transdifferentiation of LECs into fibroblast-like cells is induced in residual LECs, expressing high levels of fibronectin, procollagen, Tpm, and αSMA, resembling the wound healing process [68,69,86,108].
Aberrant TGF-β signaling plays a central role in the pathobiology of human ASF in the lens [57,110,111,112] and PCO [68,69,108,113,114]. As with TGF-β, the biological effects of FGF2 on the development of PCO may be either detrimental or beneficial since FGF2 stim-ulates LEC proliferation and mitosis and collagen synthesis but reduces the expression of αSMA [115,116,117]. FGF2 stimulates lens fiber differentiation in a dose-dependent manner [118,119] and activates LEC mitosis, increasing collagen formation [120]. The concentra-tion of FGF2 increases in the LECs after cataract surgery [121]. Aberrant TGF-β2 signaling also promotes LEC migration, proliferation, stress fiber formation, and EMT, inducing the appearance of αSMA-positive myofibroblasts, which is modulated by FGF2 [68].
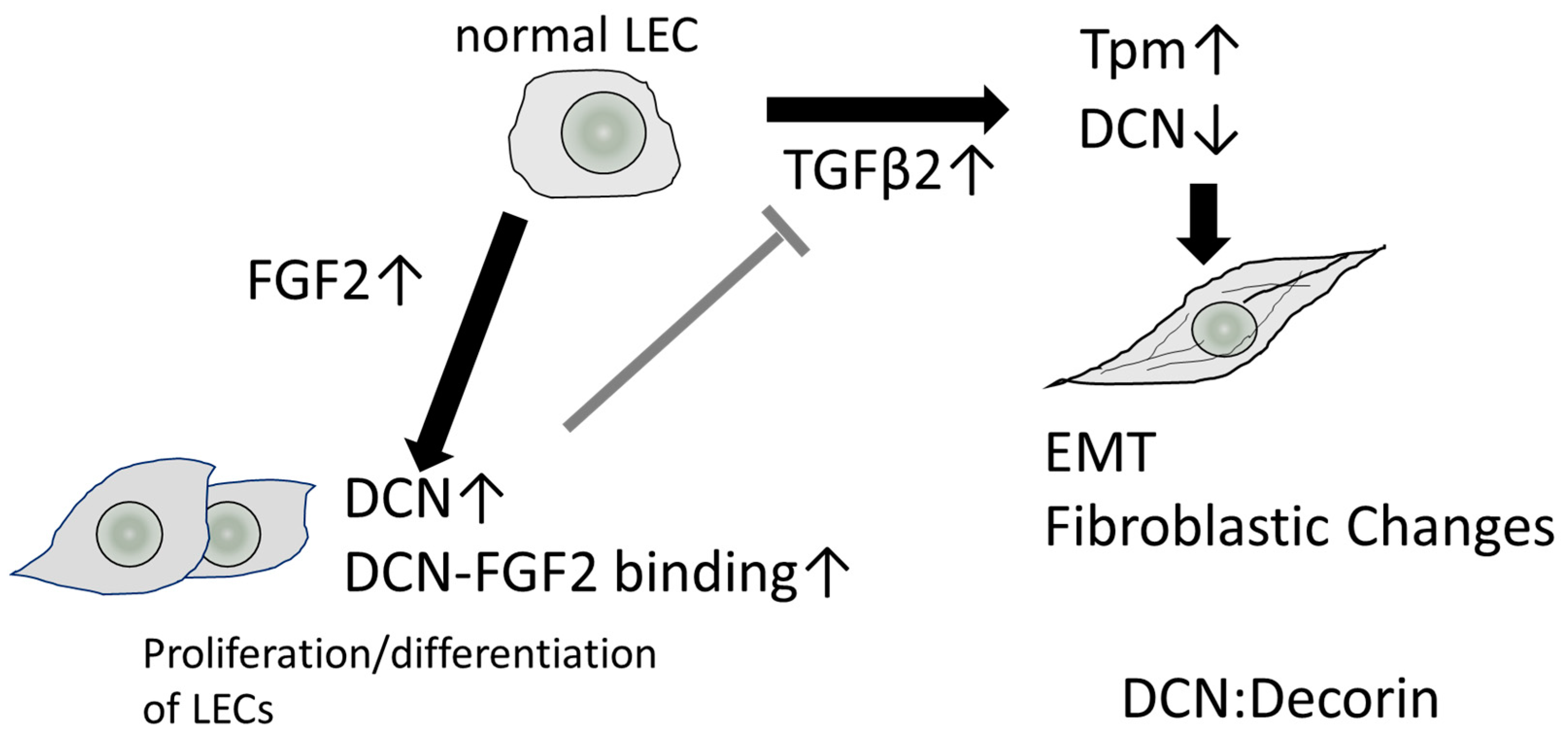
There are only a few reports on the role of decorin in the lens and its involvement in lens-related diseases. Under normal physiological conditions, the expression of decorin in LECs is low; however, the mRNA levels of decorin are increased in human LEC tissues (HLECs) with ASF obtained during cataract surgery [2]. Our recent study revealed that decorin may play a significant role in PCO and wound healing in the lens [7]. The expres-sion of many genes related to the ECM, including decorin, collagen, and fibronectin, were upregulated in LECs obtained from a mouse PCO model using a microarray-based ap-proach after extracapsular clear lens extraction (ECLE) [7]. Increased expression of decorin during EMT was confirmed in both rat and mouse PCO models via real-time quantitative PCR and Western blotting, suggesting that decorin is related to the cell proliferation, dif-ferentiation, fibrosis, and EMT of LECs observed in ASF and PCO [7]. Therefore, we ana-lyzed the interaction of decorin with TGF-β and FGF2 in the lens. In cultured mouse LECs (MLECs) and HLECs, the expression of decorin mRNA levels increased in response to FGF2 and decreased in response to TGFβ-2 [7]. Similar to our report, reduced expressions of decorin protein and mRNA after TGF-β treatment have been observed in many cell types, including fibroblasts [122,123]. It has been reported that the decorin gene contains TIE as a transcriptional silencer [76,77,78]. Thus, the addition of TGF-β may transcriptionally inhibit the expression of decorin in cultured LECs.
On the other hand, the mechanisms underlying the transcriptional regulation of decorin by FGF have not been elucidated, and the effect of FGF2 on the expression of decorin differed among various cell types [124,125,126,127]. Previous studies reported that decorin is secreted from skin fibroblasts and binds to FGF2 and FGF-7 in the human wound fluids collected within 24 h of surgery, activating cellular proliferation through the activation of the MAPK pathway [128,129]. Thus, the interaction of FGF2 and decorin may induce cell proliferation in PCO tissues. hDCN overexpression in HLECs suppressed TGF-β-induced Tpm1 upregulation [7]. Tpms are known as proteins that stabilize F-actin filaments, regulating the dynamic and structural properties of these filaments by controlling their interactions with actin binding proteins [130,131]. In our previous study, we observed that Tpm1 and Tpm2 are involved in regulating EMT and stabilizing actin stress fibers [68]. Both Tpm1 and Tpm2 are induced by TGFβ-2 during EMT in LECs and have been used as EMT markers in lens cells [68,132]. Thus, the overexpression of decorin in the lens may suppress TGFβ-mediated EMT. Moreover, FGF2 also acts as an antagonist of TGF-β-mediated EMT [68]. We reported that Tpm1 and Tpm2 expression was repressed after adding FGF2 to cultured MLECs and HLECs [133]. In a previous in vivo functional study, the extent of decorin binding to TGF-β inhibits hepatic fibrosis [20]. In other words, there is a complex relationship between decorin and TGF-β in that TGF-β can mutually repress decorin gene expression in fibroblasts and LECs. This suggests that a complex feedback loop exists between these two factors to maintain cellular homeostasis after surgery and during wound healing (Figure 3).
In a human PCO model using capsular bags from donor eyes that received an IOL, the levels of MMP2 and MMP9 were transiently elevated [134]. Additionally, pro-MMP2 and pro-MMP9 levels were increased and sustained upon treatment with active TGF-β2 [134]. Furthermore, the levels of pro-form MMP2 and MMP9 and active MMP2 were in-creased in the bathing media upon treatment with TGF-β in an organ-cultured lens from a rat ASF model [135]. In addition, the broad-spectrum MMP inhibitor GM6001 and a spe-cific MMP2/9 inhibitor suppressed the TGF-β induced-ASF [135]. From these studies, we speculate that decorin may be cleaved by MMPs in ASF and PCO and accelerate the acti-vation and release of TGF-β.
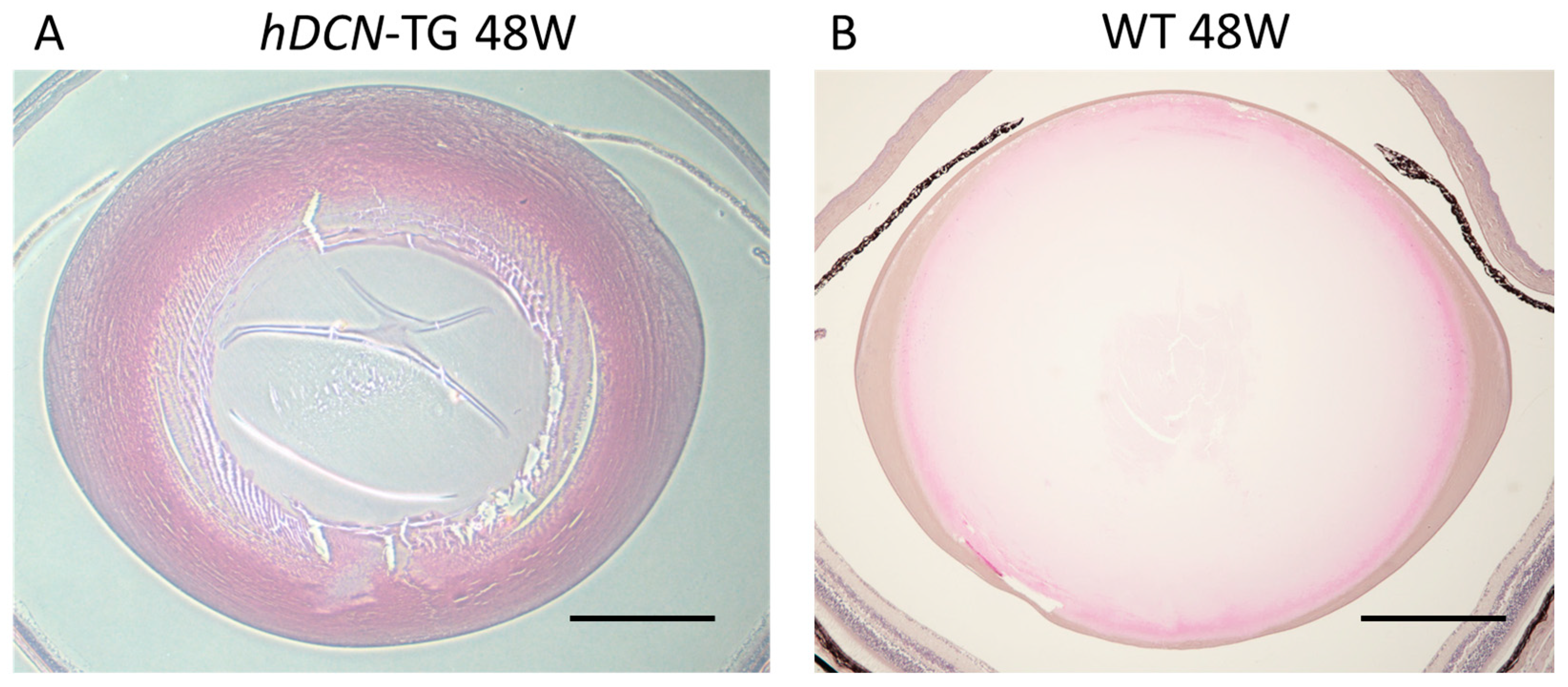
In the context of wound healing [6], it has been suggested that decorin interacts by localizing TGF-β, FGF2, and EGF close to the receptor. In the pathogenesis of PCO and other fibrotic eye diseases, decorin expression may be upregulated by FGF2, suppressing EMT and inducing normal cell proliferation and differentiation in LECs (Figure 3). We previously established a mouse PCO model with injury-induced EMT and fibrosis in the lens capsule to analyze the effect of decorin overexpression in wound healing in the lens using lens-specific human decorin conditional transgenic mice (hDCN-Tg) [7,133]. To examine the effect of decorin over-expression on the eye lens in vivo, we generated lens-specific hDCN-Tg using a plasmid in which Pax6-human αA-crystallin composite promoter (CPV14) [136] drives hDCN cDNA, as described in our previous study [7]. When the anterior capsule was injured via needle puncture, LECs differentiated into fibroblasts during the healing process via epithelial-myoblastic transition (EMyoT) [94,95] and transformed into αSMA-positive myofibroblasts. Wound healing was enhanced and fibrosis was suppressed after lens injury in hDCN-Tg mice compared with those in the wild-type (WT) [7]. It has been reported that cutaneous excisional wound healing in decorin-deficient mice (Dcn−/−) was delayed compared with that in WT (Dcn+/+) controls [6]. Thus, it is clear that the overexpression and addition of decorin may serve as an excellent therapeutic approach for improving wound healing and PCO. Moreover, the lenses of hDCN-Tg mice were clear and did not show any phenotypic abnormalities, such as abnormal arrangement or swelling of lens fibers and cell loss or abnormal growth of LECs through 48 weeks of age (Figure 4). Hence, decorin does not adversely affect the normal development and differentiation of the mouse lens.
In another study on the lens, decorin was found to inhibit high glucose-induced ROS production and apoptosis by reducing the induction of p22phox and phospho-p38 in a cultured human LEC cell line [137]. It was also reported that decorin could increase the levels of endogenous antioxidants, such as glutathione peroxidase and superoxide dis-mutase (SOD), and decrease the levels of tissue malondialdehyde, nitric oxide, and caspase-3 in the rat cerebrum after post-traumatic brain injuries [138]. Furthermore, decorin could protect retinal pigment epithelial (RPE) cells from H2O2-induced oxidative stress and apoptosis by promoting autophagy [11]. Furthermore, ROS induces the secretion and activation of TGF-β1 and is essential for TGF-β-mediated myofibroblast differentiation under different pathological conditions in various cells [56,139,140,141]. TGF-β also increases intracellular ROS content in different cell types, including LECs and trabecular meshwork (TM) cells by downregulating the expression of antioxidant-related genes, including SOD, catalase, glutaredoxin, and lens epithelium-derived growth factor (LEDGF) [139,142,143,144]. This redox imbalance is an important contributor to the pathophysiological effects of TGF-β, including fibrosis [145]. Thus, decorin may work as an antioxidant and anti-TGFβ1 agent and maintain redox balance in lens tissues.
6. The Role of Decorin in Other Eye Diseases
6.1. Cornea
There are many reports on the role and function of decorin in the cornea. Decorin was first identified in the chicken cornea [146]. In humans, mutations in the decorin gene could cause congenital stromal corneal dystrophy (CSCD) when truncated decorin binds to collagen, resulting in aggregation in the stroma [100,147]. CSCD is the only human disease associated with mutations in decorin. CSCD is a rare autosomal dominant disorder characterized by small opacities found throughout the stroma [100,148]. Decorin is believed to inhibit corneal neovascularization under normal physiological conditions and is expected to maintain avascularity in the cornea [29]. The overexpression of decorin in cultured human corneal fibroblasts inhibited the TGF-β-induced expression of fibrogenic ECM genes such as fibronectin; alpha collagen type-I, -III, and -IV; and αSMA, a known EMT marker. In addition, the overexpression of decorin via tissue-targeted adeno-associated virus (AAV) 5-mediated delivery of the Decorin (Dcn) gene reduced corneal fibrosis caused by photorefractive keratectomy and corneal neovascularization in rabbits in vivo [149,150]. In addition, AAV5–Dcn gene delivery in the rabbit cornea revealed that it is safe and non-toxic to the eye in vivo during the 6-month follow-up period [151]. Thus, decorin plays a significant role in maintaining corneal transparency.
Decorin has also been reported to have therapeutic applications for various diseases of the cornea. For example, sustained topical delivery of a fluid gel eye drop containing human decorin to the cornea promoted wound healing and reduced corneal scarring and opacity in a murine model of pseudomonas-induced keratitis [152]. Furthermore, topical decorin treatment also promoted corneal epithelial nerve regeneration after corneal abra-sion injury using Cx3cr1-deficient mice spontaneously lacking resident corneal epithelial dendritic cells [153]. From these results, it was hypothesized that decorin induces corneal nerve regeneration through the activation of dendritic cells [153]. Thus, decorin gene therapy or the optical administration of decorin has a high potential for treating corneal fibrotic and angiogenic diseases.
6.2. Glaucoma
Intraocular pressure (IOP) is the cardinal modifiable important risk factor for the on-set and progression of glaucoma [154,155]. TM and Schlemm’s canal provide the major route for the outflow of the aqueous humor from the anterior segment of the eye and are responsible for the raised IOP associated with POAG due to increased outflow resistance [156]. Here, the TM exhibits marked contractile properties, and TM contraction is associ-ated with reduced aqueous humor drainage through the outflow system [157]. Decorin has extenisve anti-fibrotic effects in glaucoma. The aqueous level of decorin was found to be decreased in eyes with glaucoma compared with non-glaucomatous eyes [88]. Moreo-ver, it has been reported that soluble decorin attenuated the generation and expression of typical downstream target genes, such as CCN2/CTGF, Fn, and Col IV, in human TM cells by TGF-β2. The negative reciprocal regulation of decorin and TGF-β was also observed, confirming the dramatic downregulation of decorin in the canonical outflow pathway of patients with POAG [14]. Expression of decorin mRNA was also decreased in the endo-thelial cells of the Schlemm canal derived from glaucomatous patients [158]. Intracameral injection of human recombinant decorin considerably reduced fibrosis in the TM and IOP and inhibited retinal ganglion cell loss in a rodent model of TM fibrosis via repeated in-tracameral injection of TGF-β [8]. Thus, decorin may be useful in developing an effective therapy for eyes with ocular hypertension and POAG associated with the fibrosis of the TM.
In POAG, the disturbed ECM metabolism and ECM remodeling are observed in the human optic nerve (ON) head, which is particularly obvious in the lamina cribrosa (LC) layer [159]. The remodeling processes in the ON head of glaucomatous patients are ac-companied by a disruption of the homeostatic balance of growth factors such as TGF-β [160]. In oligonucleotide microarray analysis using cultured LC cells from POAG human donors, expression of decorin was decreased [159]. In decorin-deficient mice, it was re-ported that an increase in IOP and a loss of ON axons [14] as well as a dramatic upreg-ulation of CTGF/CCN2, TGF-β-1, and TGF-β-2 were found [161]. In ON astrocytes, decorin negativery regulated TGF-β and CTGF/CCN2 [161]. Thus, the downregulation of decorin in the glaucomatous Schlemm canal endothelial cells, LC cells, and optic nerve head may affect the progression of POAG. The treatment of decorin may attenuate the remodeling process in the ON head observed during the progression of glaucoma and may reduce IOP.
Furthermore, in patients with uncontrolled glaucoma with progressive visual field loss due to glaucoma medication, IOP reduction by glaucoma surgery may be necessary. Glaucoma filtration surgery is a useful technique in reducing the IOP. Although this tech-nique immediately reduces IOP, the long-term effect is often eclipsed by the postoperative wound-healing process [162,163,164]. After glaucoma filtration treatments, the subconjuncti-val injection of decorin in rabbits showed markedly reduced conjunctival scarring and maintained lower IOP levels [165]. Thus, the application of decorin can inhibit the ob-struction of the fistula, which in turn inhibits fibroproliferative processes in fibroblasts from Tenon’s capsule and the episclera after filtration surgery [165].
The protein expression of decorin was also found to be increased in LECs of patients with PXG compared with those with pseudoexfoliation syndrome [106]. Simultaneously, TGFβ1 and FGF2 expression is elevated in the LECs of patients with PXG [106,166]. Thus, through the protein interaction networks of TGF-β and FGF2 that induce ECM remodeling, decorin may play a potential role in PXG and may be used as a novel therapeutic target for glaucoma.
6.3. Retinal Diseases
SLRPs are crucial in many important biological functions involved in the mainte-nance of retinal homeostasis [13]. Decorin localizes in all layers of the human retina, in-cluding the RPE, Bruch’s membrane, choroidal stroma, and choroidal blood vessel walls [12]. A key step in PDR is neo-vascularization, beginning with the activation of the angio-genic switch after ischemic changes. Wet AMD is characterized by the so-called angio-genesis choroidal neovascularization or retinal aingiomatous proliferation [167]. Decorin is a key regulator for angiogenic responses by acting as an inhibitor for multiple RTKs and suppressing VEGFR through various mechanisms in choroid-retinal endothelial cells [26,168]. Moreover, many reports have demonstrated the anti-angiogenic effects of decorin in the retina [168,169]. A decreased expression of decorin has been reported in the retina of animal models of ischemic disease in the retinal vessels, such as oxygen-induced retinopathy in rats [170,171]. In another study, decorin treatment in rats with oxygen-induced retinopathy inhibited retinal neovascularization by reducing neovascular endothelial cell proliferation and the immunoreactivity of VEGF and TNF-α in the retina [172]. Additionally, intravitreal injection of decorin extensively suppressed VEGF and TNF-α expression and inhibited TGF-β expression in the RPE-choroid complex in a mouse model of laser-induced choroidal neovascularization, a model of AMD [169]. Thus, the administration of decorin into the eyes may inhibit VEGF-induced retinal diseases related to angiogenesis. Moreover, PVR is a complex process involving cell proliferation and ECM remodeling [173]. TGF-β is over-expressed in the vitreous humor of patients with PVR and PDR [55]. TGF-β is also presumed to be involved in the contraction of proliferative membranes and subretinal strands in PVR [174]. In an experimental model of rabbit traumatic PVR, intravitreal injection of decorin during vitrectomy reduced the development of fibrosis and tractional retinal detachment [175]. Therefore, decorin is also expected to prevent or regress neovascularization, EMT, and fibrosis in ischemic retinal diseases such as retinopathy of prematurity, proliferative DR, PVR, retinal detachment, and AMD as a TGF-β antagonist [9,11,107,168,172,175].
7. Summary
In summary, decorin can neutralize several important mediators of fibrosis and an-giogenesis in the eyes. Decorin has been shown to have anti-fibrotic effects in various an-imal models of eye diseases, and no toxicity has been reported in these studies. Decorin regulates the activity of multiple growth factors, receptor tyrosine kinases, and extracellu-lar matrix components. Although preclinical studies have been conducted, decorin has not yet been clinically applied as a drug due to its short half-life in circulation and diffi-culty in mass production since high doses are required for effective treatment. In the future, it is expected that GAG-unbound decorin core proteins, which are easier to create, will be produced in large quantities for use in pharmaceuticals [27,176]. Therefore, decorin is expected to be clinically used in the treatment of various ocular diseases needing adjuvant peptide therapies, including PCO, due to its anti-fibrotic, anti-inflammatory, anti-angiogenic, and antioxidant effects by affecting several growth factor pathways in the eyes. Hence, the development of effective intraocular delivery methods for decorin is warranted.
Author Contributions
E.K. wrote the manuscript. H.S. and S.S. guided the planning. D.P.S. wrote the review manuscript. T.S. and D.P.S. critically amended the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant provided by the JSPS Grants-in-Aid for Scientific Research C 23592588 (to S.S.), the National Eye Institute, National Institute of Health [grant number EY024589, (to D.P.S.); and Bridge Grant, UNMC (to D.P.S.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mohan, R.R.; Tovey, J.C.; Gupta, R.; Sharma, A.; Tandon, A. Decorin biology, expression, function and therapy in the cornea. Curr. Mol. Med. 2011, 11, 110–128. [Google Scholar] [CrossRef] [PubMed]
- Azuma, N.; Hara, T.; Hara, T. Extracellular matrix of opacified anterior capsule after endocapsular cataract surgery. Graefes Arch. Clin. Exp. Ophthalmol. 1998, 236, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Villodre, E.S.; Larson, R.; Rahal, O.M.; Wang, X.; Gong, Y.; Song, J.; Krishnamurthy, S.; Ueno, N.T.; Tripathy, D.; et al. Decorin-mediated suppression of tumorigenesis, invasion, and metastasis in inflammatory breast cancer. Commun. Biol. 2021, 4, 72. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V. Matrix proteoglycans: From molecular design to cellular function. Annu. Rev. Biochem. 1998, 67, 609–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iozzo, R.V.; Schaefer, L. Proteoglycans in health and disease: Novel regulatory signaling mechanisms evoked by the small leucine-rich proteoglycans. FEBS J. 2010, 277, 3864–3875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvelainen, H.; Puolakkainen, P.; Pakkanen, S.; Brown, E.L.; Hook, M.; Iozzo, R.V.; Sage, E.H.; Wight, T.N. A role for decorin in cutaneous wound healing and angiogenesis. Wound Repair Regen. 2006, 14, 443–452. [Google Scholar] [CrossRef]
- Shibata, S.; Shibata, N.; Ohtsuka, S.; Yoshitomi, Y.; Kiyokawa, E.; Yonekura, H.; Singh, D.P.; Sasaki, H.; Kubo, E. Role of decorin in posterior capsule opacification and eye lens development. Cells 2021, 10, 863. [Google Scholar] [CrossRef]
- Hill, L.J.; Mead, B.; Blanch, R.J.; Ahmed, Z.; De Cogan, F.; Morgan-Warren, P.J.; Mohamed, S.; Leadbeater, W.; Scott, R.A.; Berry, M.; et al. Decorin reduces intraocular pressure and retinal ganglion cell loss in rodents through fibrolysis of the scarred trabecular meshwork. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3743–3757. [Google Scholar] [CrossRef]
- Low, S.W.Y.; Vaidya, T.; Gadde, S.G.K.; Mochi, T.B.; Kumar, D.; Kassem, I.S.; Costakos, D.M.; Ahmad, B.; Sethu, S.; Ghosh, A.; et al. Decorin concentrations in aqueous humor of patients with diabetic retinopathy. Life 2021, 11, 1421. [Google Scholar] [CrossRef]
- Oruc, Y.; Keser, S.; Yusufoglu, E.; Celik, F.; Sahin, I.; Yardim, M.; Aydin, S. Decorin, tenascin C, total antioxidant, and total oxidant level changes in patients with pseudoexfoliation syndrome. J. Ophthalmol. 2018, 2018, 7459496. [Google Scholar] [CrossRef]
- Xie, X.; Li, D.; Cui, Y.; Xie, T.; Cai, J.; Yao, Y. Decorin protects retinal pigment epithelium cells from oxidative stress and apoptosis via AMPK-mTOR-regulated autophagy. Oxid. Med. Cell. Longev. 2022, 2022, 3955748. [Google Scholar] [CrossRef] [PubMed]
- Keenan, T.D.; Clark, S.J.; Unwin, R.D.; Ridge, L.A.; Day, A.J.; Bishop, P.N. Mapping the differential distribution of proteoglycan core proteins in the adult human retina, choroid, and sclera. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7528–7538. [Google Scholar] [CrossRef] [PubMed]
- Low, S.W.Y.; Connor, T.B.; Kassem, I.S.; Costakos, D.M.; Chaurasia, S.S. Small leucine-rich proteoglycans (SLRPs) in the retina. Int. J. Mol. Sci. 2021, 22, 7293. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Pawlak, R.; Weber, G.R.; Dillinger, A.E.; Kuespert, S.; Iozzo, R.V.; Quigley, H.A.; Ohlmann, A.; Tamm, E.R.; Fuchshofer, R. A novel ocular function for decorin in the aqueous humor outflow. Matrix Biol. 2021, 97, 1–19. [Google Scholar] [CrossRef]
- Vogel, K.G.; Paulsson, M.; Heinegard, D. Specific inhibition of type I and type II collagen fibrillogenesis by the small proteoglycan of tendon. Biochem. J. 1984, 223, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Nareyeck, G.; Seidler, D.G.; Troyer, D.; Rauterberg, J.; Kresse, H.; Schonherr, E. Differential interactions of decorin and decorin mutants with type I and type VI collagens. Eur. J. Biochem. 2004, 271, 3389–3398. [Google Scholar] [CrossRef]
- Ruhland, C.; Schonherr, E.; Robenek, H.; Hansen, U.; Iozzo, R.V.; Bruckner, P.; Seidler, D.G. The glycosaminoglycan chain of decorin plays an important role in collagen fibril formation at the early stages of fibrillogenesis. FEBS J. 2007, 274, 4246–4255. [Google Scholar] [CrossRef]
- Neill, T.; Schaefer, L.; Iozzo, R.V. Decorin: A guardian from the Matrix. Am. J. Pathol. 2012, 181, 380–387. [Google Scholar] [CrossRef] [Green Version]
- Merline, R.; Moreth, K.; Beckmann, J.; Nastase, M.V.; Zeng-Brouwers, J.; Tralhao, J.G.; Lemarchand, P.; Pfeilschifter, J.; Schaefer, R.M.; Iozzo, R.V.; et al. Signaling by the matrix proteoglycan decorin controls inflammation and cancer through PDCD4 and MicroRNA-21. Sci. Signal. 2011, 4, ra75. [Google Scholar] [CrossRef] [Green Version]
- Baghy, K.; Iozzo, R.V.; Kovalszky, I. Decorin-TGFbeta axis in hepatic fibrosis and cirrhosis. J. Histochem. Cytochem. 2012, 60, 262–268. [Google Scholar] [CrossRef]
- Bi, X.; Pohl, N.M.; Qian, Z.; Yang, G.R.; Gou, Y.; Guzman, G.; Kajdacsy-Balla, A.; Iozzo, R.V.; Yang, W. Decorin-mediated inhibition of colorectal cancer growth and migration is associated with E-cadherin in vitro and in mice. Carcinogenesis 2012, 33, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V.; Sanderson, R.D. Proteoglycans in cancer biology, tumour microenvironment and angiogenesis. J. Cell. Mol. Med. 2011, 15, 1013–1031. [Google Scholar] [CrossRef] [PubMed]
- Gubbiotti, M.A.; Neill, T.; Frey, H.; Schaefer, L.; Iozzo, R.V. Decorin is an autophagy-inducible proteoglycan and is required for proper in vivo autophagy. Matrix Biol. 2015, 48, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Young, M.F.; Chakravarti, S.; Birk, D.E. Interclass small leucine-rich repeat proteoglycan interactions regulate collagen fibrillogenesis and corneal stromal assembly. Matrix Biol. 2014, 35, 103–111. [Google Scholar] [CrossRef]
- Santra, M.; Eichstetter, I.; Iozzo, R.V. An anti-oncogenic role for decorin. Down-regulation of ErbB2 leads to growth suppression and cytodifferentiation of mammary carcinoma cells. J. Biol. Chem. 2000, 275, 35153–35161. [Google Scholar] [CrossRef] [Green Version]
- Csordas, G.; Santra, M.; Reed, C.C.; Eichstetter, I.; McQuillan, D.J.; Gross, D.; Nugent, M.A.; Hajnoczky, G.; Iozzo, R.V. Sustained down-regulation of the epidermal growth factor receptor by decorin. A mechanism for controlling tumor growth in vivo. J. Biol. Chem. 2000, 275, 32879–32887. [Google Scholar] [CrossRef] [Green Version]
- Jarvinen, T.A.; Prince, S. Decorin: A growth factor antagonist for tumor growth inhibition. BioMed Res. Int. 2015, 2015, 654765. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, L.; Schaefer, R.M. Proteoglycans: From structural compounds to signaling molecules. Cell Tissue Res. 2010, 339, 237–246. [Google Scholar] [CrossRef]
- Gubbiotti, M.A.; Vallet, S.D.; Ricard-Blum, S.; Iozzo, R.V. Decorin interacting network: A comprehensive analysis of decorin-binding partners and their versatile functions. Matrix Biol. 2016, 55, 7–21. [Google Scholar] [CrossRef]
- Stander, M.; Naumann, U.; Wick, W.; Weller, M. Transforming growth factor-beta and p-21: Multiple molecular targets of decorin-mediated suppression of neoplastic growth. Cell Tissue Res. 1999, 296, 221–227. [Google Scholar] [CrossRef]
- Chen, S.; Birk, D.E. Focus on molecules: Decorin. Exp. Eye Res. 2011, 92, 444–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, M.W.; Chamberlain, C.G.; McAvoy, J.W. Binding of FGF-1 and FGF-2 to heparan sulphate proteoglycans of the mammalian lens capsule. Growth Factors 1997, 14, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Seidler, D.G.; Dreier, R. Decorin and its galactosaminoglycan chain: Extracellular regulator of cellular function? IUBMB Life 2008, 60, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Hocking, A.M.; Shinomura, T.; McQuillan, D.J. Leucine-rich repeat glycoproteins of the extracellular Matrix. Matrix Biol. 1998, 17, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Danielson, K.G.; Baribault, H.; Holmes, D.F.; Graham, H.; Kadler, K.E.; Iozzo, R.V. Targeted disruption of decorin leads to abnormal collagen fibril morphology and skin fragility. J. Cell Biol. 1997, 136, 729–743. [Google Scholar] [CrossRef] [Green Version]
- Dunkman, A.A.; Buckley, M.R.; Mienaltowski, M.J.; Adams, S.M.; Thomas, S.J.; Satchell, L.; Kumar, A.; Pathmanathan, L.; Beason, D.P.; Iozzo, R.V.; et al. Decorin expression is important for age-related changes in tendon structure and mechanical properties. Matrix Biol. 2013, 32, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, Y.; Kodama, Y.; Matsumoto, T. Bone matrix decorin binds transforming growth factor-beta and enhances its bioactivity. J. Biol. Chem. 1994, 269, 32634–32638. [Google Scholar] [CrossRef]
- Brandan, E.; Retamal, C.; Cabello-Verrugio, C.; Marzolo, M.P. The low density lipoprotein receptor-related protein functions as an endocytic receptor for decorin. J. Biol. Chem. 2006, 281, 31562–31571. [Google Scholar] [CrossRef]
- Cabello-Verrugio, C.; Brandan, E. A novel modulatory mechanism of transforming growth factor-beta signaling through decorin and LRP-1. J. Biol. Chem. 2007, 282, 18842–18850. [Google Scholar] [CrossRef] [Green Version]
- Cabello-Verrugio, C.; Santander, C.; Cofre, C.; Acuna, M.J.; Melo, F.; Brandan, E. The internal region leucine-rich repeat 6 of decorin interacts with low density lipoprotein receptor-related protein-1, modulates transforming growth factor (TGF)-beta-dependent signaling, and inhibits TGF-beta-dependent fibrotic response in skeletal muscles. J. Biol. Chem. 2012, 287, 6773–6787. [Google Scholar] [CrossRef]
- Ungvari, Z.; Valcarcel-Ares, M.N.; Tarantini, S.; Yabluchanskiy, A.; Fulop, G.A.; Kiss, T.; Csiszar, A. Connective tissue growth factor (CTGF) in age-related vascular pathologies. Geroscience 2017, 39, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Lipson, K.E.; Wong, C.; Teng, Y.; Spong, S. CTGF is a central mediator of tissue remodeling and fibrosis and its inhibition can reverse the process of fibrosis. Fibrogenesis Tissue Repair 2012, 5, S24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vial, C.; Gutierrez, J.; Santander, C.; Cabrera, D.; Brandan, E. Decorin interacts with connective tissue growth factor (CTGF)/CCN2 by LRR12 inhibiting its biological activity. J. Biol. Chem. 2011, 286, 24242–24252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, L.; Iozzo, R.V. Biological functions of the small leucine-rich proteoglycans: From genetics to signal transduction. J. Biol. Chem. 2008, 283, 21305–21309. [Google Scholar] [CrossRef] [Green Version]
- Khan, G.A.; Girish, G.V.; Lala, N.; Di Guglielmo, G.M.; Lala, P.K. Decorin is a novel VEGFR-2-binding antagonist for the human extravillous trophoblast. Mol. Endocrinol. 2011, 25, 1431–1443. [Google Scholar] [CrossRef]
- Rohani, M.G.; Parks, W.C. Matrix remodeling by MMPs during wound repair. Matrix Biol. 2015, 44–46, 113–121. [Google Scholar] [CrossRef]
- Itoh, Y. Membrane-type matrix metalloproteinases: Their functions and regulations. Matrix Biol. 2015, 44–46, 207–223. [Google Scholar] [CrossRef]
- Deryugina, E.I.; Quigley, J.P. Tumor angiogenesis: MMP-mediated induction of intravasation- and metastasis-sustaining neovasculature. Matrix Biol. 2015, 44–46, 94–112. [Google Scholar] [CrossRef]
- Imai, K.; Hiramatsu, A.; Fukushima, D.; Pierschbacher, M.D.; Okada, Y. Degradation of decorin by matrix metalloproteinases: Identification of the cleavage sites, kinetic analyses and transforming growth factor-beta1 release. Biochem. J. 1997, 322 Pt 3, 809–814. [Google Scholar] [CrossRef]
- Yu, Q.; Stamenkovic, I. Cell surface-localized matrix metalloproteinase-9 proteolytically activates TGF-beta and promotes tumor invasion and angiogenesis. Genes Dev. 2000, 14, 163–176. [Google Scholar] [CrossRef]
- Dallas, S.L.; Rosser, J.L.; Mundy, G.R.; Bonewald, L.F. Proteolysis of latent transforming growth factor-beta (TGF-beta)-binding protein-1 by osteoclasts. A cellular mechanism for release of TGF-beta from bone Matrix. J. Biol. Chem 2002, 277, 21352–21360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tandon, A.; Tovey, J.C.; Sharma, A.; Gupta, R.; Mohan, R.R. Role of transforming growth factor Beta in corneal function, biology and pathology. Curr. Mol. Med. 2010, 10, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Li, M.O.; Wan, Y.Y.; Sanjabi, S.; Robertson, A.K.; Flavell, R.A. Transforming growth factor-beta regulation of immune responses. Annu. Rev. Immunol. 2006, 24, 99–146. [Google Scholar] [CrossRef] [PubMed]
- Hales, A.M.; Schulz, M.W.; Chamberlain, C.G.; McAvoy, J.W. TGF-beta 1 induces lens cells to accumulate alpha-smooth muscle actin, a marker for subcapsular cataracts. Curr. Eye Res. 1994, 13, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Kita, T.; Hata, Y.; Kano, K.; Miura, M.; Nakao, S.; Noda, Y.; Shimokawa, H.; Ishibashi, T. Transforming growth factor-beta2 and connective tissue growth factor in proliferative vitreoretinal diseases: Possible involvement of hyalocytes and therapeutic potential of Rho kinase inhibitor. Diabetes 2007, 56, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.M.; Desai, L.P. Reciprocal regulation of TGF-beta and reactive oxygen species: A perverse cycle for fibrosis. Redox Biol. 2015, 6, 565–577. [Google Scholar] [CrossRef] [Green Version]
- Lovicu, F.J.; Schulz, M.W.; Hales, A.M.; Vincent, L.N.; Overbeek, P.A.; Chamberlain, C.G.; McAvoy, J.W. TGFbeta induces morphological and molecular changes similar to human anterior subcapsular cataract. Br. J. Ophthalmol. 2002, 86, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Broekelmann, T.J.; Limper, A.H.; Colby, T.V.; McDonald, J.A. Transforming growth factor beta 1 is present at sites of extracellular matrix gene expression in human pulmonary fibrosis. Proc. Natl. Acad. Sci. USA 1991, 88, 6642–6646. [Google Scholar] [CrossRef] [Green Version]
- Massague, J. The transforming growth factor-beta family. Annu. Rev. Cell Biol. 1990, 6, 597–641. [Google Scholar] [CrossRef]
- Ask, K.; Bonniaud, P.; Maass, K.; Eickelberg, O.; Margetts, P.J.; Warburton, D.; Groffen, J.; Gauldie, J.; Kolb, M. Progressive pulmonary fibrosis is mediated by TGF-beta isoform 1 but not TGF-beta3. Int. J. Biochem. Cell Biol. 2008, 40, 484–495. [Google Scholar] [CrossRef]
- Zhou, L.; Zhu, X.; Tan, J.; Wang, J.; Xing, Y. Effect of recombinant adeno-associated virus mediated transforming growth factor-beta1 on corneal allograft survival after high-risk penetrating keratoplasty. Transpl. Immunol. 2013, 28, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.H.; Chen, D.Q.; Wang, Y.N.; Feng, Y.L.; Cao, G.; Vaziri, N.D.; Zhao, Y.Y. New insights into TGF-beta/Smad signaling in tissue fibrosis. Chem. Biol. Interact. 2018, 292, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massague, J.; Seoane, J.; Wotton, D. Smad transcription factors. Genes Dev. 2005, 19, 2783–2810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmierer, B.; Hill, C.S. TGFbeta-SMAD signal transduction: Molecular specificity and functional flexibility. Nat. Rev. Mol. Cell Biol. 2007, 8, 970–982. [Google Scholar] [CrossRef]
- Yu, L.; Hebert, M.C.; Zhang, Y.E. TGF-beta receptor-activated p38 MAP kinase mediates Smad-independent TGF-beta responses. EMBO J. 2002, 21, 3749–3759. [Google Scholar] [CrossRef] [Green Version]
- Stratton, R.; Rajkumar, V.; Ponticos, M.; Nichols, B.; Shiwen, X.; Black, C.M.; Abraham, D.J.; Leask, A. Prostacyclin derivatives prevent the fibrotic response to TGF-beta by inhibiting the Ras/MEK/ERK pathway. FASEB J. 2002, 16, 1949–1951. [Google Scholar] [CrossRef]
- Biernacka, A.; Dobaczewski, M.; Frangogiannis, N.G. TGF-beta signaling in fibrosis. Growth Factors 2011, 29, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Kubo, E.; Shibata, S.; Shibata, T.; Kiyokawa, E.; Sasaki, H.; Singh, D.P. FGF2 antagonizes aberrant TGFbeta regulation of tropomyosin: Role for posterior capsule opacity. J. Cell. Mol. Med. 2017, 21, 916–928. [Google Scholar] [CrossRef]
- Kubo, E.; Shibata, T.; Singh, D.P.; Sasaki, H. Roles of TGF beta and FGF signals in the lens: Tropomyosin regulation for posterior capsule cpacity. Int. J. Mol. Sci. 2018, 19, 3093. [Google Scholar] [CrossRef] [Green Version]
- Kurosaka, D.; Kato, K.; Nagamoto, T.; Negishi, K. Growth factors influence contractility and alpha-smooth muscle actin expression in bovine lens epithelial cells. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1701–1708. [Google Scholar]
- Yamaguchi, Y.; Mann, D.M.; Ruoslahti, E. Negative regulation of transforming growth factor-beta by the proteoglycan decorin. Nature 1990, 346, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Ruoslahti, E.; Yamaguchi, Y. Proteoglycans as modulators of growth factor activities. Cell 1991, 64, 867–869. [Google Scholar] [CrossRef] [PubMed]
- Kerr, L.D.; Miller, D.B.; Matrisian, L.M. TGF-beta 1 inhibition of transin/stromelysin gene expression is mediated through a Fos binding sequence. Cell 1990, 61, 267–278. [Google Scholar] [CrossRef] [PubMed]
- White, L.A.; Mitchell, T.I.; Brinckerhoff, C.E. Transforming growth factor beta inhibitory element in the rabbit matrix metalloproteinase-1 (collagenase-1) gene functions as a repressor of constitutive transcription. Biochim. Biophys. Acta 2000, 1490, 259–268. [Google Scholar] [CrossRef]
- Sharma, P.; Fatma, N.; Kubo, E.; Shinohara, T.; Chylack, L.T., Jr.; Singh, D.P. Lens epithelium-derived growth factor relieves transforming growth factor-beta1-induced transcription repression of heat shock proteins in human lens epithelial cells. J. Biol. Chem. 2003, 278, 20037–20046. [Google Scholar] [CrossRef] [Green Version]
- Danielson, K.G.; Siracusa, L.D.; Donovan, P.J.; Iozzo, R.V. Decorin, epiphycan, and lumican genes are closely linked on murine Chromosome 10 and are deleted in lethal steel mutants. Mamm. Genome 1999, 10, 201–203. [Google Scholar] [CrossRef] [PubMed]
- Demoor-Fossard, M.; Galera, P.; Santra, M.; Iozzo, R.V.; Pujol, J.P.; Redini, F. A composite element binding the vitamin D receptor and the retinoic X receptor alpha mediates the transforming growth factor-beta inhibition of decorin gene expression in articular chondrocytes. J. Biol. Chem. 2001, 276, 36983–36992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iozzo, R.V.; Cohen, I. Altered proteoglycan gene expression and the tumor stroma. Experientia 1993, 49, 447–455. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Moscatello, D.K.; McQuillan, D.J.; Eichstetter, I. Decorin is a biological ligand for the epidermal growth factor receptor. J. Biol. Chem. 1999, 274, 4489–4492. [Google Scholar] [CrossRef] [Green Version]
- Mauviel, A.; Santra, M.; Chen, Y.Q.; Uitto, J.; Iozzo, R.V. Transcriptional regulation of decorin gene expression. Induction by quiescence and repression by tumor necrosis factor-alpha. J. Biol. Chem. 1995, 270, 11692–11700. [Google Scholar] [CrossRef] [Green Version]
- Mauviel, A.; Korang, K.; Santra, M.; Tewari, D.; Uitto, J.; Iozzo, R.V. Identification of a bimodal regulatory element encompassing a canonical AP-1 binding site in the proximal promoter region of the human decorin gene. J. Biol. Chem. 1996, 271, 24824–24829. [Google Scholar] [CrossRef] [PubMed]
- Demoor-Fossard, M.; Redini, F.; Boittin, M.; Pujol, J.P. Expression of decorin and biglycan by rabbit articular chondrocytes. Effects of cytokines and phenotypic modulation. Biochim. Biophys. Acta 1998, 1398, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Border, W.A.; Okuda, S.; Languino, L.R.; Ruoslahti, E. Transforming growth factor-beta regulates production of proteoglycans by mesangial cells. Kidney. Int. 1990, 37, 689–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, D.Y.; Lovicu, F.J. Myofibroblast transdifferentiation: The dark force in ocular wound healing and fibrosis. Prog. Retin. Eye Res. 2017, 60, 44–65. [Google Scholar] [CrossRef] [PubMed]
- Lois, N.; Taylor, J.; McKinnon, A.D.; Smith, G.C.; van’t Hof, R.; Forrester, J.V. Effect of TGF-beta2 and anti-TGF-beta2 antibody in a new in vivo rodent model of posterior capsule opacification. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4260–4266. [Google Scholar] [CrossRef] [Green Version]
- McDonnell, P.J.; Rowen, S.L.; Glaser, B.M.; Sato, M. Posterior capsule opacification. An in vitro model. Arch. Ophthalmol. 1985, 103, 1378–1381. [Google Scholar] [CrossRef]
- Nikhalashree, S.; George, R.; Shantha, B.; Lingam, V.; Vidya, W.; Panday, M.; Sulochana, K.N.; Coral, K. Detection of proteins associated with extracellular matrix regulation in the aqueous humour of patients with primary glaucoma. Curr. Eye Res. 2019, 44, 1018–1025. [Google Scholar] [CrossRef]
- NikhalaShree, S.; Karthikkeyan, G.; George, R.; Shantha, B.; Vijaya, L.; Ratra, V.; Sulochana, K.N.; Coral, K. Lowered decorin with aberrant extracellular matrix remodeling in aqueous humor and tenon’s tissue from primary glaucoma patients. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4661–4669. [Google Scholar] [CrossRef] [Green Version]
- Guerin, C.J.; Hu, L.; Scicli, G.; Scicli, A.G. Transforming growth factor beta in experimentally detached retina and periretinal membranes. Exp. Eye Res. 2001, 73, 753–764. [Google Scholar] [CrossRef]
- Jampel, H.D.; Roche, N.; Stark, W.J.; Roberts, A.B. Transforming growth factor-beta in human aqueous humor. Curr. Eye Res. 1990, 9, 963–969. [Google Scholar] [CrossRef]
- Wallentin, N.; Wickstrom, K.; Lundberg, C. Effect of cataract surgery on aqueous TGF-beta and lens epithelial cell proliferation. Investig. Ophthalmol. Vis. Sci. 1998, 39, 1410–1418. [Google Scholar]
- Hu, D.N.; McCormick, S.A.; Lin, A.Y.; Lin, J.Y. TGF-beta2 inhibits growth of uveal melanocytes at physiological concentrations. Exp. Eye Res. 1998, 67, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, L.; Nazari, H.; Sreekumar, P.G.; Kannan, R.; Dustin, L.; Zhu, D.; Barron, E.; Hinton, D.R. TGF-beta2 secretion from RPE decreases with polarization and becomes apically oriented. Cytokine 2015, 71, 394–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokawa, N.; Sotozono, C.; Nishida, K.; Kinoshita, S. High total TGF-beta 2 levels in normal human tears. Curr. Eye. Res. 1996, 15, 341–343. [Google Scholar] [CrossRef]
- Connor, T.B., Jr.; Roberts, A.B.; Sporn, M.B.; Danielpour, D.; Dart, L.L.; Michels, R.G.; de Bustros, S.; Enger, C.; Kato, H.; Lansing, M.; et al. Correlation of fibrosis and transforming growth factor-beta type 2 levels in the eye. J. Clin. Investig. 1989, 83, 1661–1666. [Google Scholar] [CrossRef]
- Fuchshofer, R.; Tamm, E.R. The role of TGF-beta in the pathogenesis of primary open-angle glaucoma. Cell Tissue Res. 2012, 347, 279–290. [Google Scholar] [CrossRef]
- Igarashi, N.; Honjo, M.; Asaoka, R.; Kurano, M.; Yatomi, Y.; Igarashi, K.; Miyata, K.; Kaburaki, T.; Aihara, M. Aqueous autotaxin and TGF-betas are promising diagnostic biomarkers for distinguishing open-angle glaucoma subtypes. Sci. Rep. 2021, 11, 1408. [Google Scholar] [CrossRef]
- Saucedo, L.; Pfister, I.B.; Zandi, S.; Gerhardt, C.; Garweg, J.G. Ocular TGF-beta, matrix metalloproteinases, and TIMP-1 increase with the development and progression of diabetic retinopathy in Type 2 diabetes mellitus. Mediat. Inflamm. 2021, 2021, 9811361. [Google Scholar] [CrossRef]
- Yu, X.B.; Sun, X.H.; Dahan, E.; Guo, W.Y.; Qian, S.H.; Meng, F.R.; Song, Y.L.; Simon, G.J. Increased levels of transforming growth factor-betal and -beta2 in the aqueous humor of patients with neovascular glaucoma. Ophthalmic Surg. Lasers Imaging 2007, 38, 6–14. [Google Scholar] [CrossRef]
- Bredrup, C.; Knappskog, P.M.; Majewski, J.; Rodahl, E.; Boman, H. Congenital stromal dystrophy of the cornea caused by a mutation in the decorin gene. Investig. Ophthalmol. Vis. Sci. 2005, 46, 420–426. [Google Scholar] [CrossRef]
- Rodahl, E.; Van Ginderdeuren, R.; Knappskog, P.M.; Bredrup, C.; Boman, H. A second decorin frame shift mutation in a family with congenital stromal corneal dystrophy. Am. J. Ophthalmol. 2006, 142, 520–521. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Buyank, F.; Sinha, N.R.; Grant, D.G.; Sinha, P.R.; Iozzo, R.V.; Chaurasia, S.S.; Mohan, R.R. Decorin regulates collagen fibrillogenesis during corneal wound healing in mouse in vivo. Exp. Eye Res. 2022, 216, 108933. [Google Scholar] [CrossRef] [PubMed]
- Begum, G.; O’Neill, J.; Chaudhary, R.; Blachford, K.; Snead, D.R.J.; Berry, M.; Scott, R.A.H.; Logan, A.; Blanch, R.J. Altered decorin biology in proliferative vitreoretinopathy: A mechanistic and cohort study. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4929–4936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Beshri, A.S.; Edward, D.P.; Haiti, K.A.; Craven, E.R. Overhanging-dissecting blebs: Immunohistochemical characterization. J. Glaucoma 2018, 27, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, M.G.; Bressler, S.L.; Wight, T.N. The regulated synthesis of versican, decorin, and biglycan: Extracellular matrix proteoglycans that influence cellular phenotype. Crit. Rev. Eukaryot. Gene Expr. 2004, 14, 203–234. [Google Scholar] [CrossRef]
- Sahay, P.; Chakraborty, M.; Rao, A. Global and comparative proteome signatures in the lens capsule, trabecular meshwork, and iris of patients with pseudoexfoliation glaucoma. Front. Mol. Biosci. 2022, 9, 877250. [Google Scholar] [CrossRef]
- Abdullatif, A.M.; Macky, T.A.; Abdullatif, M.M.; Nassar, K.; Grisanti, S.; Mortada, H.A.; Soliman, M.M. Intravitreal decorin preventing proliferative vitreoretinopathy in perforating injuries: A pilot study. Graefes Arch. Clin. Exp. Ophthalmol. 2018, 256, 2473–2481. [Google Scholar] [CrossRef]
- Wormstone, I.M. Posterior capsule opacification: A cell biological perspective. Exp. Eye Res. 2002, 74, 337–347. [Google Scholar] [CrossRef]
- Marcantonio, J.M.; Syam, P.P.; Liu, C.S.; Duncan, G. Epithelial transdifferentiation and cataract in the human lens. Exp. Eye Res. 2003, 77, 339–346. [Google Scholar] [CrossRef]
- de Iongh, R.U.; Lovicu, F.J.; Overbeek, P.A.; Schneider, M.D.; Joya, J.; Hardeman, E.D.; McAvoy, J.W. Requirement for TGFbeta receptor signaling during terminal lens fiber differentiation. Development 2001, 128, 3995–4010. [Google Scholar] [CrossRef]
- de Iongh, R.U.; Wederell, E.; Lovicu, F.J.; McAvoy, J.W. Transforming growth factor-beta-induced epithelial-mesenchymal transition in the lens: A model for cataract formation. Cells Tissues Organs 2005, 179, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Seomun, Y.; Hwang, K.H.; Kim, J.E.; Kim, I.S.; Kim, J.H.; Joo, C.K. Overexpression of the transforming growth factor-beta-inducible gene betaig-h3 in anterior polar cataracts. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1840–1845. [Google Scholar]
- Saika, S.; Miyamoto, T.; Ishida, I.; Shirai, K.; Ohnishi, Y.; Ooshima, A.; McAvoy, J.W. TGFbeta-Smad signalling in postoperative human lens epithelial cells. Br. J. Ophthalmol. 2002, 86, 1428–1433. [Google Scholar] [CrossRef] [PubMed]
- Medvedovic, M.; Tomlinson, C.R.; Call, M.K.; Grogg, M.; Tsonis, P.A. Gene expression and discovery during lens regeneration in mouse: Regulation of epithelial to mesenchymal transition and lens differentiation. Mol. Vis. 2006, 12, 422–440. [Google Scholar]
- Hayashi, N.; Kato, H.; Kiyosawa, T.; Hayashi, H.; Oshima, K.; Yamaoka, M. [The change in immunohistochemical localization of basic fibroblast growth factor (b-FGF) around the lens capsule after extracapsular extraction]. Nihon Ganka Gakkai Zasshi 1991, 95, 621–624. [Google Scholar]
- Lovicu, F.J.; McAvoy, J.W. Localization of acidic fibroblast growth factor, basic fibroblast growth factor, and heparan sulphate proteoglycan in rat lens: Implications for lens polarity and growth patterns. Investig. Ophthalmol. Vis. Sci. 1993, 34, 3355–3365. [Google Scholar]
- Meacock, W.R.; Spalton, D.J.; Stanford, M.R. Role of cytokines in the pathogenesis of posterior capsule opacification. Br. J. Ophthalmol. 2000, 84, 332–336. [Google Scholar] [CrossRef] [Green Version]
- Chamberlain, C.G.; McAvoy, J.W. Induction of lens fibre differentiation by acidic and basic fibroblast growth factor (FGF). Growth Factors 1989, 1, 125–134. [Google Scholar] [CrossRef]
- McAvoy, J.W.; Chamberlain, C.G. Fibroblast growth factor (FGF) induces different responses in lens epithelial cells depending on its concentration. Development 1989, 107, 221–228. [Google Scholar] [CrossRef]
- Nishi, O.; Nishi, K.; Fujiwara, T.; Shirasawa, E.; Ohmoto, Y. Effects of the cytokines on the proliferation of and collagen synthesis by human cataract lens epithelial cells. Br. J. Ophthalmol. 1996, 80, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Wormstone, I.M.; Del Rio-Tsonis, K.; McMahon, G.; Tamiya, S.; Davies, P.D.; Marcantonio, J.M.; Duncan, G. FGF: An autocrine regulator of human lens cell growth independent of added stimuli. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1305–1311. [Google Scholar]
- Kahari, V.M.; Larjava, H.; Uitto, J. Differential regulation of extracellular matrix proteoglycan (PG) gene expression. Transforming growth factor-beta 1 up-regulates biglycan (PGI), and versican (large fibroblast PG) but down-regulates decorin (PGII) mRNA levels in human fibroblasts in culture. J. Biol. Chem. 1991, 266, 10608–10615. [Google Scholar] [PubMed]
- Vuorio, T.; Kahari, V.M.; Black, C.; Vuorio, E. Expression of osteonectin, decorin, and transforming growth factor-beta 1 genes in fibroblasts cultured from patients with systemic sclerosis and morphea. J. Rheumatol. 1991, 18, 247–251. [Google Scholar] [PubMed]
- Shu, C.; Smith, S.M.; Little, C.B.; Melrose, J. Use of FGF-2 and FGF-18 to direct bone marrow stromal stem cells to chondrogenic and osteogenic lineages. Future Sci. OA 2016, 2, FSO142. [Google Scholar] [CrossRef] [Green Version]
- Tan, E.M.; Hoffren, J.; Rouda, S.; Greenbaum, S.; Fox, J.W.t.; Moore, J.H., Jr.; Dodge, G.R. Decorin, versican, and biglycan gene expression by keloid and normal dermal fibroblasts: Differential regulation by basic fibroblast growth factor. Exp. Cell Res. 1993, 209, 200–207. [Google Scholar] [CrossRef]
- Wang, S.C.; Lin, C.T.; Nie, D.B.; Ouyang, H.S. The effect of basic fibroblast growth factor on the gene expression of decorin by periodontal ligament fibroblasts in culture. Hua Xi Kou Qiang Yi Xue Za Zhi 2008, 26, 352–354. [Google Scholar]
- Sonal, D. Prevention of IGF-1 and TGFbeta stimulated type II collagen and decorin expression by bFGF and identification of IGF-1 mRNA transcripts in articular chondrocytes. Matrix Biol. 2001, 20, 233–242. [Google Scholar] [CrossRef]
- Trowbridge, J.M.; Rudisill, J.A.; Ron, D.; Gallo, R.L. Dermatan sulfate binds and potentiates activity of keratinocyte growth factor (FGF-7). J. Biol. Chem 2002, 277, 42815–42820. [Google Scholar] [CrossRef]
- Penc, S.F.; Pomahac, B.; Winkler, T.; Dorschner, R.A.; Eriksson, E.; Herndon, M.; Gallo, R.L. Dermatan sulfate released after injury is a potent promoter of fibroblast growth factor-2 function. J. Biol. Chem. 1998, 273, 28116–28121. [Google Scholar] [CrossRef] [Green Version]
- Gunning, P. Emerging issues for tropomyosin structure, regulation, function and pathology. Adv. Exp. Med. Biol. 2008, 644, 293–298. [Google Scholar]
- Gunning, P.W.; Schevzov, G.; Kee, A.J.; Hardeman, E.C. Tropomyosin isoforms: Divining rods for actin cytoskeleton function. Trends Cell Biol. 2005, 15, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Shu, D.Y.; Lovicu, F.J. Enhanced EGF receptor-signaling potentiates TGFbeta-induced lens epithelial-mesenchymal transition. Exp. Eye Res. 2019, 185, 107693. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Shibata, S.; Ishigaki, Y.; Kiyokawa, E.; Ikawa, M.; Singh, D.P.; Sasaki, H.; Kubo, E. Tropomyosin 2 heterozygous knockout in mice using CRISPR-Cas9 system displays the inhibition of injury-induced epithelial-mesenchymal transition, and lens opacity. Mech. Ageing Dev. 2018, 171, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Wormstone, I.M.; Tamiya, S.; Anderson, I.; Duncan, G. TGF-beta2-induced matrix modification and cell transdifferentiation in the human lens capsular bag. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2301–2308. [Google Scholar]
- Dwivedi, D.J.; Pino, G.; Banh, A.; Nathu, Z.; Howchin, D.; Margetts, P.; Sivak, J.G.; West-Mays, J.A. Matrix metalloproteinase inhibitors suppress transforming growth factor-beta-induced subcapsular cataract formation. Am. J. Pathol. 2006, 168, 69–79. [Google Scholar] [CrossRef]
- Zhao, H.; Yang, Y.; Rizo, C.M.; Overbeek, P.A.; Robinson, M.L. Insertion of a Pax6 consensus binding site into the alphaA-crystallin promoter acts as a lens epithelial cell enhancer in transgenic mice. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1930–1939. [Google Scholar] [CrossRef] [Green Version]
- Du, S.; Shao, J.; Xie, D.; Zhang, F. Decorin inhibits glucose-induced lens epithelial cell apoptosis via suppressing p22phox-p38 MAPK signaling pathway. PLoS ONE 2020, 15, e0224251. [Google Scholar] [CrossRef]
- Ozay, R.; Turkoglu, E.; Gurer, B.; Dolgun, H.; Evirgen, O.; Erguder, B.I.; Hayirli, N.; Gurses, L.; Sekerci, Z.; Yilmaz, E.R. Does Decorin Protect Neuronal Tissue via Its Antioxidant and Antiinflammatory Activity from Traumatic Brain Injury? An Experimental Study. World Neurosurg. 2017, 97, 407–415. [Google Scholar] [CrossRef]
- Fatma, N.; Kubo, E.; Sharma, P.; Beier, D.R.; Singh, D.P. Impaired homeostasis and phenotypic abnormalities in Prdx6-/- mice lens epithelial cells by reactive oxygen species: Increased expression and activation of TGFbeta. Cell Death Differ. 2005, 12, 734–750. [Google Scholar] [CrossRef]
- Bondi, C.D.; Manickam, N.; Lee, D.Y.; Block, K.; Gorin, Y.; Abboud, H.E.; Barnes, J.L. NAD(P)H oxidase mediates TGF-beta1-induced activation of kidney myofibroblasts. J. Am. Soc. Nephrol. 2010, 21, 93–102. [Google Scholar] [CrossRef]
- Fatma, N.; Kubo, E.; Toris, C.B.; Stamer, W.D.; Camras, C.B.; Singh, D.P. PRDX6 attenuates oxidative stress- and TGFbeta-induced abnormalities of human trabecular meshwork cells. Free Radic. Res. 2009, 43, 783–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohba, M.; Shibanuma, M.; Kuroki, T.; Nose, K. Production of hydrogen peroxide by transforming growth factor-beta 1 and its involvement in induction of egr-1 in mouse osteoblastic cells. J. Cell Biol. 1994, 126, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Herrera, B.; Murillo, M.M.; Alvarez-Barrientos, A.; Beltran, J.; Fernandez, M.; Fabregat, I. Source of early reactive oxygen species in the apoptosis induced by transforming growth factor-beta in fetal rat hepatocytes. Free Radic. Biol. Med. 2004, 36, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Herrera, B.; Alvarez, A.M.; Sanchez, A.; Fernandez, M.; Roncero, C.; Benito, M.; Fabregat, I. Reactive oxygen species (ROS) mediates the mitochondrial-dependent apoptosis induced by transforming growth factor (beta) in fetal hepatocytes. FASEB J. 2001, 15, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, A.; Kozlov, A.V. Biological Activities of Reactive Oxygen and Nitrogen Species: Oxidative Stress versus Signal Transduction. Biomolecules 2015, 5, 472–484. [Google Scholar] [CrossRef] [Green Version]
- Blochberger, T.C.; Cornuet, P.K.; Hassell, J.R. Isolation and partial characterization of lumican and decorin from adult chicken corneas. A keratan sulfate-containing isoform of decorin is developmentally regulated. J. Biol. Chem 1992, 267, 20613–20619. [Google Scholar] [CrossRef]
- Kamma-Lorger, C.S.; Pinali, C.; Martinez, J.C.; Harris, J.; Young, R.D.; Bredrup, C.; Crosas, E.; Malfois, M.; Rodahl, E.; Meek, K.M.; et al. Role of decorin core protein in collagen organisation in congenital stromal corneal dystrophy (CSCD). PLoS ONE 2016, 11, e0147948. [Google Scholar] [CrossRef] [Green Version]
- Bredrup, C.; Stang, E.; Bruland, O.; Palka, B.P.; Young, R.D.; Haavik, J.; Knappskog, P.M.; Rodahl, E. Decorin accumulation contributes to the stromal opacities found in congenital stromal corneal dystrophy. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5578–5582. [Google Scholar] [CrossRef] [Green Version]
- Mohan, R.R.; Tandon, A.; Sharma, A.; Cowden, J.W.; Tovey, J.C. Significant inhibition of corneal scarring in vivo with tissue-selective, targeted AAV5 decorin gene therapy. Investig. Ophthalmol. Vis. Sci. 2011, 52, 4833–4841. [Google Scholar] [CrossRef] [Green Version]
- Mohan, R.R.; Tovey, J.C.; Sharma, A.; Schultz, G.S.; Cowden, J.W.; Tandon, A. Targeted decorin gene therapy delivered with adeno-associated virus effectively retards corneal neovascularization in vivo. PLoS ONE 2011, 6, e26432. [Google Scholar] [CrossRef]
- Mohan, R.R.; Balne, P.K.; Muayad, M.S.; Tripathi, R.; Sinha, N.R.; Gupta, S.; An, J.A.; Sinha, P.R.; Hesemann, N.P. Six-Month In Vivo Safety Profiling of Topical Ocular AAV5-Decorin Gene Transfer. Transl. Vis. Sci. Technol. 2021, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Hill, L.J.; Moakes, R.J.A.; Vareechon, C.; Butt, G.; Ng, A.; Brock, K.; Chouhan, G.; Vincent, R.C.; Abbondante, S.; Williams, R.L.; et al. Sustained release of decorin to the surface of the eye enables scarless corneal regeneration. NPJ Regen. Med. 2018, 3, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Downie, L.E.; Grover, L.M.; Moakes, R.J.A.; Rauz, S.; Logan, A.; Jiao, H.; Hill, L.J.; Chinnery, H.R. The neuroregenerative effects of topical decorin on the injured mouse cornea. J. Neuroinflamm. 2020, 17, 142. [Google Scholar] [CrossRef] [PubMed]
- Collaborative Normal-Tension Glaucoma Study Group. The effectiveness of intraocular pressure reduction in the treatment of normal-tension glaucoma. Am. J. Ophthalmol. 1998, 126, 498–505. [Google Scholar] [CrossRef]
- Collaborative Normal-Tension Glaucoma Study Group. Comparison of glaucomatous progression between untreated patients with normal-tension glaucoma and patients with therapeutically reduced intraocular pressures. Am. J. Ophthalmol. 1998, 126, 487–497. [Google Scholar] [CrossRef]
- Moses, R.A. The effect of intraocular pressure on resistance to outflow. Surv. Ophthalmol. 1977, 22, 88–100. [Google Scholar] [CrossRef]
- Wiederholt, M. Direct involvement of trabecular meshwork in the regulation of aqueous humor outflow. Curr. Opin. Ophthalmol. 1998, 9, 46–49. [Google Scholar] [CrossRef]
- Overby, D.R.; Zhou, E.H.; Vargas-Pinto, R.; Pedrigi, R.M.; Fuchshofer, R.; Braakman, S.T.; Gupta, R.; Perkumas, K.M.; Sherwood, J.M.; Vahabikashi, A.; et al. Altered mechanobiology of Schlemm’s canal endothelial cells in glaucoma. Proc. Natl. Acad. Sci. USA 2014, 111, 13876–13881. [Google Scholar] [CrossRef] [Green Version]
- Kirwan, R.P.; Wordinger, R.J.; Clark, A.F.; O’Brien, C.J. Differential global and extra-cellular matrix focused gene expression patterns between normal and glaucomatous human lamina cribrosa cells. Mol. Vis. 2009, 15, 76–88. [Google Scholar]
- Pena, J.D.; Taylor, A.W.; Ricard, C.S.; Vidal, I.; Hernandez, M.R. Transforming growth factor beta isoforms in human optic nerve heads. Br. J. Ophthalmol. 1999, 83, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.; Dillinger, A.E.; Ohlmann, A.; Iozzo, R.V.; Fuchshofer, R. Decorin-an antagonist of TGF-beta in astrocytes of the optic nerve. Int. J. Mol. Sci. 2021, 22, 7660. [Google Scholar] [CrossRef] [PubMed]
- Addicks, E.M.; Quigley, H.A.; Green, W.R.; Robin, A.L. Histologic characteristics of filtering blebs in glaucomatous eyes. Arch. Ophthalmol. 1983, 101, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Jampel, H.D.; Morrison, J.; Vocci, M.; Quigley, H. Identification of fibrin/fibrinogen in glaucoma filtration surgery wounds. Ophthalmic Surg. Lasers Imaging Retin. 1988, 19, 576–579. [Google Scholar] [CrossRef]
- Jampel, H.D.; McGuigan, L.J.; Dunkelberger, G.R.; L’Hernault, N.L.; Quigley, H.A. Cellular proliferation after experimental glaucoma filtration surgery. Arch. Ophthalmol. 1988, 106, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Grisanti, S.; Szurman, P.; Warga, M.; Kaczmarek, R.; Ziemssen, F.; Tatar, O.; Bartz-Schmidt, K.U. Decorin modulates wound healing in experimental glaucoma filtration surgery: A pilot study. Investig. Ophthalmol. Vis. Sci. 2005, 46, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Sahay, P.; Reddy, S.; Prusty, B.K.; Modak, R.; Rao, A. TGFbeta1, MMPs and cytokines profiles in ocular surface: Possible tear biomarkers for pseudoexfoliation. PLoS ONE 2021, 16, e0249759. [Google Scholar] [CrossRef]
- Rattner, A.; Nathans, J. Macular degeneration: Recent advances and therapeutic opportunities. Nat. Rev. Neurosci. 2006, 7, 860–872. [Google Scholar] [CrossRef]
- Du, S.; Wang, S.; Wu, Q.; Hu, J.; Li, T. Decorin inhibits angiogenic potential of choroid-retinal endothelial cells by downregulating hypoxia-induced Met, Rac1, HIF-1alpha and VEGF expression in cocultured retinal pigment epithelial cells. Exp. Eye Res. 2013, 116, 151–160. [Google Scholar] [CrossRef]
- Wang, X.; Ma, W.; Han, S.; Meng, Z.; Zhao, L.; Yin, Y.; Wang, Y.; Li, J. TGF-beta participates choroid neovascularization through Smad2/3-VEGF/TNF-alpha signaling in mice with Laser-induced wet age-related macular degeneration. Sci. Rep. 2017, 7, 9672. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.E.; Park, Y.J.; Chung, I.Y.; Seo, S.W.; Park, J.M.; Yoo, J.M.; Song, J.K. Gene expression changes in a rat model of oxygen-induced retinopathy. Korean J. Ophthalmol. 2011, 25, 42–47. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.J.; Kim, Y.H.; Choi, W.S.; Chung, I.Y.; Yoo, J.M. Treatment with triamcinolone acetonide prevents decreased retinal levels of decorin in a rat model of oxygen-induced retinopathy. Curr. Eye Res. 2010, 35, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Duran, G.L.S.; Balbaba, M.; Colakoglu, N.; Bulmus, O.; Ulas, F.; Eroksuz, Y. Effect of Decorin and Bevacizumab on oxygen-induced retinopathy in rat models: A comparative study. Indian J. Ophthalmol. 2021, 69, 369–373. [Google Scholar] [CrossRef]
- Ryan, S.J. The pathophysiology of proliferative vitreoretinopathy in its management. Am. J. Ophthalmol. 1985, 100, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.; Hoerauf, H. TGF-ss and RPE-derived cells in taut subretinal strands from patients with proliferative vitreoretinopathy. Eur. J. Ophthalmol. 2011, 21, 422–426. [Google Scholar] [CrossRef]
- Nassar, K.; Luke, J.; Luke, M.; Kamal, M.; Abd El-Nabi, E.; Soliman, M.; Rohrbach, M.; Grisanti, S. The novel use of decorin in prevention of the development of proliferative vitreoretinopathy (PVR). Graefes Arch. Clin. Exp. Ophthalmol. 2011, 249, 1649–1660. [Google Scholar] [CrossRef]
- Jarvinen, T.A. Design of target-seeking antifibrotic compounds. Methods Enzymol. 2012, 509, 243–261. [Google Scholar] [CrossRef]
Figure 1.
Structural and putative functional features of the decorin (DCN) molecule. Mammalian decorin contains core proteins and a glycosaminoglycan (GAG) side chain attached to a serine residue near the N terminus. Four domains of core proteins (I–IV) of DCN are shown. In domain I, the initial 16-amino-acid long signal peptide directs the incipient core protein to the rough endoplasmic reticulum and is cleaved via the co-translational pathway. In domain II, the propeptide is composed of 14 amino acids and regulates the attachment of the GAG chain. The protein core consists of tandem leucine-rich repeats (LRRs) flanking cysteine-rich disulfide domains. Twelve LRRs in domain III comprise the major central domain. Most decorin ligands share an interface with decorin at its core protein, an important hub for the extracellular matrix, cell surface receptors, and growth factors. Decorin has a high-affinity binding site for collagen (Col) at LRRs 4–6 and a low-affinity site at the C-terminus. The binding sites of TGF-β isoforms are LRR4 and LRR5 at the decorin core protein in domain II. The LRR7 domain binds to EGFR and ErB4 and activates the MAPK pathway. Decorin interacts with multiple growth factor signaling pathways. The carboxyl terminus domain and binding sites for fibronectin Fn and collagen Col can be found in domain IV.
Figure 1.
Structural and putative functional features of the decorin (DCN) molecule. Mammalian decorin contains core proteins and a glycosaminoglycan (GAG) side chain attached to a serine residue near the N terminus. Four domains of core proteins (I–IV) of DCN are shown. In domain I, the initial 16-amino-acid long signal peptide directs the incipient core protein to the rough endoplasmic reticulum and is cleaved via the co-translational pathway. In domain II, the propeptide is composed of 14 amino acids and regulates the attachment of the GAG chain. The protein core consists of tandem leucine-rich repeats (LRRs) flanking cysteine-rich disulfide domains. Twelve LRRs in domain III comprise the major central domain. Most decorin ligands share an interface with decorin at its core protein, an important hub for the extracellular matrix, cell surface receptors, and growth factors. Decorin has a high-affinity binding site for collagen (Col) at LRRs 4–6 and a low-affinity site at the C-terminus. The binding sites of TGF-β isoforms are LRR4 and LRR5 at the decorin core protein in domain II. The LRR7 domain binds to EGFR and ErB4 and activates the MAPK pathway. Decorin interacts with multiple growth factor signaling pathways. The carboxyl terminus domain and binding sites for fibronectin Fn and collagen Col can be found in domain IV.

Figure 2.
Schematic representation of the human decorin promoter. Consensus sequences for cis-regulatory elements are labeled.
Figure 2.
Schematic representation of the human decorin promoter. Consensus sequences for cis-regulatory elements are labeled.

Figure 3.
Schematic representation of the role of decorin (DCN) in the LECs. FGF2 induces the expression of DCN, but its mechanism is unclear. FGF2 binds to the GAG chain of DCN enhancing the activity of FGF2 and potentially promoting LEC proliferation and differentiation. Higher levels of DCN bind to TGF-β and inhibit its activity. On the other hand, TGF-β binds to the promoter of DCN and represses decorin expression, possibly promoting EMT and fibrosis. Increased expression of decorin inhibits the expression of Tpm1 and Tpm2 and may promote normal cell growth and wound healing. DCN, decorin; LECs, lens epithelial cells; FGF2, fibroblast growth factor 2; GAG, glycosamino-glycan; TGF-β, transforming growth factor-β; EMT, epithelial–mesenchymal transition.
Figure 3.
Schematic representation of the role of decorin (DCN) in the LECs. FGF2 induces the expression of DCN, but its mechanism is unclear. FGF2 binds to the GAG chain of DCN enhancing the activity of FGF2 and potentially promoting LEC proliferation and differentiation. Higher levels of DCN bind to TGF-β and inhibit its activity. On the other hand, TGF-β binds to the promoter of DCN and represses decorin expression, possibly promoting EMT and fibrosis. Increased expression of decorin inhibits the expression of Tpm1 and Tpm2 and may promote normal cell growth and wound healing. DCN, decorin; LECs, lens epithelial cells; FGF2, fibroblast growth factor 2; GAG, glycosamino-glycan; TGF-β, transforming growth factor-β; EMT, epithelial–mesenchymal transition.

Figure 4.
Histochemical analysis of lenses in 48-week-old (W) hDCN-Tg mice. Adult lenses in 48 W hDCN-Tg animals (A) did not show lens fiber damage and pathological changes and did not differ from those in WT mice (B). Overexpression of hDCN did not affect the histological changes in lens devel-opment. Scale bar = 200 µm.
Figure 4.
Histochemical analysis of lenses in 48-week-old (W) hDCN-Tg mice. Adult lenses in 48 W hDCN-Tg animals (A) did not show lens fiber damage and pathological changes and did not differ from those in WT mice (B). Overexpression of hDCN did not affect the histological changes in lens devel-opment. Scale bar = 200 µm.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kubo, E.; Shibata, S.; Shibata, T.; Sasaki, H.; Singh, D.P. Role of Decorin in the Lens and Ocular Diseases. Cells 2023, 12, 74. https://doi.org/10.3390/cells12010074
AMA Style
Kubo E, Shibata S, Shibata T, Sasaki H, Singh DP. Role of Decorin in the Lens and Ocular Diseases. Cells. 2023; 12(1):74. https://doi.org/10.3390/cells12010074
Chicago/Turabian StyleKubo, Eri, Shinsuke Shibata, Teppei Shibata, Hiroshi Sasaki, and Dhirendra P. Singh. 2023. "Role of Decorin in the Lens and Ocular Diseases" Cells 12, no. 1: 74. https://doi.org/10.3390/cells12010074
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.