Modeling Biofilms in Water Systems with New Variables: A Review

1
School of Environmental Science and Engineering, Shandong University, Jinan 250100, China
2
School of Municipal and Environmental Engineering, Shandong Jianzhu University, Jinan 250101, China
*
Authors to whom correspondence should be addressed.
Water 2017, 9(7), 462; https://doi.org/10.3390/w9070462
Submission received: 30 April 2017
/
Revised: 8 June 2017
/
Accepted: 23 June 2017
/
Published: 4 July 2017
(This article belongs to the Special Issue Modeling of Water Systems)
{kind=link}
{kind=link}
Abstract
:In nature, microorganisms mainly live in biofilms rather than planktonically to defend against various environmental stimuli. Understanding and predicting the dynamics and mechanisms of biofilms is of grand importance to human life. Besides experimental approaches, modeling provides a powerful tool to describe biofilms mathematically, and the in silico simulation offers a deep insight into the underlying functional machineries of biofilms. In this review, we briefly summarized the main advances in biofilm modeling, including the 1D model, multidimensional model, as well as the incorporation of extracellular polymeric substance and quorum sensing signals. Specifically, we focus on recent experimental advances in biofilms, which may challenge the current modeling systems via newly discovered properties and novel participants in natural water systems. We hope this article could build a bridge between in silico modeling and new experimental discoveries, and, in particular, we wish to attract more attention to biofilm-related-environmental issues at various scales.
1. Introduction
Microorganisms tend to grow multicellularly rather than singularly to improve the survivability in varying environments with stimuli [1,2]. Biofilm is the dominant multicellular structure in nature, and it exists in almost all ecological niches and engineered systems, like in wastewater treatment processes [3,4] and on medical equipment [5,6]. In all cases, biofilms are structured by the matrix of microbes, extracellular polymeric substances (EPS), and other abiotic materials, including nutrients, signal molecules, and minerals [1]. The properties and functions of biofilms are essential to human society at various aspects and scales. In natural water systems, the cyanobacterial biofilm regulates the biocalcification, a fundamental bioprocess which contributes to local and global carbon cycles [7,8,9]. In addition, well-structured and functional biofilms or microbial aggregates facilitate the degradation and removal of pollutants [10,11]. Therefore, a systematic investigation of the biofilm dynamics benefits various aspects of human life.
The properties and functions of biofilms are attributed to complex structures and microbial metabolic networks. Specifically, the biofilm matrix results in gradients of chemicals across the biofilm as well as physical and chemical benefits to biofilm-hosting microbes [2]. For instance, microbes in biofilms have much higher tolerance, sometimes hundreds of times higher, toward antibiotics than the planktonic ones, thus jeopardizing the utilization of antimicrobial agents [12,13]. Moreover, it has been reported that microbes in biofilms have different genetic properties, and even the transcription profiles of the same microbial strain vary temporally and spatially [14,15,16]. These variations are putatively due to the distribution of nutrients, O2, pH, and stressors in biofilms. One theory proposed that these varied genetic properties result from the structure of biofilms as well [17]. Therefore, an in-depth understanding and description of biofilms at different scales and aspects is quite necessary but challenging in order to harness the biofilms for desired purposes.
Besides laboratory approaches and ‘-omic’ information, modeling has exhibited promising potential and promoted the elucidation of biofilm dynamics and underlying mechanisms [18]. Moreover, modeling possesses the capability of not only mathematic description but also prediction of the evolution of biofilms at microbial community levels. Since 40 years ago, researchers have invested lots of efforts to develop theories and models to simulate biofilms [19,20]. Based on the spatial and temporal properties of biofilms, several models have been purposed, including the one-dimensional (1D) model, which addresses the mass transfer, reaction/diffusion, and detachments, as well as multidimensional models, which allow the diversity of microbial ecology to be considered [19,21].
As environmental conditions trigger the formation of biofilms, many models have considered environmental stimuli when describing the multicellular structure. However, biofilms have encountered or revealed new ‘variables’ in both natural and engineered systems as human society advances, like new stressors [22,23,24,25,26] and novel properties of EPS [27]. Moreover, several issues need to be addressed via not only model microorganism—like Escherichia coli, Bacillus subtilis and Pseudomonas aeruginosa—but also candidates with special ecological functions, such as cyanobacteria Synechococcus sp. and Synechocystis sp. [7]. In this review, we summarized recent advances in biofilm modeling, especially biofilms in natural water systems with novel challenges. We hope this article provides a bridge between in silico modeling and new discoveries in laboratory, and, in particular, we want to attract more attention to biofilm-related-environmental issues.
2. Modeling the Dynamics of Biofilms
2.1. The Evolution of Biofilms
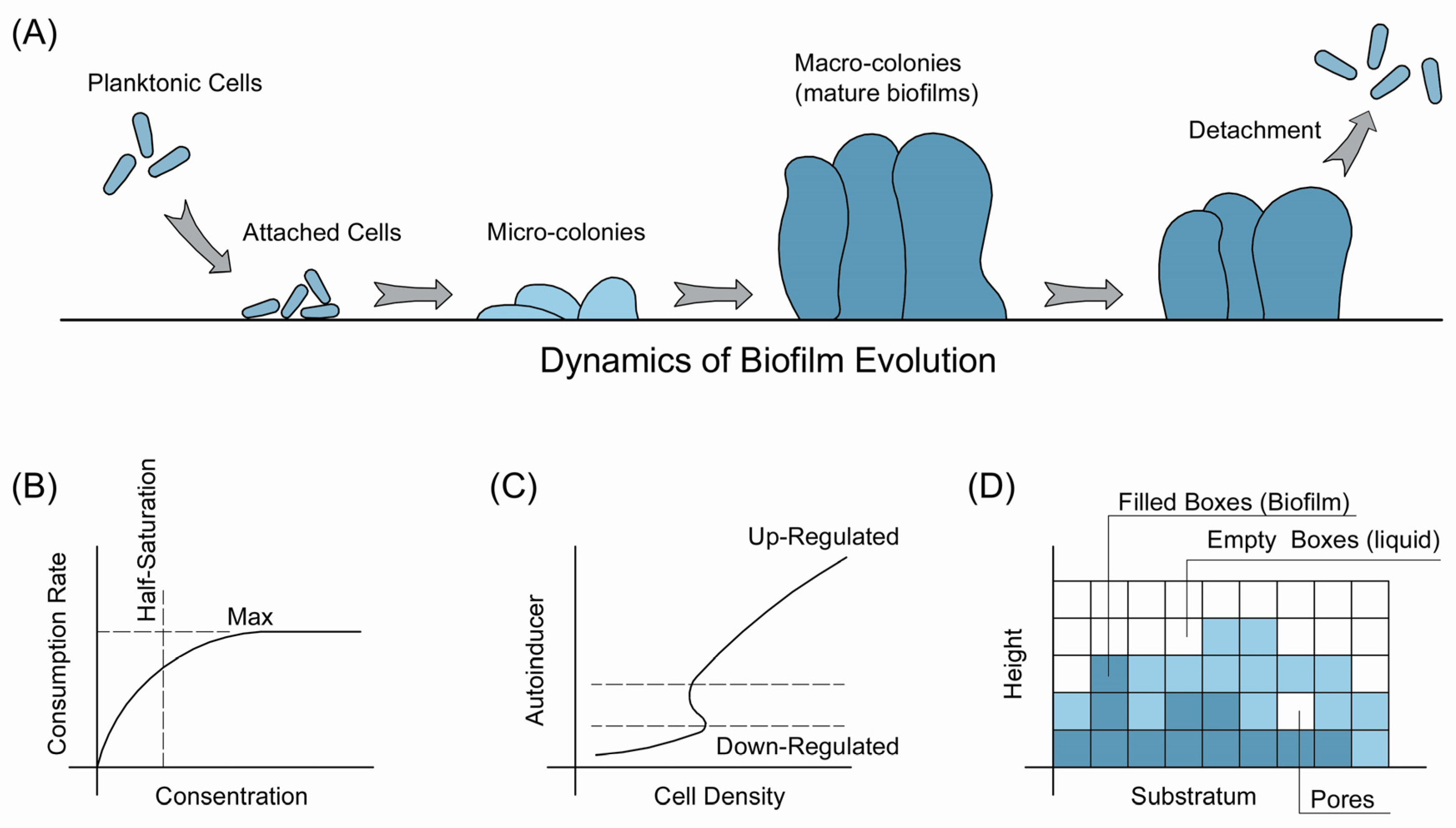
The formation of biofilms typically follows a defined scenario (Figure 1) [12,21,28]. In certain external environmental conditions, planktonic cells attached to the substratum, which also refers to adhesion [29]. Two types of adhesion occur: reversible and irreversible adhesion. Cells swim away when reversibly attached to a surface, while irreversibly attached cells begin to grow and form microcolonies. This is the initial stage of a biofilm. Then, the biofilm matures to macrocolonies with mushroom-like tower structures [30,31]. Finally, the biofilm reaches a pseudo-steady state, and the growth and detachment of cells reach equilibrium.
Two dominant theories have been purposed to explain this dynamic. One is the developmental theory suggested by O’Toole et al. [17,28]. The main idea is that the formation of biofilms is genetically programmed. Under certain environmental conditions, a genetic pathway will be triggered and the transcription was initiated to regulate the formation of biofilms. Concerns about the developmental model consist of the failures to identify a designed genetic pathway coding the formation of biofilms [32,33], plus the lacking of checkpoints of different stages during development [17]. Another theory hypothesizes that the formation of biofilms is the ecological adaption of individual members via cellular plasticity [34,35]. According to this theory, microorganisms optimize their surface properties to facilitate colonization and mediate the cell interactions in response to environmental conditions, and the biofilm matrix attributed to the accumulative phenotypes of the individual adaption. Supporters of this hypothesis also suggested the apparently cooperative phenotypes of biofilm could be better explained by the physiology of single cells [17].
Despite the fundamental differences between these two theories, typical biofilms go through the abovementioned five stages: adhesion, growth, microcolony, microcolony, and detachment/erosion (Figure 1A). The 1D model is a well-established mathematical description of these stages, especially biofilms colonized by one microbial strain [21]. The first step of settling the 1D model is to determine the concentrations of nutrients on the surface of biofilm, which is also the interface between bulk liquid and biofilm. To do so, researchers have assumed a boundary layer between the external environment and biofilm, where diffusion and advection dominate the mass transfer. Thus, the film theory can be employed to simulate the concentrations of interested factors. After predicting substrate on the surface, the growth of biofilms can be simulated based on two principles. One is that the conversion of substrates to biomass is regulated by the diffusion of substrates, together with the assumption of the steady-state of the biofilm, where concentrations of substrates maintain stability (Figure 1B). In addition, the 1D model was became more reliable via dividing the substratum to segments and the distribution of biomass on substratum could be simulated [36,37].
The 1D model successfully mimicked the mass transfer and distribution within biofilms. However, the dynamics of biofilms cannot be best simulated by this model, because biofilms exhibited complex structures and are influenced by various inner and environmental factors [38,39]. More importantly, biofilms in both natural and engineered systems usually consist of different microorganisms rather than one single strain. Therefore, multidimensional models were developed to simulate various ‘biophases’ with distinct properties, including species, nutrients, water, etc. [40]. Two main multidimensional models have been applied recently: the cellular automata (CA) model and individual based model [41,42]. In the CA model, the biofilm matrix is divided into several compartments with fixed volumes (Figure 1D). The biomass in the biofilm matrix will move into the compartment and, if the room is full, the extra biomass will move to available compartments nearby. The CA model describes this process mathematically and displays the spatial heterogeneity using a discrete formulation [43,44]. The individual based model, on the other hand, relates the properties of single cells with the biofilms [45]. It is actually a developed CA model without artificially defined compartments but microbes or microbial aggregates as the simulation unit, such as the BacSim model by Kreft et al. [46].
2.2. Extracellular Polymeric Substances
EPS are of great significance in biofilms. The microbes only account for 10% of the dry weight of biofilms, while EPS contribute to the rest of the mass [1]. Typically, EPS consist of exoproteins (PN), exopolysaccharides (PS), extracellular DNAs (eDNA), as well as other abiotic biomolecules [47,48]. These macromolecules are excreted by the biofilm-hosting microbes or come from the cell lysis or the hydrolysis of macromolecules. EPS construct the scaffolds of biofilms, maintain the architectures, and regulate the adhesion and cohesion of inside cells via both the physical networks and chemical groups [1].
Because of the importance of EPS, researchers have embedded these features into current models based on the molecular knowledge of EPS, and several assumptions have been made [49]. First, the production of EPS is associated with cell growth and the concentrations of substrates. Secondly, EPS impact the density of biofilms because of their dominant volumetric fraction. Thirdly, EPS are the predominant adsorbents for chemicals and particles in the matrix. Additionally, EPS may serve as energy sources in starving conditions. The model was developed and conducted via the partial differential equations by Horn et al. [49]. Interestingly, the simulation results of an EPS-integrated model generated similar results to those that merely consider biofilms. Nevertheless, these models benefit from the understanding of distribution of chemicals in the biofilm matrix, such as heavy metals and chlorinated organic substances, which are quite important in the natural ecosystems [19].
These models confront challenges as knowledge of EPS advances. At the initial stage of EPS-related research, PS were considered as the most significant component for maintaining the structure of biofilms [1,50]. Therefore, early concepts usually regard PS as the ‘glues’, which keep integrity of biofilms. For instance, the model microorganism for biofilm study, P. aeruginosa, produces three types of PS: alginate, Psl and Pel [51]. Alginate is the most studied PS and often used as the model chemical to evaluate the functions of PS [52]. It has been reported that alginate contributes to the initial stage of biofilm formation and the mechanical stability of mature biofilms, and, without the well-produced alginate, biofilms cannot be formed as well as the wildtype strains do.
However, the functions of EPS are not constrained in ‘glues’. Recent discoveries reported that some of EPS exhibit antibiofilm properties. In 2006, Valle et al., discovered that the biofilm by E. coli MG1655 would be inhibited when co-cultured with the uropathogenic CFT073 strain [53]. The inhibitory effects resulted from the Group II capsular polysaccharides in the supernatant of the co-culture, which was produced by E. coli. A systematic screening of various E. coli strains revealed that strains that are capable of synthesizing the Group II capsular polysaccharides conferred similar deficiency on the formation of biofilms [53]. Another study suggested that a linear polysaccharide, PAM galactan, with the structure →3)-β-d-Galf-(1→6)-β-d-Galf-(1→, exhibited broad-spectrum inhibition activity on biofilms, including biofilms by Aggregatibacter actinomycetemcomitans, Klebsiella pneumoniae, Staphylococcus aureus, Staphylococcus epidermidis, Candida albicans, as well as its producing strain K. kingae [54]. Besides the antibiofilm effects on their producing strains, these antibiofilm-PS usually work as weapons to protect the biofilms and defend the invading strains. Interestingly, the abovementioned ‘glues’—Psl and Pel—also exhibited antibiofilm effects on competing strains. The PS from wild-type P. aeruginosa PAO1 showed reduced biofilm dispersal effects on S. epidermids, while those from Pel and Psl mutant strain do not possess these properties, indicating the roles of Psl and Pel on the competition of environmental strains when forming biofilms [55].
In addition, the roles of PN have been discovered [1]. PN contain various functional enzymes and work on the degradation of EPS, such as PS, eDNA, cellulose, or lipids. These enzymatic functions usually assist with the digestion of substrate for biofilm-dwelling microbes. Moreover, PN also show structural functions to biofilms. Those non-enzymatic PN consist of cell surface-associated and carbohydrate-binding proteins, which bind the cell surface and EPS as well as largely contribute to the surface properties of cells. For example, the galactose-specific lectin LecA and the fructose-specific lectin LecB are categorized as structural proteins [56,57]. Also, recent study discovered that the ratio between PN and PS contributes even more to the surface properties of microbe in the matrix than those alone [48,52]. Furthermore, Smith et al. [58] found that PN not only work as structural components, but also have the recruitment function to new cells. The PN of this new function, RbmA, was discovered in the diarrheal pathogen Vibrio cholerae. The activity of RbmA is both activated intracellularly and depends on the interaction between RbmA and biofilms. Therefore, these newly discovered properties might require novel model systems to describe these new functions.
2.3. Quorum Sensing (QS)
Quorum sensing (QS) is a population-density-dependent cell-to-cell communicating process. It regulates the activity of a group of microbes when working toward a shared objective [59]. One of these phenotypes is the formation of biofilms. Briefly, the working mechanism of QS follows the following pattern. The signal molecules, known as autoinducers, are produced and released with the growth of cells, and, when the concentrations of these autoinducers achieve the threshold value, certain genetic networks will be upregulated. These networks typically consist of genes associated with luminescence, biofilm formation, plus virulence factor production. When modeling the process, the QS system is considered as a switch in that when the concentrations of autoinducers are low, the phenotype of microbes are stable, while, if the levels of autoinducers reach the threshold value, the phenotype changes to another statue [19]. As such, the production kinetics of autoinducers by intracellular enzymes, the inside and outside cell concentrations of autoinducers should be determined. Several assumptions have been made. First, the substrate for the production of autoinducers are sufficient. Second, the production of autoinducers, such as 3-oxo-C12-HSL and C4-HSL, and autoinducer-producing enzymes—such as LasR, LasI, RhlR, and RsaL—are following the Michaelis–Menten kinetics. Then, the autoinducers travel freely across the cell membrane. Klapper and Dockery summarized this model [19], and they pointed out that one important problem is to determine how cell density regulates the QS systems (Figure 1C). The local volume fraction of cells (ρ) was introduced and a bifurcation diagram of the model was identified in silico where ρ functioned as the switch between two stable phases of cells as abovementioned.
The first model coupling QS into modeling of biofilms was reported by Chopp et al. [60,61]. The ordinary differential equation-based spatiotemporal model discussed the correlations between the physical parameters and autoinducers in the biofilms matrix, where the biofilms were categorized into biotic part, consisting of living cells and abiotic materials, including EPS and dead cells. Then, Ward et al. [62] considered the cell growth during the interaction between QS and biofilm formation. This model was based on a previous work, which described the QS system by three components: switch-off cells, switch-on cells and the switch, autoinducers, and assumed that higher concentrations of autoinducers increased the proportion of switch-on cells [63]. Unlike the model by Chopp et al. [60,61], this newly purposed model hypothesized that the biofilms are thin layers rather than a one dimensional description. The modeling results were in agreement with the experimental data, indicating an intermediate phase of biofilms, in which there were still switch-off cells in mature biofilms, and these cells would soon be upregulated along with the increased concentrations of autoinducers. Ward and King [64] further developed this model by assuming the biofilms as viscous fluid with low Reynolds' number. In this model, two distinct boundary conditions have been purposed: one is the shear stress-free condition, and the other is the no-slip condition. This model also allows more variables to be incorporated to better simulate the formation of biofilms and evaluate the effects of these variables on biofilm dynamics.
These models, however, are based on the typical QS systems in P. aeruginosa, which contain two pairs of autoinducers. However, more autoinducer pairs have been discovered, and the functions of QS have been elucidated more. For example, Papenfort et al. [65] identified another QS signal pairs, the 3,5-dimethylpyrazin-2-ol-VqmA pair, in Vibrio cholerae. The working mechanisms are similar but different to the Lux systems, as this system not only regulates the biofilm formation of its producing strain, but also may influence those by competing strains in the same niche. It is necessary to combine these new signal pairs and their properties in biofilm modeling when considering QS.
3. Biofilms in Water Systems Challenged by Novel Variables
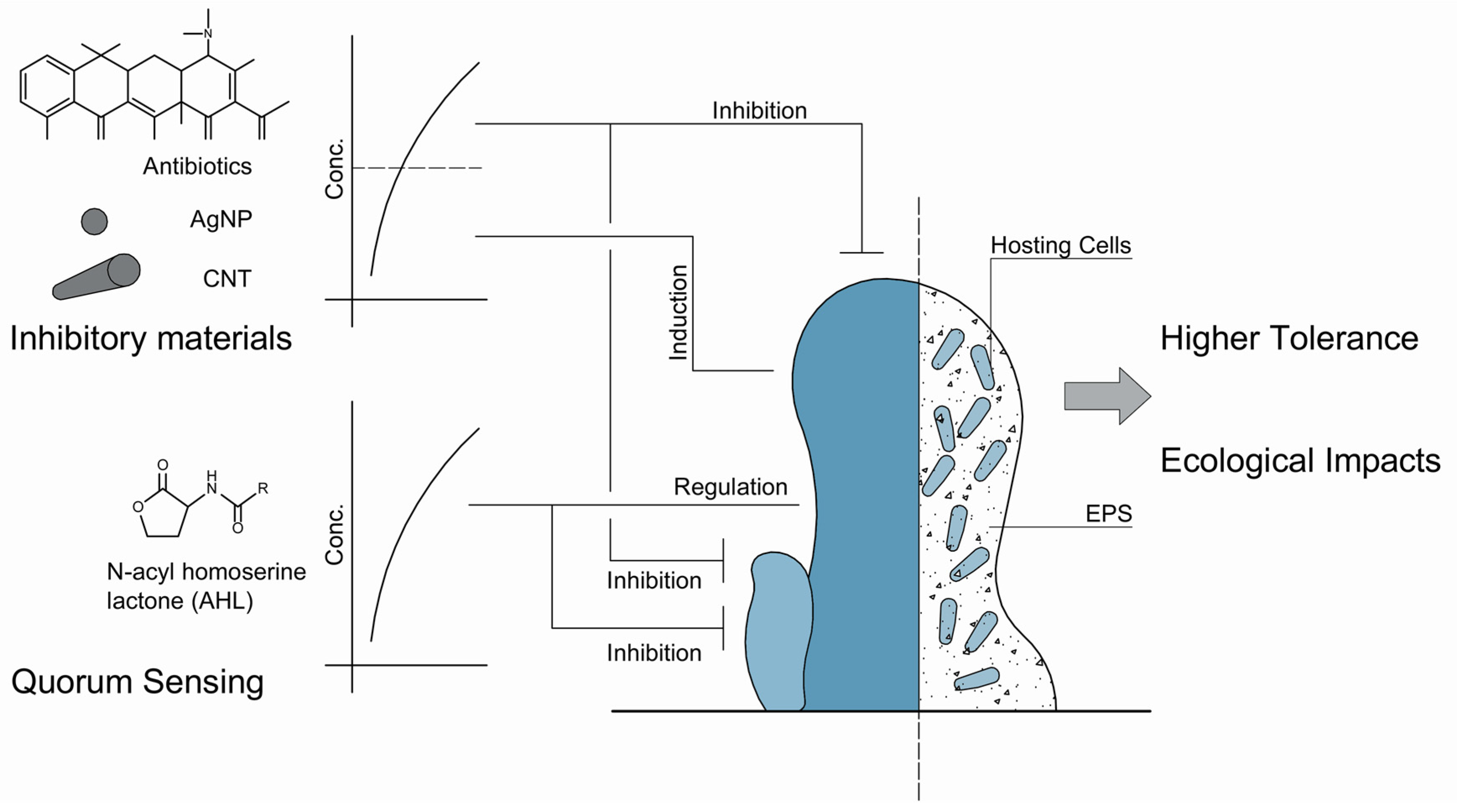
Biofilms are triggered by environmental stimuli. Recent models have considered the effects of stressors when mimicking the biofilms [19]. Generally, microbes in biofilms exhibit much higher tolerance toward environmental stressors than the planktonic cells. This is a critical issue, especially in medicine. As we mentioned previously, cells in biofilms are hundreds of times more tolerant to antimicrobial agents [12]. Using higher dosage of antibiotics is one solution but with extremely high cost and therapeutic risk [19]. Therefore, mathematical descriptions of the fate and transport of these antimicrobial stressors are necessary and of grand importance to manipulate biofilms (Figure 2).
To mimic the high tolerance of biofilms toward antibiotics, four mechanisms have been purposed. The first one is that the biofilm matrix serves as a barrier, which prevents the diffusion of antibiotic via the physical obstruction and chemical interaction, such as adsorption by EPS, reaction with biofilm materials, and catalytic degradation [48,52,66]. If the antibiotics penetrate the biofilm, the antimicrobial effects still becomes limited. These may contribute to the changed microenvironments, including pH, oxygen, and the activity of the dwelling microbes. The efficiency of antibiotics is much lower toward inactive cells [67]. Another mechanism is the adaptive responses of the microbes. When antibiotics, particularly sublethal levels of antibiotics, attack the cells after penetrating the first two barriers, microbes will produce protective enzymes or extra EPS to protect the cells [68,69]. At last, even the antibiotics work very well, some microorganisms survive. These special microorganisms are known as ‘persisters’ [70]. Interestingly, these persisters are genetically the same as the overall population of the strain, and the survival may be considered a switched-up phenotype. These mechanisms are not constrained in antibiotics but other inhibitory materials, such as nanoparticles and organic solvents. Xia et al., discovered that, in the presence of the widely used solvent, n-butanol, E. coli excreted more EPS and, specifically, the over-secreted EPS varied the hydrophobicity and chaotropicity of exposed cells to protect the cells [48]. Li et al., found that the photoautotrophic microbes—S. elongatus and M. aeruginosa—secrete more EPS under osmotic stress and these EPS serve as binding sites for the adsorption of external chemicals [71]. Based on these four hypothesized mechanisms, the transport and fate of external stressors have been modeled and the mathematical details can be found in the review by Klapper and Dockery [19].
3.1. The Inducing Effects of Sublethal Levels of Stressors on Biofilms
These inhibitory materials do not always work as killers. In natural ecosystems, particularly in water systems, various gradients of antibiotics have been detected because of the utility of antibiotics on human and livestock [72,73]. These detected antibiotics, although ubiquitous, are typically not of high enough concentrations to eliminate the encountered microbes. However, these sublethal or low-level concentrations of antibiotics exert even more profound impacts, as they allow microbes to grow and accelerate the evolution of antibiotic resistance genes [74]. On behalf of biofilm modeling, the effects of low-level antibiotics are fundamentally different from those above MIC, as the sublethal levels of antibiotics induce the formation of biofilms (Figure 2). This effect makes antibiotics a bifunctional chemical, and they also challenge the mathematical description and prediction of biofilm dynamics in natural ecosystems.
Hoffman et al. [75] discovered that the formation of biofilms by P. aeruginosa and E. coli would be induced in the presence of subinhibitory concentrations of aminoglycoside antibiotics. In P. aeruginosa, this inducing effect is attributed to the activity of the arr gene, the product of which is an inner membrane phosphodiesterase catalyzing the second messenger, cyclic di-guanosine monophosphate (c-di-GMP), and regulating the cell surface properties. Later, more research papers discussed the phenomenon that low-level antibiotics induce the formation of biofilms. For instance, Kaplan et al. [76] discovered that the low levels of β-lactam antibiotics—including methicillin, ampicillin, amoxicillin, and cloxacillin—would induce the formation of biofilms by various Staphylococcus aureus strains, a well-known human pathogen. The biomass in the induced biofilms was 10 times higher than those without the exposure of antibiotics. Bleich et al. [77] found that the thiazolyl peptide antibiotics promote the formation of biofilms by gram-positive bacteria B. subtilis and the inducing effects of this specific antibiotics are universal. Tan et al. [78] also discovered that the aminoglycoside antibiotics promote the biofilm formation by the photoautotrophic microorganism, S. elongatus, via upregulating the expression of EPS related genes. Recently, it has been commonly accepted that low-levels of antibiotics induce the formation of biofilms. Moreover, these inducing effects also work as competing mechanisms for antibiotic producing strains to survive the invasion and competition by other biofilm forming strains [79]. Therefore, how to bridge the novel discoveries associated with these antimicrobial agents is an urgent request for the modeling of biofilms.
These inducing phenomenon of low-level inhibitory materials are not constrained in antibiotics. These have also been identified in nanoparticles (Figure 2). For instance, the silver nanoparticles (AgNPs) showed promising antimicrobial activity on various microbial strains [80]. Recently, Yang et al. [81] discovered that the sublethal concentrations of AgNPs promoted the formation of biofilms by P. aeruginosa via upregulating a complex genetic networks associated with QS, EPS, as well as the antibiotic resistance genes. Upadhyayulaa and Gadhamshettyb [82] reported that the carbon nanotubes and their composites possess the capability of inducing biofilm formation as well at low level concentrations as well, while they were traditionally regarded as antimicrobial materials.
3.2. Ecological Impacts of Interfered Biofilms
In natural ecosystems, the biofilms are not only a ‘living style’ for better survival but also have ecological and biogeochemical roles. As discussed above, the low-level antibiotics would promote the biofilm formation by cyanobacterium, S. elongatus, and the cyanobacterial biofilm is the house for the biocalcification process, a fundamental bioprocess which contributes to local and global carbon cycles [78]. As reported, about 41.9% of the total carbon was attributed to carbonate, and a quite large proportion of carbonate is of biogenic origin [9]. It has been reported that the formation of the cyanobacterial biofilms would be promoted by subinhibitory concentrations of kanamycin with a higher amount of EPS excretion, both of which provide more functional groups for Ca2+ to bind. Meanwhile, the icfA gene, which encodes the carbonic anhydrase, was upregulated and subsequently increased the pH in the microenvironments in biofilms and facilitates the formation of calcium carbonate. As the dynamics of carbonate were essential in the biocalcification process, the interfered biofilms exert ecological and biogeochemical impacts through the combined effects of biofilm and icfA on biocalcification. As reported, enhanced biocalcification might increase atmospheric CO2 concentrations via the changed equilibrium [83]. Cyanobacteria are not the only microbes harboring ecological and biogeochemical roles in natural ecosystems. S. oneidensis and Geobacter sp. are also examples. These two strains mediate the redox balance through extracellular electron transfer and these processes typically need pili or EPS for cell–cell communication and transportation, which also depends on the formation of biofilms [84,85,86]. Therefore, development of mathematic models not only describes the dynamics of biofilms but also links the environmental functions which are of grand importance. Furthermore, it may also help to better evaluate the environmental and ecological risks of emerging pollutants.
4. Challenges in the New Era
In conclusion, the typical models for the simulation of biofilms are summarized in this review, and we also reviewed recent discoveries in the integration and determination of new variables during the process, such as QS, EPS, and inhibitory materials. Our objective is to bring more attention to link the new discoveries of biofilms—especially on behalf of natural water ecosystems—with current mathematical models to facilitate the understanding and prediction of biofilms, as well as the related environmental impacts. In our opinion, a better description of the bifunctional key components in biofilm matrix—like EPS, QS signals, antibiotics, as well as other emerging pollutants—should be established. More importantly, a mathematical link bridging the biofilm and its ecological functions is necessary.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 21476130 and No. 21676161). The authors would like to thank Chao Song for his valuable suggestions and comments on this paper.
Author Contributions
S.-G.W., Q.L., and P.-F.X. developed the idea and outlines of the review. Q.L. and P.-F.X. wrote the paper. Q.L., P.-F.X., and Z.-Y.T. revised the manuscript. P.-F.X. and Z.-Y.T. further proofread the main text.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Huerta, B.; Rodriguez-Mozaz, S.; Nannou, C.; Nakis, L.; Ruhi, A.; Acuña, V.; Sabater, S.; Barcelo, D. Determination of a broad spectrum of pharmaceuticals and endocrine disruptors in biofilm from a waste water treatment plant-impacted river. Sci. Total Environ. 2016, 540, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Hwang, G.; El-Din, M.G.; Liu, Y. Effect of reactor configuration and microbial characteristics on biofilm reactors for oil sands process-affected water treatment. Int. Biodeterior. Biodegrad. 2014, 89, 74–81. [Google Scholar] [CrossRef]
- Desrousseaux, C.; Sautou, V.; Descamps, S.; Traoré, O. Modification of the surfaces of medical devices to prevent microbial adhesion and biofilm formation. J. Hosp. Infect. 2013, 85, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Sandler, N.; Salmela, I.; Fallarero, A.; Rosling, A.; Khajeheian, M.; Kolakovic, R.; Genina, N.; Nyman, J.; Vuorela, P. Towards fabrication of 3D printed medical devices to prevent biofilm formation. Int. J. Pharm. 2014, 459, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.N.; Li, X.M.; Umar, A.; Fan, W.H.; Wang, Y. Insight into calcification of Synechocystis sp. enhanced by extracellular carbonic anhydrase. RSC Adv. 2016, 6, 29811–29817. [Google Scholar] [CrossRef]
- Dhami, N.K.; Reddy, M.S.; Mukherjee, A. Application of calcifying bacteria for remediation of stones and cultural heritages. Front. Microbiol. 2014, 5, 304. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Dittrich, M. Carbonate precipitation through microbial activities in natural environment, and their potential in biotechnology: A review. Front. Bioeng. Biotechnol. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-L.; Fang, W.; Wang, Y.-P.; Sheng, G.-P.; Zeng, R.J.; Li, W.-W.; Yu, H.-Q. Phosphorus removal in an enhanced biological phosphorus removal process: Roles of extracellular polymeric substances. Environ. Sci. Technol. 2013, 47, 11482–11489. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-W.; Zhang, H.-L.; Sheng, G.-P.; Yu, H.-Q. Roles of extracellular polymeric substances in enhanced biological phosphorus removal process. Water Res. 2015, 86, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Mah, T.-F.C.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Stewart, P.S.; Franklin, M.J. Physiological heterogeneity in biofilms. Nat. Rev. Microbiol. 2008, 6, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Schmeisser, C.; Stöckigt, C.; Raasch, C.; Wingender, J.; Timmis, K.; Wenderoth, D.; Flemming, H.-C.; Liesegang, H.; Schmitz, R.; Jaeger, K.-E. Metagenome survey of biofilms in drinking-water networks. Appl. Environ. Microbiol. 2003, 69, 7298–7309. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.E.; O’toole, G.A. Microbial biofilms: From ecology to molecular genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef] [PubMed]
- Monds, R.D.; O’Toole, G.A. The developmental model of microbial biofilms: Ten years of a paradigm up for review. Trends Microbiol. 2009, 17, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, S.R.; Bernstein, H.C.; Song, H.-S.; Fredrickson, J.K.; Fields, M.W.; Shou, W.; Johnson, D.R.; Beliaev, A.S. Engineering microbial consortia for controllable outputs. ISME J. 2016, 10, 2077–2084. [Google Scholar] [CrossRef] [PubMed]
- Klapper, I.; Dockery, J. Mathematical description of microbial biofilms. SIAM Rev. 2010, 52, 221–265. [Google Scholar] [CrossRef]
- Characklis, W. Bioengineering report: Fouling biofilm development: A process analysis. Biotechnol. Bioeng. 1981, 23, 1923–1960. [Google Scholar] [CrossRef]
- Horn, H.; Lackner, S. Modeling of biofilm systems: A review. In Productive Biofilms; Springer: Berlin, Germany, 2014; pp. 53–76. [Google Scholar]
- Mueller, N.C.; Nowack, B. Exposure modeling of engineered nanoparticles in the environment. Environ. Sci. Technol. 2008, 42, 4447–4453. [Google Scholar] [CrossRef] [PubMed]
- Marambio-Jones, C.; Hoek, E.M. A review of the antibacterial effects of silver nanomaterials and potential implications for human health and the environment. J. Nanopart. Res. 2010, 12, 1531–1551. [Google Scholar] [CrossRef]
- Scown, T.; Van Aerle, R.; Tyler, C. Review: Do engineered nanoparticles pose a significant threat to the aquatic environment? Crit. Rev. Toxicol. 2010, 40, 653–670. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.L. Antibiotics and antibiotic resistance genes in natural environments. Science 2008, 321, 365–367. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Martínez, J.-L.; Cantón, R. Antibiotics and antibiotic resistance in water environments. Curr. Opin. Biotechnol. 2008, 19, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Rendueles, O.; Kaplan, J.B.; Ghigo, J. Antibiofilm polysaccharides. Environ. Microbiol. 2013, 15, 334–346. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm formation as microbial development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.; Sun, X.; Taylor, A.A.; Walker, S.L.; Wang, Y.; Wang, S. D-Amino acids inhibit initial bacterial Adhesion: Thermodynamic evidence. Biotechnol. Bioeng. 2015, 112, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Solano, C.; Echeverz, M.; Lasa, I. Biofilm dispersion and quorum sensing. Curr. Opin. Microbiol. 2014, 18, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Rao, S.; Bansal, A.; Dang, S.; Gupta, S.; Gabrani, R. Pseudomonas aeruginosa biofilm: Potential therapeutic targets. Biologicals 2014, 42, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lazazzera, B.A. Lessons from DNA microarray analysis: The gene expression profile of biofilms. Curr. Opin. Microbiol. 2005, 8, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Ghigo, J.-M. Are there biofilm-specific physiological pathways beyond a reasonable doubt? Res. Microbiol. 2003, 154, 1–8. [Google Scholar] [CrossRef]
- Klausen, M.; Gjermansen, M.; Kreft, J.-U.; Tolker-Nielsen, T. Dynamics of development and dispersal in sessile microbial communities: Examples from Pseudomonas aeruginosa and Pseudomonas putida model biofilms. FEMS Microbiol. Lett. 2006, 261, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lardon, L.A.; Merkey, B.V.; Martins, S.; Dötsch, A.; Picioreanu, C.; Kreft, J.; Smets, B.F. iDynoMiCS: Next-generation individual-based modelling of biofilms. Environ. Microbiol. 2011, 13, 2416–2434. [Google Scholar] [CrossRef] [PubMed]
- Morgenroth, E.; Eberl, H.J.; Van Loosdrecht, M.C.M.; Noguera, D.R.; Pizarro, G.E.; Picioreanu, C.; Rittmann, B.E.; Schwarz, A.O.; Wanner, O. Comparing biofilm models for a single species biofilm system. Water Sci. Technol. 2004, 49, 145–154. [Google Scholar] [PubMed]
- Horn, H.; Hempel, D.C. Growth and decay in an auto-/heterotrophic biofilm. Water Res. 1997, 31, 2243–2252. [Google Scholar] [CrossRef]
- Cogan, N.; Keener, J.P. The role of the biofilm matrix in structural development. Math. Med. Biol. 2004, 21, 147–166. [Google Scholar] [CrossRef] [PubMed]
- Duddu, R.; Bordas, S.; Chopp, D.; Moran, B. A combined extended finite element and level set method for biofilm growth. Int. J. Numer. Methods Eng. 2008, 74, 848–870. [Google Scholar] [CrossRef]
- Wanner, O.; Reichert, P. Mathematical modeling of mixed-culture biofilms. Biotechnol. Bioeng. 1996, 49, 172–184. [Google Scholar] [CrossRef]
- Picioreanu, C.; Van Loosdrecht, M.C.; Heijnen, J.J. Mathematical modeling of biofilm structure with a hybrid differential-discrete cellular automaton approach. Biotechnol. Bioeng. 1998, 58, 101–116. [Google Scholar] [CrossRef]
- Kreft, J.-U.; Picioreanu, C.; Wimpenny, J.W.; van Loosdrecht, M.C. Individual-based modelling of biofilms. Microbiology 2001, 147, 2897–2912. [Google Scholar] [CrossRef] [PubMed]
- Laspidou, C.; Kungolos, A.; Samaras, P. Cellular-automata and individual-based approaches for the modeling of biofilm structures: Pros and cons. Desalination 2010, 250, 390–394. [Google Scholar] [CrossRef]
- Skoneczny, S. Cellular automata-based modelling and simulation of biofilm structure on multi-core computers. Water Sci. Technol. 2015, 72, 2071–2081. [Google Scholar] [CrossRef] [PubMed]
- Hellweger, F.L.; Bucci, V. A bunch of tiny individuals—Individual-based modeling for microbes. Ecol. Model. 2009, 220, 8–22. [Google Scholar] [CrossRef]
- Kreft, J.-U.; Booth, G.; Wimpenny, J.W. BacSim, a simulator for individual-based modelling of bacterial colony growth. Microbiology 1998, 144, 3275–3287. [Google Scholar] [CrossRef] [PubMed]
- Sheng, G.-P.; Yu, H.-Q.; Li, X.-Y. Extracellular polymeric substances (EPS) of microbial aggregates in biological wastewater treatment systems: A review. Biotechnol. Adv. 2010, 28, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.-F.; Li, Q.; Tan, L.-R.; Sun, X.-F.; Song, C.; Wang, S.-G. Extracellular polymeric substances protect Escherichia coli from organic solvents. RSC Adv. 2016, 6, 59438–59444. [Google Scholar] [CrossRef]
- Horn, H.; Neu, T.; Wulkow, M. Modelling the structure and function of extracellular polymeric substances in biofilms with new numerical techniques. Water Sci. Technol. 2001, 43, 121–127. [Google Scholar] [PubMed]
- Vu, B.; Chen, M.; Crawford, R.J.; Ivanova, E.P. Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 2009, 14, 2535–2554. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Hu, Y.; Liu, Y.; Zhang, J.; Ulstrup, J.; Molin, S. Distinct roles of extracellular polymeric substances in Pseudomonas aeruginosa biofilm development. Environ. Microbiol. 2011, 13, 1705–1717. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Sun, X.-F.; Xing, S.-F.; Xia, P.-F.; Shi, Y.-J.; Wang, S.-G. Characterization of the interactions between tetracycline antibiotics and microbial extracellular polymeric substances with spectroscopic approaches. Environ. Sci. Pollut. Res. 2014, 21, 1786–1795. [Google Scholar] [CrossRef] [PubMed]
- Valle, J.; Da Re, S.; Henry, N.; Fontaine, T.; Balestrino, D.; Latour-Lambert, P.; Ghigo, J.-M. Broad-spectrum biofilm inhibition by a secreted bacterial polysaccharide. Proc. Natl. Acad. Sci. USA 2006, 103, 12558–12563. [Google Scholar] [CrossRef] [PubMed]
- Bendaoud, M.; Vinogradov, E.; Balashova, N.V.; Kadouri, D.E.; Kachlany, S.C.; Kaplan, J.B. Broad-spectrum biofilm inhibition by Kingella kingae exopolysaccharide. J. Bacteriol. 2011, 193, 3879–3886. [Google Scholar] [CrossRef] [PubMed]
- Colvin, K.M.; Gordon, V.D.; Murakami, K.; Borlee, B.R.; Wozniak, D.J.; Wong, G.C.; Parsek, M.R. The pel polysaccharide can serve a structural and protective role in the biofilm matrix of Pseudomonas aeruginosa. PLoS Pathog. 2011, 7, e1001264. [Google Scholar] [CrossRef] [PubMed]
- Diggle, S.P.; Stacey, R.E.; Dodd, C.; Cámara, M.; Williams, P.; Winzer, K. The galactophilic lectin, LecA, contributes to biofilm development in Pseudomonas aeruginosa. Environ. Microbiol. 2006, 8, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Tielker, D.; Hacker, S.; Loris, R.; Strathmann, M.; Wingender, J.; Wilhelm, S.; Rosenau, F.; Jaeger, K.-E. Pseudomonas aeruginosa lectin LecB is located in the outer membrane and is involved in biofilm formation. Microbiology 2005, 151, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; Maestre-Reyna, M.; Lee, G.; Gerard, H.; Wang, A.H.-J.; Watnick, P.I. In situ proteolysis of the Vibrio cholerae matrix protein RbmA promotes biofilm recruitment. Proc. Natl. Acad. Sci. USA 2015, 112, 10491–10496. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed]
- Chopp, D.L.; Kirisits, M.J.; Moran, B.; Parsek, M.R. A mathematical model of quorum sensing in a growing bacterial biofilm. J. Ind. Microbiol. Biotechnol. 2002, 29, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Chopp, D.L.; Kirisits, M.J.; Moran, B.; Parsek, M.R. The dependence of quorum sensing on the depth of a growing biofilm. Bull. Math. Biol. 2003, 65, 1053–1079. [Google Scholar] [CrossRef]
- Ward, J.P.; King, J.R.; Koerber, A.J.; Croft, J.M.; Sockett, R.E.; Williams, P. Early development and quorum sensing in bacterial biofilms. J. Math. Biol. 2003, 47, 23–55. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.P.; King, J.R.; Koerber, A.; Williams, P.; Croft, J.; Sockett, R. Mathematical modelling of quorum sensing in bacteria. Math. Med. Biol. 2001, 18, 263–292. [Google Scholar] [CrossRef]
- Ward, J.P.; King, J.R. Thin-film modelling of biofilm growth and quorum sensing. J. Eng. Math. 2012, 73, 71–92. [Google Scholar] [CrossRef]
- Papenfort, K.; Silpe, J.E.; Schramma, K.R.; Cong, J.-P.; Seyedsayamdost, M.R.; Bassler, B.L. A Vibrio cholerae autoinducer-receptor pair that controls biofilm formation. Nat. Chem. Biol. 2017, 13, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Xing, S.; Wang, X.; Wang, S. Changes of the reactor performance and the properties of granular sludge under tetracycline (TC) stress. Bioresour. Technol. 2013, 139, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Walters, M.C.; Roe, F.; Bugnicourt, A.; Franklin, M.J.; Stewart, P.S. Contributions of antibiotic penetration, oxygen limitation, and low metabolic activity to tolerance of Pseudomonas aeruginosa biofilms to ciprofloxacin and tobramycin. Antimicrob. Agents Chemother. 2003, 47, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Bagge, N.; Schuster, M.; Hentzer, M.; Ciofu, O.; Givskov, M.; Greenberg, E.P.; Høiby, N. Pseudomonas aeruginosa biofilms exposed to imipenem exhibit changes in global gene expression and β-lactamase and alginate production. Antimicrob. Agents Chemother. 2004, 48, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Elkins, J.G.; Hassett, D.J.; Stewart, P.S.; Schweizer, H.P.; McDermott, T.R. Protective role of catalase in Pseudomonas aeruginosa biofilm resistance to hydrogen peroxide. Appl. Environ. Microbiol. 1999, 65, 4594–4600. [Google Scholar] [PubMed]
- Lewis, K. Persister cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Xia, P.-F.; Tan, L.-R.; Wang, Y.; Sun, X.-F.; Wang, S.-G. Inducible microbial osmotic responses enable enhanced biosorption capability of cyanobacteria. Biochem. Eng. J. 2017, 120, 113–117. [Google Scholar] [CrossRef]
- Carvalho, I.T.; Santos, L. Antibiotics in the aquatic environments: A review of the European scenario. Environ. Int. 2016, 94, 736–757. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Courvalin, P.; Dantas, G.; Davies, J.; Eisenstein, B.; Huovinen, P.; Jacoby, G.A.; Kishony, R.; Kreiswirth, B.N.; Kutter, E. Tackling antibiotic resistance. Nat. Rev. Microbiol. 2011, 9, 894–896. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.I.; Hughes, D. Microbiological effects of sublethal levels of antibiotics. Nat. Rev. Microbiol. 2014, 12, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.R.; D’argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside antibiotics induce bacterial biofilm formation. Nature 2005, 436, 1171–1175. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.B.; Izano, E.A.; Gopal, P.; Karwacki, M.T.; Kim, S.; Bose, J.L.; Bayles, K.W.; Horswill, A.R. Low levels of β-lactam antibiotics induce extracellular DNA release and biofilm formation in Staphylococcus aureus. MBio 2012, 3, e00198-12. [Google Scholar] [CrossRef] [PubMed]
- Bleich, R.; Watrous, J.D.; Dorrestein, P.C.; Bowers, A.A.; Shank, E.A. Thiopeptide antibiotics stimulate biofilm formation in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2015, 112, 3086–3091. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.-R.; Xia, P.-F.; Sun, X.-F.; Guo, N.; Song, C.; Li, Q.; Wang, S.-G. Ecological insights into low-level antibiotics interfered biofilms of Synechococcus elongatus. RSC Adv. 2016, 6, 78132–78135. [Google Scholar] [CrossRef]
- Oliveira, N.M.; Martinez-Garcia, E.; Xavier, J.; Durham, W.M.; Kolter, R.; Kim, W.; Foster, K.R. Biofilm formation as a response to ecological competition. PLoS Biol. 2015, 13, e1002191. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.-F.; Qin, J.; Xia, P.-F.; Guo, B.-B.; Yang, C.-M.; Song, C.; Wang, S.-G. Graphene oxide–silver nanoparticle membrane for biofouling control and water purification. Chem. Eng. J. 2015, 281, 53–59. [Google Scholar] [CrossRef]
- Yang, Y.; Alvarez, P.J. Sublethal concentrations of silver nanoparticles stimulate biofilm development. Environ. Sci. Technol. Lett. 2015, 2, 221–226. [Google Scholar] [CrossRef]
- Upadhyayula, V.K.; Gadhamshetty, V. Appreciating the role of carbon nanotube composites in preventing biofouling and promoting biofilms on material surfaces in environmental engineering: A review. Biotechnol. Adv. 2010, 28, 802–816. [Google Scholar] [CrossRef] [PubMed]
- Ridgwell, A.; Zeebe, R.E. The role of the global carbonate cycle in the regulation and evolution of the Earth system. Earth Planet. Sci. Lett. 2005, 234, 299–315. [Google Scholar] [CrossRef]
- Shi, L.; Richardson, D.J.; Wang, Z.; Kerisit, S.N.; Rosso, K.M.; Zachara, J.M.; Fredrickson, J.K. The roles of outer membrane cytochromes of Shewanella and Geobacter in extracellular electron transfer. Environ. Microbiol. Rep. 2009, 1, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Bond, D.R.; Strycharz-Glaven, S.M.; Tender, L.M.; Torres, C.I. On electron transport through Geobacter biofilms. ChemSusChem 2012, 5, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Pirbadian, S.; Barchinger, S.E.; Leung, K.M.; Byun, H.S.; Jangir, Y.; Bouhenni, R.A.; Reed, S.B.; Romine, M.F.; Saffarini, D.A.; Shi, L. Shewanella oneidensis MR-1 nanowires are outer membrane and periplasmic extensions of the extracellular electron transport components. Proc. Natl. Acad. Sci. USA 2014, 111, 12883–12888. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schemes of biofilm evolution (A); the simulation of substrate consumption (B); and quorum sensing (C) via current 1D model; and the illustration of the multidimensional cellular automata model (D).
Figure 1.
Schemes of biofilm evolution (A); the simulation of substrate consumption (B); and quorum sensing (C) via current 1D model; and the illustration of the multidimensional cellular automata model (D).

Figure 2.
Biofilms in the presence of signals and stimuli in natural water systems, and the regulation and inhibition of these factors on biofilm matrix.
Figure 2.
Biofilms in the presence of signals and stimuli in natural water systems, and the regulation and inhibition of these factors on biofilm matrix.

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, Q.; Xia, P.-F.; Tao, Z.-Y.; Wang, S.-G. Modeling Biofilms in Water Systems with New Variables: A Review. Water 2017, 9, 462. https://doi.org/10.3390/w9070462
AMA Style
Li Q, Xia P-F, Tao Z-Y, Wang S-G. Modeling Biofilms in Water Systems with New Variables: A Review. Water. 2017; 9(7):462. https://doi.org/10.3390/w9070462
Chicago/Turabian StyleLi, Qian, Peng-Fei Xia, Ze-Yu Tao, and Shu-Guang Wang. 2017. "Modeling Biofilms in Water Systems with New Variables: A Review" Water 9, no. 7: 462. https://doi.org/10.3390/w9070462
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.