
Protective Effect of Natural Antioxidant Compounds on Methimazole Induced Oxidative Stress in a Feline Kidney Epithelial Cell Line (CRFK)

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Cell Culture and Treatment
2.3. Cell Viability Assay
2.4. Reactive Oxygen Species (ROS) Production Assay
2.5. Determination of Reduced Glutathione (GSH) Content
2.6. Quantitative RT-PCR (q-PCR) Assays
2.7. Statistical Analysis
3. Results
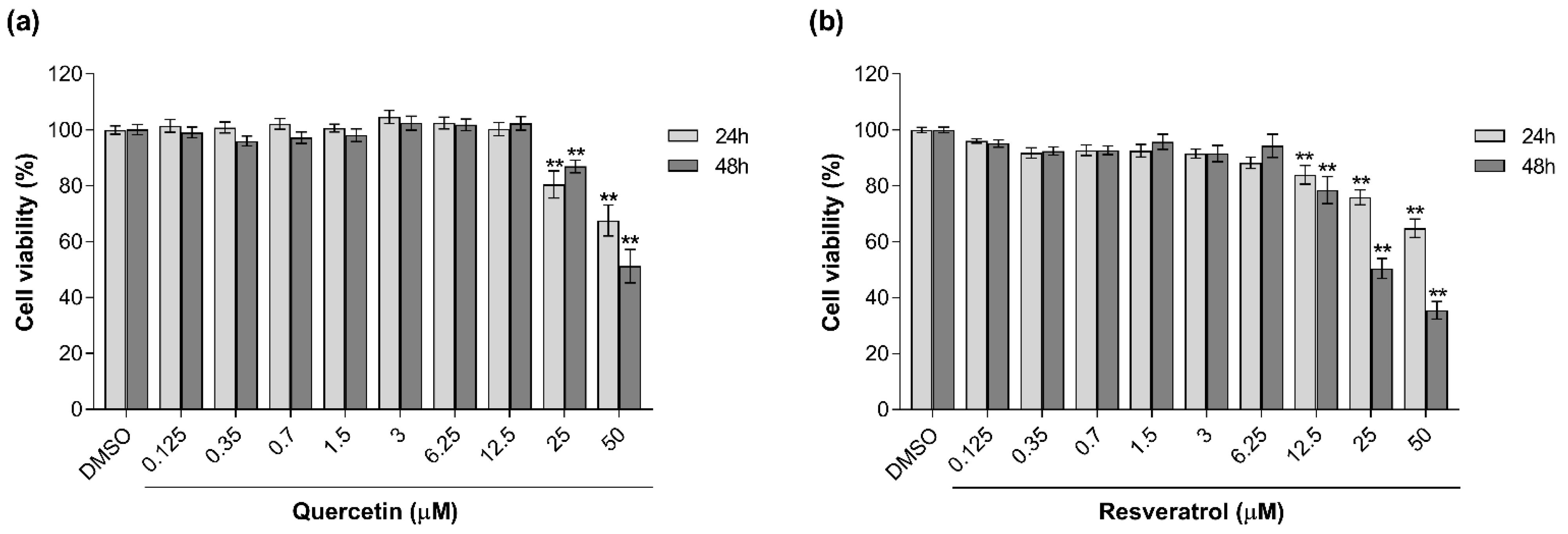
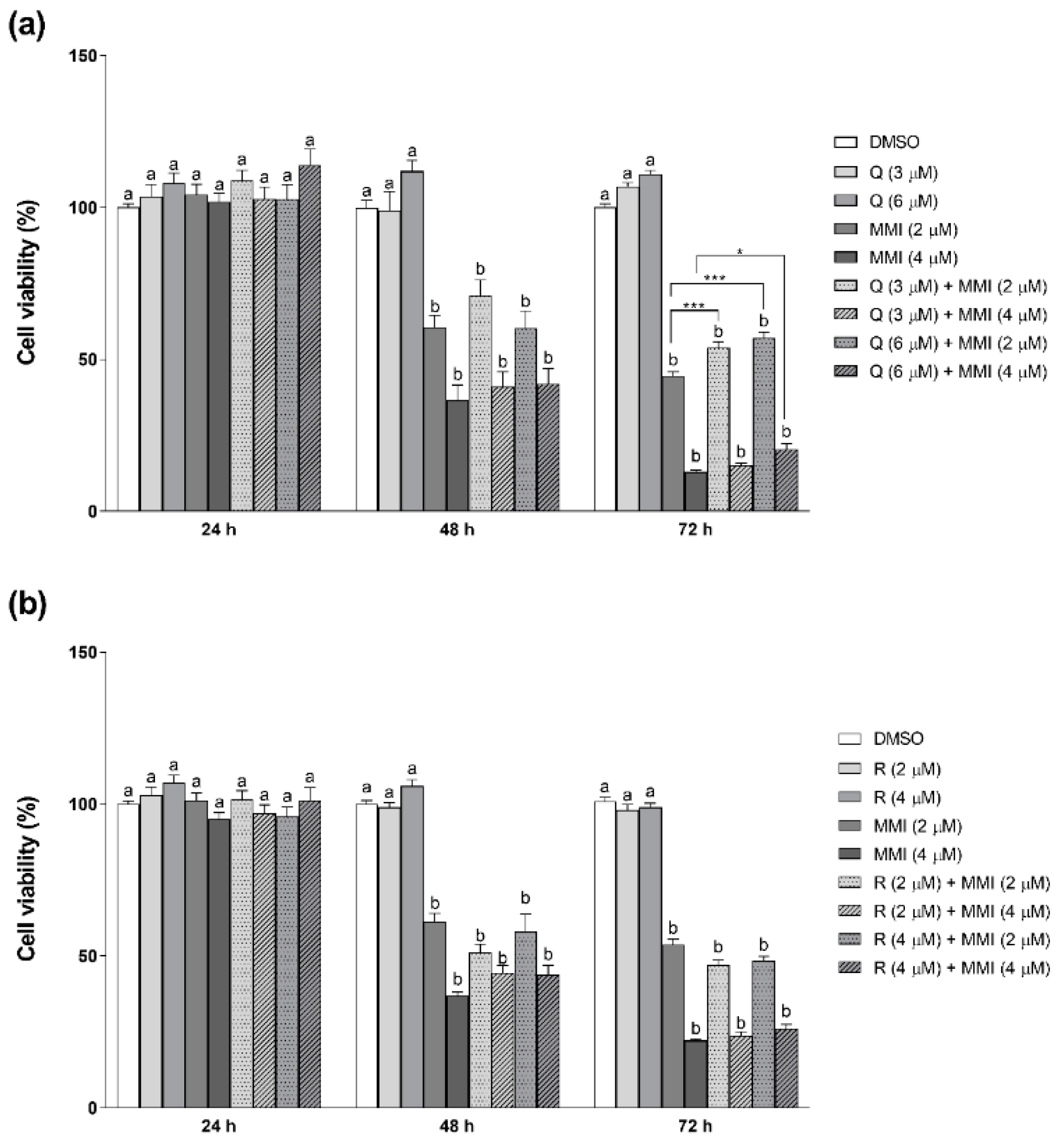
3.1. Effects of Natural Antioxidants on MMI-Induced Cytotoxicity
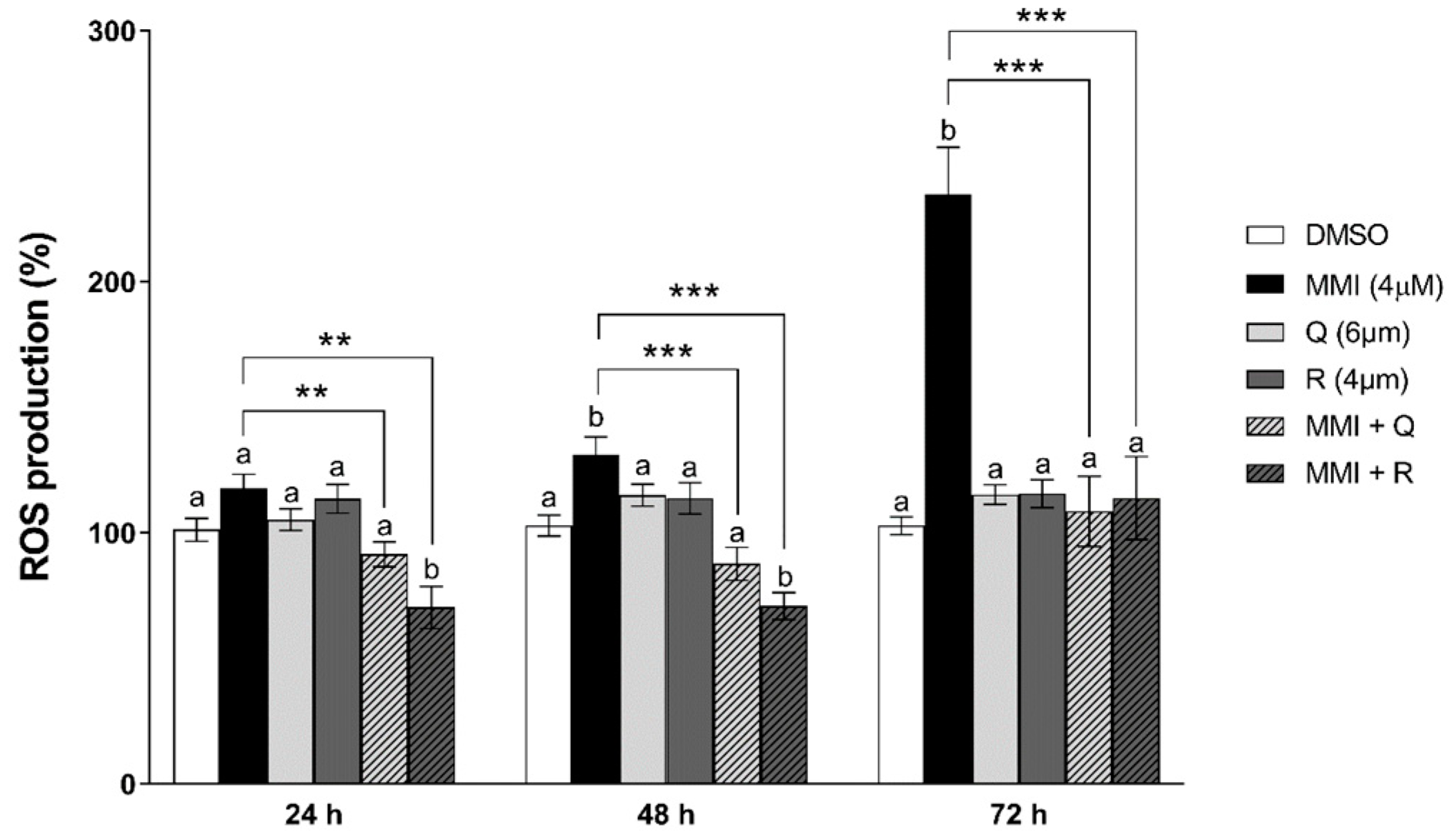
3.2. Effects of Natural Antioxidants on MMI-Induced ROS Production
3.3. Modulation of Reduced GSH Content by MMI and Antioxidants
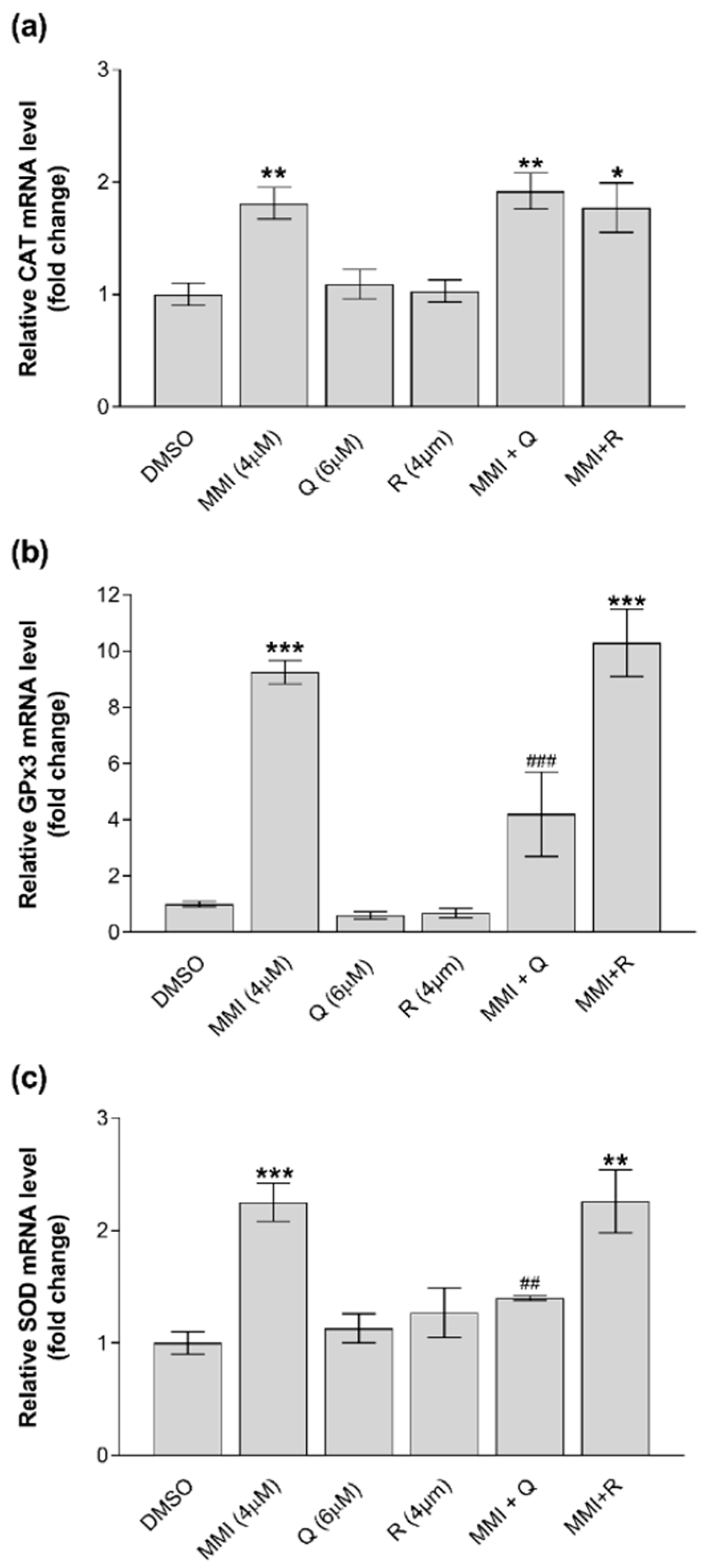
3.4. Modulation of Gene Expression by MMI and Antioxidants
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peterson, M. Hyperthyroidism in Cats. J. Feline Med. Surg. 2012, 14, 804–818. [Google Scholar] [CrossRef]
- Grauer, G.F.; Schermerhorn, T.; Armbrust, L.; Vaske, H. Diagnosis and management of feline hyperthyroidism: Current perspectives. Vet. Med. Res. Rep. 2014, 5, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Syme, H.M. Cardiovascular and Renal Manifestations of Hyperthyroidism. Vet. Clin. North Am. Small Anim. Pr. 2007, 37, 723–743. [Google Scholar] [CrossRef]
- Daminet, S.; Kooistra, H.S.; Fracassi, F.; Graham, P.A.; Hibbert, A.; Lloret, A.; Mooney, C.T.; Neiger, R.; Rosenberg, D.; Syme, H.M.; et al. Best practice for the pharmacological management of hyperthyroid cats with antithyroid drugs. J. Small Anim. Pr. 2013, 55, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Vaske, H.H.; Schermerhorn, T.; Grauer, G.F. Effects of feline hyperthyroidism on kidney function: A review. J. Feline Med. Surg. 2015, 18, 55–59. [Google Scholar] [CrossRef]
- Trepanier, L.A. Pharmacologic Management of Feline Hyperthyroidism. Vet. Clin. North Am. Small Anim. Pr. 2007, 37, 775–788. [Google Scholar] [CrossRef] [PubMed]
- Carney, H.C.; Ward, C.R.; Bailey, S.J.; Bruyette, D.; Dennis, S.; Ferguson, D.; Hinc, A.; Rucinsky, A.R. 2016 AAFP Guidelines for the Management of Feline Hyperthyroidism. J. Feline Med. Surg. 2016, 18, 400–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burch, H.B.; Cooper, D.S. Antithyroid drug therapy: 70 years later. Eur. J. Endocrinol. 2018, 179, R261–R274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shell, A.; Sullivan, J.W. Acute Kidney Injury Following Methimazole Initiation: A Case Report. J. Pharm. Pract. 2018, 33, 99–101. [Google Scholar] [CrossRef] [PubMed]
- Becker, T.J.; Graves, T.K.; Kruger, J.M.; Braselton, W.E.; Nachreiner, R.F. Effects of methimazole on renal function in cats with hyperthyroidism. J. Am. Anim. Hosp. Assoc. 2000, 36, 215–223. [Google Scholar] [CrossRef]
- Heidari, R.; Niknahad, H.; Jamshidzadeh, A.; Abdoli, N. Factors affecting drug-induced liver injury: Antithyroid drugs as instances. Clin. Mol. Hepatol. 2014, 20, 237–248. [Google Scholar] [CrossRef]
- Heidari, R.; Niknahad, H.; Jamshidzadeh, A.; Eghbal, M.A.; Abdoli, N. An Overview on the Proposed Mechanisms of Antithyroid Drugs-Induced Liver Injury. Adv. Pharm. Bull. 2015, 5, 1–11. [Google Scholar] [CrossRef]
- Kocak, M.; Akarsu, E.; Korkmaz, H.; Taysi, S. The Effect of Antithyroid Drugs on Osteopontin and Oxidative Stress in Graves’ Disease. Acta Endocrinol. 2019, 15, 221–224. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Lanzolla, G.; Marcocci, C.; Marinò, M. Oxidative Stress in Graves Disease and Graves Orbitopathy. Eur. Thyroid. J. 2020, 9, 40–50. [Google Scholar] [CrossRef]
- Mancini, A.; Di Segni, C.; Raimondo, S.; Olivieri, G.; Silvestrini, A.; Meucci, E.; Curro’, D. Thyroid Hormones, Oxidative Stress, and Inflammation. Mediat. Inflamm. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ademoğlu, E.; Özbey, N.; Erbil, Y.; Tanrikulu, S.; Barbaros, U.; Yanik, B.T.; Bozbora, A.; Özarmağan, S. Determination of oxidative stress in thyroid tissue and plasma of patients with Graves’ disease. Eur. J. Intern. Med. 2006, 17, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Komosinska-Vassev, K.; Olczyk, K.; Kucharz, E.J.; Marcisz, C.; Winsz-Szczotka, K.; Kotulska, A. Free radical activity and antioxidant defense mechanisms in patients with hyperthyroidism due to Graves’ disease during therapy. Clin. Chim. Acta 2000, 300, 107–117. [Google Scholar] [CrossRef]
- Candellone, A.; Gianella, P.; Ceccarelli, L.; Raviri, G.; Badino, P.; Roncone, S.; Kooistra, H.S.; Meineri, G. Redox unbalance in the hyperthyroid cat: A comparison with healthy and non-thyroidal diseased cats. BMC Vet. Res. 2019, 15, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candellone, A.; Badino, P.; Gianella, P.; Girolami, F.; Raviri, G.; Saettone, V.; Meineri, G. Evaluation of Antioxidant Supplementation on Redox Unbalance in Hyperthyroid Cats Treated with Methimazole: A Blinded Randomized Controlled Trial. Antioxidants 2019, 9, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I. Antioxidant activity of food constituents: An overview. Arch. Toxicol. 2012, 86, 345–391. [Google Scholar] [CrossRef] [PubMed]
- Li, A.-N.; Li, S.; Zhang, Y.-J.; Xu, X.-R.; Chen, Y.-M.; Li, H.-B. Resources and Biological Activities of Natural Polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-J.; Gan, R.-Y.; Li, S.; Zhou, Y.; Li, A.-N.; Xu, D.-P.; Li, H.-B. Antioxidant Phytochemicals for the Prevention and Treatment of Chronic Diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef]
- Crandell, R.A.; Fabricant, C.G.; Nelson-Rees, W.A. Development, characterization, and viral susceptibility of a feline (Felis catus) renal cell line (CRFK). Vitr. Cell. Dev. Biol. Anim. 1973, 9, 176–185. [Google Scholar] [CrossRef]
- Lawson, J.; Syme, H.; Wheeler-Jones, C.; Elliott, J. Characterisation of Crandell-Rees Feline Kidney (CRFK) cells as mesenchymal in phenotype. Res. Vet. Sci. 2019, 127, 99–102. [Google Scholar] [CrossRef]
- Koike, M.; Yutoku, Y.; Koike, A. Inhibition of Crandell-Rees Feline Kidney cell proliferation by X-ray-induced senescence. J. Vet. Med Sci. 2021, 83, 798–804. [Google Scholar] [CrossRef]
- Van Beusekom, C.D.; Zimmering, T.M. Profibrotic effects of angiotensin II and transforming growth factor beta on feline kidney epithelial cells. J. Feline Med. Surg. 2018, 21, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Trepanier, L.; Peterson, M.; Aucoin, D. Pharmacokinetics of methimazole in normal cats and cats with hyperthyroidism. Res. Vet. Sci. 1991, 50, 69–74. [Google Scholar] [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Ugazio, G.; Burdino, E.; Dacasto, M.; Bosio, A.; Klooster, G.V.; Nebbia, C. Induction of hepatic drug metabolizing enzymes and interaction with carbon tetrachloride in rats after a single oral exposure to atrazine. Toxicol. Lett. 1993, 69, 279–288. [Google Scholar] [CrossRef]
- Penning, L.C.; Vrieling, H.E.; Brinkhof, B.; Riemers, F.M.; Rothuizen, J.; Rutteman, G.R.; Hazewinkel, H.A. A validation of 10 feline reference genes for gene expression measurements in snap-frozen tissues. Vet. Immunol. Immunopathol. 2007, 120, 212–222. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cano-Europa, E.; Blas-Valdivia, V.; Franco-Colin, M.; Gallardo-Casas, C.A.; Ortiz-Butrón, R. Methimazole-induced hypothyroidism causes cellular damage in the spleen, heart, liver, lung and kidney. Acta Histochem. 2011, 113, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Cano-Europa, E.; Blas-Valdivia, V.; Lopez-Galindo, G.E.; Franco-Colin, M.; Pineda-Reynoso, M.; Hernandez-Garcia, A.; Ortiz-Butron, R. Methimazole-induced hypothyroidism causes alteration of the REDOX environment, oxidative stress, and hepatic damage; events not caused by hypothyroidism itself. Ann. Hepatol. 2010, 9, 80–88. [Google Scholar] [CrossRef]
- Heidari, R.; Babaei, H.; Eghbal, M.A. Cytoprotective effects of organosulfur compounds against methimazole induced toxicity in isolated rat hepatocytes. Adv. Pharm. Bull. 2013, 3, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, M.E.; Broome, M.R. Hyperthyroid cats on long-term medical treatment show a progressive increase in the prevalence of large thyroid tumors, intrathoracic thyroid masses and suspected thyroid carcinoma. J. Vet. Intern. Med. 2012, 26, 1523. [Google Scholar]
- Woods, J.S.; Ellis, M.E. Up-regulation of glutathione synthesis in rat kidney by methyl mercury: Relationship to mercury-induced oxidative stress. Biochem. Pharmacol. 1995, 50, 1719–1724. [Google Scholar] [CrossRef]
- Olson, G.E.; Whitin, J.C.; Hill, K.E.; Winfrey, V.P.; Motley, A.K.; Austin, L.M.; Deal, J.; Cohen, H.J.; Burk, R.F. Extracellular glutathione peroxidase (Gpx3) binds specifically to basement membranes of mouse renal cortex tubule cells. Am. J. Physiol. Physiol. 2010, 298, F1244–F1253. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhao, Q.; Zhang, K.; Sun, W.; Jia, X.; Yang, Y.; Yin, J.; Tang, C.; Zhang, J. Se deficiency induces renal pathological changes by regulating selenoprotein expression, disrupting redox balance, and activating inflammation. Metallomics 2020, 12, 1576–1584. [Google Scholar] [CrossRef]
- Zhang, H.; Luan, L.; Bi, M.; Zhao, L.; Yuan, L.; Feng, J.; Liu, R. In vivo and in vitro studies on inactivation of selenium containing protein- glutathione peroxidase 3 in mice nephrocytes caused by lead. Ecotoxicol. Environ. Saf. 2020, 203, 111008. [Google Scholar] [CrossRef] [PubMed]
- Santi, A.; Baldissareli, J.; Murussi, C.R.; Dias, G.R.M.; de Menezes, C.C.; Zanini, D.; Abdalla, F.H.; Thomé, G.R.; Martins, C.C.; Schetinger, M.R.C.; et al. Effects of Quercetin on Oxidative Stress Biomarkers in Methimazole—Induced Hypothyroid Rats. Exp. Clin. Endocrinol. Diabetes 2014, 122, 533–539. [Google Scholar] [CrossRef]
- Chi, K.-K.; Zhang, W.-H.; Chen, Z.; Cui, Y.; He, W.; Wang, S.-G.; Zhang, C.; Chen, J.; Wang, G.-C. Comparison of quercetin and resveratrol in the prevention of injury due to testicular torsion/detorsion in rats. Asian J. Androl. 2015, 18, 908–912. [Google Scholar] [CrossRef]
- Chuang, C.-C.; Martinez, K.; Xie, G.; Kennedy, A.; Bumrungpert, A.; Overman, A.; Jia, W.; McIntosh, M.K. Quercetin is equally or more effective than resveratrol in attenuating tumor necrosis factor-α–mediated inflammation and insulin resistance in primary human adipocytes. Am. J. Clin. Nutr. 2010, 92, 1511–1521. [Google Scholar] [CrossRef] [PubMed]
- Chekalina, N.I.; Kazakov, Y.M.; Mamontova, T.V.; Vesnina, L.E.; Kaidashev, I.P. Resveratrol more effectively than quercetin reduces endothelium degeneration and level of necrosis factor α in patients with coronary artery disease. Wiad. Lek. 2016, 69, 475–479. [Google Scholar] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free. Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Jackson, S.J.; Singletary, K.W.; Murphy, L.L.; Venema, R.C.; Young, A.J. Phytonutrients Differentially Stimulate NAD(P)H:Quinone Oxidoreductase, Inhibit Proliferation, and Trigger Mitotic Catastrophe in Hepa1c1c7 Cells. J. Med. Food 2016, 19, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. | Sequence (5′–>3′) | Amplicon Size (bp) |
|---|---|---|---|
| CAT | XM_003993157.4 | FW: ACGCCTGTGTGAGAACATTG REV: TCACTGAAGTTCTTGACCG | 82 |
| GPx1 | XM_004001361.5 | FW: GGGCATCAGGAGAACGCTAA REV: CGCCATTTACCTCGCACTTC | 115 |
| GPx3 | XM_003981387.5 | FW: AACGGGGAGAAAGAGCAGAA REV: TTCCCAGAAGAGGCGGTTAG | 96 |
| GPx4 | XM_023242756.1 | FW: TCACCAAGTTCCTCATTGACA REV: TAGAGGTAGCAGGGCAAGTC | 100 |
| GSTA2 | XM_011282429.3 | FW: ATGTGGAAGAGCTTGACCCC REV: CGGGAGGGAGATTGCTGATT | 84 |
| GSTM3 | XM_003990413.5 | FW: CCGTTTTGAGGCTTTGGAGA REV: TTGGGCCATCTTGTTGTTGA | 85 |
| GSTP1 | XM_023240130.1 | FW: TCGCAGCAAATACATCACCC REV: GTCTCGAAAGGCTTCAGGTG | 96 |
| SOD1 | XM_006935922.3 | FW: CATCATTGGCCGCACGAT REV: ATGACACCACAAGCCAAACG | 114 |
| YWHAZ | XM_006943327.4 | FW: GAAGAGTCCTACAAAGACAGCACGC REV: AATTTTCCCCTCCTTCTCCTGC | 115 |
| GSH Content (µg GSH/mg Protein) | |||||
|---|---|---|---|---|---|
| 4 h | 8 h | 24 h | 48 h | 72 h | |
| DMSO | 129.3 ± 25.2 | 106.2 ± 9.8 | 122.6 ± 19.3 | 85.0 ± 14.1 | 73.88 ± 12.0 |
| MMI | 118.9 ± 15.4 | 145.3 ± 10.7 | 176.1 ± 20.1 | 151.6 ± 10.5 * | 154.4 ± 15.5 * |
| Q | 117.4 ± 23.9 | 215.0 ± 26.4 * | 226.2 ± 16.8 * | 119.8 ± 13.6 | 100.7 ± 10.1 |
| R | 109.1 ± 19.1 | 130.1 ± 10.7 | 142.84 ± 18.6 | 99.1 ± 12.8 | 94.8 ± 17.6 |
| MMI + Q | 120.8 ± 24.4 | 220.8 ± 5.4 *# | 268.6 ± 4.9 *# | 208.5 ± 23.5 * | 224.8 ± 24.7 *# |
| MMI + R | 117.5 ± 16.6 | 200.1 ± 19.2 * | 219.7 ± 7.0 * | 192.3 ± 13.2 * | 197.2 ± 12.0 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Girolami, F.; Candellone, A.; Jarriyawattanachaikul, W.; Meineri, G.; Nebbia, C.; Badino, P. Protective Effect of Natural Antioxidant Compounds on Methimazole Induced Oxidative Stress in a Feline Kidney Epithelial Cell Line (CRFK). Vet. Sci. 2021, 8, 220. https://doi.org/10.3390/vetsci8100220
Girolami F, Candellone A, Jarriyawattanachaikul W, Meineri G, Nebbia C, Badino P. Protective Effect of Natural Antioxidant Compounds on Methimazole Induced Oxidative Stress in a Feline Kidney Epithelial Cell Line (CRFK). Veterinary Sciences. 2021; 8(10):220. https://doi.org/10.3390/vetsci8100220
Chicago/Turabian StyleGirolami, Flavia, Alessia Candellone, Watanya Jarriyawattanachaikul, Giorgia Meineri, Carlo Nebbia, and Paola Badino. 2021. "Protective Effect of Natural Antioxidant Compounds on Methimazole Induced Oxidative Stress in a Feline Kidney Epithelial Cell Line (CRFK)" Veterinary Sciences 8, no. 10: 220. https://doi.org/10.3390/vetsci8100220