Vaccination of Mice with a Novel Trypsin from Trichinella spiralis Elicits the Immune Protection against Larval Challenge

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasite, Animal and Antigens
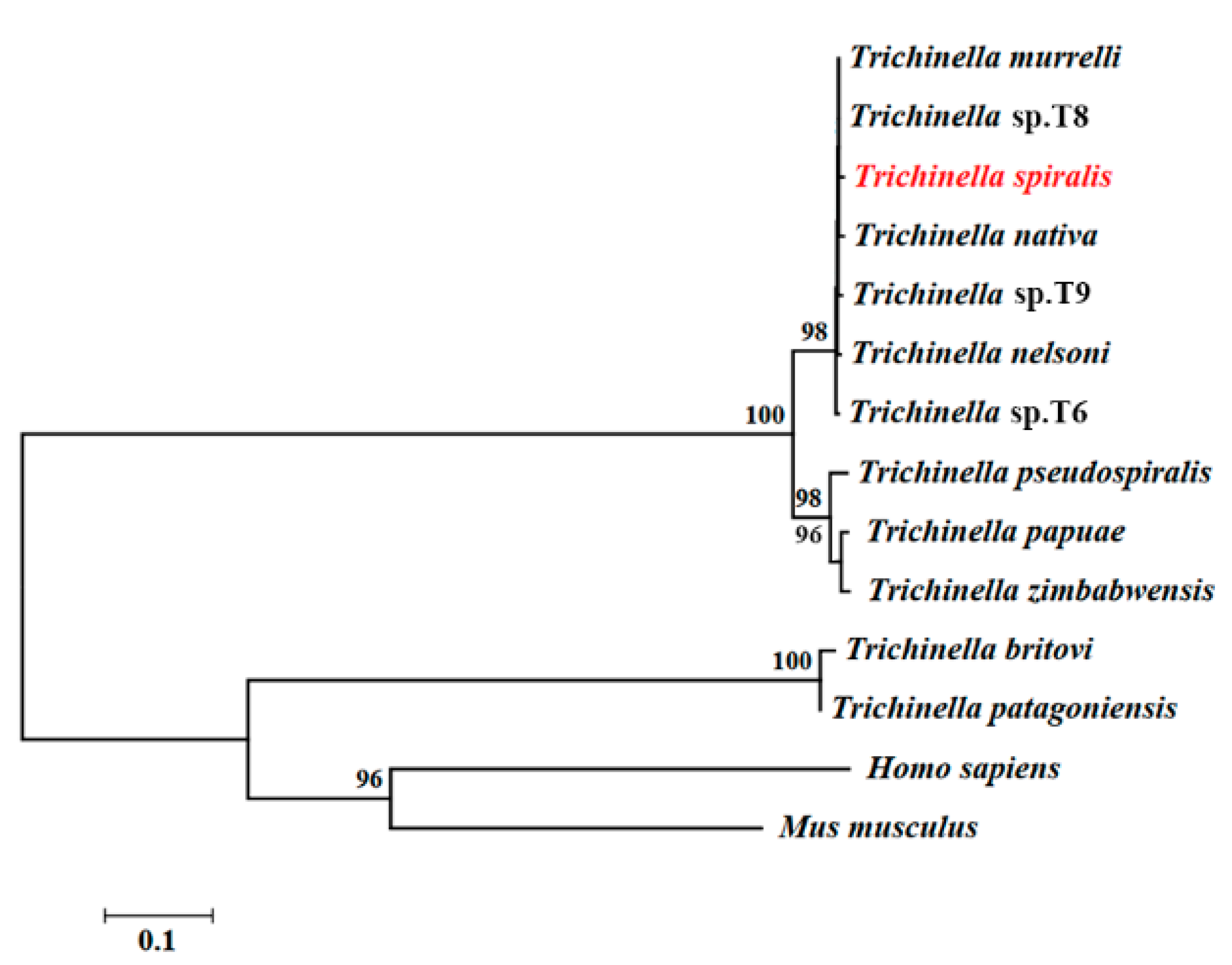
2.2. Bioinformatics Analysis of TsT Gene
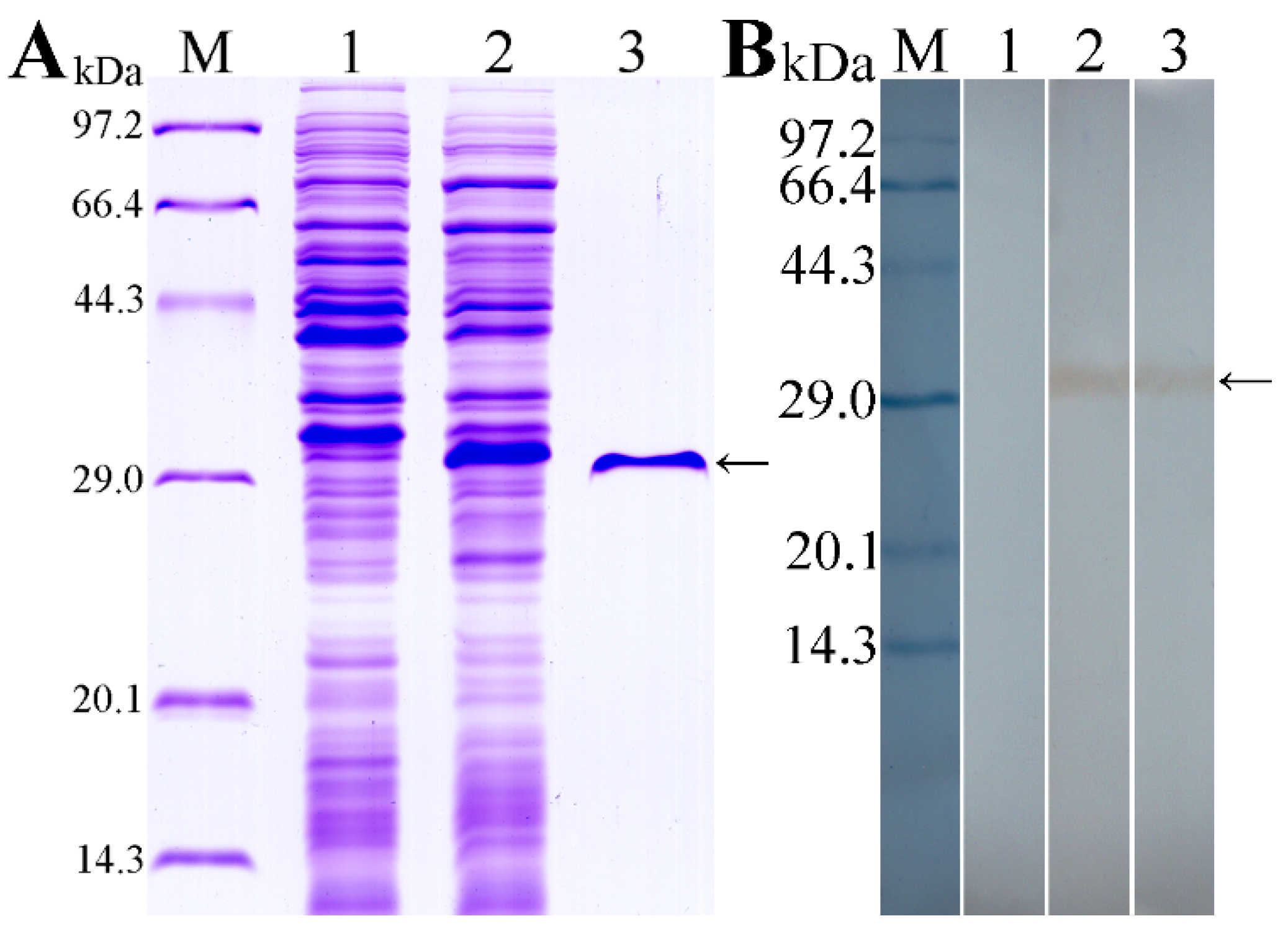
2.3. Cloning, Expression and Identification of TsT
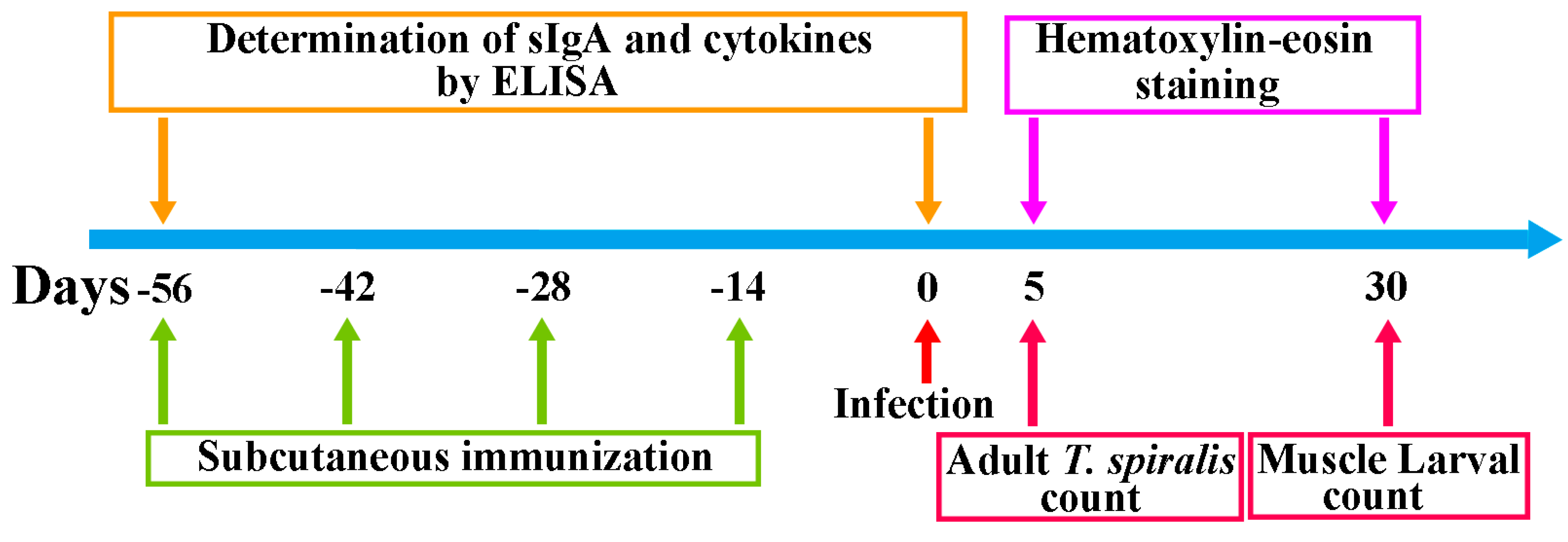
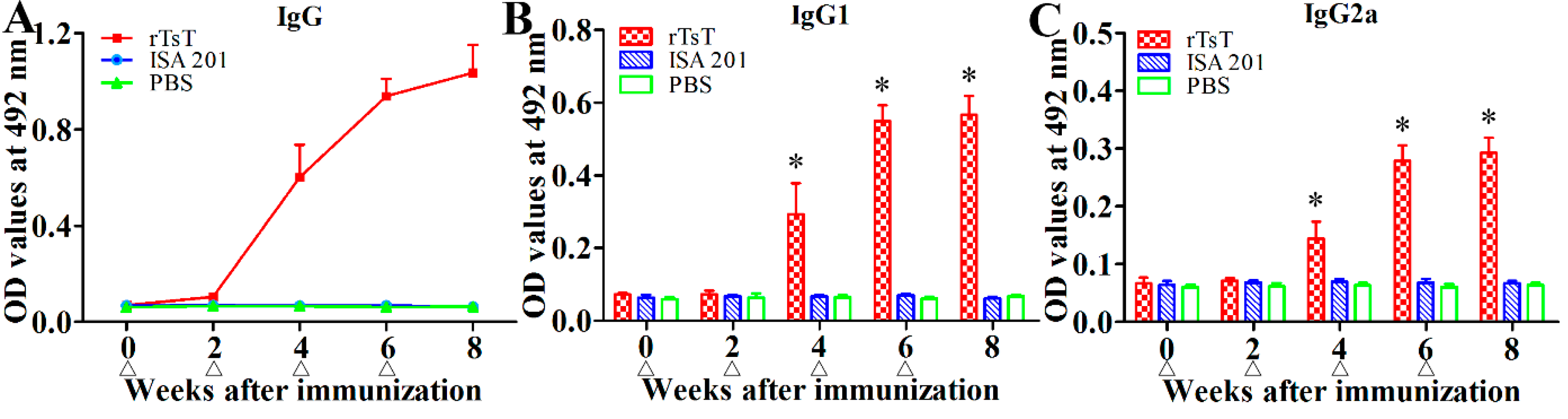
2.4. Immunization of Mice and ELISA Determination of Anti-rTsT Antibodies
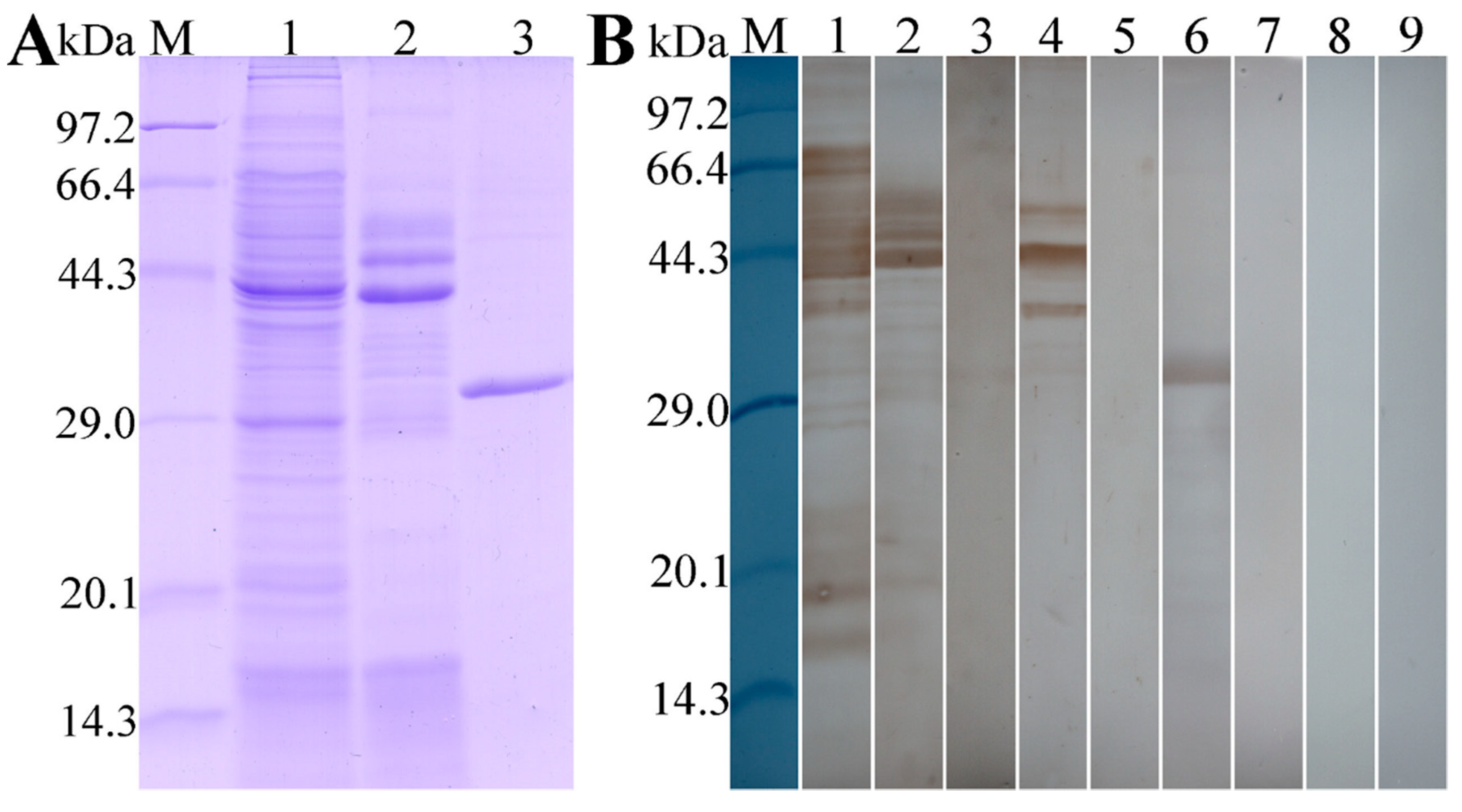
2.5. Western Blotting Analysis of rTsT Antigenicity
2.6. RT-PCR Analysis of TsT mRNA Expression in Diverse T. spiralis Stages
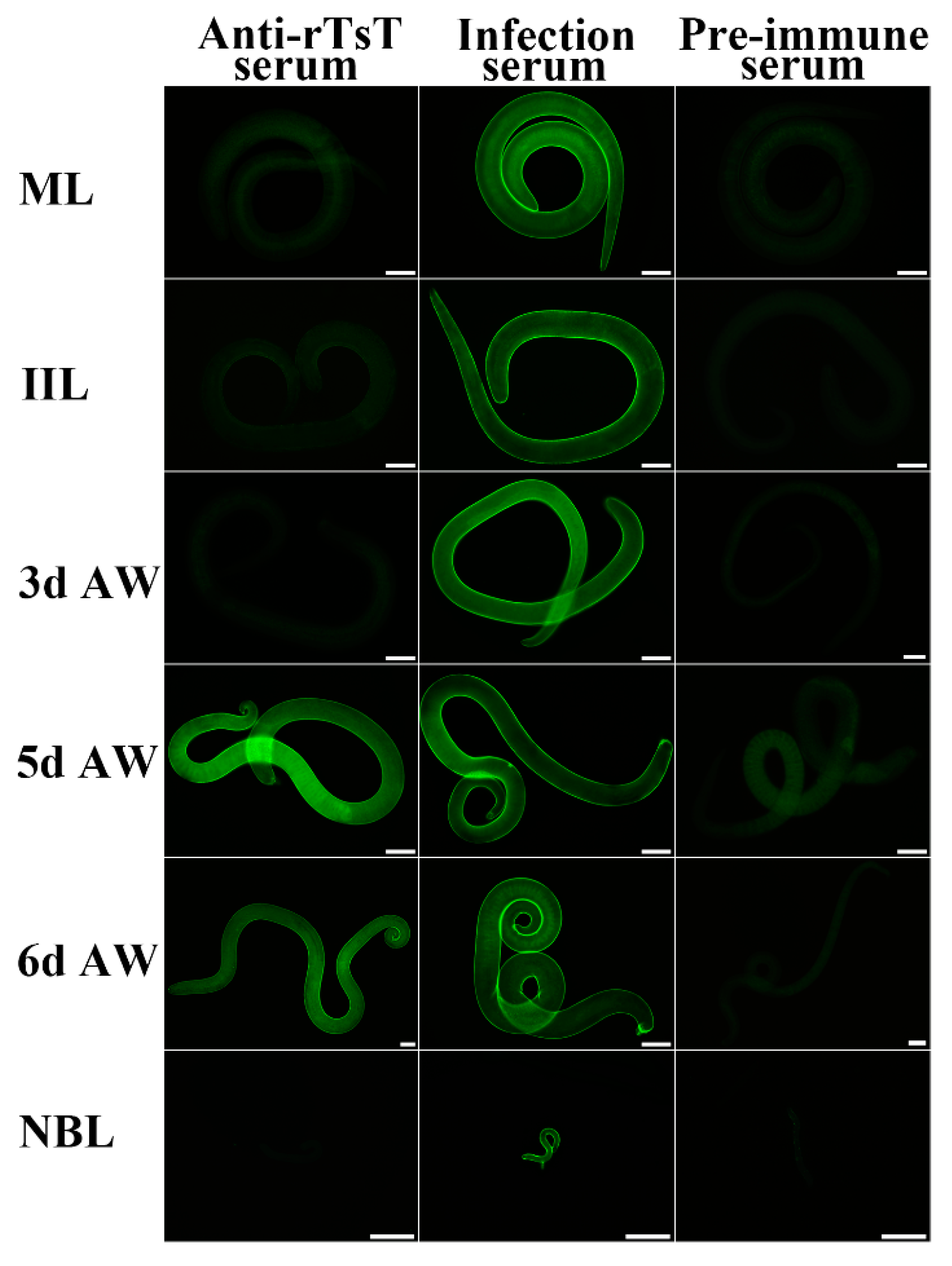
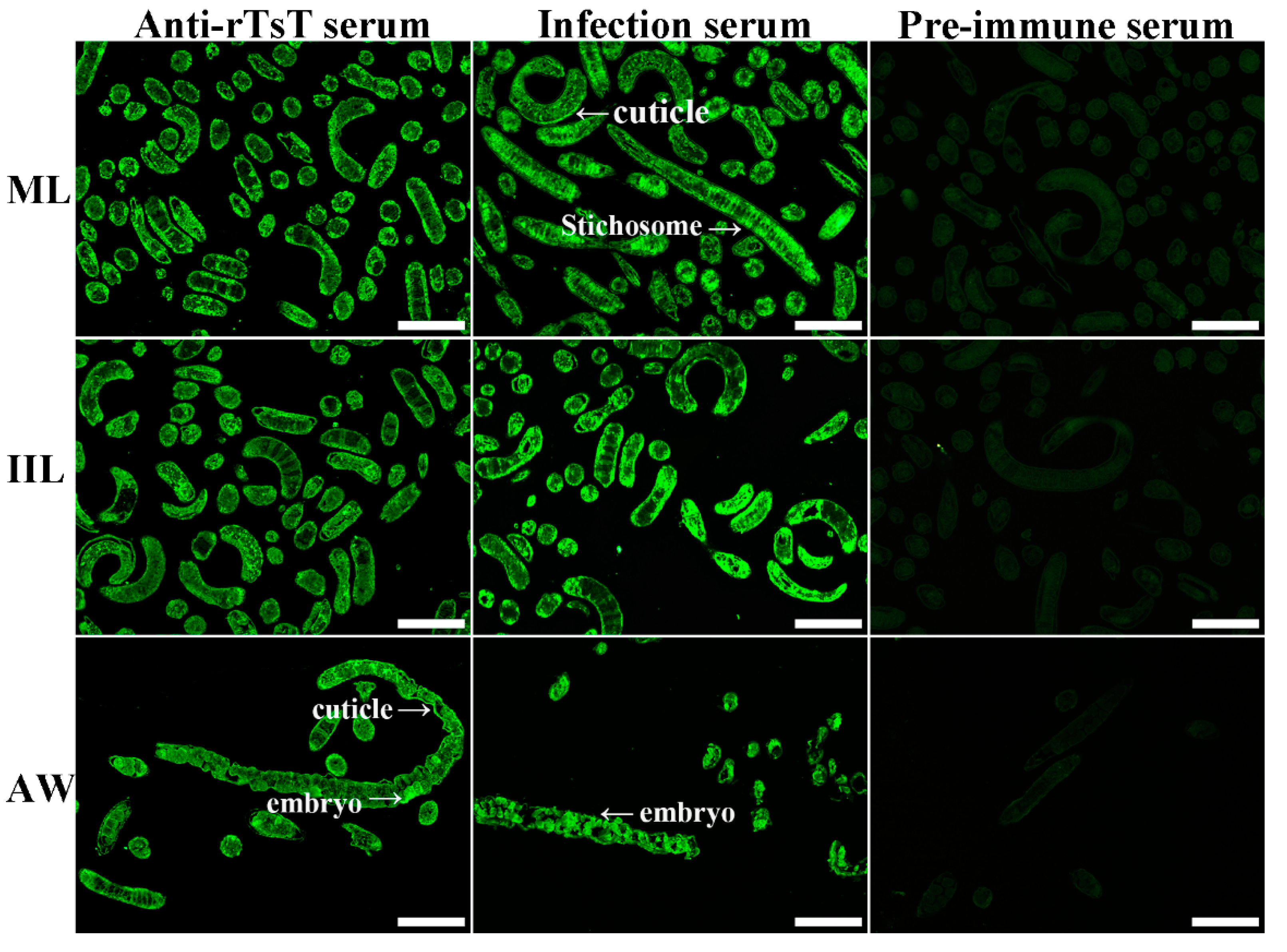
2.7. Indirect Immunofluorescence Test (IIFT)
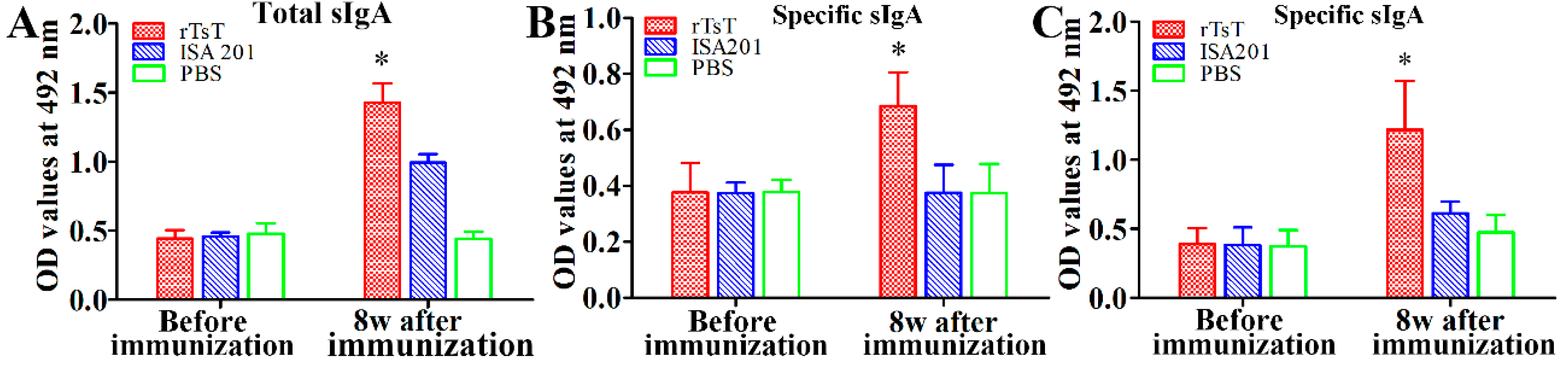
2.8. Assay of Total IgA and TsT-Specific IgA in Enteral Fluid
2.9. Assay of TsT-Specific Cytokines
2.10. Antibody-Dependent Cellular Cytotoxicity (ADCC)
2.11. Larval Challenge and Evaluation of Protection Efficacy
2.12. Histopathological Examination of Muscle and Intestines from Infected Mice
2.13. Statistical Analysis
3. Results
3.1. Bioinformatics Analysis of TsT
3.2. Expression and Western Blot Analysis of rTsT
3.3. TsT Transcription in Diverse T. Spiralis Stages
3.4. Expression and Localization of Native TsT in Various T. spiralis Stages
3.5. Anti-TsT Antibody Response
3.6. Enteral Mucosal Immune Response Induced by rTsT Immunization
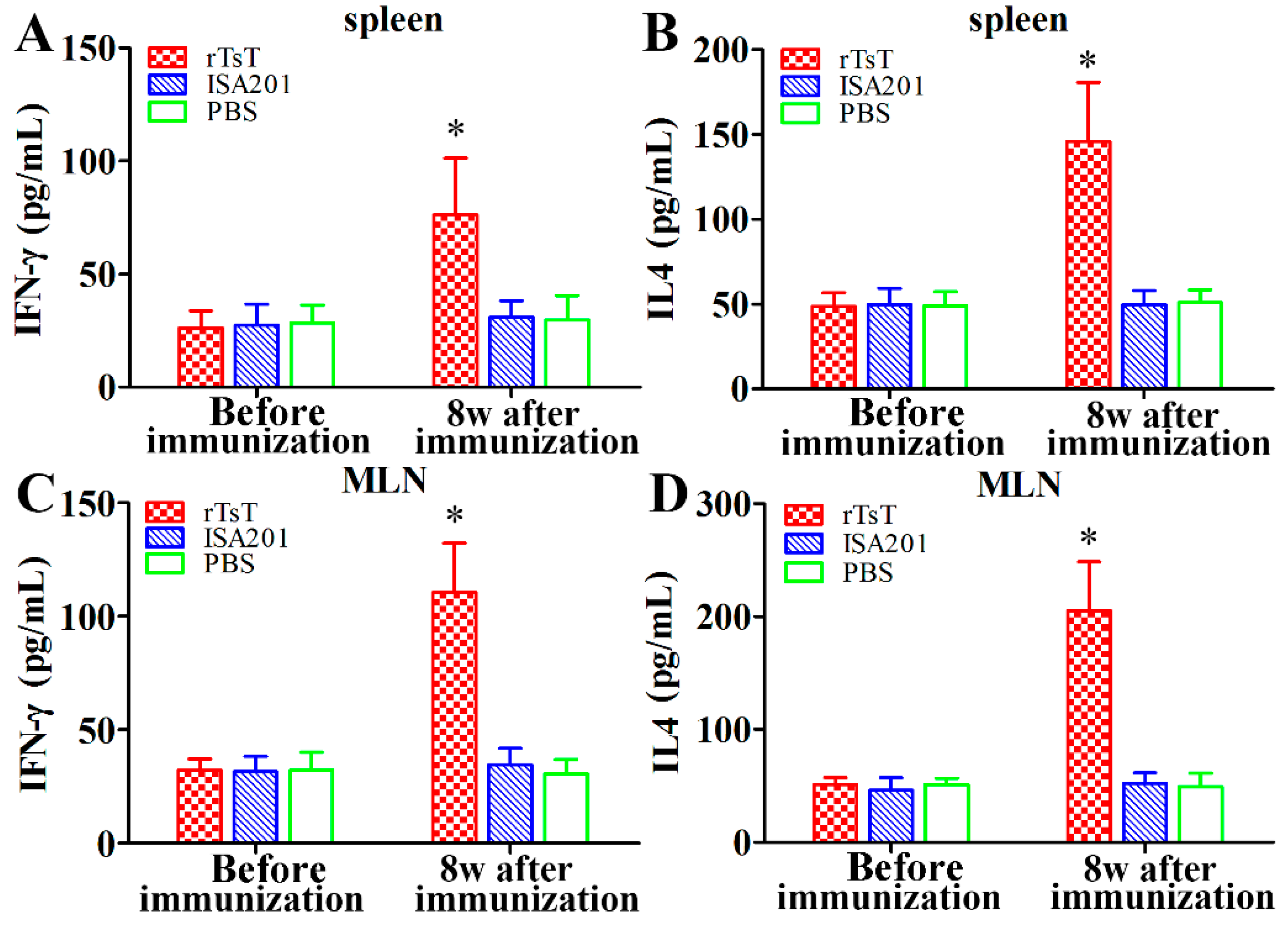
3.7. Cytokine Responses to the rTsT Immunization
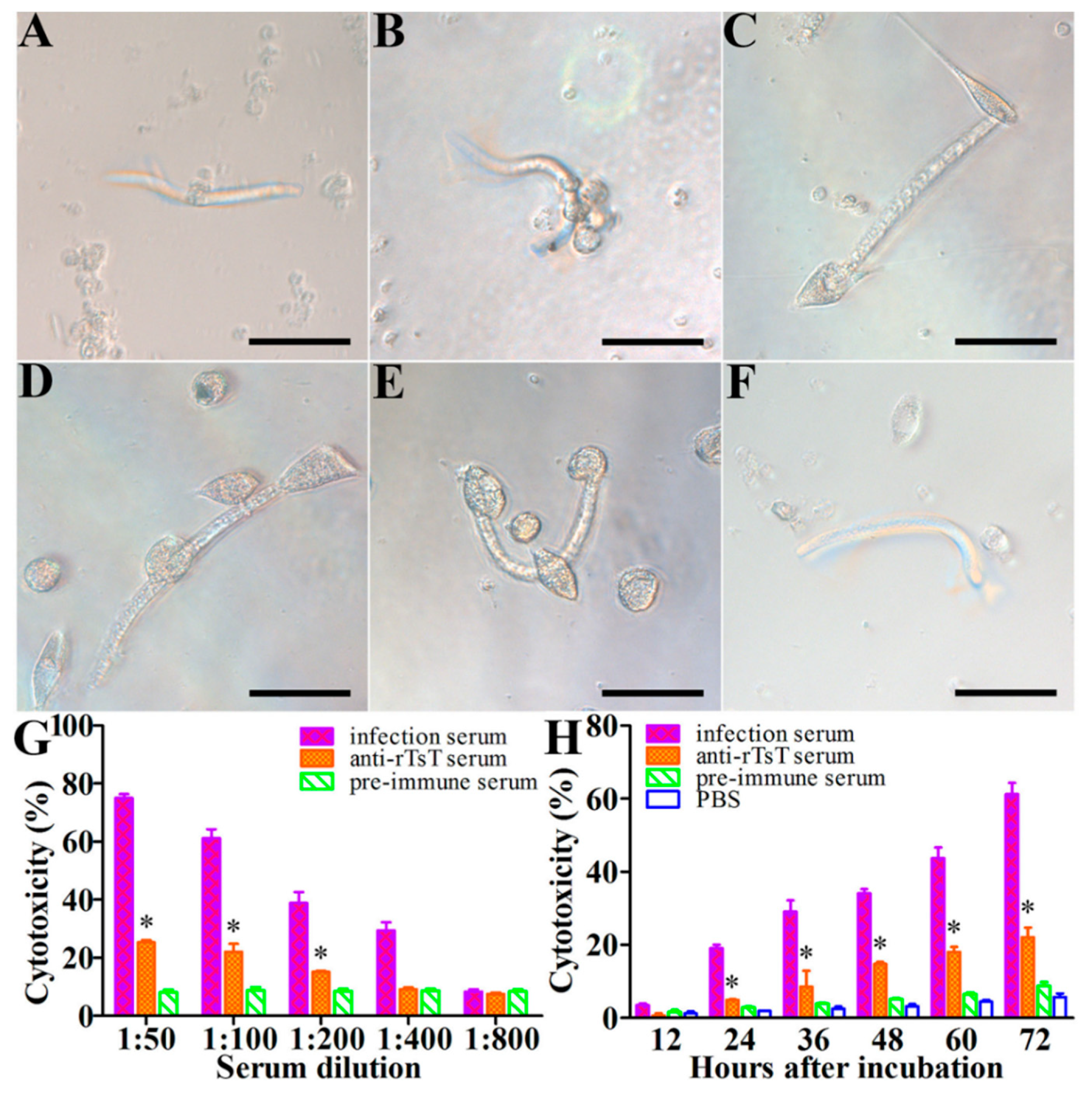
3.8. Killing and Destruction of the NBL by PECs and Anti-rTsT Immune Serum
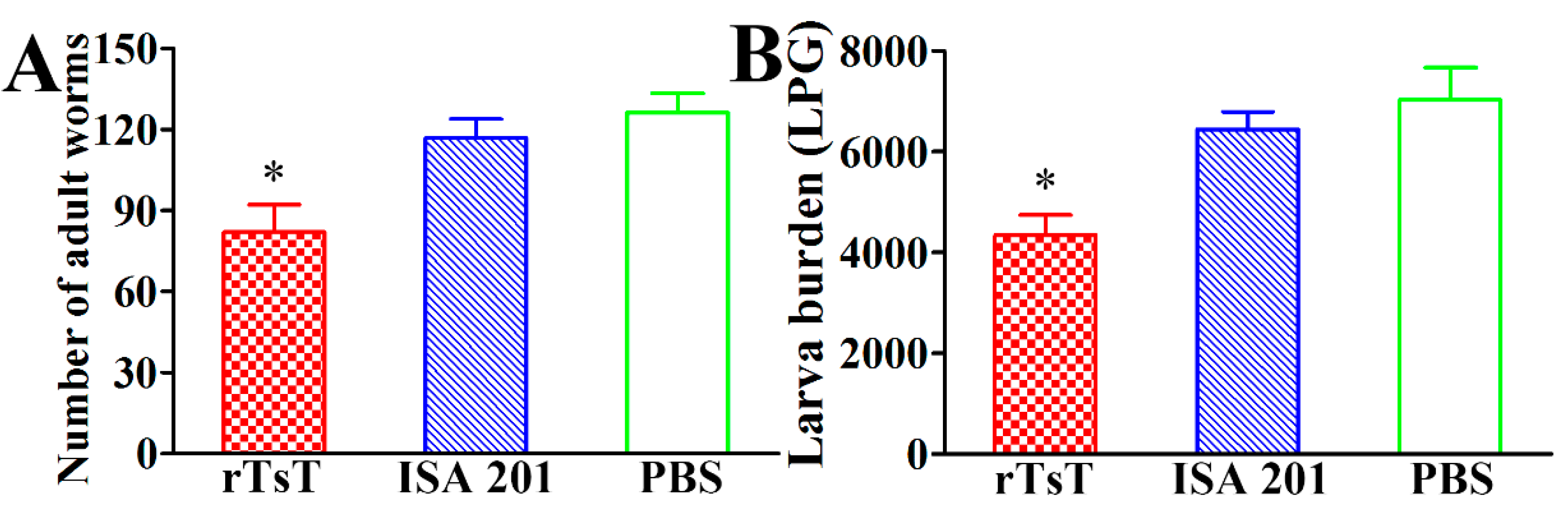
3.9. Immune Protection Induced by rTsT Immunization
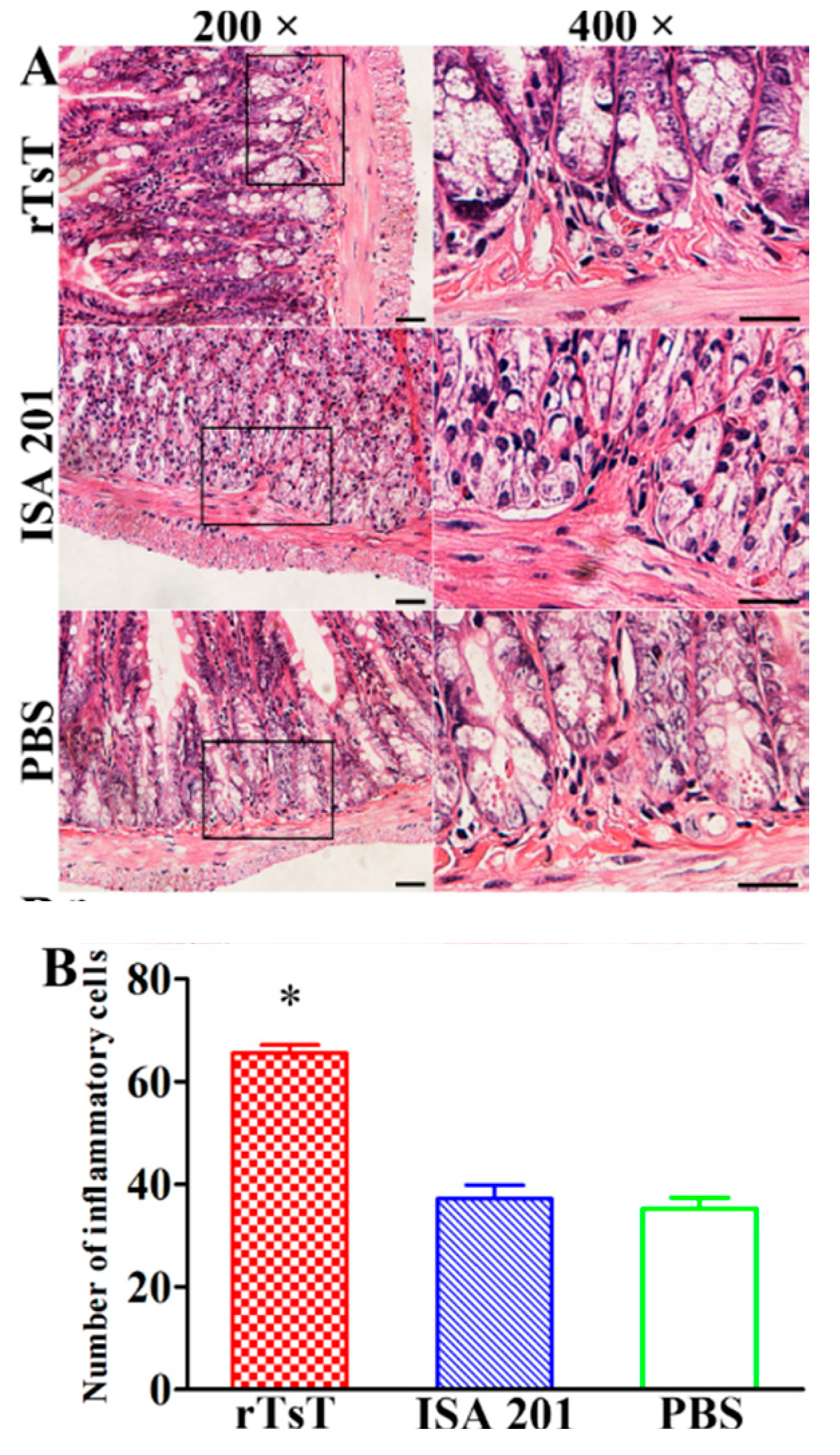
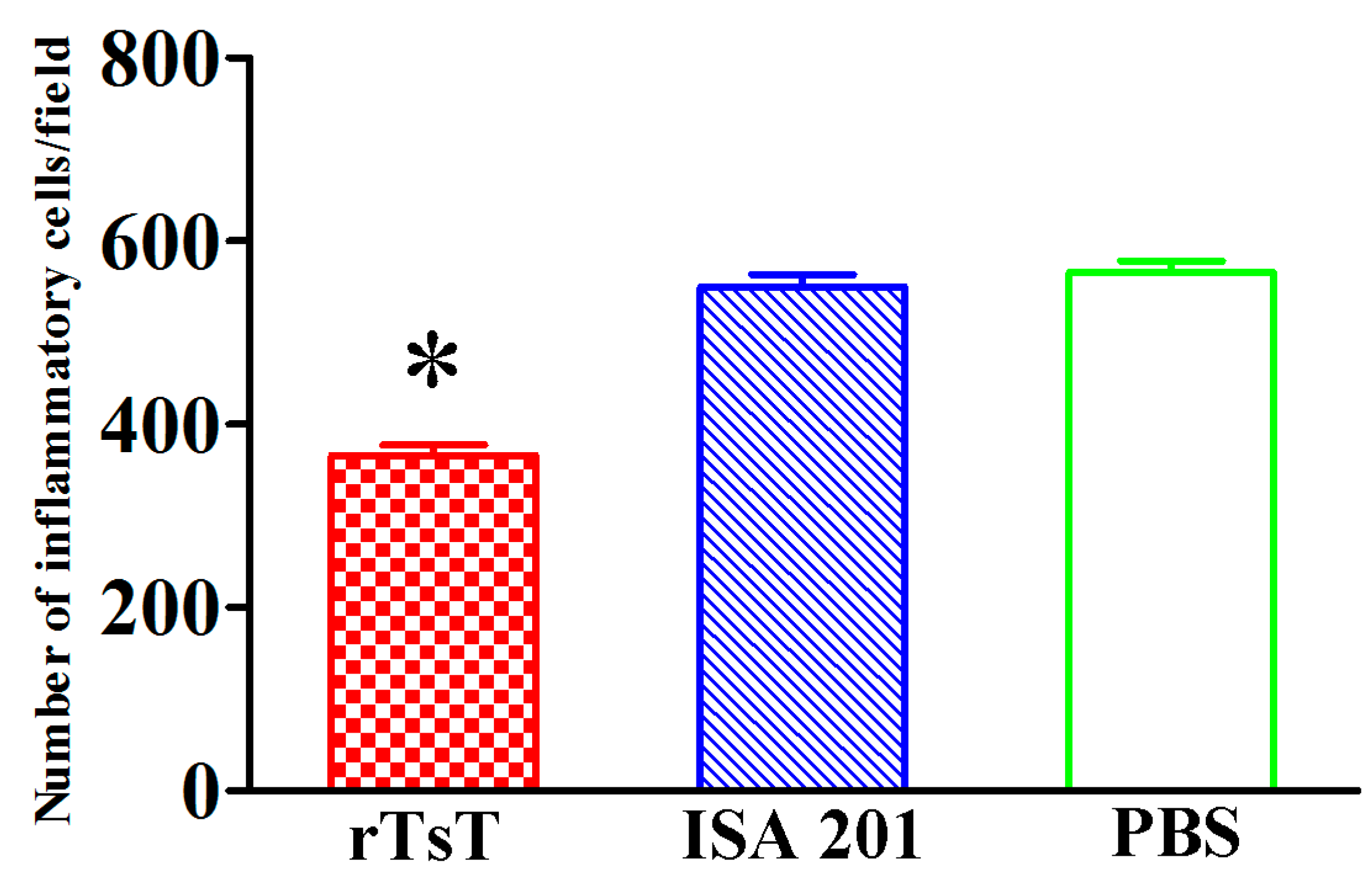
3.10. Intestinal and Muscle Histopathological Change of Infected Mice
4. Discussion
5. Conclusions
6. Ethics Approval and Consent to Participate
Author Contributions
Funding
Conflicts of Interest
References
- Pozio, E. World distribution of Trichinella spp. infections in animals and humans. Vet. Parasitol. 2007, 149, 3–21. [Google Scholar] [CrossRef]
- FAO/WHO. Multicriteria-based Ranking for Risk Management of Food-Borne Parasites; Food and Agriculture Organization of the United Nations/World Health Organization: Rome, Italy, 2014. [Google Scholar]
- Cui, J.; Jiang, P.; Liu, L.N.; Wang, Z.Q. Survey of Trichinella infections in domestic pigs from northern and eastern Henan, China. Vet. Parasitol. 2013, 194, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Zhang, X.; Wang, L.A.; Han, L.H.; Yang, M.; Duan, J.Y.; Sun, G.G.; Qi, X.; Liu, R.D.; Wang, Z.Q.; et al. Survey of Trichinella infection from domestic pigs in the historical endemic areas of Henan province, central China. Parasitol. Res. 2016, 115, 4707–4709. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Wang, Z.Q.; Xu, B.L. The epidemiology of human trichinellosis in China during 2004–2009. Acta Trop. 2011, 118, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murrell, K.D.; Pozio, E. Worldwide occurrence and impact of human trichinellosis, 1986–2009. Emerg. Infect. Dis. 2011, 17, 2194–2202. [Google Scholar] [CrossRef] [PubMed]
- Rostami, A.; Gamble, H.R.; Dupouy-Camet, J.; Khazan, H.; Bruschi, F. Meat sources of infection for outbreaks of human trichinellosis. Food Microbiol. 2017, 64, 65–71. [Google Scholar] [CrossRef]
- Liu, P.; Wang, Z.Q.; Liu, R.D.; Jiang, P.; Long, S.R.; Liu, L.N.; Zhang, X.Z.; Cheng, X.C.; Yu, C.; Ren, H.J.; et al. Oral vaccination of mice with Trichinella spiralis nudix hydrolase DNA vaccine delivered by attenuated Salmonella elicited protective immunity. Exp. Parasitol. 2015, 153, 29–38. [Google Scholar] [CrossRef]
- Zhang, N.; Li, W.; Fu, B. Vaccines against Trichinella spiralis: Progress, challenges and future prospects. Transbound Emerg. Dis. 2018, 65, 1447–1458. [Google Scholar] [CrossRef]
- Ren, H.J.; Cui, J.; Wang, Z.Q.; Liu, R.D. Normal mouse intestinal epithelial cells as a model for the in vitro invasion of Trichinella spiralis infective larvae. PLoS ONE 2011, 6, e27010. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.D.; Wang, Z.Q.; Wang, L.; Long, S.R.; Ren, H.J.; Cui, J. Analysis of differentially expressed genes of Trichinella spiralis larvae activated by bile and cultured with intestinal epithelial cells using real-time PCR. Parasitol. Res. 2013, 112, 4113–4120. [Google Scholar] [CrossRef]
- Despommier, D.D. How does Trichinella spiralis make itself at home? Parasitol. Today 1998, 14, 318–323. [Google Scholar] [CrossRef]
- Long, S.R.; Wang, Z.Q.; Jiang, P.; Liu, R.D.; Qi, X.; Liu, P.; Ren, H.J.; Shi, H.N.; Cui, J. Characterization and functional analysis of Trichinella spiralis nudix hydrolase. Exp. Parasitol. 2015, 159, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Liu, R.D.; Sun, G.G.; Song, Y.Y.; Jiang, P.; Zhang, X.; Cui, J. Proteomic analysis of Trichinella spiralis adult worm excretory-secretory proteins recognized by sera of patients with early trichinellosis. Front. Microbiol. 2017, 8, 986. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yang, F.; Yang, D.Q.; Jiang, P.; Liu, R.D.; Zhang, X.; Cui, J.; Wang, Z.Q. Molecular characterization of Trichinella spiralis galectin and its participation in larval invasion of host’s intestinal epithelial cells. Vet. Res. 2018, 49, 79. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Liu, R.D.; Bai, S.J.; Hao, H.N.; Yue, W.W.; Xu, Y.X.Y.; Long, S.R.; Cui, J.; Wang, Z.Q. Molecular characterization of a Trichinella spiralis aspartic protease and its facilitation role in larval invasion of host intestinal epithelial cells. PLoS Negl. Trop. Dis. 2020, 14, e0008269. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Li, L.G.; Liu, R.D.; Sun, G.G.; Liu, C.Y.; Zhang, S.B.; Jiang, P.; Zhang, X.; Ren, H.J.; Wang, Z.Q.; et al. Molecular identification and characterization of Trichinella spiralis proteasome subunit beta type-7. Parasit. Vectors 2015, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Bolas-Fernandez, F.; Corral Bezara, L.D. TSL-1 antigens of Trichinella: An overview of their potential role in parasite invasion, survival and serodiagnosis of trichinellosis. Res. Vet. Sci. 2006, 81, 297–303. [Google Scholar] [CrossRef]
- Wang, B.; Wang, Z.Q.; Jin, J.; Ren, H.J.; Liu, L.N.; Cui, J. Cloning, expression and characterization of a Trichinella spiralis serine protease gene encoding a 35.5 kDa protein. Exp. Parasitol. 2013, 134, 148–154. [Google Scholar] [CrossRef]
- Bien, J.; Cabaj, W.; Moskwa, B. Proteomic analysis of potential immunoreactive proteins from muscle larvae and adult worms of Trichinella spiralis in experimentally infected pigs. Folia Parasitol. 2015, 62, 22. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.D.; Cui, J.; Liu, X.L.; Jiang, P.; Sun, G.G.; Zhang, X.; Long, S.R.; Wang, L.; Wang, Z.Q. Comparative proteomic analysis of surface proteins of Trichinella spiralis muscle larvae and intestinal infective larvae. Acta Trop. 2015, 150, 79–86. [Google Scholar] [CrossRef]
- Liu, R.D.; Jiang, P.; Wen, H.; Duan, J.Y.; Wang, L.A.; Li, J.F.; Liu, C.Y.; Sun, G.G.; Wang, Z.Q.; Cui, J. Screening and characterization of early diagnostic antigens in excretory-secretory proteins from Trichinella spiralis intestinal infective larvae by immunoproteomics. Parasitol. Res. 2016, 115, 615–622. [Google Scholar] [CrossRef]
- Liu, R.D.; Qi, X.; Sun, G.G.; Jiang, P.; Zhang, X.; Wang, L.A.; Liu, X.L.; Wang, Z.Q.; Cui, J. Proteomic analysis of Trichinella spiralis adult worm excretory-secretory proteins recognized by early infection sera. Vet. Parasitol. 2016, 231, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cui, J.; Hu, D.D.; Liu, R.D.; Wang, Z.Q. Identification of early diagnostic antigens from major excretory-secretory proteins of Trichinella spiralis muscle larvae using immunoproteomics. Parasit. Vectors 2014, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Wang, L.; Cui, J. Proteomic analysis of Trichinella spiralis proteins in intestinal epithelial cells after culture with their larvae by shotgun LC-MS/MS approach. J. Proteom. 2012, 75, 2375–2383. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Z.Q.; Cui, J. Proteomic analysis of the changed proteins of Trichinella spiralis infective larvae after co-culture in vitro with intestinal epithelial cells. Vet. Parasitol. 2013, 194, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Nagano, I.; Wu, Z.; Takahashi, Y. Functional genes and proteins of Trichinella spp. Parasitol. Res. 2009, 104, 197–207. [Google Scholar] [CrossRef]
- Sun, G.G.; Ren, H.N.; Liu, R.D.; Song, Y.Y.; Qi, X.; Hu, C.X.; Yang, F.; Jiang, P.; Zhang, X.; Wang, Z.Q.; et al. Molecular characterization of a putative serine protease from Trichinella spiralis and its elicited immune protection. Vet. Res. 2018, 49, 59. [Google Scholar] [CrossRef] [Green Version]
- Li, J.F.; Guo, K.X.; Qi, X.; Lei, J.J.; Han, Y.; Yan, S.W.; Jiang, P.; Yu, C.; Cheng, X.C.; Wang, Z.Q.; et al. Protective immunity against Trichinella spiralis in mice elicited by oral vaccination with attenuated Salmonella-delivered TsSP1.2 DNA. Vet. Res. 2018, 49, 87. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.G.; Lei, J.J.; Ren, H.N.; Zhang, Y.; Guo, K.X.; Long, S.R.; Liu, R.D.; Jiang, P.; Wang, Z.Q.; Cui, J. Intranasal immunization with recombinant Trichinella spiralis serine protease elicits protective immunity in BALB/c mice. Exp. Parasitol. 2019, 201, 1–10. [Google Scholar] [CrossRef]
- Xu, J.; Bai, X.; Wang, L.B.; Shi, H.N.; Van Der Giessen, J.W.B.; Boireau, P.; Liu, M.Y.; Liu, X.L. Immune responses in mice vaccinated with a DNA vaccine expressing serine protease-like protein from the new-born larval stage of Trichinella spiralis. Parasitology 2017, 144, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Z.; Sun, X.Y.; Bai, Y.; Song, Y.Y.; Hu, C.X.; Li, X.; Cui, J.; Wang, Z.Q. Protective immunity in mice vaccinated with a novel elastase-1 significantly decreases Trichinella spiralis fecundity and infection. Vet. Res. 2020, 51, 43. [Google Scholar] [CrossRef] [Green Version]
- Mitreva, M.; Jasmer, D.P.; Zarlenga, D.S.; Wang, Z.; Abubucker, S.; Martin, J.; Taylor, C.M.; Yin, Y.; Fulton, L.; Minx, P.; et al. The draft genome of the parasitic nematode Trichinella spiralis. Nat. Genet. 2011, 43, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.Q.; Li, L.Z.; Jiang, P.; Liu, L.N.; Cui, J. Molecular identification and phylogenetic analysis of Trichinella isolates from different provinces in mainland China. Parasitol. Res. 2012, 110, 753–757. [Google Scholar] [CrossRef]
- Jiang, P.; Wang, Z.Q.; Cui, J.; Zhang, X. Comparison of artificial digestion and Baermann’s methods for detection of Trichinella spiralis pre-encapsulated larvae in muscles with low-level infections. Foodborne Pathog. Dis. 2012, 9, 27–31. [Google Scholar] [CrossRef]
- Sun, G.G.; Wang, Z.Q.; Liu, C.Y.; Jiang, P.; Liu, R.D.; Wen, H.; Qi, X.; Wang, L.; Cui, J. Early serodiagnosis of trichinellosis by ELISA using excretory–secretory antigens of Trichinella spiralis adult worms. Parasit. Vectors 2015, 8, 484. [Google Scholar] [CrossRef] [Green Version]
- Li, L.G.; Wang, Z.Q.; Liu, R.D.; Yang, X.; Liu, L.N.; Sun, G.G.; Jiang, P.; Zhang, X.; Zhang, G.Y.; Cui, J. Trichinella spiralis: Low vaccine potential of glutathione S-transferase against infections in mice. Acta Trop. 2015, 146, 25–32. [Google Scholar] [CrossRef]
- Wu, Z.; Nagano, I.; Takahashi, Y.; Maekawa, Y. Practical methods for collecting Trichinella parasites and their excretory-secretory products. Parasitol. Int. 2016, 65, 591–595. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Z.Q.; Cui, J. Protein changes in Trichinella spiralis muscle larvae in vitro induced by bovine bile. Vet. Parasitol. 2013, 194, 164–167. [Google Scholar] [CrossRef]
- Long, S.R.; Wang, Z.Q.; Liu, R.D.; Liu, L.N.; Li, L.G.; Jiang, P.; Zhang, X.; Zhang, Z.F.; Shi, H.N.; Cui, J. Molecular identification of Trichinella spiralis nudix hydrolase and its induced protective immunity against trichinellosis in BALB/c mice. Parasit. Vectors 2014, 7, 600. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.N.; Wang, Z.Q.; Zhang, X.; Jiang, P.; Qi, X.; Liu, R.D.; Zhang, Z.F.; Cui, J. Characterization of Spirometra erinaceieuropaei plerocercoid cysteine protease and potential application for serodiagnosis of sparganosis. PLoS Negl. Trop. Dis. 2015, 9, e0003807. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Yue, X.; Han, Y.; Jiang, P.; Yang, F.; Lei, J.J.; Liu, R.D.; Zhang, X.; Wang, Z.Q.; Cui, J. Characterization of two Trichinella spiralis adult-specific DNase II and their capacity to induce protective immunity. Front. Microbiol. 2018, 9, 2504. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.Y.; Song, Y.Y.; Ren, H.N.; Sun, G.G.; Liu, R.D.; Jiang, P.; Long, S.R.; Zhang, X.; Wang, Z.Q.; Cui, J. Cloning and expression of a Trichinella spiralis putative glutathione S-transferase and its elicited protective immunity against challenge infections. Parasit. Vectors 2017, 10, 448. [Google Scholar] [CrossRef]
- Sun, G.G.; Song, Y.Y.; Jiang, P.; Ren, H.N.; Yan, S.W.; Han, Y.; Liu, R.D.; Zhang, X.; Wang, Z.Q.; Cui, J. Characterization of a Trichinella spiralis putative serine protease. Study of its potential as sero-diagnostic tool. PLoS Negl. Trop. Dis. 2018, 12, e0006485. [Google Scholar] [CrossRef]
- Wang, S.W.; Wang, Z.Q.; Cui, J. Protein change of intestinal epithelial cells induced in vitro by Trichinella spiralis infective larvae. Parasitol. Res. 2011, 108, 593–599. [Google Scholar] [CrossRef]
- Liu, R.D.; Cui, J.; Wang, L.; Long, S.R.; Zhang, X.; Liu, M.Y.; Wang, Z.Q. Identification of surface proteins of Trichinella spiralis muscle larvae using immunoproteomics. Trop. Biomed. 2014, 31, 579–591. [Google Scholar]
- Hu, C.X.; Jiang, P.; Yue, X.; Zeng, J.; Zhang, X.Z.; Song, Y.Y.; Liu, R.D.; Zhang, X.; Wang, Z.Q.; Cui, J. Molecular characterization of a Trichinella spiralis elastase-1 and its potential as a diagnostic antigen for trichinellosis. Parasit. Vectors 2020, 13, 97. [Google Scholar] [CrossRef]
- Cui, J.; Ren, H.J.; Liu, R.D.; Wang, L.; Zhang, Z.F.; Wang, Z.Q. Phage-displayed specific polypeptide antigens induce significant protective immunity against Trichinella spiralis infection in BALB/c mice. Vaccine 2013, 31, 1171–1177. [Google Scholar] [CrossRef]
- Cui, J.; Wang, L.; Sun, G.G.; Liu, L.N.; Zhang, S.B.; Liu, R.D.; Zhang, X.; Jiang, P.; Wang, Z.Q. Characterization of a Trichinella spiralis 31 kDa protein and its potential application for the serodiagnosis of trichinellosis. Acta Trop. 2015, 142, 57–63. [Google Scholar] [CrossRef]
- Sun, G.G.; Liu, R.D.; Wang, Z.Q.; Jiang, P.; Wang, L.; Liu, X.L.; Liu, C.Y.; Zhang, X.; Cui, J. New diagnostic antigens for early trichinellosis: The excretory-secretory antigens of Trichinella spiralis intestinal infective larvae. Parasitol. Res. 2015, 114, 4637–4644. [Google Scholar] [CrossRef]
- Liu, C.Y.; Ren, H.N.; Song, Y.Y.; Sun, G.G.; Liu, R.D.; Jiang, P.; Long, S.R.; Zhang, X.; Wang, Z.Q.; Cui, J. Characterization of a putative glutathione S-transferase of the parasitic nematode Trichinella spiralis. Exp. Parasitol. 2018, 187, 59–66. [Google Scholar] [CrossRef]
- Han, Y.; Yue, X.; Hu, C.X.; Liu, F.; Liu, R.D.; He, M.M.; Long, S.R.; Cui, J.; Wang, Z.Q. Interaction of a Trichinella spiralis cathepsin B on enterocytes promotes the larval intrusion into the cells. Res. Vet. Sci. 2020, 130, 110–117. [Google Scholar] [CrossRef]
- Yang, F.; Guo, K.X.; Yang, D.Q.; Liu, R.D.; Long, S.R.; Zhang, X.; Jiang, P.; Cui, J.; Wang, Z.Q. Functional analysis of Trichinella spiralis serine protease 1.2 by siRNA mediated RNA interference. Trop. Biomed. 2020, 37, 457–470. [Google Scholar]
- Xu, J.; Liu, R.D.; Long, S.R.; Song, Y.Y.; Jiang, P.; Zhang, X.; Cui, J.; Wang, Z.Q. Characterization of a chymotrypsin-like enzyme from Trichinella spiralis and its facilitation of larva penetration into the host’s enteral epithelial cells. Res. Vet. Sci. 2020, 128, 1–8. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Li, L.; Cui, J. Molecular characterization of Trichinella spiralis aminopeptidase and its potential as a novel vaccine candidate antigen against trichinellosis in BALB/c mice. Parasit. Vectors 2013, 6, 246. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Han, Y.; Yue, X.; Liu, F.; Song, Y.Y.; Yan, S.W.; Lei, J.J.; Zhang, X.; Jiang, P.; Wang, Z.Q. Vaccination of mice with a recombinant novel cathepsin B inhibits Trichinella spiralis development, reduces the fecundity and worm burden. Parasit. Vectors 2019, 12, 581. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Han, Y.; Jiang, P.; Yue, X.; Ren, H.N.; Sun, G.G.; Long, S.R.; Yu, C.; Cheng, X.C.; Cui, J.; et al. Oral vaccination with Trichinella spiralis DNase II DNA vaccine delivered by attenuated Salmonella induces a protective immunity in BALB/c mice. Vet. Res. 2018, 49, 119. [Google Scholar] [CrossRef] [Green Version]
- Guo, K.X.; Bai, Y.; Ren, H.N.; Sun, X.Y.; Song, Y.Y.; Liu, R.D.; Long, S.R.; Zhang, X.; Jiang, P.; Wang, Z.Q.; et al. Characterization of a Trichinella spiralis aminopeptidase and its participation in invasion, development and fecundity. Vet. Res. 2020, 51, 78. [Google Scholar] [CrossRef]
- Bermudez-Cruz, R.M.; Fonseca-Linan, R.; Grijalva-Contreras, L.E.; Mendoza-Hernandez, G.; Ortega-Pierres, M.G. Proteomic analysis and immunodetection of antigens from early developmental stages of Trichinella spiralis. Vet. Parasitol. 2016, 231, 22–31. [Google Scholar] [CrossRef]
- Liu, L.N.; Jing, F.J.; Cui, J.; Fu, G.Y.; Wang, Z.Q. Detection of circulating antigen in serum of mice infected with Trichinella spiralis by an IgY-IgM mAb sandwich ELISA. Exp. Parasitol. 2013, 133, 150–155. [Google Scholar] [CrossRef]
- Dea-Ayuela, M.A.; Rama-Iniguez, S.; Bolas-Fernandez, F. Vaccination of mice against intestinal Trichinella spiralis infections by oral administration of antigens microencapsulated in methacrilic acid copolymers. Vaccine 2006, 24, 2772–2780. [Google Scholar] [CrossRef]
- Liu, P.; Cui, J.; Liu, R.D.; Wang, M.; Jiang, P.; Liu, L.N.; Long, S.R.; Li, L.G.; Zhang, S.B.; Zhang, X.Z.; et al. Protective immunity against Trichinella spiralis infection induced by TsNd vaccine in mice. Parasit. Vectors 2015, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Sun, X.; Li, B.; Huang, J.; Zhan, B.; Zhu, X. Vaccination with a paramyosin-based multi-epitope vaccine elicits significant protective immunity against Trichinella spiralis infection in mice. Front. Microbiol. 2017, 8, 1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moskwa, B. Trichinella spiralis: In vitro cytotoxicity of peritoneal cells against synchronous newborn larvae of different age. Parasitol. Res. 1999, 85, 59–63. [Google Scholar] [CrossRef]
- Cui, J.; Li, L.G.; Jiang, P.; Liu, R.D.; Yang, X.; Liu, L.N.; Liu, P.; Zhang, S.B.; Wang, Z.Q. Biochemical and functional characterization of the glutathione S-transferase from Trichinella spiralis. Parasitol. Res. 2015, 114, 2007–2013. [Google Scholar] [CrossRef]
- Song, Y.Y.; Zhang, Y.; Ren, H.N.; Sun, G.G.; Qi, X.; Yang, F.; Jiang, P.; Zhang, X.; Cui, J.; Wang, Z.Q. Characterization of a serine protease inhibitor from Trichinella spiralis and its participation in larval invasion of host’s intestinal epithelial cells. Parasit. Vectors 2018, 11, 499. [Google Scholar] [CrossRef]
- Song, Y.Y.; Wang, L.A.; Na Ren, H.; Qi, X.; Sun, G.G.; Liu, R.D.; Jiang, P.; Zhang, X.; Cui, J.; Wang, Z.Q. Cloning, expression and characterisation of a cysteine protease from Trichinella spiralis. Folia Parasitol. 2018, 65. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Zhang, S.B.; Jiang, P.; Liu, R.D.; Long, S.R.; Zhang, X.; Ren, H.J.; Cui, J. The siRNA-mediated silencing of Trichinella spiralis nudix hydrolase results in reduction of larval infectivity. Parasitol. Res. 2015, 114, 3551–3557. [Google Scholar] [CrossRef]
- Yang, F.; Yang, D.Q.; Song, Y.Y.; Guo, K.X.; Li, Y.L.; Long, S.R.; Jiang, P.; Cui, J.; Wang, Z.Q. In vitro silencing of a serine protease inhibitor suppresses Trichinella spiralis invasion, development, and fecundity. Parasitol. Res. 2019, 118, 2247–2255. [Google Scholar] [CrossRef]
- Gu, Y.; Sun, X.; Huang, J.; Zhan, B.; Zhu, X. A multiple antigen peptide vaccine containing CD4(+) t cell epitopes enhances humoral immunity against Trichinella spiralis infection in mice. J. Immunol. Res. 2020, 2020, 2074803. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Tang, B.; Yang, Y.; Cai, X.; Jia, W.; Luo, X.; Yan, H.; Zhang, Z.; Lin, J.; Liu, M.; et al. Vaccination with a DNase II recombinant protein against Trichinella spiralis infection in pigs. Vet. Parasitol. 2020. [Google Scholar] [CrossRef]
- Dzik, J.M. Molecules released by helminth parasites involved in host colonization. Acta Biochim. Pol. 2006, 53, 33–64. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wen, Y.; Cai, Y.N.; Vallee, I.; Boireau, P.; Liu, M.Y.; Cheng, S.P. Serine proteases of parasitic helminths. Korean J. Parasitol. 2015, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Tang, B.; Bai, X.; Wang, L.; Wu, X.; Shi, H.; Wang, X.; Liu, X.; Liu, M. Characterization of an antigenic serine protease in the Trichinella spiralis adult. Exp. Parasitol. 2018, 195, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, D.I.; Crawford, C.R.; Duce, I.R.; Behnke, J.M. Antigen stripping from the nematode epicuticle using the cationic detergent cetyltrimethylammonium bromide (CTAB). Parasite Immunol. 1985, 7, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Pompa-Mera, E.N.; Arroyo-Matus, P.; Ocana-Mondragon, A.; Gonzalez-Bonilla, C.R.; Yepez-Mulia, L. Protective immunity against enteral stages of Trichinella spiralis elicited in mice by live attenuated Salmonella vaccine that secretes a 30-mer parasite epitope fused to the molecular adjuvant C3d-P28. Res. Vet. Sci. 2014, 97, 533–545. [Google Scholar] [CrossRef]
- Della Bella, C.; Benagiano, M.; De Gennaro, M.; Gomez-Morales, M.A.; Ludovisi, A.; D′Elios, S.; Luchi, S.; Pozio, E.; D′Elios, M.M.; Bruschi, F. T-cell clones in human trichinellosis: Evidence for a mixed Th1/Th2 response. Parasite Immunol. 2017, 39. [Google Scholar] [CrossRef]
- Bell, R.G.; Appleton, J.A.; Negrao-Correa, D.A.; Adams, L.S. Rapid expulsion of Trichinella spiralis in adult rats mediated by monoclonal antibodies of distinct IgG isotypes. Immunology 1992, 75, 520–527. [Google Scholar]
- Inaba, T.; Sato, H.; Kamiya, H. Monoclonal IgA antibody-mediated expulsion of Trichinella from the intestine of mice. Parasitology 2003, 126, 591–598. [Google Scholar] [CrossRef]
- Ren, H.N.; Guo, K.X.; Zhang, Y.; Liu, R.D.; Jiang, P.; Zhang, X.; Wang, L.; Cui, J.; Wang, Z.Q. Molecular characterization of a 31 kDa protein from Trichinella spiralis and its induced immune protection in BALB/c mice. Parasit. Vectors 2018, 11, 625. [Google Scholar] [CrossRef] [Green Version]
- Helmby, H.; Grencis, R.K. Contrasting roles for IL-10 in protective immunity to different life cycle stages of intestinal nematode parasites. Eur. J. Immunol. 2003, 33, 2382–2390. [Google Scholar] [CrossRef]
- Castillo-Alvarez, A.M.; Vaquero-Vera, A.; Fonseca-Liñán, R.; Ruiz-Pérez, F.; Villegas-Sepúlveda, N.; Ortega-Pierres, G. A prime-boost vaccination of mice with attenuated Salmonella expressing a 30-mer peptide from the Trichinella spiralis gp43 antigen. Vet. Parasitol. 2013, 194, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Parkhouse, R.M.; Ortega-Pierres, G. Stage-specific antigens of Trichinella spiralis. Parasitology 1984, 88, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.N.; Liu, R.D.; Song, Y.Y.; Zhuo, T.X.; Guo, K.X.; Zhang, Y.; Jiang, P.; Wang, Z.Q.; Cui, J. Label-free quantitative proteomic analysis of molting-related proteins of Trichinella spiralis intestinal infective larvae. Vet. Res. 2019, 50, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]















© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zeng, J.; Song, Y.Y.; Long, S.R.; Liu, R.D.; Jiang, P.; Zhang, X.; Cui, J.; Wang, Z.Q. Vaccination of Mice with a Novel Trypsin from Trichinella spiralis Elicits the Immune Protection against Larval Challenge. Vaccines 2020, 8, 437. https://doi.org/10.3390/vaccines8030437
Zhang Y, Zeng J, Song YY, Long SR, Liu RD, Jiang P, Zhang X, Cui J, Wang ZQ. Vaccination of Mice with a Novel Trypsin from Trichinella spiralis Elicits the Immune Protection against Larval Challenge. Vaccines. 2020; 8(3):437. https://doi.org/10.3390/vaccines8030437
Chicago/Turabian StyleZhang, Yao, Jie Zeng, Yan Yan Song, Shao Rong Long, Ruo Dan Liu, Peng Jiang, Xi Zhang, Jing Cui, and Zhong Quan Wang. 2020. "Vaccination of Mice with a Novel Trypsin from Trichinella spiralis Elicits the Immune Protection against Larval Challenge" Vaccines 8, no. 3: 437. https://doi.org/10.3390/vaccines8030437