Hydrostatic Filtration Enables Large-Scale Production of Outer Membrane Vesicles That Effectively Protect Chickens against Gallibacterium anatis

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Immunogens Production
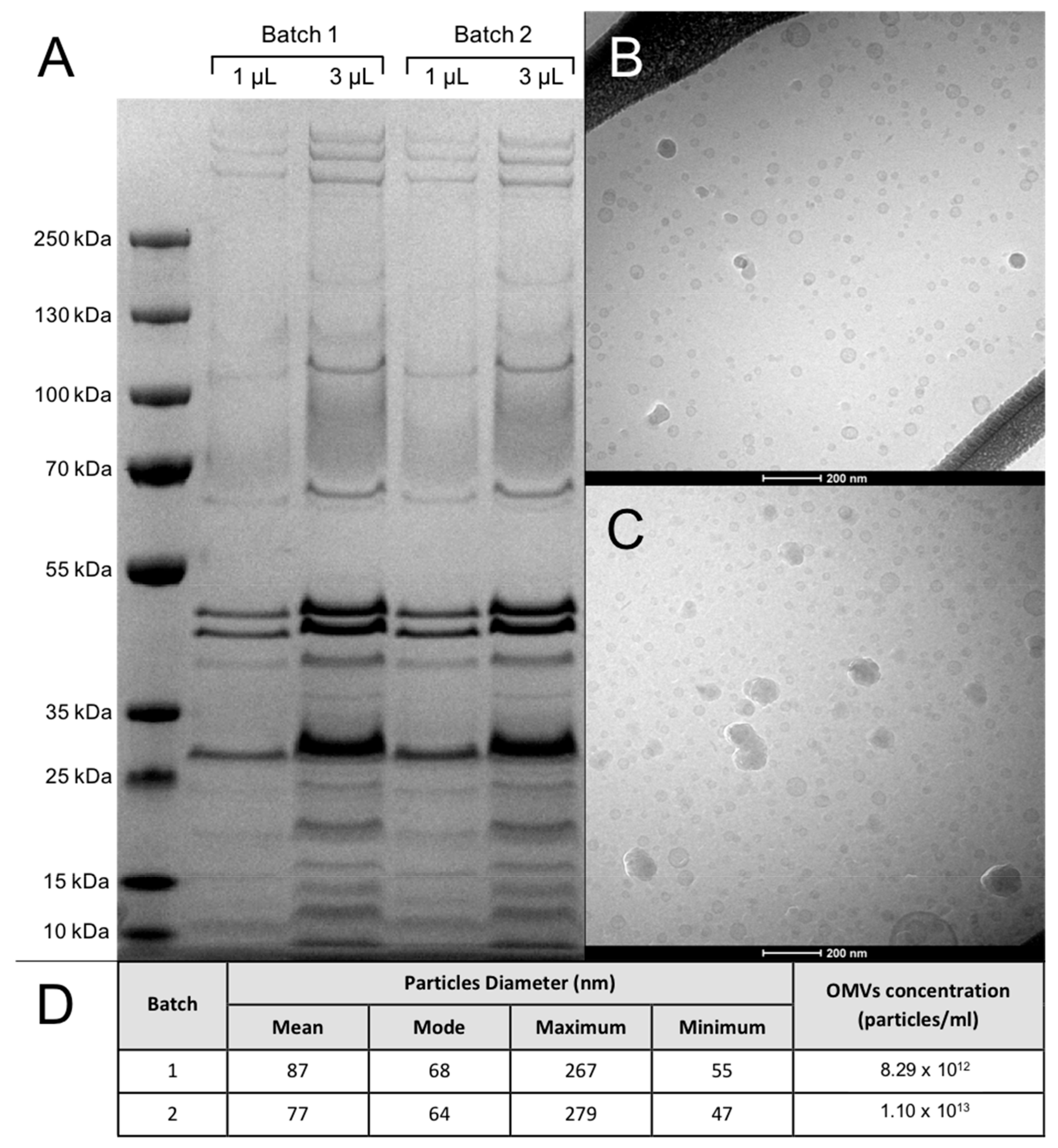
2.2. Characterization of OMV Batches
- (i)
- Different OMV concentrations were loaded on 10% NuPAGE® Novex® Bis–Tris gels (Invitrogen, Carlsbad, CA, USA) and run under reducing conditions. PageRulerTM Plus Prestained Protein Ladder (Thermofisher Scientific, Waltham, MA, USA) was used for size determination. Protein gels were then stained with SimplyBlueTM SafeStain (Invitrogen). Protein concentration was qualitatively estimated by comparison with a protein standard of known concentration (not shown).
- (ii)
- Of the nanodispersed OMVs 3 µL was applied on a hydrophilized lacey carbon 300 mesh copper grid (Ted Pella Inc., Redding, CA, USA). The excess sample on the grid was then blotted with filter paper at a blotting time of 3 s, blotting force 0, temperature 4 °C and 100% humidity (FEI Vitrobot IV), and was rapidly plunged into liquid nitrogen cooled ethane (−180 °C). Sample observations were performed using a Tecnai G2 20 transmission electron microscope (FEI) at a voltage of 200 kV under a low-dose rate. Images were recorded with an FEI Eagle camera 4 k × 4 k at variable nominal magnifications.
- (iii)
- OMVs batches were quantified by TRPS using a qNano device (Izon Sciences Ltd., Christchurch, New Zealand) following a standard protocol [33]. Data were analyzed using the data capture and analysis software, Izon Control Suite V.3.3.2.2001.
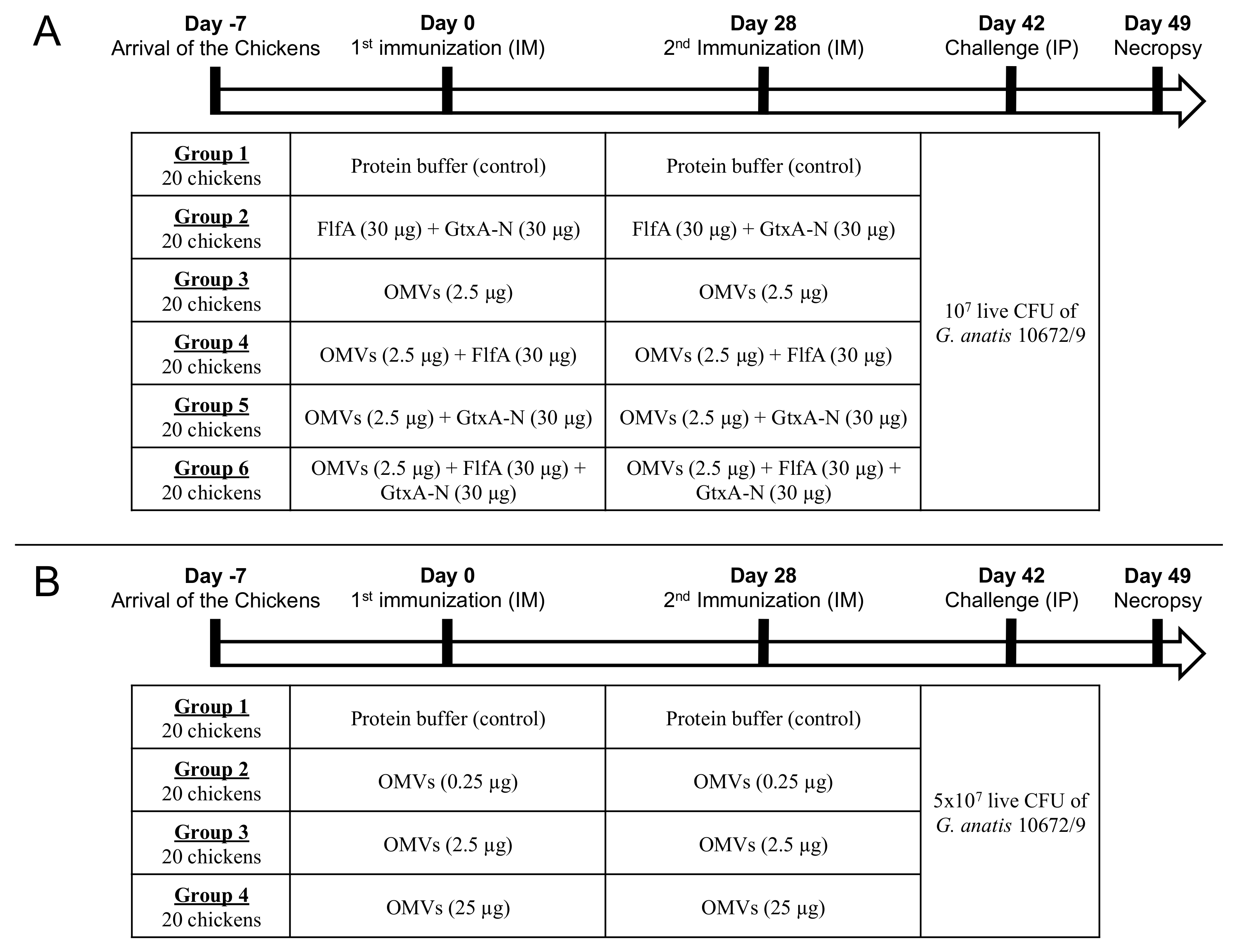
2.3. In Vivo Studies
2.4. Animal Model
2.5. Immunization
2.6. Challenge
2.7. Post-Mortem Examination
2.8. Serum Preparation
2.9. IgY Extraction from Egg Yolk
2.10. Antibody Titration
3. Results
3.1. OMV Batches Analysis
3.2. Concentration of Challenge Inocula
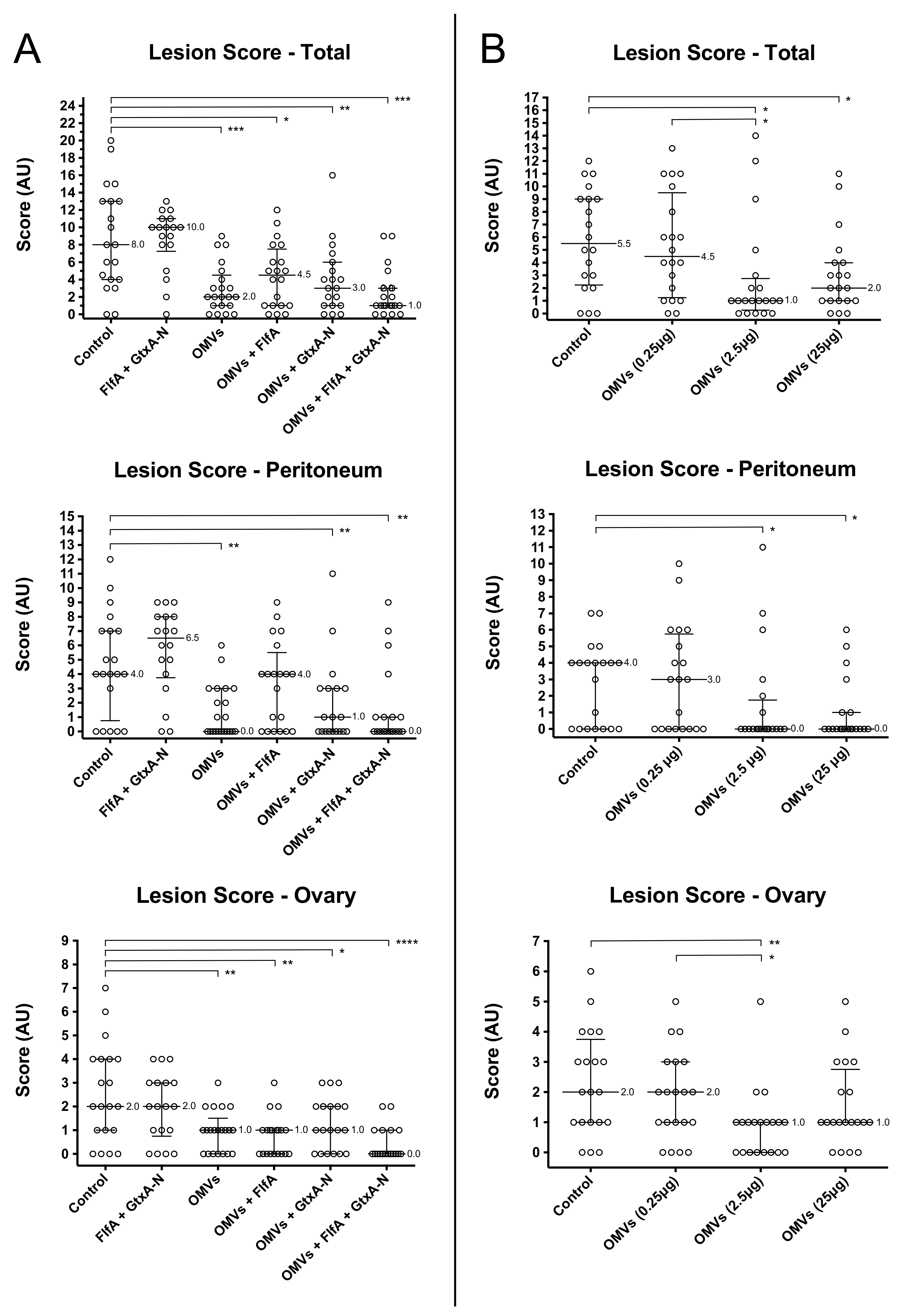
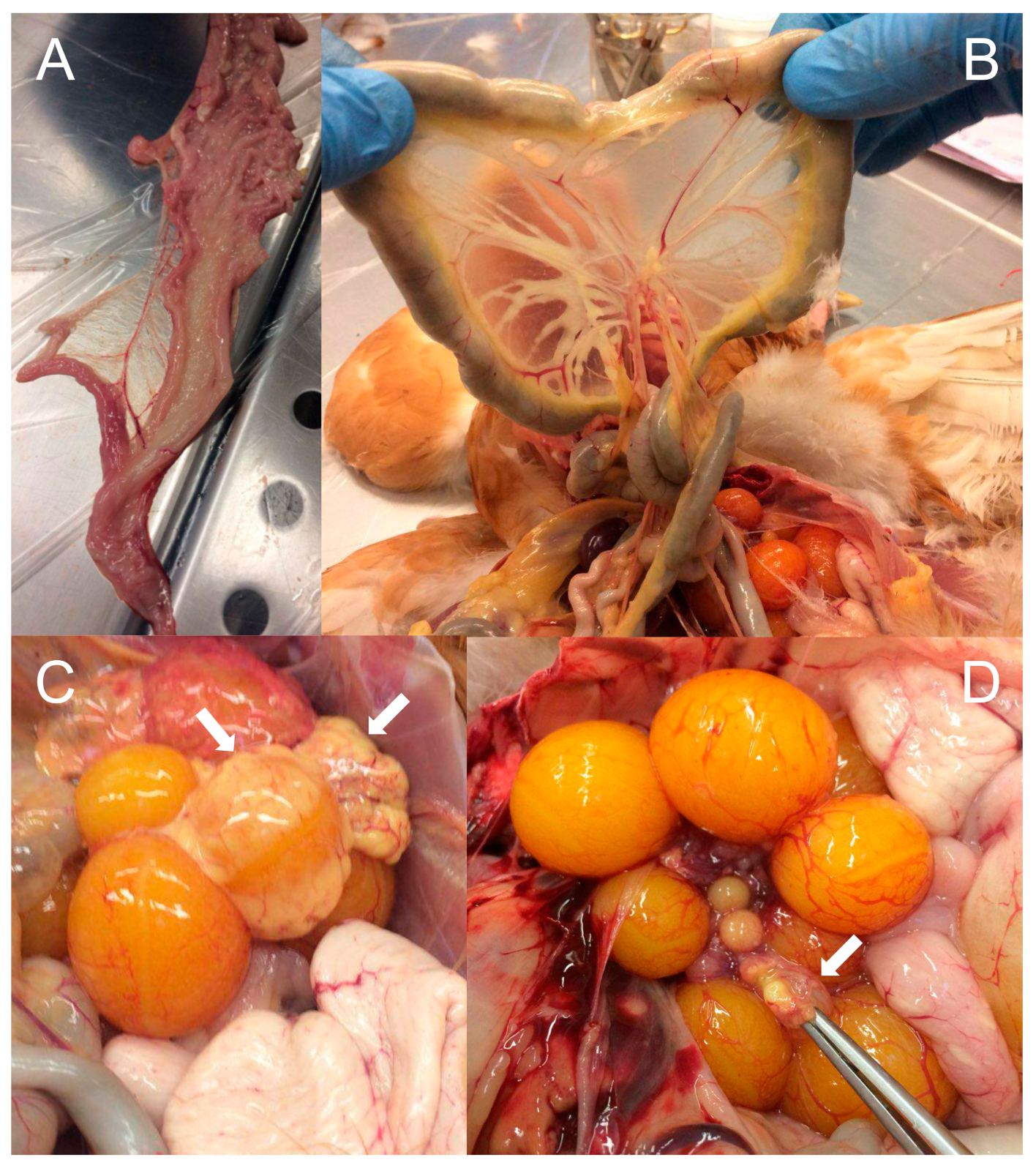
3.3. Pathological Analysis
- Study 1: Animal groups that received OMV-based immunizations (groups 3–6) presented significantly different total and ovary lesion scores, respectively, as compared to the control group (group 1). On the contrary, immunization with FlfA and GtxA-N alone (group 2) failed instead to produce any observable effect on lesion scores when compared to the control group.
- Study 2: Animals that were immunized with 2.5 μg OMVs (group 3) presented significantly different peritoneum, ovary and total lesion scores as compared to the unprotected control group (group 1). Animals immunized with 25 μg OMVs (group 4) only differed significantly from the control group in the peritoneum and total lesion scores. Immunization with 0.25 μg OMVs (group 2) failed to produce any observable reduction in lesion scores as compared to the control group.
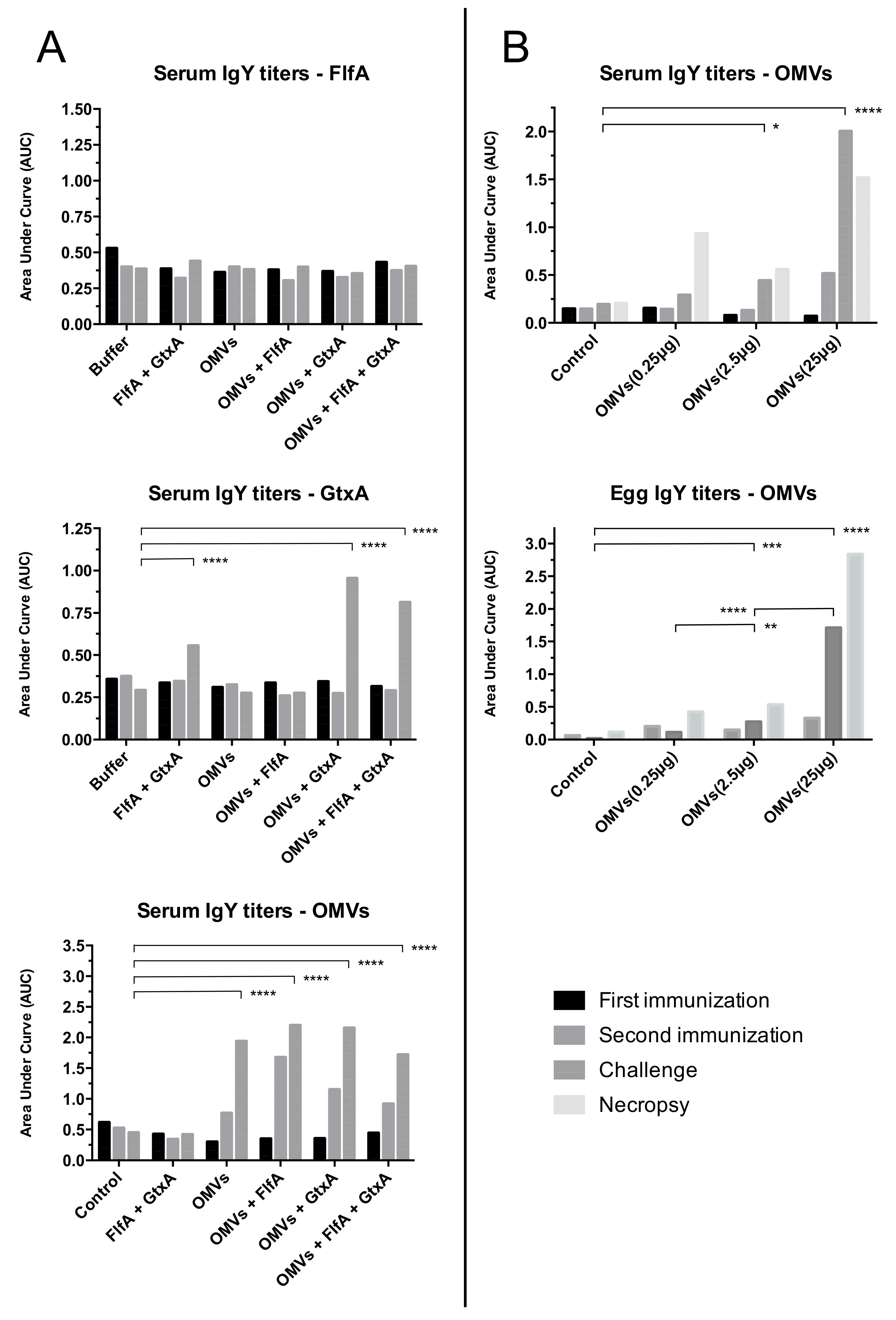
3.4. Antibody Response to Immunization
- Study 1: Immunization with G. anatis OMVs (groups 3–6) induced significantly higher OMV-specific IgY titers, even after first immunization (T1), than the controls. None of the animals immunized with FlfA however developed any detectable FlfA-specific IgY titers. Immunization with GtxA-N (groups 2, 5 and 6) induced significantly increased GtxA-N-specific IgY titers, although not until the second immunization was completed (T2).
- Study 2: Immunization with 2.5 μg and 25 μg OMVs (group 3 and 4, respectively) induced significantly increased OMV-specific serum IgY titers of immunized animals. Similar titers were also detected in pooled yolks from the eggs laid by animals immunized with 2.5 μg and 25 μg OMVs. The lower immunization dose of 0.25 μg (group 3) did not appear to induce detectable serum or yolk specific IgY titers.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bojesen, A.M.; Christensen, J.P.; Bisgaard, M. Gallibacterium infections and other avian Pasteurellaceae. In Poultry Diseases; Elsevier Health Sciences: New York, NY, USA, 2008; p. 632. ISBN 9780702028625. [Google Scholar]
- Persson, G.; Bojesen, A.M. Bacterial determinants of importance in the virulence of Gallibacterium anatis in poultry. Vet. Res. 2015, 46, 57. [Google Scholar] [CrossRef] [Green Version]
- Bojesen, A.M.; Nielsen, O.L.; Christensen, J.P.; Bisgaard, M. In vivo studies of Gallibacterium anatis infection in chickens. Avian Pathol. 2004, 33, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Paudel, S.; Liebhart, D.; Hess, M.; Hess, C. Pathogenesis of Gallibacterium anatis in a natural infection model fulfils Koch’s postulates: 1. Folliculitis and drop in egg production are the predominant effects in specific pathogen free layers. Avian Pathol. 2014, 43, 443–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, F.T.W.; Williams, N.J.; Wattret, A.; Jones, T. Observations on salpingitis, peritonitis and salpingoperitonitis in a layer breeder flock. Vet. Rec. 2005, 157, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, C.; De Souza-Pilz, M.; Bojesen, A.M.; Bisgaard, M.; Hess, M. Tissue distribution of haemolytic Gallibacterium anatis isolates in laying birds with reproductive disorders. Avian Pathol. 2009, 38, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.H.; Thornton, J.K.; Zhang, Y.; Mauel, M.J. A 5-year retrospective report of Gallibacterium anatis and pasteurella multocida isolates from chickens in Mississippi. Poult. Sci. 2013, 92, 3166–3171. [Google Scholar] [CrossRef]
- Varan Singh, S.; R Singh, B. Gallibacterium anatis: An Emerging Pathogen of Poultry Birds and Domiciled Birds. J. Vet. Sci. Technol. 2016, 7, 324. [Google Scholar] [CrossRef]
- Elhamid, H.; Mataried, N.; Elbestawy, A.; Bekheet, A.; Lakany, H. Pathogenicity of Ten Gallibacterium anatis isolates in commercial broiler chickens. Alex. J. Vet. Sci. 2016, 49, 42–49. [Google Scholar] [CrossRef]
- Paudel, S.; Hess, M.; Hess, C. Coinfection of Avibacterium paragallinarum and Gallibacterium anatis in Specific-Pathogen-Free Chickens Complicates Clinical Signs of Infectious Coryza, Which Can Be Prevented by Vaccination. Avian Dis. 2017, 61, 55–63. [Google Scholar] [CrossRef]
- Christensen, H.; Bisgaard, M.; Bojesen, A.M.; Mutters, R.; Olsen, J.E. Genetic relationships among avian isolates classified as Pasteurella haemolytica, “Actinobacillus salpingitidis” or Pasteurella anatis with proposal of Gallibacterium anatis gen. nov., comb. nov. and description of additional genomospecies within Gallibacterium gen. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 275–287. [Google Scholar]
- Johnson, T.J.; Danzeisen, J.L.; Trampel, D.; Nolan, L.K.; Seemann, T.; Bager, R.J.; Bojesen, A.M. Genome Analysis and Phylogenetic Relatedness of Gallibacterium anatis Strains from Poultry. PLoS ONE 2013, 8, e54844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef] [Green Version]
- Kulp, A.; Kuehn, M.J. Biological Functions and Biogenesis of Secreted Bacterial Outer Membrane Vesicles. Annu. Rev. Microbiol. 2012, 64, 163–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Pol, L.; Stork, M.; van der Ley, P. Outer membrane vesicles as platform vaccine technology. Biotechnol. J. 2015, 10, 1689–1706. [Google Scholar] [CrossRef] [PubMed]
- Vernikos, G.; Medini, D. Bexsero® chronicle. Pathog. Glob. Health 2014, 108, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Antenucci, F.; Fougeroux, C.; Deeney, A.; Ørskov, C.; Rycroft, A.; Holst, P.J.; Bojesen, A.M. In vivo testing of novel vaccine prototypes against Actinobacillus pleuropneumoniae. Vet. Res. 2018, 49, 4. [Google Scholar] [CrossRef] [Green Version]
- Persson, G.; Pors, S.E.; Thøfner, I.C.N.; Bojesen, A.M. Vaccination with outer membrane vesicles and the fimbrial protein FlfA offers improved protection against lesions following challenge with Gallibacterium anatis. Vet. Microbiol. 2018, 217, 104–111. [Google Scholar] [CrossRef]
- Pritsch, M.; Ben-Khaled, N.; Chaloupka, M.; Kobold, S.; Berens-Riha, N.; Peter, A.; Liegl, G.; Schubert, S.; Hoelscher, M.; Löscher, T.; et al. Comparison of Intranasal Outer Membrane Vesicles with Cholera Toxin and Injected MF59C.1 as Adjuvants for Malaria Transmission Blocking Antigens AnAPN1 and Pfs48/45. J. Immunol. Res. 2016, 2016. [Google Scholar] [CrossRef]
- Tan, K.; Li, R.; Huang, X.; Liu, Q. Outer Membrane Vesicles: Current Status and Future Direction of These Novel Vaccine Adjuvants. Front. Microbiol. 2018, 9, 783. [Google Scholar] [CrossRef] [Green Version]
- Bager, R.J.; Persson, G.; Nesta, B.; Soriani, M.; Serino, L.; Jeppsson, M.; Nielsen, T.K.; Bojesen, A.M. Outer membrane vesicles reflect environmental cues in Gallibacterium anatis. Vet. Microbiol. 2013, 167, 565–572. [Google Scholar] [CrossRef]
- Bager, R.J.; Nesta, B.; Pors, S.E.; Soriani, M.; Serino, L.; Boyce, J.D.; Adler, B.; Bojesen, A.M. The fimbrial protein FlfA from gallibacterium anatis is a virulence factor and vaccine candidate. Infect. Immun. 2013, 81, 1964–1973. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, I.J.; Pors, S.E.; Bager Skjerning, R.J.; Nielsen, S.S.; Bojesen, A.M. Immunogenic and protective efficacy of recombinant protein GtxA-N against Gallibacterium anatis challenge in chickens. Avian Pathol. 2015, 44, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Pors, S.E.; Skjerning, R.B.; Flachs, E.M.; Bojesen, A.M. Recombinant proteins from Gallibacterium anatis induces partial protection against heterologous challenge in egg-laying hens. Vet. Res. 2016, 47, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, I.J.; Persson, G.; Bojesen, A.M.; Pors, S.E.; Skjerning, R.B.; Thøfner, I.C.N. Outer membrane vesicles of Gallibacterium anatis induce protective immunity in egg-laying hens. Vet. Microbiol. 2016, 195, 123–127. [Google Scholar]
- Linhartová, I.; Bumba, L.; Mašín, J.; Basler, M.; Osička, R.; Kamanová, J.; Procházková, K.; Adkins, I.; Hejnová-Holubová, J.; Sadílková, L.; et al. RTX proteins: A highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, B.M.; Frees, D.; Bojesen, A.M. GtxA from Gallibacterium anatis, a cytolytic RTX-toxin with a novel domain organisation. Vet. Res. 2010, 41, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristensen, B.M.; Frees, D.; Bojesen, A.M. Expression and secretion of the RTX-toxin GtxA among members of the genus Gallibacterium. Vet. Microbiol. 2011, 153, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Klimentova, J.; Stulik, J. Methods of isolation and purification of outer membrane vesicles from gram-negative bacteria. Microbiol. Res. 2015, 170, 1–9. [Google Scholar] [CrossRef]
- Musante, L.; Tataruch, D.; Gu, D.; Benito-Martin, A.; Calzaferri, G.; Aherne, S.; Holthofer, H. A simplified method to recover urinary vesicles for clinical applications, and sample banking. Sci. Rep. 2014, 4, 7532. [Google Scholar] [CrossRef]
- Antenucci, F.; Fougeroux, C.; Bossé, J.T.; Magnowska, Z.; Roesch, C.; Langford, P.; Holst, P.J.; Bojesen, A.M. Identification and characterization of serovar-independent immunogens in Actinobacillus pleuropneumoniae. Vet. Res. 2017, 48, 74. [Google Scholar] [CrossRef] [Green Version]
- Bager, R.J.; Kudirkiene, E.; da Piedade, I.; Seemann, T.; Nielsen, T.K.; Pors, S.E.; Mattsson, A.H.; Boyce, J.D.; Adler, B.; Bojesen, A.M. In silico prediction of Gallibacterium anatis pan-immunogens. Vet. Res. 2014, 45, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blundell, E.L.C.J.; Vogel, R.; Platt, M. Particle-by-Particle Charge Analysis of DNA-Modified Nanoparticles Using Tunable Resistive Pulse Sensing. Langmuir 2016, 32, 1082–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bojesen, A.M.; Nielsen, S.S.; Bisgaard, M. Prevalence and transmission of haemolytic Gallibacterium species in chicken production systems with different biosecurity levels. Avian Pathol. 2003, 32, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Pauly, D.; Chacana, P.A.; Calzado, E.G.; Brembs, B.; Schade, R. IgY technology: Extraction of chicken antibodies from egg yolk by polyethylene glycol (PEG) precipitation. J. Vis. Exp. 2011, 51, e3084. [Google Scholar] [CrossRef]
- Bitto, N.; Kaparakis-Liaskos, M. The Therapeutic Benefit of Bacterial Membrane Vesicles. Int. J. Mol. Sci. 2017, 18, 1287. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Jang, H.; Nho, S.W.; Cha, I.S.; Hikima, J.; Ohtani, M.; Aoki, T.; Jung, T.S. Outer Membrane Vesicles as a Candidate Vaccine against Edwardsiellosis. PLoS ONE 2011, 6, e17629. [Google Scholar] [CrossRef]
- Bishop, A.L.; Schild, S.; Patimalla, B.; Klein, B.; Camilli, A. Mucosal immunization with Vibrio cholerae outer membrane vesicles provides maternal protection mediated by antilipopolysaccharide antibodies that inhibit bacterial motility. Infect. Immun. 2010, 78, 4402–4420. [Google Scholar] [CrossRef] [Green Version]
- Tsolakos, N.; Lie, K.; Bolstad, K.; Maslen, S.; Kristiansen, P.A.; Høiby, E.A.; Wallington, A.; Vipond, C.; Skehel, M.; Tang, C.M.; et al. Characterization of meningococcal serogroup B outer membrane vesicle vaccines from strain 44/76 after growth in different media. Vaccine 2010, 28, 3211–3218. [Google Scholar] [CrossRef]
- Hanly, W.C.; Artwohl, J.E.; Bennett, B.T. Review of Polyclonal Antibody Production Procedures in Mammals and Poultry. ILAR J. 1995, 37, 93–118. [Google Scholar] [CrossRef] [Green Version]
- Cronkite, D.A.; Strutt, T.M. The Regulation of Inflammation by Innate and Adaptive Lymphocytes. J. Immunol. Res. 2018, 2018, 1–14. [Google Scholar] [CrossRef]
- Uchiyama, T.; Kamagata, Y.; Yoshioka, M. Mechanism of lipopolysaccharide-induced immunosuppression: Immunological activity of B cell subsets responding to T-dependent or T-independent antigens in lipopolysaccharide-preinjected mice. Infect. Immun. 1984, 45, 367–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Giudice, G.; Podda, A.; Rappuoli, R. What are the limits of adjuvanticity? Vaccine 2001, 20, S38–S41. [Google Scholar] [CrossRef]
- Downey-Slinker, E.D.; Ridpath, J.F.; Sawyer, J.E.; Skow, L.C.; Herring, A.D. Antibody titers to vaccination are not predictive of level of protection against a BVDV type 1b challenge in Bos indicus—Bos taurus steers. Vaccine 2016, 34, 5053–5059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [Green Version]
- Hassan, J.O.; Curtiss, R., 3rd. Effect of vaccination of hens with an avirulent strain of Salmonella typhimurium on immunity of progeny challenged with wild-Type Salmonella strains. Infect. Immun. 1996, 64, 938–944. [Google Scholar] [CrossRef] [Green Version]
- Faulkner, O.B.; Estevez, C.; Yu, Q.; Suarez, D.L. Passive antibody transfer in chickens to model maternal antibody after avian influenza vaccination. Vet. Immunol. Immunopathol. 2013, 152, 341–347. [Google Scholar] [CrossRef]
- Al-Natour, M.Q.; Ward, L.A.; Saif, Y.M.; Stewart-Brown, B.; Keck, L.D. Effect of Different Levels of Maternally Derived Antibodies on Protection Against Infectious Bursal Disease Virus. Avian Dis. 2004, 48, 177–182. [Google Scholar] [CrossRef]
- Rauw, F.; Gardin, Y.; Palya, V.; van Borm, S.; Gonze, M.; Lemaire, S.; van den Berg, T.; Lambrecht, B. Humoral, cell-mediated and mucosal immunity induced by oculo-nasal vaccination of one-day-old SPF and conventional layer chicks with two different live Newcastle disease vaccines. Vaccine 2009, 27, 3631–3642. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Inoculum | Administered to Group | CFU/mL (Pre-Challenge) | CFU/mL (Post-Challenge) |
|---|---|---|---|---|
| 1 | 1 | 1, 2 | 0.92 × 107 | 0.8 × 107 |
| 2 | 3, 4 | 0.8 × 107 | 0.81 × 107 | |
| 3 | 5, 6 | 1.25 × 107 | 1.10 × 107 | |
| 2 | 1 | 1, 2 | 5.58 × 107 | 3.83 × 107 |
| 2 | 3, 4 | 5.79 × 107 | 4.9 × 107 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antenucci, F.; Arak, H.; Gao, J.; Allahgadry, T.; Thøfner, I.; Bojesen, A.M. Hydrostatic Filtration Enables Large-Scale Production of Outer Membrane Vesicles That Effectively Protect Chickens against Gallibacterium anatis. Vaccines 2020, 8, 40. https://doi.org/10.3390/vaccines8010040
Antenucci F, Arak H, Gao J, Allahgadry T, Thøfner I, Bojesen AM. Hydrostatic Filtration Enables Large-Scale Production of Outer Membrane Vesicles That Effectively Protect Chickens against Gallibacterium anatis. Vaccines. 2020; 8(1):40. https://doi.org/10.3390/vaccines8010040
Chicago/Turabian StyleAntenucci, Fabio, Homa Arak, Jianyang Gao, Toloe Allahgadry, Ida Thøfner, and Anders Miki Bojesen. 2020. "Hydrostatic Filtration Enables Large-Scale Production of Outer Membrane Vesicles That Effectively Protect Chickens against Gallibacterium anatis" Vaccines 8, no. 1: 40. https://doi.org/10.3390/vaccines8010040