
DNA Vaccines: Their Formulations, Engineering and Delivery

1
The Jake Gittlen Laboratories for Cancer Research, College of Medicine, Pennsylvania State University, Hershey, PA 17033, USA
2
The Department of Pathology and Laboratory Medicine, College of Medicine, Pennsylvania State University, Hershey, PA 17033, USA
*
Authors to whom correspondence should be addressed.
Vaccines 2024, 12(1), 71; https://doi.org/10.3390/vaccines12010071
Submission received: 13 December 2023
/
Revised: 2 January 2024
/
Accepted: 8 January 2024
/
Published: 11 January 2024
(This article belongs to the Special Issue Feature Papers of DNA and mRNA Vaccines)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The concept of DNA vaccination was introduced in the early 1990s. Since then, advancements in the augmentation of the immunogenicity of DNA vaccines have brought this technology to the market, especially in veterinary medicine, to prevent many diseases. Along with the successful COVID mRNA vaccines, the first DNA vaccine for human use, the Indian ZyCovD vaccine against SARS-CoV-2, was approved in 2021. In the current review, we first give an overview of the DNA vaccine focusing on the science, including adjuvants and delivery methods. We then cover some of the emerging science in the field of DNA vaccines, notably efforts to optimize delivery systems, better engineer delivery apparatuses, identify optimal delivery sites, personalize cancer immunotherapy through DNA vaccination, enhance adjuvant science through gene adjuvants, enhance off-target and heritable immunity through epigenetic modification, and predict epitopes with bioinformatic approaches. We also discuss the major limitations of DNA vaccines and we aim to address many theoretical concerns.
Keywords:
DNA vaccine; vector; adjuvant; delivery; TH1; TH2; immune responses; innate immunity; adaptive immunity; integration; tolerance; inflammatory; gene therapy1. Introduction
The advent of cell culture in the early 1900s [1], the development of the electron microscope in the early 1930s [2,3], the discovery of DNA structure and the central dogma in the 1950s [4] and sequencing technology in the 1970s [5] have contributed significantly to the development of vaccines for numerous previously morbid and mortal pathogens [6,7,8].
The most salient vaccine technology of the 21st century is gene vaccination. Gene vaccines work by harnessing advances in biochemistry, molecular biology, genetics and chemistry to deliver selected portions of the deoxyribonucleic acid (DNA), or messenger ribonucleic acid (mRNA), of the pathogen of interest. In theory, the engineering can eliminate any pathogenicity, thus reducing the morbidity of vaccination and obviating any mortality of vaccination owing to inoculum pathogenesis [9,10].
DNA vaccines, first developed in the 1990s [11], are scalable, stable, easily manipulable and amenable to stockpiling [12]. They produce comprehensively robust adaptive immune responses comparable to those seen in attenuated pathogen vaccines with the safety profile of a subunit vaccine and can so can be given to immunocompromised persons [13]. The earliest gene vaccines consisted only of DNA as plasmids, which were taken up by cells via different delivery methods, transported to the nucleus, transcribed to mRNA and then translated into proteins, which are processed to antigens that stimulate immune responses [11,12].
The mRNA vaccine is technically different, because mRNA is not packaged into a plasmid; however, the mRNA vaccine offers significant advantages over the DNA vaccine, the two most important advantages being that the antigen to be presented is post-RNA-processing and integration into the host genome is substantially less likely [14,15].
While gene vaccination is not a household term, ‘mRNA vaccine’ certainly is owing to the incredible successes of two mRNA vaccines during the SARS-CoV-2 global pandemic [16,17]. DNA and mRNA vaccines are both gene vaccines, and thus fundamentally converge at the point of immunogenic protein translation within the cell; however, DNA and mRNA vaccines are not entirely alike—a DNA vaccine does not simply become an mRNA vaccine when the nucleic acid sequence enters a cell and undergoes transcription, because the processing of DNA to RNA in vivo includes some degree of variability at the local (cell-specific) level [18], and RNA translation, likewise, is subject to local factors [19].
DNA vaccines work by delivering antigens in their genetic form, as DNA plasmids, to cells. Both DNA and mRNA vaccines need to cross the cell membrane, and, as hydrophilic, polar and charged molecules, they do not freely cross the cellular membrane. DNA must additionally cross the nuclear envelope to the nucleus to be transcribed to mRNA before the antigenic protein can be translated in order to stimulate an immune response. mRNA vaccines are post-processing, do not need to cross the nuclear envelope and can be immediately translated to the antigenic protein once in the cytoplasm [20].
DNA vaccines were first put forth in the early 1990s when it was discovered that intramuscular plasmid DNA injection could induce protein expression, and importantly, it was concurrently shown that no special delivery system was necessary for most cells to take up DNA and route it to the nucleus [21]. Shortly after, this idea was adapted to the delivery of antigens of interest [22], and it has since been shown that naked DNA delivered to the cell results in relatively low and inconsistent gene expression [23,24], so newer formulations were adapted and developed to increase expression. Technological advances have made the viral vector one of the platforms of choice for DNA delivery by placing the gene of interest within the genome of a modified virus [25]. The virus is then taken up by cells, in much the same way a viral infection would occur, after which the replicative property of the virus is harnessed for the expression of the delivered gene sequence [26].
It would be more than thirty years from discovery until the first DNA vaccine was approved for use in humans: the Indian ZyCovD [27,28,29], which is a vaccine against SARS-CoV-2 [27]. The first DNA vaccine to make it to a phase I clinical trial was an HIV-1 vaccine in the late 1990s [30]. While the HIV vaccine never made it to market, the proof of concept was satisfied and many of the safety concerns about the technology were tempered by the trial [31]. One of the principle hurdles to effective HIV vaccination is the rapid rate of mutation of HIV, a retrovirus with a famously error-prone reverse transcriptase and short generation time [32,33,34,35]. Mutating pathogens are a moving target for vaccination, and this is one of the principle reasons for the success of the COVID-19 vaccines: the spike protein is highly conserved [36]. As of today, hundreds of DNA vaccines are in active clinical trials, or have completed clinical trials, for conditions including: influenza, zika, breast cancer, HIV, prostate cancer, malaria, HSV, melanoma, hepatitis and others [37].
Had it not been for the pandemic, a different DNA vaccine would likely have made it to market first. Although, which vaccine this might have been is difficult to answer; several technical improvements have contributed to a renewed interest and expanded promise. These include more effective delivery approaches, adjuvant advancements, promoter and gene optimization strategies and improved nucleic acid design [30]. DNA vaccines are in wide use in veterinary medicine and many of the technological and engineering advances in trial today have come from veterinary research [12,38]. Human applications of the technology have lagged, largely due to sub-optimal immunogenicity when compared to conventional vaccine approaches [39]. Many technological advances have brought DNA vaccination forward, and more is yet to come.
DNA vaccines in clinical trials, and through clinical trials, are summarized in other reviews [40,41,42,43,44,45,46]. This review will focus specifically on DNA vaccine technology, including its development, applications and emerging technologies. However, many of the technologies being developed can also be readily applied to mRNA vaccines.
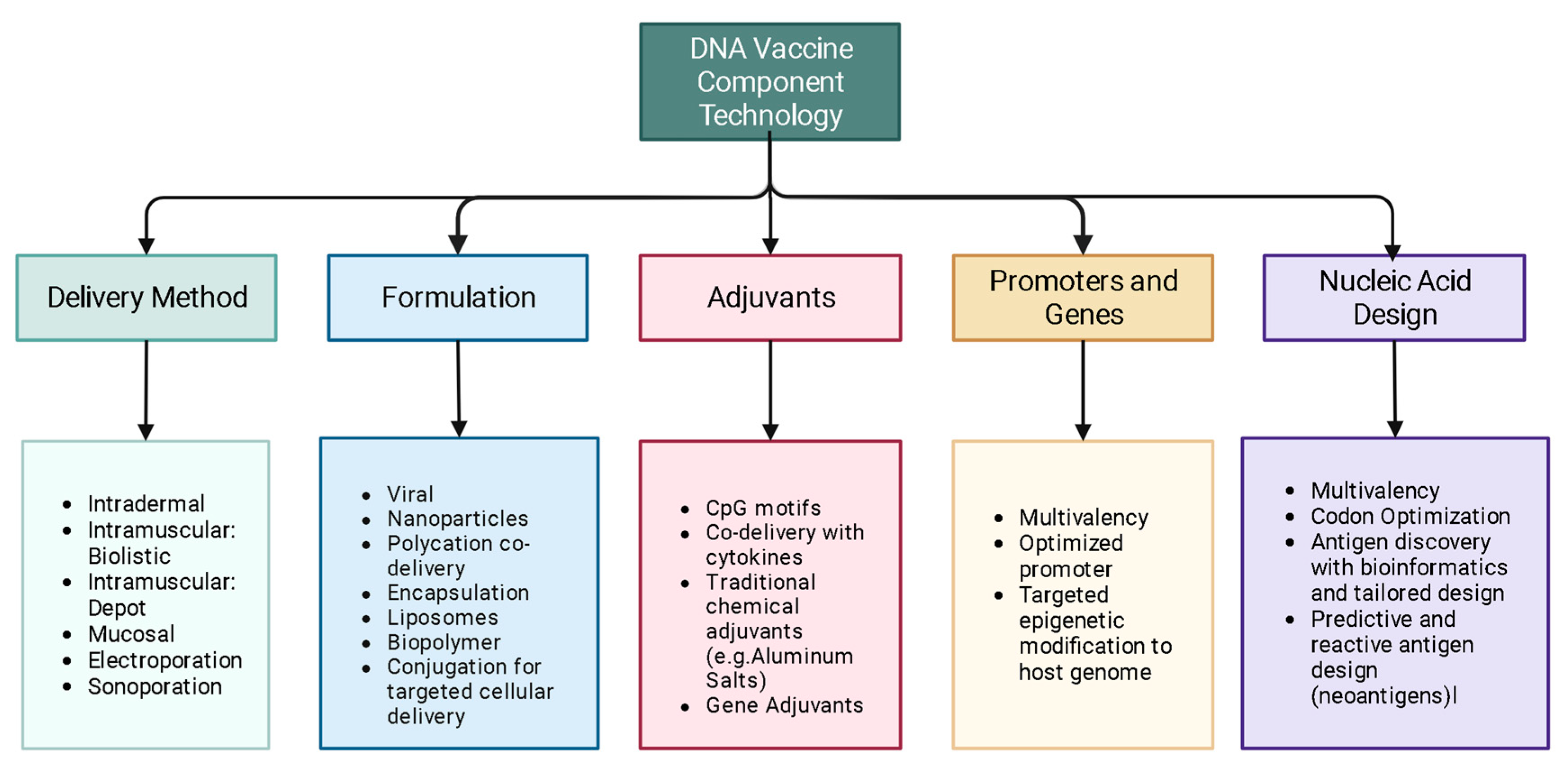
Some of the emerging science in the field of DNA vaccines, covered in this review, aims to: optimize delivery systems, better engineer delivery apparatuses, improve delivery techniques, personalize cancer immunotherapy through DNA vaccination, enhance adjuvant science through gene adjuvants, enhance off-target and heritable immunity through epigenetic modification, as well as to model and predict epitopes with bioinformatic approaches (Figure 1).
2. Mechanism
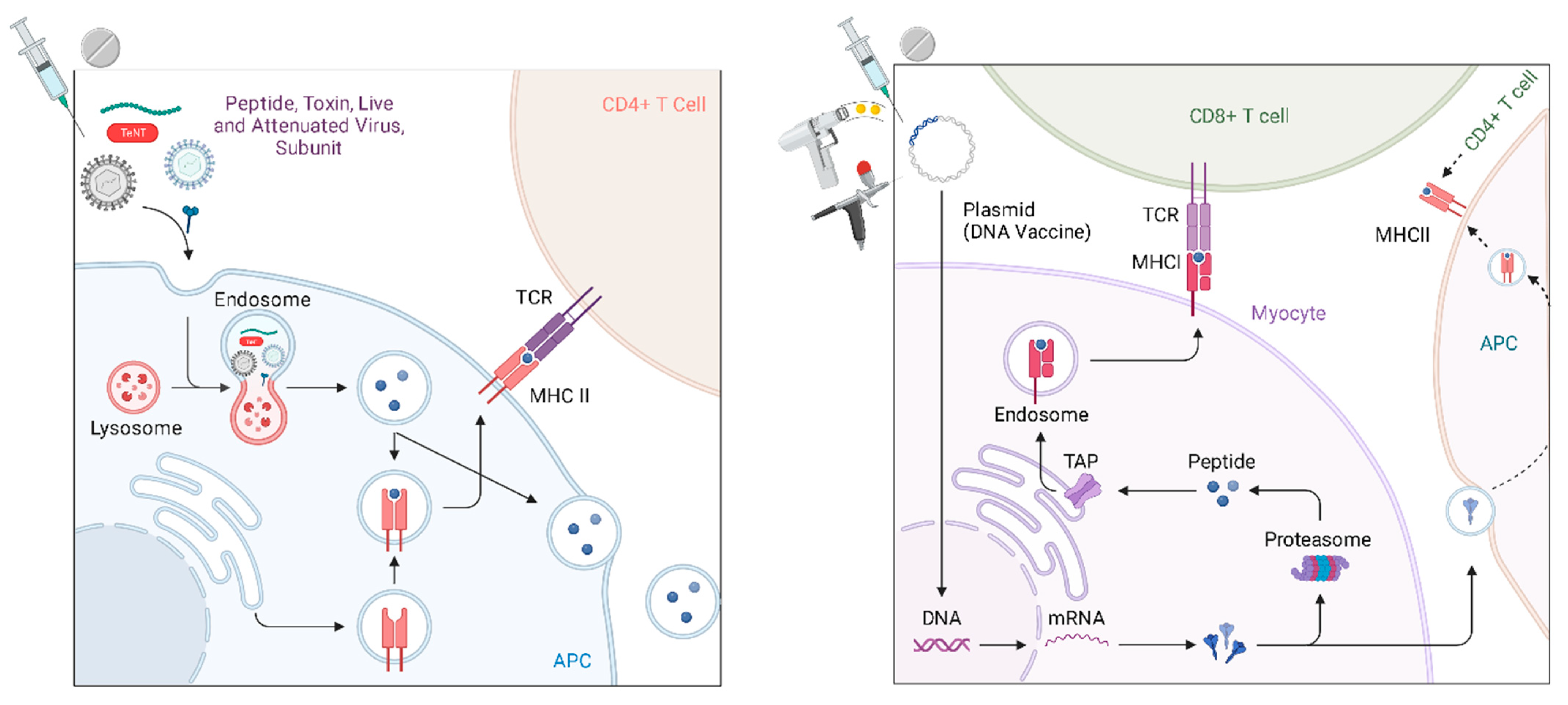
Like all vaccines, the mechanism by which DNA vaccines generate immunogenicity is by activating the adaptive immune response. When a plasmid is delivered near to a cell and then taken up (either passively or by facilitation), the plasmid DNA can then be recognized and expressed by the native cellular machinery to generate the target antigen within the cell that has taken it up. From there, antigens (usually varying lengths of peptides) are presented on the cell surface for interaction with the immune cells by one of two pathways, either the major histocompatibility complex (MHC) class I (MHC I) or class II (MHC II) pathways (Figure 2). MHCI, which is present in all nucleated cells [47], is most frequently thought to be the presentation mechanism for endogenous antigens (most commonly peptides), while MHCII is thought to be the classical pathway for the expression of exogenous antigens, such as bacteria, fungi, protozoa and free viruses that the cell has endocytosed [48,49,50]. Because plasmids are taken up by the cell and the antigen to be presented is then generated intracellularly through the transcription and translation of the delivered DNA, the most common mechanism of the antigen presentation in DNA vaccination is MHC Class I [51,52].
Furthermore, MHC Class I presentation is associated with the activation of CD8 T lymphocytes; CD8 serves as an adhesion molecule for MHC I [52,53,54]. In sequence, DNA vaccines generate intracellular peptides, which are, in this pathway, endogenous proteins, and are most commonly presented on MHCI and activate CD8 T cells. Concurrently, the translated proteins are also exocytosed and subsequently taken up by other antigen presenting cells, which drain to regional lymph nodes [55]. These include macrophages, dendritic cells and other monocytes. These antigen presenting cells (APCs) endocytose and/or phagocytose materials from the extracellular environment and then process and present by MHCII, which preferentially activates CD4 T cells. Two types of CD4 T helper cells (Th1 and Th2) can further activate CD8 T cells or promote B cell differentiation to plasma cells to generate an humoral immune response, respectively [55]. The relative balance of both cytotoxic and humoral immunity [56] sets gene vaccines apart from other vaccination technologies, that have historically relied more heavily on humoral responses and have generated attenuated cytotoxic effects by antigen cross-presentation on MHCI [57] without necessarily ensuring a robust cell-mediated response [52,58]. Additionally, plasmid DNA has historically been derived from bacteria. As DNA, and because of its fundamental design, plasmids can have a number of CpG motifs. A CpG motif is a known adjuvant that activates TLR9-bearing B cells and plasmacytoid dendritic cells to promote strong Th1-type immune responses and these CpG motifs can be optimally engineered into DNA vaccines [59,60,61]. Likewise, viral vectors are chosen for their intrinsic immunostimulatory effects and for their facility in transporting their genomic cargo into cells.
The immune responses generated by a vaccine are an essential consideration. The specific T cell and B cell activation is especially crucial when determining a vaccine’s effectiveness. T cells have T-cell receptors (TCRs) that recognize processed antigen fragments presented on MHC by other cells [62], either cells that have endocytosed the antigen (bacteria) or cells that are producing the antigen (tumors or virally infected cells). These T cells need to differentiate between self and foreign peptides, and are only activated upon MHC presentation of foreign epitopes [63]. Regarding vaccination, T cells respond to a cell that is infected to primarily eliminate infected cells. So, T cells are primarily reactionary in the scope of a pathogenic invasion. On the other hand, B cells have receptors, specific to an epitope, which can bind to extracellular antigens. And, the B cell receptor has the same specificity as the antibodies that the terminally differentiated plasma cell secretes [62]. In conventional peptide, subunit, or inactivated pathogen vaccinology, antibody titers have been the measure of efficacy and a surrogate for preventative protection [64].
Plasma cells secrete neutralizing antibodies against the pathogen of interest, resulting in elimination and prevention. Some of these plasma cells then become memory plasma cells that surveil and prevent re-infection by rapid reactivation upon re-exposure to the antigen. This is to say that conventional vaccines have focused quantitatively on the humoral response [65] as a measure of prevention and protection with acknowledgment that the cellular response mostly contributes to infection control [64]. The cellular response is much better understood today than when antibody titers were first drawn, and it is known that most viral clearance relies heavily on cytotoxic function [66], which has informed the direction of current vaccine development [52,67,68,69,70,71]. The DNA vaccine has shown its ability of inducing strong T cell-mediated immune responses, which is crucial for eliminating infected cells as well as for anti-tumor effects [72]. Interestingly, the delivery method may direct the immune responses for DNA vaccination. For example, the proliferation of CD4 T helper cells toward Th1 or Th2 has been shown to be somewhat directable, with intradermal (ID) delivery suggested to preferentially induce the Th1-immune response by recruiting more APCs and inducing a more polyfunctional cellular immune response, while intramuscular (IM) delivery results in a stronger humoral, or Th2-mediated, immune response with an advantage in antigen expression [65,73,74,75,76]. However, an argument has been made that the delivery method (e.g., gene gun vs. IM depot) is likely to be a more directive factor for the DNA vaccine [75,77]. But, the issue seems to be only supplementary, because adjuvants are also capable of specific stimulatory effects directing CD4+ T cells toward either, or both, Th-1 and Th-2 [78].
Lastly, unlike injected protein or subunit antigens, which have a comparatively short half-life [23,79,80], plasmid DNA can provide tissue expression of antigens over much longer periods of time, potentially offering a better priming of the immune system [39]. Interestingly, in conventional vaccine development, DNA is considered a contaminant since a DNA plasmid is often used to generate a protein or to knock a gene into a bacterium but the plasmid and/or DNA is then intentionally removed in the final vaccine product [81].
3. Design
Conventional, non-nucleic acid, vaccine platforms rely on attenuated and/or inactivated pathogens or components of pathogens. The idea of epitope-based vaccine engineering was first put forth in 2014 [82]. Gene vaccines allow the antigen to be designed within the parameters of the intrinsic biochemical machinery; the antigen is customizable within this framework. For most existing pathogens, a target antigen is chosen with an eye towards an antigen that is easily translated and convenient to manufacture, of which the ‘spike’ protein of the SARS-2 coronavirus is an example [83]. However, the antigen selection process can be optimized, and recent advances in both computing power and global collaborative efforts have generated repositories of antigen structure, peptide sequence and nucleic acid sequence, in order to accelerate vaccine development [84,85,86,87,88,89]. The National Institute of Health (NIH) has developed a number of these repositories through the National Center for Biotechnology Information (NCBI) while other repositories are hosted by academic and corporate organizations. Additionally, epitopes can be predicted by simulation, and this offers expedient development for emerging pathogens [84].
Immunotherapy is the principle of harnessing the body’s immune system to treat disease, including cancer [90]. Chimeric antigen receptor T (CAR-T) cells [91,92], as an example, have revolutionized the treatment of hematologic malignancy. One of the most exciting applications of DNA vaccines is immunotherapy. Other anti-cancer vaccines exist, but to date these are all preventative cancer vaccines. The widely used vaccines against HPV, a known cause of cervical and oropharyngeal cancers, prevent viral infection, thus preventing HPV-associated cancers [93]. However, the cancer vaccine in the immunotherapy realm more commonly refers to vaccination given against an incident cancer, a therapeutic cancer vaccine [13,43].
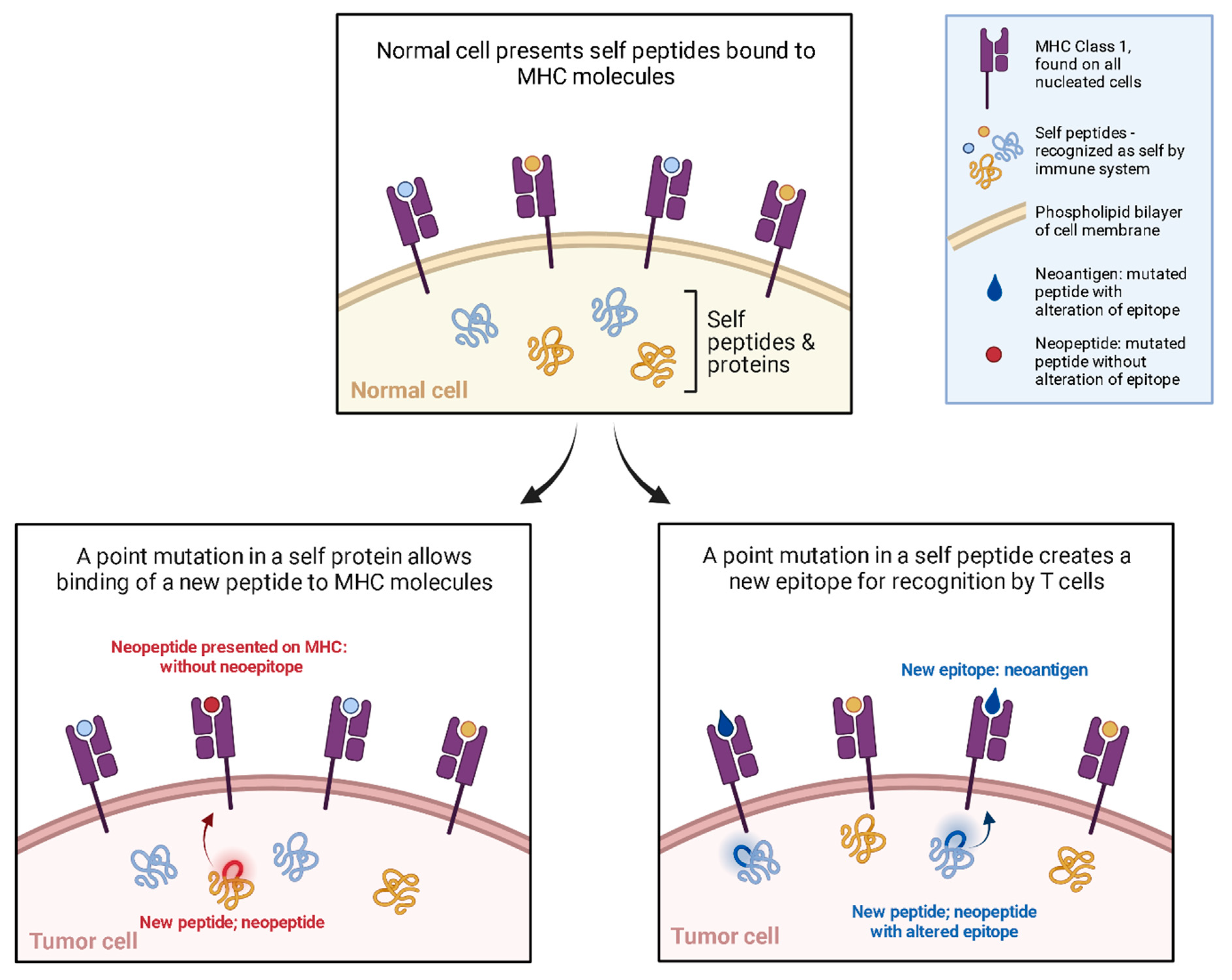
Immuno-oncology research now includes DNA vaccines. In contrast to the traditional DNA vaccine targeting infectious pathogens, DNA cancer vaccines target cancer cells to induce a generalized as well as a tailored immune response. Cancer cells survive by evading the immune system, this is termed the tolerogenic effect [94]. The most famously shown mechanisms are CTLA-4 and PD-1 [95]. However, cancer cells are constantly forming new peptides by means of genomic mutation, dysregulated RNA splicing, disordered post-translational modification, and integrated viral open reading frames [96]. These new peptides are called neopeptides (Figure 3); some of these neopeptides are ultimately presented on MHC in such an orientation as to present a novel portion of this neopeptide, resulting in a neoantigen. It is these neoantigens that offer a potential target for immunotherapy. The advances in sequencing, and the aforementioned explosion in bioinformatics offer potential personalized anti-tumor immunotherapy targets [97,98].
In the cancer-immunity cycle [99], it is the cytotoxic T cells which provide the greatest anti-tumor effects. While many solid tumors are resectable, metastases and diffuse tumor burdens are often beyond surgical options. While it has been shown that natural killer (NK) cells are more effective for metastatic cancers than cytotoxic T cells are, the addition of intrinsically generated, specific, cytotoxic T cells by immunotherapy offers a promising addition to the repertoire. Many hurdles remain, including waning immunity with age, immunosuppressive treatment regimens and the rapid mutation of tumor cells [100].
In cancer vaccine practice, a tumor cell sample is gathered and the somatic mutations from the whole genome or exome are identified. From there, experiments and algorithms help to identify the most suitable ‘neopeptides’, and the appropriate DNA sequences are loaded in to a viral vector or the gene is loaded in to a plasmid to be injected into the patient [101]. This is the framework in which ‘personalized’ DNA vaccines [102] are presented and the idea behind anti-tumor vaccines [96,97,103], many of which are currently in clinical trial [104]. Similarly, known tumor features and markers can be targeted, such as ERBB2 in some breast cancers [40].
Existing immunotherapies, like checkpoint inhibition, may also benefit from this additional neoantigen DNA vaccine against cancer cells in much the same way that highly active antiretroviral therapy (HAART) works as a cocktail therapy; to not only reduce the viral load in HIV patients but also to reduce the likelihood of resistance to any individual therapy [105,106,107].
Epigenetic changes to oncogenes, proto-oncogenes, tumor suppressor genes and other related DNA sequences are known causes of cancers [108,109]. Vaccines have been shown to have heterologous, or ‘non-specific’ effects, which are additional effects beyond the specific protection against the targeted disease [110,111,112]. While the exact mechanisms are only beginning to be understood, there is emerging evidence that epigenetic mechanisms are involved [113], including DNA imprinting, as has been shown in HIV and BCG vaccine patient analyses [114,115]. The data from the BCG cohorts indicates that many of these ‘non-specific’ effects can be especially useful, having proven benefit in protecting against autoimmune diseases including multiple sclerosis and type 1 diabetes. Interestingly, the protection conferred accumulates gradually over years [115,116,117]. This is especially interesting in light of recent evidence for the transmission of trained immunity in mammals linked to the epigenetic modification of cell lines, termed, ‘trained immunity’ [118].
In principle, epigenetic effects hold promise; however, concerns abound. DNA interaction, especially plasmid integration, is the most feared consequence of DNA vaccination [119]. Although integration has been proven to be exceedingly rare [119,120,121], it is poignant for its catastrophic potential. While DNA integration is feared, activity above the genome may yet prove useful with directed action. This is still an emerging field, and no DNA vaccines to date intentionally target epigenetic changes.
DNA vaccines are, like all DNA, identifiable and functional based on the sequence of nucleic acids. The deployment of the desired antigen necessarily must consider the conveyance apparatus, such as a plasmid or viral vector, as well as diffusion facilitators and adjuvants [122], while also considering the route of administration. The size of the DNA sequence is an important consideration as well. Meanwhile, some small, non-polar molecules (about 40 kDa) likely can diffuse freely across the nuclear envelope [123]. However, for larger molecules (AAV viral proteins are all larger than 60 kDa [124]) the most important mechanism for larger nuclear translocation involves nuclear pore complexes (NPCs) [24,125,126,127].
In theory, a DNA vaccine consists of the DNA sequence encoding the potential antigen under the control of a eukaryotic promoter to drive gene expression in the host cell. To date, most DNA vaccines have been bacterial plasmids engineered for gene expression in eukaryotic cells by the selection of appropriate promoter and termination sequences, as well as a nuclear localization signal [128,129]. Plasmids used experimentally, or therapeutically, to deliver gene products are called plasmid vectors. In addition to the gene product, these plasmids also contain an enhancer, a promoter, a transcription termination and/or a polyadenylation signal sequence, and an origin of replication. Additionally, plasmids may contain an antibiotic-selectable marker to enable growth and manipulation in bacterial cells and a marker sequence to identify transfected cells [130]. Engineering plasmids for optimal transfection efficacy and immunogenicity has shown that the promoter is an especially important consideration. Virally derived promoters have been shown to provide greater gene expression than other eukaryotic promoters and, in particular, the CMV promoter, have often been shown to direct the highest level of transgene expression compared with other promoters [131].
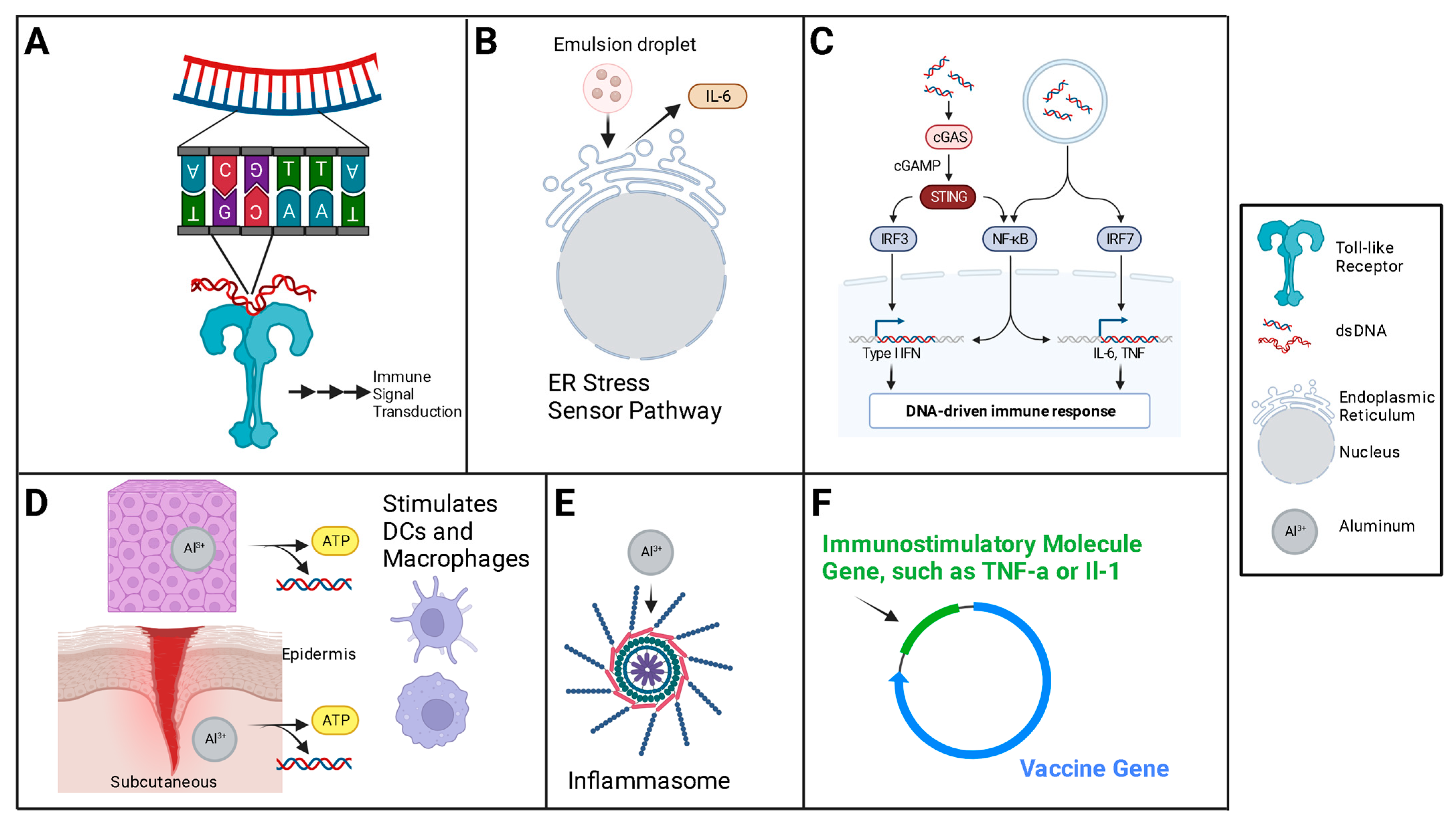
Natural elements of plasmid DNA are also important potentiators and modulators of the innate immune response preemptory of the intended, adaptive response. The innate immune system uses pattern-recognition receptors to sense pathogens and then induce the downstream production of interferons and cytokines. One such example is the Toll-like receptor-9 (TLR9), a cytosolic receptor, that binds DNA sequences containing unmethylated cytosine-guanine (CpG) motifs, leading to the activation of signaling pathways [132].
Viral vectors were first elucidated in the 1970’s as a way to deliver new genetic material to cells [133]. Naturally, the technology has been deployed in basic science and gene therapy, especially where integration is the goal. When a chosen gene is engineered in to the viral genome, it is considered recombinant. The main classes of viral vectors (including RNA and DNA, single stranded and double-stranded) can be categorized into two groups according to whether their genomes integrate into the host (retroviruses and lentivirus) or persist in the cell nucleus predominantly as extrachromosomal episomes (adeno-associated virus, adenovirus, and herpes viruses) [134,135]. Additionally, each type of vector has characteristics which make it’s use more or less desirable in a given situation, AAV vectors carry a small ssRNA genome, are not very immunogenic but can only accommodate about 5 kilobase (kb) inserts which can integrate in to the genome [136,137,138]. Naked dsDNA adenoviruses possess a larger packaging capacity of about 7.5 kb of foreign DNA, and possess the endogenous virus nuclear localization signals [129]. Importantly these do not integrate and result in short-term episomal expression of the gene of interest in a relatively broad range of host cells, which can be targeted to specific cell populations with structural modifications. The original adenovirus vectors generated strong immune responses, whereas the so-called gutted second and third generation vectors containing deletions have proven to elicit substantially reduced immunogenicity [139], which may be good in gene therapy, but in vaccines the viral capsid immunogenicity is considered advantageous for the early, innate immune priming of the immune system. Previous exposure and immunity to the vector viruses can inhibit vector efficacy. Comprehensive reviews of specific viral vectors exist [134].
The first ZyCov-D, the first DNA vaccine approved for human use, is a plasmid vector [29]. However, both plasmid and viral vector DNA vaccines are currently in pre-clinical or clinical trials [25,140,141].
The engineering of the nucleic acid sequence is a source of continued advancement. The improved optimization of the promoter sequence can increase the production of antigens, resulting in a more robust immune response. Likewise, codon optimization, the selection of codon triplets that have the highest tRNA utilization frequency in the cytoplasm, can increase translation rates and mRNA stability [12]. Additionally, DNA vaccines can easily be made multivalent, simply by adding additional antigenic genes to the vector, and this has been shown to enhance their efficacy [12,142]. Likewise, an alternative approach involves co-injecting with adjuvant gene vectors [30].
Differentiating Infected from Vaccinated (DIV) strategies are widely used in animal vaccination campaigns to assort herds in disease control and eradication. Previously called ‘marker vaccines’, these are typically specific protein antigens not present in the natural pathogen, so that an assayable result indicates the animal has been vaccinated as opposed to infected [143,144]. DNA vaccines are perfectly situated for the adoption of ‘markers’, whereby an additional gene product can be used to screen for vaccination, assisting in both the long-term research of vaccination success while also providing readily identifiable vaccination history when uncertain.
4. Delivery
Viral vectors often harness the existing pathogenicity, facilitated uptake and existing affinity for permissive cell attachment inherent to the virus [25]. Modifications can be made to the capsid by the attachment or removal of ligand molecules. Because viruses naturally infect cells, most of the molecular engineering concerns plasmid delivery. Optimizing the route of the administration of the vaccine, be it oral, intramuscular or subcutaneous, applies to all formulations.
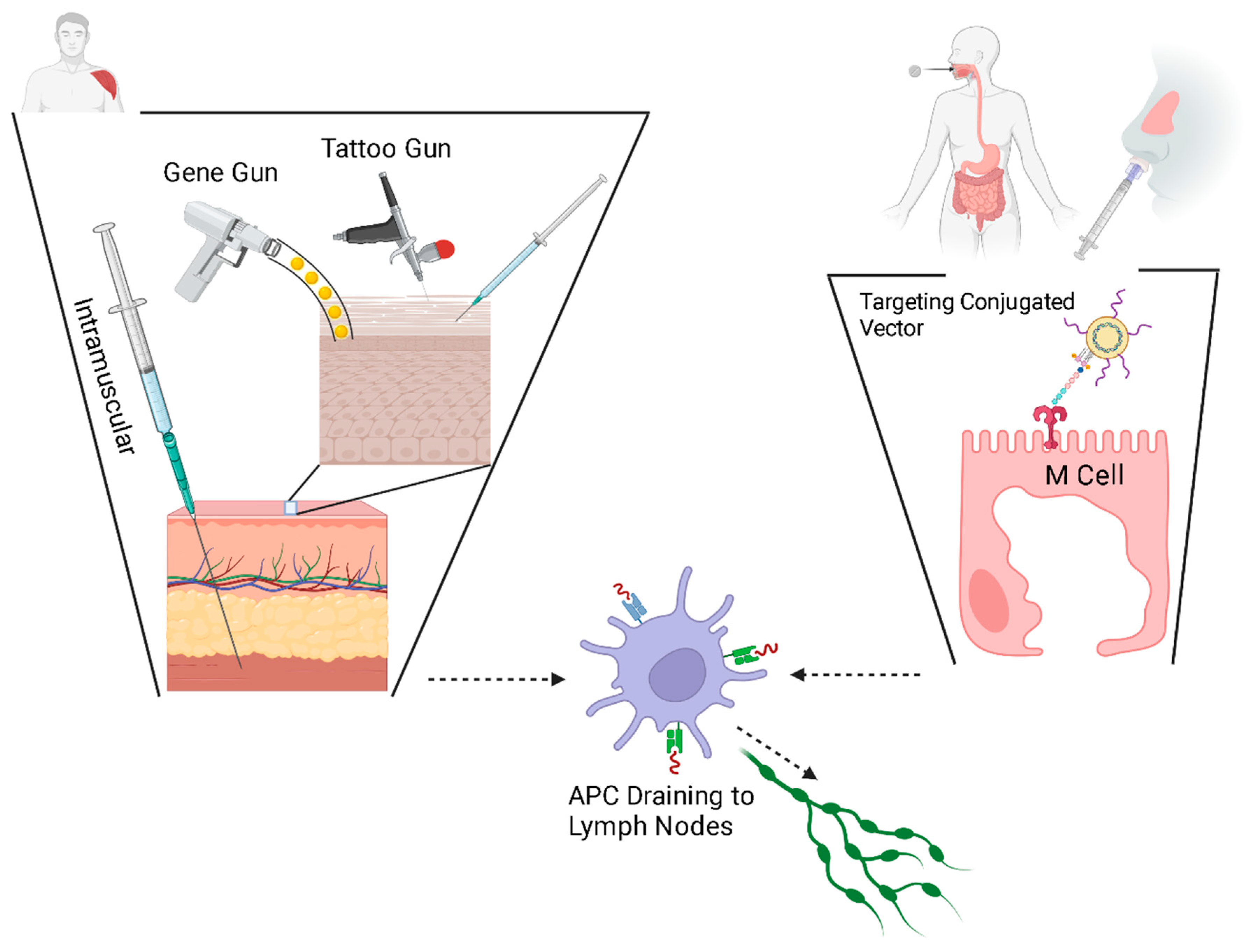
Interestingly, the seminal DNA vaccine research employed a technology that is still in use today: the biolistic system, also known as the ‘gene gun’ [22]. The earliest DNA vaccines, small plasmids, resulted in relatively low immunogenicity, and this is believed to have been the result of limited plasmid uptake [42]. Several possible reasons have been theorized and substantial work has been done to develop newer delivery methods and to enhance transfection. Physical methods of delivery are thought to be one method to increase the transfection efficiency of DNA vaccines. These methods include particle bombardment and high-pressure delivery, dermal patches (the microneedle array), epidermal tattoo and electroporation (EP).
The particle bombardment, or ‘gene gun’, approach uses a highly pressurized stream of microscopic heavy metal beads coated with DNA vaccines, similar to the biolistics of the earliest DNA vaccine, to deliver DNA plasmids, linked to microscopic, coated, gold particles [145,146,147] into the skin in a dry powder formulation. High-pressure delivery is conceptually similar to particle bombardment by forcing vaccines in liquid form through a tiny opening to create a fine, high-pressure stream that penetrates the skin [6], without the need for a needle stick [148]. Ultrasound is also being developed for a needleless delivery system through a process called ‘cavitation’, whereby openings are generated in the dermis, through which the vaccine can be delivered without a needle stick [149].
Another concept under investigation, tattoo gun delivery, harnesses the unique properties of the immune system within the epidermal layer. While intradermal vaccination dates to Jenner’s smallpox vaccine [150,151] and is not new, the epidermal idea has increased the deliverable dose by expanding the delivery area to a larger surface by moving across the epidermis [152]. Tattooing creates a dermal and epidermal inflammatory trauma and thus an inflammatory response [152]. Harnessing the natural inflammatory response as the optimal time to deliver plasmids has demonstrated robust immunogenicity [153,154].
At the same time as delivery, electroporation works by delivering short electrical impulses, which are thought to disrupt the plasma membrane of cells to facilitate plasmid uptake [153,155]. Likewise, ‘sonoporation’ uses ultrasound waves to generate openings in the cell membrane [156,157].
New research seeks to target specific cells and tissues to protect the nucleic acid vaccine against degradation. Molecularly, lysosomal inhibitors have been shown to protect nucleic acid vaccines [158]. Genetically, this can be achieved by encoding specific proteins, especially fusion proteins, to preferentially engage APCs, thus facilitating the immune response [159,160,161,162]. The mucosal delivery of the DNA vaccine has been hampered by limited vaccine uptake [163,164]; mucosal M cells can be targeted by the conjugation of stimulatory and/or localizing ligands to selected viral vectors, allowing for specific uptake at mucosal surfaces [161]. This method of conjugation can be applied to any conveyance method, so that specific stimulatory molecules can be attached to liposomes or to nanoparticles as well. In a similar way, gene adjuvants, discussed later, are another convenient mechanism for stimulating local immune cells.
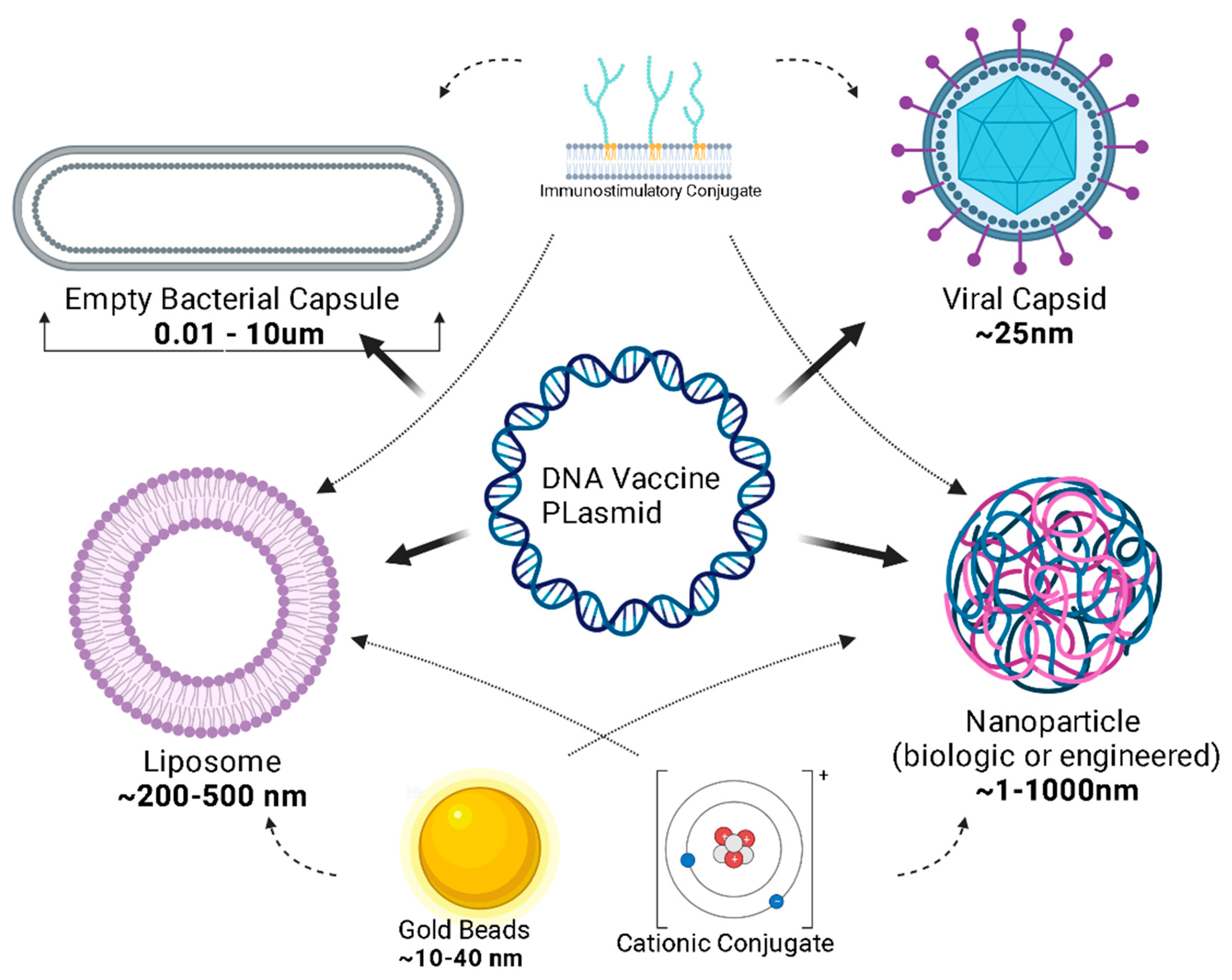
Plasmids must cross the phospholipid cellular membrane, avoid breakdown by cytosolic organelles and nucleases, and translocate across the nuclear envelope. Nanoparticles and viral vectors harness existing technology to optimize the delivery of the vaccine to its site of action, the nucleus [165]. Liposomes are a popular delivery mechanism because of their diffusion across the phospholipid membrane; liposomes also have adjuvant properties [165]. Like liposomes, engineered nanoparticles can be designed for lipophilicity and can be constructed to micrometer diameter sizes (Figure 4).
Nucleic acids carry a negative charge and DNA vaccine plasmids carry a net negative charge, and since most membranes of living cells are considered to have a resting negative charge, similar in principle to the idea of electroporation [153], it is likely that the attenuation of the charge or the masking of the charge will facilitate a better uptake of the plasmid [12,166]. Polycations have been trialed; however, encapsulation has proven, thus far, to be a better solution [12].
Before plasmid uptake, engineering the vaccine vector is a field of exciting growth. Previously, naked plasmids needed to be deposited in relatively large concentrations. Novel ideas about the delivery vector have expanded the scope and efficacy of DNA vaccines [167]. The typical DNA plasmid can be attached to a nanoparticle (<100 nm), which is a microscopic synthetic polymer such as starch, cellulose, silk, collagen, gelatin, albumin and chitosan (from chitin) chosen for biocompatibility, biodegradability, and low toxicity. In recent years, several studies have focused on the advances in this field [168]. Alternatively, cytoplasm-free, non-denatured Gram-negative bacterial cell envelopes, known as “bacterial ghosts’, are being explored for their inherent cell and tissue affinity, ease of production, and storage capabilities that do not require refrigeration, as well as their potential to inherently induce the innate immune system [39,167].
And, as mentioned before, the site of DNA vaccine delivery has been shown to affect the immune response and thus the efficacy of the vaccine [169].
5. Adjuvants
DNA is itself an adjuvant when exposed in the cytosol (Figure 5). The interplay of the many recognition molecules converges on a major molecule in the relay pathway to the innate immune response, the STING, or the stimulator of the interferon gene [171]. The two major classes of ‘exogenous’ adjuvants used for DNA vaccines are the traditional, chemical [10] adjuvants, and, second, the gene-encoded adjuvants—proteins encoded by the DNA vaccine or in plasmids delivered with the DNA vaccine.
Traditional adjuvants, such as aluminum salts and adjuvant systems, which are commonly used in conventional vaccines, activate the local innate immune system at the site of delivery and/or delay the removal of the vaccine from the site of delivery. Nucleic acid vaccine plasmids can be modified to include gene products in addition to the vaccine antigen. Genetic adjuvants, or gene adjuvants, are most commonly cytokine genes placed in the plasmid, or a separate plasmid, to be transcribed with the antigen [165,172,173,174]; other gene products are possible, with Toll-like receptor (TLR) ligands a common choice as well [175].
Plasmid DNA has historically been derived from bacteria. Because of its fundamental design, DNA plasmids have a number of CpG motifs. These are often unmethylated in non-mammals [39], and it is specifically unmethylated CpG which is immunostimulatory [176,177], capable of stimulating innate immune responses via the TLRs [178]. Specifically, the unmethylated CpG motif represents a pathogen-associated molecular pattern (PAMP), which is recognized by a specific pattern recognition system in the TLR-9 of antigen presenting cells [60,179]. Interestingly, TLR activation has been shown to have anti-tumor effects, and this synergy has inspired much of the cancer vaccine research [180].
Co-delivery with cytokines and/or cytokine genes, especially interleukin (IL) -2, -12 and -15 has been shown to augment the adaptive immune response by stimulating APCs [165,181]. Likewise, immunopotentiators, like TLR ligands, have been shown to similarly enhance the initial and long-term immune response after DNA vaccine delivery [132,182,183]. Traditional adjuvants, like aluminum-based compounds, still have a place in the DNA vaccine world, as do co-delivered CpG nucleic acid motifs [184,185,186], which are ironically mostly methylated in mammalian genomes (silencing genes) [187], but can be engineered into DNA plasmids unmethylated [39,188,189].
6. Limitations/Concerns
The great fear with DNA vaccines is genome integration; this could be a partial or complete plasmid genome integration, which can result in insertional mutagenesis, possibly in the activation of oncogenes or the inactivation of tumor suppressor genes [190]. The FDA designed guidelines for DNA vaccines, stating that the frequency of plasmid integration needs to be lower than the spontaneous mutation rate [81]. All of the data to date suggest that while integration can occur, it occurs at a frequency below the rate of spontaneous mutagenesis [120,121,191]. DNA plasmids can remain at the site of injection for months after injection, typically longer than RNA can remain, due to the protection afforded by double stranding [41]. DNA poses a greater risk than RNA for genome integration because RNA requires a reverse transcriptase to generate cDNA prior to potential integration. While most cells have a reverse transcriptase, it is not frequently expressed at very high levels [192,193]. The potential integration of the DNA vaccine remains a concern in the public consciousness [194,195,196], and we cannot discredit the stochastic evidence since the integration rate is additive to the spontaneous rate of mutation, which is, per gene, on the order of one mutation per one million cell divisions in humans [197,198]. Necessarily, a harmful mutation must be even less frequent. Nevertheless, published data overwhelmingly support the safety of DNA vaccination, the convincing example being the approval of the first DNA vaccine, ZyCovD [28]. Nonetheless, public concern is a serious impediment to gene vaccine uptake, as evidenced during the COVID pandemic [199,200,201].
Many approved vaccines including the Tetanus, Diptheria and acellular Pertussis (TDaP), and the Measles, Mumps, Rubella and Varicella (MMRV), have much higher (although still low) serious adverse event rates [202]. Public awareness campaigns as well as long-term COVID DNA vaccine safety experience and data will likely exonerate DNA vaccines in this regard, although this will need to be re-demonstrated with many of the newer technological advancements as alterations of the plasmid DNA, adjuvants and alternative delivery vehicles could theoretically affect the probability of integration.
Second, DNA plasmids are often developed in bacterial lines, and these lines are often selected for by inserting an antibiotic resistance gene into the plasmid. This has led to a concern about antibiotic resistance as a possible adverse effect of DNA vaccination should the antibiotic resistance gene be integrated [203]. This has not been shown in any trials to date and has been proven unlikely [204]. Given the very low rate of insertion and the absence of single gene transfection to date [205,206,207], this issue will benefit from concerted educational campaigns and repeated evidence. Repeat data may be harder to generate because the FDA has proposed that DNA vaccines prepared using a plasmid DNA previously documented to have an acceptable DNA biodistribution/integration profile could waive biodistribution/persistence studies. Additionally, newer bacterial-plasmid systems without antibiotic resistance genes are under investigation [208]. One bacterial-free DNA amplification system, the doggybone, has demonstrated an equivalent immune response from an entirely enzymatically constructed linear DNA vaccine [209].
Another concern is possible antigen tolerance after continuous exposure to DNA vaccine products. There is evidence that peripheral immunity can develop, thus having the opposite effect to vaccination [210]. This is very rare but still possible. As noted previously, nucleic acids are available for transcription until degradation, and while DNA vaccine plasmids are designed to be non-replicating [30], plasmids can be transcribed multiple times and can remain at the site for months after delivery [41]. This has purposeful application in the prevention and treatment of autoimmune disorders by intended, tolerizing DNA vaccines [211,212,213,214]. However, after DNA vaccines deliver nucleic acid to cells, the lifespan of the DNA vaccine is finite but dependent on numerous factors [215]. Repeated exposure to antigens has the potential to result in immunological tolerance [210,216,217]; the failure to recognize an antigen as a non-host due to prolonged exposure, in which case the immune system does not mount a response [192,198,199]. Tolerance could be problematic not only at the individual level but at the population level as well, because those vaccinated could ultimately become permanent carriers [218]. It might reasonably be assumed that injected plasmids could also induce anti-DNA antibodies, but this has been proven untrue to date in published studies [219]. Further investigation needs to be conducted to determine the impact of the prolonged expression of antigens by DNA vaccines on host responses.
7. Conclusions
DNA vaccine approval was accelerated by a worldwide need for a scalable and stable vaccine against SARS-2-CoV in 2020 [27,220]. However, DNA vaccine technology has been rapidly improving and advancing since first proposed in the early 1990s. Plasmid engineering has advanced, with robust promoters, immunostimulatory CpG motifs and immunostimulatory gene adjuvants now part of the design. BLAST sequencing and other bioinformatic tools [221,222] allow for rapid selection of robustly immunogenic antigens, and plasmids can be easily made polyvalent. Viral vectors are widely used in many DNA vaccines in development, hindered mainly by their limited load size and immunogenicity. Plasmid vectors can be transported in empty Gram-negative bacterial cell envelopes for their immunostimulatory effects, or plasmids can be attached to liposomes or nanoparticles to facilitate diffusion across the plasma membrane. The biolistic method, whereby DNA is forced into the cell, from the first successful study of DNA vaccines, remains under investigation. Likewise, the plasma membrane can be disrupted by sound and current. Some vectors now include cell-population-targeting motifs, most specifically to facilitate mucous membrane delivery. The immune micro-environment of different tissues is now better understood than ever before, with tattooed delivery potentially yielding immune response advantages in epidermal APC populations (Figure 6). These technologies can be combined in seemingly infinite ways. For instance, polycations are attached to liposomes to engage both the hydrophobic external plasma membrane as well as the typically negative membrane potential of most cells.
DNA vaccine technology is an emerging tool in the fight against cancer in the form of immunotherapy. Personalized medicine is a buzzword. Neoantigens may be appropriate targets for stimulation of the immune system against tumors and leukemias, and DNA vaccines can be assembled specifically and rapidly [223]. Likewise, epigenetic changes may be able to impart protective factors, both for the pathogen of interest and for future disease. Exciting evidence suggests that induced epigenetic changes may even be heritable, suggesting the possibility of lineage vaccination.
DNA vaccines hold demonstrated promise for global vaccination campaigns. They are stable, compact, scalable and reasonably simple to administer, depending on the delivery method. The discovery and development of optimal antigens is the key challenge; many exciting scientific and engineering advances occur regularly, beckoned by enormous potential. The great fear of genome integration is seemingly over-weighted, it being incredibly rare. Just as with all other vaccines approved for the general population, the continued adherence to, and updating of the FDA regulations [224,225] for monitoring adverse outcomes will lead to step-wise improvements in the safety of DNA vaccines. Compared to conventional peptide or subunit vaccines, the common side effect profile is somewhat more severe. However, the COVID vaccines provide the only gene vaccines for comparison to date [226], and it is likely that the specific antigen contributes more substantially to the varying side effects of different vaccines than do the vaccine technologies themselves. Conversely, at the population level, the serious adverse event rate is much lower for gene vaccines compared to most conventional vaccines [202]. However, at the individual level, the potential side effects of DNA vaccines are potentially more catastrophic, and this is a relatively unique paradox; a lower risk of a serious event but a higher risk of a catastrophic event. The advantages of DNA vaccines are clear, and the trajectory is one of rapid ascent. Careful policies, including ethical considerations and clearly defined measurables regarding both persons and populations, are essential to avoid anything like the decades-long delay suffered by gene therapy in the early 2000s [227,228,229].
Author Contributions
Conceptualization: J.H. and M.K.; First draft: M.K.; Critical revision of the article: M.K. and J.H.; Final approval of the version to be published: M.K. and J.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Institutes of Health (grant R01 CA47622), research initiation grants in the Department of Pathology and Laboratory medicine, Pennsylvania State University College of Medicine, and the Jake Gittlen Memorial Golf Tournament.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors have no conflicting interests to declare.
References
- Jedrzejczak-Silicka, M. History of Cell Culture. In New Insights into Cell Culture Technology; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- The Development of the Electron Microscope|American Association for the Advancement of Science (AAAS). Available online: https://www.aaas.org/development-electron-microscope (accessed on 11 September 2023).
- Shaber, L. The History of the Electron Microscope. Advancing Materials. Available online: https://www.thermofisher.com/blog/materials/the-history-of-the-electron-microscope/ (accessed on 11 September 2023).
- Cobb, M. 60 years ago, Francis Crick changed the logic of biology. PLoS Biol. 2017, 15, e2003243. [Google Scholar] [CrossRef] [PubMed]
- The Sequence of Sequencers: The History of Sequencing DNA—PMC. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4727787/ (accessed on 11 September 2023).
- Plotkin, S. History of vaccination. Proc. Natl. Acad. Sci. USA 2014, 111, 12283–12287. [Google Scholar] [CrossRef] [PubMed]
- Vaccine Timeline|History of Vaccines. The College of Physicians of Philadelphia. Available online: https://historyofvaccines.org/history/vaccine-timeline/overview (accessed on 18 January 2023).
- Vaccine History Timeline. Immunize.org. Available online: https://www.immunize.org/vaccines/vaccine-timeline/ (accessed on 11 December 2023).
- Srivastava, I.K.; Liu, M.A. Gene vaccines. Ann. Intern. Med. 2003, 138, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M.; Hu, J. The Integrated Consideration of Vaccine Platforms, Adjuvants, and Delivery Routes for Successful Vaccine Development. Vaccines 2023, 11, 695. [Google Scholar] [CrossRef]
- Cui, Z. DNA vaccine. Adv. Genet. 2005, 54, 257–289. [Google Scholar] [CrossRef]
- Jazayeri, S.D.; Poh, C.L. Recent advances in delivery of veterinary DNA vaccines against avian pathogens. Vet. Res. 2019, 50, 78. [Google Scholar] [CrossRef]
- Grimmett, E.; Al-Share, B.; Alkassab, M.B.; Zhou, R.W.; Desai, A.; Rahim, M.M.; Woldie, I. Cancer vaccines: Past, present and future; a review article. Discov. Oncol. 2022, 13, 31. [Google Scholar] [CrossRef]
- Liu, M.A. A Comparison of Plasmid DNA and mRNA as Vaccine Technologies. Vaccines 2019, 7, 37. [Google Scholar] [CrossRef]
- Leitner, W.W.; Ying, H.; Restifo, N.P. DNA and RNA-based vaccines: Principles, progress and prospects. Vaccine 1999, 18, 765–777. [Google Scholar] [CrossRef]
- Decades in the Making: mRNA COVID-19 Vaccines. NIH COVID-19 Research. Available online: https://covid19.nih.gov/nih-strategic-response-covid-19/decades-making-mrna-covid-19-vaccines (accessed on 11 September 2023).
- Fang, E.; Liu, X.; Li, M.; Zhang, Z.; Song, L.; Zhu, B.; Wu, X.; Liu, J.; Zhao, D.; Li, Y. Advances in COVID-19 mRNA vaccine development. Sig. Transduct Target Ther. 2022, 7, 94. [Google Scholar] [CrossRef]
- Perez-Rueda, E.; Hernandez-Guerrero, R.; Martinez-Nuñez, M.A.; Armenta-Medina, D.; Sanchez, I.; Ibarra, J.A. Abundance, diversity and domain architecture variability in prokaryotic DNA-binding transcription factors. PLoS ONE 2018, 13, e0195332. [Google Scholar] [CrossRef]
- Margaliot, M.; Huleihel, W.; Tuller, T. Variability in mRNA translation: A random matrix theory approach. Sci. Rep. 2021, 11, 5300. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.P.; Hossain, M.K.; Beladiya, J.; Apostolopoulos, V. Nucleic Acid Vaccines for COVID-19: A Paradigm Shift in the Vaccine Development Arena. Biologics 2021, 1, 337–356. [Google Scholar] [CrossRef]
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science 1990, 247 Pt 1, 1465–1468. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.C.; DeVit, M.; Johnston, S.A. Genetic immunization is a simple method for eliciting an immune response. Nature 1992, 356, 152–154. [Google Scholar] [CrossRef]
- Osborn, M.J.; McElmurry, R.T.; Lees, C.J.; DeFeo, A.P.; Chen, Z.Y.; Kay, M.A.; Naldini, L.; Freeman, G.; Tolar, J.; Blazar, B.R. Minicircle DNA-based gene therapy coupled with immune modulation permits long-term expression of α-L-iduronidase in mice with mucopolysaccharidosis type I. Mol. Ther. 2011, 19, 450–460. [Google Scholar] [CrossRef]
- Lechardeur, D.; Verkman, A.S.; Lukacs, G.L. Intracellular routing of plasmid DNA during non-viral gene transfer. Adv. Drug Deliv. Rev. 2005, 57, 755–767. [Google Scholar] [CrossRef]
- Travieso, T.; Li, J.; Mahesh, S.; Mello, J.D.F.R.E.; Blasi, M. The use of viral vectors in vaccine development. npj Vaccines 2022, 7, 75. [Google Scholar] [CrossRef]
- Hanke, T. New vector and vaccine platforms: mRNA, DNA, viral vectors. Curr. Opin. HIV AIDS 2022, 17, 338. [Google Scholar] [CrossRef]
- Mallapaty, S. India’s DNA COVID Vaccine Is a World First—More Are Coming. 2 September 2021. Available online: https://www.nature.com/articles/d41586-021-02385-x (accessed on 20 January 2023).
- Khobragade, A.; Bhate, S.; Ramaiah, V.; Deshpande, S.; Giri, K.; Phophle, H.; Supe, P.; Godara, I.; Revanna, R.; Nagarkar, R.; et al. Efficacy, safety, and immunogenicity of the DNA SARS-CoV-2 vaccine (ZyCoV-D): The interim efficacy results of a phase 3, randomised, double-blind, placebo-controlled study in India. Lancet 2022, 399, 1313–1321. [Google Scholar] [CrossRef]
- Blakney, A.K.; Bekker, L.-G. DNA vaccines join the fight against COVID-19. Lancet 2022, 399, 1281–1282. [Google Scholar] [CrossRef]
- Kutzler, M.A.; Weiner, D.B. DNA vaccines: Ready for prime time? Nat. Rev. Genet. 2008, 9, 776–788. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, R.R.; Boyer, J.D.; Ugen, K.E.; Lacy, K.E.; Gluckman, S.J.; Bagarazzi, M.L.; Chattergoon, M.A.; Baine, Y.; Higgins, T.J.; Ciccarelli, R.B.; et al. First human trial of a DNA-based vaccine for treatment of human immunodeficiency virus type 1 infection: Safety and host response. J. Infect Dis. 1998, 178, 92–100. [Google Scholar] [CrossRef]
- Andrews, S.M.; Rowland-Jones, S. Recent advances in understanding HIV evolution. F1000Research 2017, 6, 597. [Google Scholar] [CrossRef]
- Sekaly, R.-P. The failed HIV Merck vaccine study: A step back or a launching point for future vaccine development? J. Exp. Med. 2008, 205, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.E. The repeated setbacks of HIV vaccine development laid the groundwork for SARS-CoV-2 vaccines. Health Policy Technol. 2022, 11, 100619. [Google Scholar] [CrossRef] [PubMed]
- Cohen, K.W.; Fiore-Gartland, A.; Walsh, S.R.; Yusim, K.; Frahm, N.; Elizaga, M.L.; Maenza, J.; Scott, H.; Mayer, K.H.; Goepfert, P.A.; et al. Trivalent mosaic or consensus HIV immunogens prime humoral and broader cellular immune responses in adults. J. Clin. Investig. 2023, 133, e163338. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.; Xu, W.; Liu, S. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 9. [Google Scholar] [CrossRef]
- Home—ClinicalTrials.gov. Available online: https://classic.clinicaltrials.gov/ct2/home (accessed on 28 September 2023).
- Aida, V.; Pliasas, V.C.; Neasham, P.J.; North, J.F.; McWhorter, K.L.; Glover, S.R.; Kyriakis, C.S. Novel Vaccine Technologies in Veterinary Medicine: A Herald to Human Medicine Vaccines. Front. Vet. Sci. 2021, 8, 654289. Available online: https://www.frontiersin.org/articles/10.3389/fvets.2021.654289 (accessed on 16 October 2023). [CrossRef]
- Li, L.; Petrovsky, N. Molecular mechanisms for enhanced DNA vaccine immunogenicity. Expert Rev. Vaccines 2016, 15, 313–329. [Google Scholar] [CrossRef]
- Disis, M.L.; Guthrie, K.A.; Liu, Y.; Coveler, A.L.; Higgins, D.M.; Childs, J.S.; Dang, Y.; Salazar, L.G. Safety and Outcomes of a Plasmid DNA Vaccine Encoding the ERBB2 Intracellular Domain in Patients with Advanced-Stage ERBB2-Positive Breast Cancer: A Phase 1 Nonrandomized Clinical Trial. JAMA Oncol. 2023, 9, 71–78. [Google Scholar] [CrossRef]
- Shafaati, M.; Saidijam, M.; Soleimani, M.; Hazrati, F.; Mirzaei, R.; Amirheidari, B.; Tanzadehpanah, H.; Karampoor, S.; Kazemi, S.; Yavari, B.; et al. A brief review on DNA vaccines in the era of COVID-19. Future Virol. 2022, 17, 49–66. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, B.; Morrow, M.P.; Hutnick, N.A.; Shin, T.H.; Lucke, C.E.; Weiner, D.B. Clinical Applications of DNA Vaccines: Current Progress. Clin. Infect Dis. 2011, 53, 296–302. [Google Scholar] [CrossRef]
- Lopes, A.; Vandermeulen, G.; Préat, V. Cancer DNA vaccines: Current preclinical and clinical developments and future perspectives. J. Exp. Clin. Cancer Res. 2019, 38, 146. [Google Scholar] [CrossRef]
- Search for: DNA Vaccine|List Results|ClinicalTrials.gov. Available online: https://clinicaltrials.gov/search?intr=DNA%20Vaccine&viewType=Table&limit=100&page=1 (accessed on 1 January 2024).
- Gary, E.N.; Weiner, D.B. DNA vaccines: Prime time is now. Curr. Opin. Immunol. 2020, 65, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Maslow, J.N.; Kwon, I.; Kudchodkar, S.B.; Kane, D.; Tadesse, A.; Lee, H.; Park, Y.K.; Muthumani, K.; Roberts, C.C. DNA Vaccines for Epidemic Preparedness: SARS-CoV-2 and Beyond. Vaccines 2023, 11, 1016. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, E.W. The MHC class I antigen presentation pathway: Strategies for viral immune evasion. Immunology 2003, 110, 163–169. [Google Scholar] [CrossRef]
- Blum, J.S.; Wearsch, P.A.; Cresswell, P. Pathways of Antigen Processing. Annu. Rev. Immunol. 2013, 31, 443–473. [Google Scholar] [CrossRef]
- Delamarre, L.; Holcombe, H.; Mellman, I. Presentation of Exogenous Antigens on Major Histocompatibility Complex (MHC) Class I and MHC Class II Molecules Is Differentially Regulated during Dendritic Cell Maturation. J. Exp. Med. 2003, 198, 111–122. [Google Scholar] [CrossRef]
- Harding, C.V. Class I MHC presentation of exogenous antigens. J. Clin. Immunol. 1996, 16, 90–96. [Google Scholar] [CrossRef]
- Khan, K.H. DNA vaccines: Roles against diseases. Germs 2013, 3, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Leifert, J.A.; Whitton, J.L. Immune Responses to DNA Vaccines: Induction of CD8 T Cells. In Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2013. Available online: https://www.ncbi.nlm.nih.gov/books/NBK6523/ (accessed on 28 September 2023).
- Daniels, M.A.; Devine, L.; Miller, J.D.; Moser, J.M.; Lukacher, A.E.; Altman, J.D.; Kavathas, P.; Hogquist, K.A.; Jameson, S.C. CD8 binding to MHC class I molecules is influenced by T cell maturation and glycosylation. Immunity 2001, 15, 1051–1061. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.J.; Wu, M.S.; Barr, L.J.; Fuller, J.T.; Tussey, L.G.; Speller, S.; Culp, J.; Burkholder, J.K.; Swain, W.F.; Dixon, R.M.; et al. Induction of antigen-specific CD8+ T cells, T helper cells, and protective levels of antibody in humans by particle-mediated administration of a hepatitis B virus DNA vaccine. Vaccine 2000, 19, 764–778. [Google Scholar] [CrossRef] [PubMed]
- Hobernik, D.; Bros, M. DNA Vaccines—How Far From Clinical Use? Int. J. Mol. Sci. 2018, 19, 3605. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Yao, Y.; Zhang, X.; Liu, H.; Gao, G.; Peng, Y.; Chen, M.; Zhao, J.; Zhang, X.; Yin, C.; et al. Both chimpanzee adenovirus-vectored and DNA vaccines induced long-term immunity against Nipah virus infection. npj Vaccines 2023, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Embgenbroich, M.; Burgdorf, S. Current Concepts of Antigen Cross-Presentation. Front. Immunol. 2018, 9, 1643. Available online: https://www.frontiersin.org/articles/10.3389/fimmu.2018.01643 (accessed on 28 September 2023). [CrossRef] [PubMed]
- Gilbert, S.C. T-cell-inducing vaccines—what’s the future. Immunology 2012, 135, 19–26. [Google Scholar] [CrossRef]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D.M. CpG DNA as a vaccine adjuvant. Expert Rev. Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef]
- Krieg, A.M.; Yi, A.K.; Matson, S.; Waldschmidt, T.J.; Bishop, G.A.; Teasdale, R.; Koretzky, G.A.; Klinman, D.M. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 1995, 374, 546–549. [Google Scholar] [CrossRef]
- Hartl, A.; Kiesslich, J.; Weiss, R.; Bernhaupt, A.; Mostböck, S.; Scheiblhofer, S.; Ebner, C.; Ferreira, F.; Thalhamer, J. Immune responses after immunization with plasmid DNA encoding Bet v 1, the major allergen of birch pollen. J. Allergy Clin. Immunol. 1999, 103 Pt 1, 107–113. [Google Scholar] [CrossRef]
- Janeway, J.C.; Travers, P.; Walport, M.; Shlomchik, M.J. Antigen Recognition by B-cell and T-cell Receptors. In Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. Available online: https://www.ncbi.nlm.nih.gov/books/NBK10770/ (accessed on 28 September 2023).
- Szeto, C.; Lobos, C.A.; Nguyen, A.T.; Gras, S. TCR Recognition of Peptide–MHC-I: Rule Makers and Breakers. Int. J. Mol. Sci. 2020, 22, 68. [Google Scholar] [CrossRef] [PubMed]
- Amanna, I.J.; Slifka, M.K. Contributions of humoral and cellular immunity to vaccine-induced protection in humans. Virology 2011, 411, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Shedlock, D.J.; Weiner, D.B. DNA vaccination: Antigen presentation and the induction of immunity. J. Leukoc. Biol. 2000, 68, 793–806. [Google Scholar] [CrossRef] [PubMed]
- Koup, R.A.; Douek, D.C. Vaccine Design for CD8 T Lymphocyte Responses. Cold Spring Harb. Perspect. Med. 2011, 1, a007252. [Google Scholar] [CrossRef] [PubMed]
- Tsang, T.K.; Lam, K.T.; Liu, Y.; Fang, V.J.; Mu, X.; Leung, N.H.; Peiris, J.M.; Leung, G.M.; Cowling, B.J.; Tu, W. Investigation of CD4 and CD8 T cell-mediated protection against influenza A virus in a cohort study. BMC Med. 2022, 20, 230. [Google Scholar] [CrossRef]
- Reinscheid, M.; Luxenburger, H.; Karl, V.; Graeser, A.; Giese, S.; Ciminski, K.; Reeg, D.B.; Oberhardt, V.; Roehlen, N.; Lang-Meli, J. COVID-19 mRNA booster vaccine induces transient CD8+ T effector cell responses while conserving the memory pool for subsequent reactivation. Nat. Commun. 2022, 13, 4631. [Google Scholar] [CrossRef]
- Liu, J.; Yu, J.; McMahan, K.; Jacob-Dolan, C.; He, X.; Giffin, V.; Wu, C.; Sciacca, M.; Powers, O.; Nampanya, F.; et al. CD8 T cells contribute to vaccine protection against SARS-CoV-2 in macaques. Sci. Immunol. 2022, 7, eabq7647. [Google Scholar] [CrossRef]
- Dolina, J.S.; Lee, J.; Brightman, S.E.; McArdle, S.; Hall, S.M.; Thota, R.R.; Zavala, K.S.; Lanka, M.; Premlal, A.L.; Greenbaum, J.A.; et al. Linked CD4+/CD8+ T cell neoantigen vaccination overcomes immune checkpoint blockade resistance and enables tumor regression. J. Clin. Investig. 2023, 133, e164258. [Google Scholar] [CrossRef]
- Fierer, J.; Waters, C.; Walls, L. Both CD4+ and CD8+ T Cells Can Mediate Vaccine-Induced Protection against Coccidioides immitis Infection in Mice. J. Infect. Dis. 2006, 193, 1323–1331. [Google Scholar] [CrossRef]
- Oh, D.Y.; Fong, L.; Newell, E.W.; Turk, M.J.; Chi, H.; Chang, H.Y.; Satpathy, A.T.; Fairfax, B.; Silva-Santos, B.; Lantz, O. Toward a better understanding of T cells in cancer. Cancer Cell 2021, 39, 1549–1552. [Google Scholar] [CrossRef]
- Franck, C.O.; Fanslau, L.; Popov, A.B.; Tyagi, P.; Fruk, L. Biopolymer-based Carriers for DNA Vaccine Design. Angew. Chem. Int. Ed. 2021, 60, 13225–13243. [Google Scholar] [CrossRef] [PubMed]
- Weiss, R.; Leitner, W.W.; Scheiblhofer, S.; Chen, D.; Bernhaupt, A.; Mostböck, S.; Thalhamer, J.; Lyon, J.A. Genetic Vaccination against Malaria Infection by Intradermal and Epidermal Injections of a Plasmid Containing the Gene Encoding the Plasmodium berghei Circumsporozoite Protein. Infect. Immun. 2000, 68, 5914–5919. [Google Scholar] [CrossRef] [PubMed]
- Feltquate, D.M.; Heaney, S.; Webster, R.G.; Robinson, H.L. Different T helper cell types and antibody isotypes generated by saline and gene gun DNA immunization. J. Immunol. 1997, 158, 2278–2284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, Z.; Wang, Y.; Wu, S.; Wang, B.; Zhang, J.; Song, X.; Chen, Y.; Lv, P.; Hou, L. Comparative immunogenicity analysis of intradermal versus intramuscular immunization with a recombinant human adenovirus type 5 vaccine against Ebola virus. Front. Immunol. 2022, 13, 963049. Available online: https://www.frontiersin.org/articles/10.3389/fimmu.2022.963049 (accessed on 12 October 2023). [CrossRef] [PubMed]
- Fynan, E.F.; Webster, R.G.; Fuller, D.H.; Haynes, J.R.; Santoro, J.C.; Robinson, H.L. DNA vaccines: Protective immunizations by parenteral, mucosal, and gene-gun inoculations. Proc. Natl. Acad. Sci. USA 1993, 90, 11478–11482. [Google Scholar] [CrossRef] [PubMed]
- Black, M.; Trent, A.; Tirrell, M.; Olive, C. Advances in the design and delivery of peptide subunit vaccines with a focus on Toll-like receptor agonists. Expert Rev. Vaccines 2010, 9, 157–173. [Google Scholar] [CrossRef]
- Sarkar, B.; Ullah, M.A.; Araf, Y.; Rahman, M.S. Engineering a novel subunit vaccine against SARS-CoV-2 by exploring immunoinformatics approach. Inform. Med. Unlocked 2020, 21, 100478. [Google Scholar] [CrossRef]
- Krauson, A.J.; Casimero, F.V.C.; Siddiquee, Z.; Stone, J.R. Duration of SARS-CoV-2 mRNA vaccine persistence and factors associated with cardiac involvement in recently vaccinated patients. npj Vaccines 2023, 8, 141. [Google Scholar] [CrossRef]
- Klinman, D.M.; Klaschik, S.; Tross, D.; Shirota, H.; Steinhagen, F. FDA guidance on prophylactic DNA vaccines: Analysis and recommendations. Vaccine 2010, 28, 2801–2805. [Google Scholar] [CrossRef]
- Correia, B.E.; Bates, J.T.; Loomis, R.J.; Baneyx, G.; Carrico, C.; Jardine, J.G.; Rupert, P.; Correnti, C.; Kalyuzhniy, O.; Vittal, V.; et al. Proof of principle for epitope-focused vaccine design. Nature 2014, 507, 201–206. [Google Scholar] [CrossRef]
- Antonopoulou, T.; Athanassakis, I. SARS-CoV-2 immunogenicity: Is S protein the best target for vaccination? Vaccine 2022, 40, 3093–3095. [Google Scholar] [CrossRef]
- Yurina, V.; Adianingsih, O.R. Predicting epitopes for vaccine development using bioinformatics tools. Ther. Adv. Vaccines Immunother. 2022, 10, 25151355221100218. [Google Scholar] [CrossRef]
- Yurina, V.; Yudani, T.; Raras, M. Design and construction of DNA vaccine expressing lectin-like oxidize-LDL receptor-1 (LOX-1) as atherosclerosis vaccine candidate. J. Biotech. Res. 2017, 1, 103–112. [Google Scholar]
- Home—Protein—NCBI. Available online: https://www.ncbi.nlm.nih.gov/protein (accessed on 3 October 2023).
- Adianingsih, O.R.; Kharisma, V.D. Study of B cell epitope conserved region of the Zika virus envelope glycoprotein to develop multi-strain vaccine. J. Appl. Pharm. Sci. 2019, 9, 98–103. [Google Scholar]
- Sitompul, L.S.; Widodo, N.; Djati, M.S.; Utomo, D.H. Epitope mapping of gp350/220 conserved domain of epstein barr virus to develop nasopharyngeal carcinoma (npc) vaccine. Bioinformation 2012, 8, 479–482. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Racz, R.; Sayers, S.; Lin, Y.; Todd, T.; Hur, J.; Li, X.; Patel, M.; Zhao, B.; Chung, M.; et al. Updates on the web-based VIOLIN vaccine database and analysis system. Nucl. Acids Res. 2014, 42, D1124–D1132. [Google Scholar] [CrossRef]
- Immunotherapy for Cancer—NCI. Available online: https://www.cancer.gov/about-cancer/treatment/types/immunotherapy (accessed on 3 October 2023).
- June, C.H.; O’Connor, R.S.; Kawalekar, O.U.; Ghassemi, S.; Milone, M.C. CAR T cell immunotherapy for human cancer. Science 2018, 359, 1361–1365. [Google Scholar] [CrossRef]
- Melenhorst, J.J.; Chen, G.M.; Wang, M.; Porter, D.L.; Chen, C.; Collins, M.A.; Gao, P.; Bandyopadhyay, S.; Sun, H.; Zhao, Z.; et al. Decade-long leukaemia remissions with persistence of CD4+ CAR T cells. Nature 2022, 602, 7897. [Google Scholar] [CrossRef]
- Lei, J.; Ploner, A.; Elfström, K.M.; Wang, J.; Roth, A.; Fang, F.; Sundström, K.; Dillner, J.; Sparén, P. HPV Vaccination and the Risk of Invasive Cervical Cancer. N. Engl. J. Med. 2020, 383, 1340–1348. [Google Scholar] [CrossRef]
- Yaddanapudi, K.; Mitchell, R.A.; Eaton, J.W. Cancer vaccines. Oncoimmunology 2013, 2, e23403. [Google Scholar] [CrossRef]
- Snyder, A.; Makarov, V.; Merghoub, T.; Yuan, J.; Zaretsky, J.M.; Desrichard, A.; Walsh, L.A.; Postow, M.A.; Wong, P.; Ho, T.S.; et al. Genetic Basis for Clinical Response to CTLA-4 Blockade in Melanoma. N. Engl. J. Med. 2014, 371, 2189–2199. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.; Shen, G.; Gao, W.; Huang, Z.; Huang, C.; Fu, L. Neoantigens: Promising targets for cancer therapy. Sig. Transduct Target Ther. 2023, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xiao, Z.; Wang, D.; Jia, L.; Nie, S.; Zeng, X.; Hu, W. The screening, identification, design and clinical application of tumor-specific neoantigens for TCR-T cells. Mol. Cancer 2023, 22, 141. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.J.; Svensson-Arvelund, J.; Lubitz, G.S.; Marabelle, A.; Melero, I.; Brown, B.D.; Brody, J.D. Cancer vaccines: The next immunotherapy frontier. Nat. Cancer 2022, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Mellman, I. Oncology meets immunology: The cancer-immunity cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef]
- Finn, O.J. Cancer vaccines: Between the idea and the reality. Nat. Rev. Immunol. 2003, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Biswas, N.; Chakrabarti, S.; Padul, V.; Jones, L.D.; Ashili, S. Designing neoantigen cancer vaccines, trials, and outcomes. Front Immunol. 2023, 14, 1105420. [Google Scholar] [CrossRef]
- Fritah, H.; Rovelli, R.; Chiang, C.L.-L.; Kandalaft, L.E. The current clinical landscape of personalized cancer vaccines. Cancer Treat. Rev. 2022, 106, 102383. [Google Scholar] [CrossRef]
- Mattos-Arruda, L.D.; Blanco-Heredia, J.; Aguilar-Gurrieri, C.; Carrillo, J.; Blanco, J. New emerging targets in cancer immunotherapy: The role of neoantigens. ESMO Open 2019, 4, e000684. [Google Scholar] [CrossRef]
- Niemi, J.V.L.; Sokolov, A.V.; Schiöth, H.B. Neoantigen Vaccines; Clinical Trials, Classes, Indications, Adjuvants and Combinatorial Treatments. Cancers 2022, 14, 5163. [Google Scholar] [CrossRef]
- Eggleton, J.S.; Nagalli, S. Highly Active Antiretroviral Therapy (HAART). In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK554533/ (accessed on 2 November 2023).
- Report of the NIH Panel to Define Principles of Therapy of HIV Infection. Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/00052295.htm (accessed on 2 November 2023).
- Maggiolo, F.; Leone, S. Is HAART modifying the HIV epidemic? Lancet 2010, 376, 492–493. [Google Scholar] [CrossRef] [PubMed]
- Baylin, S.B.; Jones, P.A. Epigenetic Determinants of Cancer. Cold Spring Harb. Perspect Biol. 2016, 8, a019505. [Google Scholar] [CrossRef] [PubMed]
- Grønbæk, K.; Hother, C.; Jones, P.A. Epigenetic changes in cancer. APMIS 2007, 115, 1039–1059. [Google Scholar] [CrossRef]
- Agrawal, B. Heterologous Immunity: Role in Natural and Vaccine-Induced Resistance to Infections. Front Immunol. 2019, 10, 2631. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, H.S.; Ahmed, S.S.; Curtis, N.; Kollmann, T.R.; Levy, O.; Netea, M.G.; Pollard, A.J.; van Crevel, R.; Wilson, C.B. Harnessing the beneficial heterologous effects of vaccination. Nat. Rev. Immunol. 2016, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Saadatian-Elahi, M.; Aaby, P.; Shann, F.; Netea, M.G.; Levy, O.; Louis, J.; Picot, V.; Greenberg, M.; Warren, W. Heterologous vaccine effects. Vaccine 2016, 34, 3923–3930. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.J.; Finn, A.; Curtis, N. Non-specific effects of vaccines: Plausible and potentially important, but implications uncertain. Arch. Dis. Child 2017, 102, 1077–1081. [Google Scholar] [CrossRef]
- Oriol-Tordera, B.; Esteve-Codina, A.; Berdasco, M.; Rosás-Umbert, M.; Goncalves, E.; Duran-Castells, C.; Catala-Moll, F.; Llano, A.; Cedeno, S.; Puertas, M.C.; et al. Epigenetic landscape in the kick-and-kill therapeutic vaccine BCN02 clinical trial is associated with antiretroviral treatment interruption (ATI) outcome. EBioMedicine 2022, 78, 103956. [Google Scholar] [CrossRef]
- Takahashi, H.; Kühtreiber, W.M.; Keefe, R.C.; Lee, A.H.; Aristarkhova, A.; Dias, H.F.; Ng, N.; Nelson, K.J.; Bien, S.; Scheffey, D.; et al. BCG vaccinations drive epigenetic changes to the human T cell receptor: Restored expression in type 1 diabetes. Sci. Adv. 2022, 8, eabq7240. [Google Scholar] [CrossRef]
- Faustman, D.L.; Wang, L.; Okubo, Y.; Burger, D.; Ban, L.; Man, G.; Zheng, H.; Schoenfeld, D.; Pompei, R.; Avruch, J.; et al. Proof-of-concept, randomized, controlled clinical trial of Bacillus-Calmette-Guerin for treatment of long-term type 1 diabetes. PLoS ONE 2012, 7, e41756. [Google Scholar] [CrossRef]
- Kühtreiber, W.M.; Faustman, D.L. BCG Therapy for Type 1 Diabetes: Restoration of Balanced Immunity and Metabolism. Trends Endocrinol. Metab. 2019, 30, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Katzmarski, N.; Domínguez-Andrés, J.; Cirovic, B.; Renieris, G.; Ciarlo, E.; Le Roy, D.; Lepikhov, K.; Kattler, K.; Gasparoni, G.; Händler, K.; et al. Transmission of trained immunity and heterologous resistance to infections across generations. Nat. Immunol. 2021, 22, 11. [Google Scholar] [CrossRef]
- Ledwith, B.J.; Manam, S.; Troilo, P.J.; Barnum, A.B.; Pauley, C.J.; Griffiths, I.I.T.G.; Harper, L.B.; Beare, C.M.; Bagdon, W.J.; Nichols, W.W. Plasmid DNA vaccines: Investigation of integration into host cellular DNA following intramuscular injection in mice. Intervirology 2000, 43, 258–272. [Google Scholar] [CrossRef] [PubMed]
- Ledwith, B.J.; Manam, S.; Troilo, P.J.; Barnum, A.B.; Pauley, C.J.; Griffiths, T.G., 2nd; Harper, L.B.; Schock, H.B.; Zhang, H.; Faris, J.E.; et al. Plasmid DNA vaccines: Assay for integration into host genomic DNA. Dev. Biol. 2000, 104, 33–43. [Google Scholar]
- Kang, K.K.; Choi, S.M.; Choi, J.H.; Lee, D.S.; Kim, C.Y.; Ahn, B.O.; Kim, B.M.; Kim, W.B. Safety evaluation of GX-12, a new HIV therapeutic vaccine: Investigation of integration into the host genome and expression in the reproductive organs. Intervirology 2003, 46, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Iurescia, S.; Fioretti, D.; Rinaldi, M. A blueprint for DNA vaccine design. Methods Mol. Biol. 2014, 1143, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Shahin, V. Gatekeepers of the nucleus. Nat. Nanotechnol. 2016, 11, 8. [Google Scholar] [CrossRef]
- Warrington, K.H.; Gorbatyuk, O.S.; Harrison, J.K.; Opie, S.R.; Zolotukhin, S.; Muzyczka, N. Adeno-Associated Virus Type 2 VP2 Capsid Protein Is Nonessential and Can Tolerate Large Peptide Insertions at Its N Terminus. J. Virol. 2004, 78, 6595–6609. [Google Scholar] [CrossRef]
- Panté, N.; Kann, M. Nuclear Pore Complex Is Able to Transport Macromolecules with Diameters of ∼39 nm. Mol. Biol. Cell 2002, 13, 425–434. [Google Scholar] [CrossRef]
- Tran, E.J.; Wente, S.R. Dynamic nuclear pore complexes: Life on the edge. Cell 2006, 125, 1041–1053. [Google Scholar] [CrossRef]
- Azzam, I.; Liashkovich, I.; Luchtefeld, I.; Kouzel, I.U.; Shahin, V. Facilitating plasmid nuclear delivery by interfering with the selective nuclear pore barrier. Bioeng. Transl. Med. 2019, 4, e10136. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wu, T.; Zhang, B.; Liu, S.; Song, W.; Qiao, J.; Ruan, H. Types of nuclear localization signals and mechanisms of protein import into the nucleus. Cell Commun. Signal. 2021, 19, 60. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Qi, X.; Hu, Z.; Tang, Q. Mechanisms Mediating Nuclear Trafficking Involved in Viral Propagation by DNA Viruses. Viruses 2019, 11, 1035. [Google Scholar] [CrossRef] [PubMed]
- Alderton, M.R.; Gray, P.J.; Prolf, D.F. Genetic Vaccination: Can Plasmid DNA deliver its expectations? JMVH 2001, 10, 59–65. [Google Scholar]
- Garmory, H.S.; Brown, K.A.; Titball, R.W. DNA vaccines: Improving expression of antigens. Genet. Vaccines Ther. 2003, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Hemmi, H.; Takeuchi, O.; Kawai, T.; Kaisho, T.; Sato, S.; Sanjo, H.; Matsumoto, M.; Hoshino, K.; Wagner, H.; Takeda, K.; et al. A Toll-like receptor recognizes bacterial DNA. Nature 2000, 408, 740–745. [Google Scholar] [CrossRef]
- Finer, M.; Glorioso, J. A brief account of viral vectors and their promise for gene therapy. Gene Ther. 2017, 24, 1. [Google Scholar] [CrossRef]
- Lundstrom, K. Viral Vectors in Gene Therapy. Diseases 2018, 6, 42. [Google Scholar] [CrossRef]
- Ghosh, S.; Brown, A.M.; Jenkins, C.; Campbell, K. Viral Vector Systems for Gene Therapy: A Comprehensive Literature Review of Progress and Biosafety Challenges. Appl. Biosaf. 2020, 25, 7–18. [Google Scholar] [CrossRef]
- Mingozzi, F.; High, K.A. Immune responses to AAV vectors: Overcoming barriers to successful gene therapy. Blood 2013, 122, 23–36. [Google Scholar] [CrossRef]
- Samulski, R.J.; Muzyczka, N. AAV-Mediated Gene Therapy for Research and Therapeutic Purposes. Annu. Rev. Virol. 2014, 1, 427–451. [Google Scholar] [CrossRef]
- Chen, Y.H.; Keiser, M.S.; Davidson, B.L. Viral Vectors for Gene Transfer. Curr. Protoc. Mouse Biol. 2018, 8, e58. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, Z.; Tian, H.; Qi, M.; Zhai, Z.; Li, S.; Li, R.; Zhang, H.; Wang, W.; Fu, S.; et al. Biodistribution and Safety Assessment of Bladder Cancer Specific Recombinant Oncolytic Adenovirus in Subcutaneous Xenografts Tumor Model in Nude Mice. Curr. Gene Ther. 2012, 12, 67–76. [Google Scholar] [CrossRef]
- Liu, M.A.; Ulmer, J.B. Human clinical trials of plasmid DNA vaccines. Adv. Genet. 2005, 55, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Clinical Trials Using pTVG-HP Plasmid DNA Vaccine—NCI. Available online: https://www.cancer.gov/research/participate/clinical-trials/intervention/ptvg-hp-plasmid-dna-vaccine (accessed on 30 December 2023).
- Lim, K.L.; Jazayeri, S.D.; Yeap, S.K.; Alitheen, N.B.; Bejo, M.H.; Ideris, A.; Omar, A.R. Antibody and T cell responses induced in chickens immunized with avian influenza virus N1 and NP DNA vaccine with chicken IL-15 and IL-18. Res. Vet. Sci. 2013, 95, 1224–1234. [Google Scholar] [CrossRef]
- Capua, I.; Terregino, C.; Cattoli, G.; Mutinelli, F.; Rodriguez, J.F. Development of a DIVA (Differentiating Infected from Vaccinated Animals) strategy using a vaccine containing a heterologous neuraminidase for the control of avian influenza. Avian Pathol. 2003, 32, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wu, X.; Li, L.; Ge, S.; Liu, Z.; Wang, Z. Virus-like particles: Promising platforms with characteristics of DIVA for veterinary vaccine design. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 343–352. [Google Scholar] [CrossRef]
- Mandal, D.; Maran, A.; Yaszemski, M.J.; Bolander, M.E.; Sarkar, G. Cellular Uptake of Gold Nanoparticles Directly Cross-linked with Carrier Peptides by Osteosarcoma Cells. J. Mater. Sci. Mater. Med. 2009, 20, 347–350. [Google Scholar] [CrossRef]
- Rossi, G.; Monticelli, L. Gold nanoparticles in model biological membranes: A computational perspective. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2016, 1858, 2380–2389. [Google Scholar] [CrossRef]
- Van Lehn, R.C.; Atukorale, P.U.; Carney, R.P.; Yang, Y.S.; Stellacci, F.; Irvine, D.J.; Alexander-Katz, A. Effect of Particle Diameter and Surface Composition on the Spontaneous Fusion of Monolayer-Protected Gold Nanoparticles with Lipid Bilayers. Nano Lett. 2013, 13, 4060–4067. [Google Scholar] [CrossRef]
- Ledesma-Feliciano, C.; Chapman, R.; Hooper, J.W.; Elma, K.; Zehrung, D.; Brennan, M.B.; Spiegel, E.K. Improved DNA Vaccine Delivery with Needle-Free Injection Systems. Vaccines 2023, 11, 280. [Google Scholar] [CrossRef]
- Walker, M. Ground-breaking Approach to Vaccine Delivery Using Ultrasound Technology. Available online: https://medriva.com/breaking-news/revolutionizing-vaccine-delivery-needle-free-and-painless-ultrasound-technology/ (accessed on 10 December 2023).
- Jenner, E. On the Origin of the Vaccine Inoculation. Med. Phys. J. 1801, 5, 505–508. [Google Scholar]
- Fulginiti, V.A.; Papier, A.; Lane, J.M.; Neff, J.M.; Henderson, D.A.; Henderson, D.A.; Inglesby, T.V., Jr.; O’Toole, T. Smallpox Vaccination: A Review, Part I. Background, Vaccination Technique, Normal Vaccination and Revaccination, and Expected Normal Reactions. Clin. Infect. Dis. 2003, 37, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Gomez, A.M.; Babuadze, G.; Plourde-Campagna, M.A.; Azizi, H.; Berger, A.; Kozak, R.; de La Vega, M.A.; Xiii, A.; Naghibosadat, M.; Nepveu-Traversy, M.E.; et al. A novel intradermal tattoo-based injection device enhances the immunogenicity of plasmid DNA vaccines. npj Vaccines 2022, 7, 172. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Sunada, Y. Plasmid DNA gene therapy by electroporation: Principles and recent advances. Curr. Gene Ther. 2011, 11, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Pokorna, D.; Rubio, I.; Müller, M. DNA-vaccination via tattooing induces stronger humoral and cellular immune responses than intramuscular delivery supported by molecular adjuvants. Genet. Vaccines Ther. 2008, 6, 4. [Google Scholar] [CrossRef]
- Otten, G.; Schaefer, M.; Doe, B.; Liu, H.; Srivastava, I.; zur Megede, J.; O’Hagan, D.; Donnelly, J.; Widera, G.; Rabussay, D.; et al. Enhancement of DNA vaccine potency in rhesus macaques by electroporation. Vaccine 2004, 22, 2489–2493. [Google Scholar] [CrossRef]
- Xu, R. Sonoporation as a Novel Delivery Platform for an HIV DNA Vaccine. Available online: https://grantome.com/grant/NIH/R03-AI141031-01A1 (accessed on 5 October 2023).
- Zhang, N.; Foiret, J.; Kheirolomoom, A.; Liu, P.; Feng, Y.; Tumbale, S.; Raie, M.; Wu, B.; Wang, J.; Fite, B.Z.; et al. Optimization of microbubble-based DNA vaccination with low-frequency ultrasound for enhanced cancer immunotherapy. Adv. Ther. 2021, 4, 2100033. [Google Scholar] [CrossRef]
- Wang, C.; Karlsson, A.; Oguin, I.I.I.T.H.; Macintyre, A.N.; Sempowski, G.D.; McCarthy, K.R.; Wang, Y.; Moody, M.A.; Yuan, F. Transient inhibition of lysosomal functions potentiates nucleic acid vaccines. Proc. Natl. Acad. Sci. USA 2023, 120, e2306465120. [Google Scholar] [CrossRef]
- Braathen, R.; Spång, H.C.; Hinke, D.M.; Blazevski, J.; Bobic, S.; Fossum, E.; Bogen, B. A DNA Vaccine That Encodes an Antigen-Presenting Cell-Specific Heterodimeric Protein Protects against Cancer and Influenza. Mol. Ther. Methods Clin. Dev. 2020, 17, 378–392. [Google Scholar] [CrossRef]
- Kawamura, H.; Berzofsky, J.A. Enhancement of antigenic potency in vitro and immunogenicity in vivo by coupling the antigen to anti-immunoglobulin. J. Immunol. 1986, 136, 58–65. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, X.; Csencsits, K.L.; Haddad, A.; Walters, N.; Pascual, D.W. M cell-targeted DNA vaccination. Proc. Natl. Acad. Sci. USA 2001, 98, 9318–9323. [Google Scholar] [CrossRef] [PubMed]
- Hillemanns, P.; Denecke, A.; Woelber, L.; Böhmer, G.; Jentschke, M.; Schjetne, K.W.; Bruins Slot, K.M.; Fredriksen, A.B. A Therapeutic Antigen-Presenting Cell-Targeting DNA Vaccine VB10.16 in HPV16-Positive High-Grade Cervical Intraepithelial Neoplasia: Results from a Phase I/IIa Trial. Clin. Cancer Res. 2022, 28, 4885–4892. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.D.; Ugen, K.E.; Wang, B.; Agadjanyan, M.; Gilbert, L.; Bagarazzi, M.L.; Chattergoon, M.; Frost, P.; Javadian, A.; Williams, W.V.; et al. Protection of chimpanzees from high-dose heterologous HIV-1 challenge by DNA vaccination. Nat. Med. 1997, 3, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Werninghaus, I.C.; Hinke, D.M.; Fossum, E.; Bogen, B.; Braathen, R. Neuraminidase delivered as an APC-targeted DNA vaccine induces protective antibodies against influenza. Mol. Ther. 2023, 31, 2188–2205. [Google Scholar] [CrossRef]
- Suschak, J.J.; Williams, J.A.; Schmaljohn, C.S. Advancements in DNA vaccine vectors, non-mechanical delivery methods, and molecular adjuvants to increase immunogenicity. Hum. Vaccin. Immunother. 2017, 13, 2837–2848. [Google Scholar] [CrossRef]
- Putnam, D.; Gentry, C.A.; Pack, D.W.; Langer, R. Polymer-based gene delivery with low cytotoxicity by a unique balance of side-chain termini. Proc. Natl. Acad. Sci. USA 2001, 98, 1200–1205. [Google Scholar] [CrossRef]
- Li, S.; Li, W.; Jin, Y.; Wu, B.; Wu, Y. Advancements in the development of nucleic acid vaccines for syphilis prevention and control. Hum. Vaccines Immunother. 2023, 19, 2234790. [Google Scholar] [CrossRef]
- Rizeq, B.R.; Younes, N.N.; Rasool, K.; Nasrallah, G.K. Synthesis, Bioapplications, and Toxicity Evaluation of Chitosan-Based Nanoparticles. Int. J. Mol. Sci. 2019, 20, 5776. [Google Scholar] [CrossRef]
- Xu, Z.; Patel, A.; Tursi, N.J.; Zhu, X.; Muthumani, K.; Kulp, D.W.; Weiner, D.B. Harnessing Recent Advances in Synthetic DNA and Electroporation Technologies for Rapid Vaccine Development Against COVID-19 and Other Emerging Infectious Diseases. Front. Med. Technol. 2020, 2. Available online: https://www.frontiersin.org/articles/10.3389/fmedt.2020.571030 (accessed on 3 October 2023). [CrossRef]
- Stenler, S.; Blomberg, P.; Smith, C.E. Safety and efficacy of DNA vaccines. Hum. Vaccines Immunother. 2014, 10, 1306–1308. [Google Scholar] [CrossRef]
- Grunwald, T.; Ulbert, S. Improvement of DNA vaccination by adjuvants and sophisticated delivery devices: Vaccine-platforms for the battle against infectious diseases. Clin. Exp. Vaccine Res. 2015, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.H.; Chen, H.W.; Tao, M.H. Modulation of immune responses to DNA vaccines by codelivery of cytokine genes. J. Formos Med. Assoc. 1999, 98, 722–729. [Google Scholar] [PubMed]
- Scheerlinck, J.P.; Casey, G.; McWaters, P.; Kelly, J.; Woollard, D.; Lightowlers, M.W.; Tennent, J.M.; Chaplin, P.J. The immune response to a DNA vaccine can be modulated by co-delivery of cytokine genes using a DNA prime-protein boost strategy. Vaccine 2001, 19, 4053–4060. [Google Scholar] [CrossRef] [PubMed]
- Kwissa, M.; Kröger, A.; Hauser, H.; Reimann, J.; Schirmbeck, R. Cytokine-facilitated priming of CD8+ T cell responses by DNA vaccination. J. Mol. Med. 2003, 81, 91–101. [Google Scholar] [CrossRef]
- Flingai, S.; Czerwonko, M.; Goodman, J.; Kudchodkar, S.B.; Muthumani, K.; Weiner, D.B. Synthetic DNA Vaccines: Improved Vaccine Potency by Electroporation and Co-Delivered Genetic Adjuvants. Front. Immunol. 2013, 4, 54. [Google Scholar] [CrossRef]
- Cornelie, S.; Poulain-Godefroy, O.; Lund, C.; Vendeville, C.; Ban, E.; Capron, M.; Riveau, G. Methylated CpG-containing plasmid activates the immune system. Scand. J. Immunol. 2004, 59, 143–151. [Google Scholar] [CrossRef]
- Krieg, A.M. CpG motifs in bacterial DNA and their immune effects. Annu. Rev. Immunol. 2002, 20, 709–760. [Google Scholar] [CrossRef]
- Wahren, B.; Liu, M.A. DNA Vaccines: Recent Developments and the Future. Vaccines 2014, 2, 785–796. [Google Scholar] [CrossRef]
- Häcker, G.; Redecke, V.; Häcker, H. Activation of the immune system by bacterial CpG-DNA. Immunology 2002, 105, 245–251. [Google Scholar] [CrossRef]
- Zhao, B.G.; Vasilakos, J.P.; Tross, D.; Smirnov, D.; Klinman, D.M. Combination therapy targeting toll like receptors 7, 8 and 9 eliminates large established tumors. J. ImmunoTherapy Cancer 2014, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Sabbaghi, A.; Ghaemi, A. Molecular Adjuvants for DNA Vaccines: Application, Design, Preparation, and Formulation. Methods Mol. Biol. 2021, 2197, 87–112. [Google Scholar] [CrossRef]
- Kalams, S.A.; Parker, S.; Jin, X.; Elizaga, M.; Metch, B.; Wang, M.; Hural, J.; Lubeck, M.; Eldridge, J.; Cardinali, M.; et al. Safety and immunogenicity of an HIV-1 gag DNA vaccine with or without IL-12 and/or IL-15 plasmid cytokine adjuvant in healthy, HIV-1 uninfected adults. PLoS ONE 2012, 7, e29231. [Google Scholar] [CrossRef]
- Luchner, M.; Reinke, S.; Milicic, A. TLR Agonists as Vaccine Adjuvants Targeting Cancer and Infectious Diseases. Pharmaceutics 2021, 13, 142. [Google Scholar] [CrossRef]
- Kramer, K.; Shields, N.J.; Poppe, V.; Young, S.L.; Walker, G.F. Intracellular Cleavable CpG Oligodeoxynucleotide-Antigen Conjugate Enhances Anti-tumor Immunity. Mol. Ther. 2017, 25, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.Y.; Lin, M.Y.; Coffman, R.L.; Campbell, J.D.; Traquina, P.; Lin, Y.J.; Liu, L.T.; Cheng, J.; Wu, Y.C.; Wu, C.C.; et al. Development of CpG-adjuvanted stable prefusion SARS-CoV-2 spike antigen as a subunit vaccine against COVID-19. Sci. Rep. 2020, 10, 20085. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.C.; Huang, M.S.; Chiu, F.F.; Chai, K.M.; Liao, C.L.; Wu, S.C.; Chen, H.W.; Liu, S.J. Co-delivery of a trimeric spike DNA and protein vaccine with aluminum hydroxide enhanced Th1-dominant humoral and cellular immunity against SARS-CoV-2. J. Med. Virol. 2023, 95, e29040. [Google Scholar] [CrossRef]
- Li, E.; Zhang, Y. DNA Methylation in Mammals. Cold Spring Harb. Perspect Biol. 2014, 6, a019133. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, G.-M. The role of bacterial DNA containing CpG motifs in diseases. J. Leukoc. Biol. 2021, 109, 991–998. [Google Scholar] [CrossRef]
- Faurez, F.; Dory, D.; Le Moigne, V.; Gravier, R.; Jestin, A. Biosafety of DNA vaccines: New generation of DNA vectors and current knowledge on the fate of plasmids after injection. Vaccine 2010, 28, 3888–3895. [Google Scholar] [CrossRef]
- Martin, T.; Parker, S.E.; Hedstrom, R.; Le, T.; Hoffman, S.L.; Norman, J.; Hobart, P.; Lew, D. Plasmid DNA malaria vaccine: The potential for genomic integration after intramuscular injection. Hum. Gene Ther. 1999, 10, 759–768. [Google Scholar] [CrossRef]
- Zhang, L.; Richards, A.; Barrasa, M.I.; Hughes, S.H.; Young, R.A.; Jaenisch, R. Reverse-transcribed SARS-CoV-2 RNA can integrate into the genome of cultured human cells and can be expressed in patient-derived tissues. Proc. Natl. Acad. Sci. USA 2021, 118, e2105968118. [Google Scholar] [CrossRef] [PubMed]
- Integration Into the Human Genome? Available online: https://www.science.org/content/blog-post/integration-human-genome (accessed on 17 October 2023).
- Rudolf, J. Eminent MIT Scientists Defend Controversial SARS-CoV-2 Genome Integration Results. GEN-Genetic Engineering and Biotechnology News. Available online: https://www.genengnews.com/insights/eminent-mit-scientists-defend-controversial-sars-cov-2-genome-integration-results/ (accessed on 17 October 2023).
- No, the COVID-19 Vaccine Will Not Change Your DNA-mlive.com. Available online: https://www.mlive.com/public-interest/2021/04/no-the-covid-19-vaccine-will-not-change-your-dna.html (accessed on 17 October 2023).
- Fox, A.; Martin, J.; Beilharz, T. Can the Pfizer or Moderna mRNA Vaccines Affect My Genetic Code? The Conversation. Available online: http://theconversation.com/can-the-pfizer-or-moderna-mrna-vaccines-affect-my-genetic-code-162590 (accessed on 17 October 2023).
- Araten, D.J.; Golde, D.W.; Zhang, R.H.; Thaler, H.T.; Gargiulo, L.; Notaro, R.; Luzzatto, L. A Quantitative Measurement of the Human Somatic Mutation Rate. Cancer Res. 2005, 65, 8111–8117. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.L.; Loeb, L.A. The mutation rate and cancer. Genetics 1998, 148, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Neely, S.R.; Eldredge, C.; Ersing, R.; Remington, C. Vaccine Hesitancy and Exposure to Misinformation: A Survey Analysis. J. Gen. Intern. Med. 2022, 37, 179–187. [Google Scholar] [CrossRef]
- Pertwee, E.; Simas, C.; Larson, H.J. An epidemic of uncertainty: Rumors, conspiracy theories and vaccine hesitancy. Nat. Med. 2022, 28, 3. [Google Scholar] [CrossRef]
- Banoun, H. mRNA: Vaccine or Gene Therapy? The Safety Regulatory Issues. Int. J. Mol. Sci. 2023, 24, 10514. [Google Scholar] [CrossRef]
- Spencer, J.P.; Pawlowski, R.H.T.; Thomas, S. Vaccine Adverse Events: Separating Myth from Reality. Am. Fam. Physician 2017, 95, 786–794. [Google Scholar]
- Mignon, C.; Sodoyer, R.; Werle, B. Antibiotic-Free Selection in Biotherapeutics: Now and Forever. Pathogens 2015, 4, 157–181. [Google Scholar] [CrossRef]
- Mairhofer, J.; Lara, A.R. Advances in host and vector development for the production of plasmid DNA vaccines. Methods Mol. Biol. 2014, 1139, 505–541. [Google Scholar] [CrossRef] [PubMed]
- Cranenburgh, R.M.; Hanak, J.A.J.; Williams, S.G.; Sherratt, D.J. Escherichia coli strains that allow antibiotic-free plasmid selection and maintenance by repressor titration. Nucleic Acids Res. 2001, 29, e26. [Google Scholar] [CrossRef] [PubMed]
- Garmory, H.S.; Leckenby, M.W.; Griffin, K.F.; Elvin, S.J.; Taylor, R.R.; Hartley, M.G.; Hanak, J.A.; Williamson, E.D.; Cranenburgh, R.M. Antibiotic-Free Plasmid Stabilization by Operator-Repressor Titration for Vaccine Delivery by Using Live Salmonella enterica Serovar Typhimurium. Infect. Immun. 2005, 73, 2005–2011. [Google Scholar] [CrossRef] [PubMed]
- Mairhofer, J.; Pfaffenzeller, I.; Merz, D.; Grabherr, R. A novel antibiotic free plasmid selection system: Advances in safe and efficient DNA therapy. Biotechnol. J. 2008, 3, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Alcolea, P.J.; Larraga, J.; Rodríguez-Martín, D.; Alonso, A.; Loayza, F.J.; Rojas, J.M.; Ruiz-García, S.; Louloudes-Lázaro, A.; Carlón, A.B.; Sánchez-Cordón, P.J.; et al. Non-replicative antibiotic resistance-free DNA vaccine encoding S and N proteins induces full protection in mice against SARS-CoV-2. Front. Immunol. 2022, 13, 1023255. [Google Scholar] [CrossRef] [PubMed]
- Walters, A.A.; Kinnear, E.; Shattock, R.J.; McDonald, J.U.; Caproni, L.J.; Porter, N.; Tregoning, J.S. Comparative analysis of enzymatically produced novel linear DNA constructs with plasmids for use as DNA vaccines. Gene Ther. 2014, 21, 7. [Google Scholar] [CrossRef]
- Niezold, T.; Storcksdieck Genannt Bonsmann, M.; Maaske, A.; Temchura, V.; Heinecke, V.; Hannaman, D.; Buer, J.; Ehrhardt, C.; Hansen, W.; Überla, K.; et al. DNA vaccines encoding DEC205-targeted antigens: Immunity or tolerance? Immunology 2015, 145, 519–533. [Google Scholar] [CrossRef]
- Ho, P.P.; Higgins, J.P.; Kidd, B.A.; Tomooka, B.; DiGennaro, C.; Lee, L.Y.; De Vegvar, H.E.; Steinman, L.; Robinson, W.H. Tolerizing DNA vaccines for autoimmune arthritis. Autoimmunity 2006, 39, 675–682. [Google Scholar] [CrossRef]
- Moorman, C.D.; Sohn, S.J.; Phee, H. Emerging Therapeutics for Immune Tolerance: Tolerogenic Vaccines, T cell Therapy, and IL-2 Therapy. Front. Immunol. 2021, 12, 657768. Available online: https://www.frontiersin.org/articles/10.3389/fimmu.2021.657768 (accessed on 18 October 2023). [CrossRef]
- Parks, R.J.; Gussoni, E. Building immune tolerance through DNA vaccination. Proc. Natl. Acad. Sci. USA 2018, 115, 9652–9654. [Google Scholar] [CrossRef]
- Umeshappa, C.S.; Singha, S.; Blanco, J.; Shao, K.; Nanjundappa, R.H.; Yamanouchi, J.; Parés, A.; Serra, P.; Yang, Y.; Santamaria, P. Suppression of a broad spectrum of liver autoimmune pathologies by single peptide-MHC-based nanomedicines. Nat. Commun. 2019, 10, 2150. [Google Scholar] [CrossRef]
- Wolff, J.A.; Ludtke, J.J.; Acsadi, G.; Williams, P.; Jani, A. Long-term persistence of plasmid DNA and foreign gene expression in mouse muscle. Hum. Mol. Genet. 1992, 1, 363–369. [Google Scholar] [CrossRef]
- Soucy, A.M.; Hurteau, G.J.; Metzger, D.W. Live Vaccination Generates Both Disease Tolerance and Host Resistance During Chronic Pulmonary Infection With Highly Virulent Francisella tularensis SchuS4. J. Infect. Dis. 2018, 218, 1802–1812. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N.; Redwan, E.M.; Makis, W.; Rubio-Casillas, A. IgG4 Antibodies Induced by Repeated Vaccination May Generate Immune Tolerance to the SARS-CoV-2 Spike Protein. Vaccines 2023, 11, 991. [Google Scholar] [CrossRef]
- Górecki, D.C.; Simons, J.P. The dangers of DNA vaccination. Nat. Med. 1999, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.; Williams, P.; Berg, R.K.; Hodgeman, B.A.; Liu, L.; Repetto, G.; Wolff, J.A. Direct gene transfer into nonhuman primate myofibers in vivo. Hum. Gene Ther. 1992, 3, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, C. First COVID-19 DNA vaccine approved, others in hot pursuit. Nat. Biotechnol. 2021, 39, 1479–1482. [Google Scholar] [CrossRef]
- Nucleotide BLAST: Search Nucleotide Databases Using a Nucleotide Query. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&BLAST_SPEC=GeoBlast&PAGE_TYPE=BlastSearch (accessed on 19 October 2023).
- Basic Local Alignment Search Tool (BLAST)|Learn Science at Scitable. Available online: https://www.nature.com/scitable/topicpage/basic-local-alignment-search-tool-blast-29096/ (accessed on 19 October 2023).
- Cai, Y.; Rodriguez, S.; Hebel, H. DNA vaccine manufacture: Scale and quality. Expert Rev. Vaccines 2009, 8, 1277–1291. [Google Scholar] [CrossRef]
- General Principles for the Development of Vaccines to Protect against Global Infectious Diseases; CDC, 21 CFR 10.115(g)(4)(i); Food and Drug Administration: Rockville, MD, USA, 2011.
- Ensuring the Safety of Vaccines in the United States|CDC. Available online: https://www.cdc.gov/vaccines/hcp/conversations/ensuring-safe-vaccines.html (accessed on 12 November 2023).
- Rabail, R.; Ahmed, W.; Ilyas, M.; Rajoka, M.S.; Hassoun, A.; Khalid, A.R.; Khan, M.R.; Aadil, R.M. The Side Effects and Adverse Clinical Cases Reported after COVID-19 Immunization. Vaccines 2022, 10, 488. [Google Scholar] [CrossRef]
- Couzin, J.; Kaiser, J. As Gelsinger Case Ends, Gene Therapy Suffers Another Blow. Science 2005, 307, 1028. [Google Scholar] [CrossRef]
- Rinde, M. The Death of Jesse Gelsinger, 20 Years Later. Science History Institute. Available online: https://sciencehistory.org/stories/magazine/the-death-of-jesse-gelsinger-20-years-later/ (accessed on 2 December 2023).
- Kaiser, J. Decades after a Tragic Failure, Gene Therapy Successfully Treats a Rare Liver Disease. Available online: https://www.science.org/content/article/decades-after-tragic-failure-gene-therapy-successfully-treats-rare-liver-disease (accessed on 2 December 2023).
Figure 1.
Some of the components of DNA vaccine technology. Many factors including delivery route, vector design, adjuvant choice and promotor/gene design all play important roles in the outcome of DNA vaccinations. Combinations of these different components make the DNA vaccine one of the most versatile yet challenging vaccine formats. Created with BioRender.com.
Figure 1.
Some of the components of DNA vaccine technology. Many factors including delivery route, vector design, adjuvant choice and promotor/gene design all play important roles in the outcome of DNA vaccinations. Combinations of these different components make the DNA vaccine one of the most versatile yet challenging vaccine formats. Created with BioRender.com.

Figure 2.
Conventional vaccine mechanism and DNA vaccine mechanism. Conventional vaccines (on the left) including peptide, subunit, live and attenuated viruses and toxins require endocytosis and intracellular processing of the pathogen. Because the pathogen is exogenous to the presenting cell, it is processed through the MHC II pathway, which preferentially engages CD4+ cells. DNA vaccine (on the right) can be endocytosed or can be engineered to passively cross the phospholipid membrane. The nucleic acid then locates to the nucleus and transcription and translation occur as if the DNA were native, which leads to presentation of the peptide through the MHC I pathway, preferentially activating CD8+ cells, additionally, the same peptide is exocytosed and then taken up by nearby cells, which then present the peptide via the MHC II pathway. Adapted from “COVID-19 DNA-Based Vaccine”, by BioRender.com (2023). Retrieved from https://app.biorender.com/biorender-templates accessed on 10 December 2023
Figure 2.
Conventional vaccine mechanism and DNA vaccine mechanism. Conventional vaccines (on the left) including peptide, subunit, live and attenuated viruses and toxins require endocytosis and intracellular processing of the pathogen. Because the pathogen is exogenous to the presenting cell, it is processed through the MHC II pathway, which preferentially engages CD4+ cells. DNA vaccine (on the right) can be endocytosed or can be engineered to passively cross the phospholipid membrane. The nucleic acid then locates to the nucleus and transcription and translation occur as if the DNA were native, which leads to presentation of the peptide through the MHC I pathway, preferentially activating CD8+ cells, additionally, the same peptide is exocytosed and then taken up by nearby cells, which then present the peptide via the MHC II pathway. Adapted from “COVID-19 DNA-Based Vaccine”, by BioRender.com (2023). Retrieved from https://app.biorender.com/biorender-templates accessed on 10 December 2023

Figure 3.
Neoantigen mechanism, a DNA vaccine immunotherapy target. Neopeptides occur when DNA mutation results in a novel transcriptional template leading to a novel polypeptide. Neoantigens occur when the neopeptide is presented on MHC, with the new portion of the polypeptide oriented for exposure to immune cells as a novel epitope. Adapted from “Neoantigen Presentation”, by BioRender.com (2023). Retrieved from https://app.biorender.com/biorender-templates accessed on 10 December 2023.
Figure 3.
Neoantigen mechanism, a DNA vaccine immunotherapy target. Neopeptides occur when DNA mutation results in a novel transcriptional template leading to a novel polypeptide. Neoantigens occur when the neopeptide is presented on MHC, with the new portion of the polypeptide oriented for exposure to immune cells as a novel epitope. Adapted from “Neoantigen Presentation”, by BioRender.com (2023). Retrieved from https://app.biorender.com/biorender-templates accessed on 10 December 2023.

Figure 4.
DNA Vaccine transport across the plasma membrane. For DNA vaccines to have the intended effect, the DNA must be transported across the plasma membrane and the DNA must then enter the nucleus. Nuclear localization signals are often intrinsic to DNA viruses but can be engineered in to plasmids. Plasmid transport across the plasma membrane can be facilitated in many ways, some of which are represented here. Clockwise from viral capsid. Viral capsid (most often as viral vector) protects and facilitates delivery of nucleic acid cargo to cytosol of cells by harnessing viral properties for cellular engagement and cargo routing. Nanoparticles can be engineered to small sizes with lipophilicity and non-polarity to facilitate transportation of plasmid across the plasma membrane. Cationic conjugates can be attached to negatively charged nucleic acids of plasmids to facilitate transport across plasma membrane. Coated gold beads are used as carrier molecules similar to engineered nanoparticles and can easily cross the phospholipid membrane to reach the cytosol. Liposomal covering allows for passive diffusion across lipid bilayers including the nuclear envelope. Empty bacterial capsules contain intrinsic immunostimulatory properties and can prevent extracellular breakdown of naked nucleic acid. Created with BioRender.com.
Figure 4.
DNA Vaccine transport across the plasma membrane. For DNA vaccines to have the intended effect, the DNA must be transported across the plasma membrane and the DNA must then enter the nucleus. Nuclear localization signals are often intrinsic to DNA viruses but can be engineered in to plasmids. Plasmid transport across the plasma membrane can be facilitated in many ways, some of which are represented here. Clockwise from viral capsid. Viral capsid (most often as viral vector) protects and facilitates delivery of nucleic acid cargo to cytosol of cells by harnessing viral properties for cellular engagement and cargo routing. Nanoparticles can be engineered to small sizes with lipophilicity and non-polarity to facilitate transportation of plasmid across the plasma membrane. Cationic conjugates can be attached to negatively charged nucleic acids of plasmids to facilitate transport across plasma membrane. Coated gold beads are used as carrier molecules similar to engineered nanoparticles and can easily cross the phospholipid membrane to reach the cytosol. Liposomal covering allows for passive diffusion across lipid bilayers including the nuclear envelope. Empty bacterial capsules contain intrinsic immunostimulatory properties and can prevent extracellular breakdown of naked nucleic acid. Created with BioRender.com.

Figure 5.
Common adjuvants and proposed mechanisms. (A) The CpG motifs stimulate TLRs, which activate signal transduction cascades, leading to nuclear upregulation of cytokines which stimulate immune cell response; most famously, the STING pathway. (B) Oil emulsions stimulate endoplasmic reticulum (ER) stress signaling pathway, which upregulates inflammatory cytokines, like interleukin-6 (IL-6), which activate and potentiate immune cells. (C) DNA stimulates signal transduction cascade, which upregulates transcription of inflammatory genes. (D) Aluminum causes inflammation and stimulates a release of ATP and DNA from injured cells, which activate DCs and macrophages. (E) Aluminum also stimulates inflammasomes. (F) Gene Adjuvants are immunostimulatory genes added to the plasmid for transcription within the cell (or can be delivered as separate plasmids alongside the vaccine), which are transcribed with the antigenic gene and work to potentiate immune cell response. Created with BioRender.com.
Figure 5.
Common adjuvants and proposed mechanisms. (A) The CpG motifs stimulate TLRs, which activate signal transduction cascades, leading to nuclear upregulation of cytokines which stimulate immune cell response; most famously, the STING pathway. (B) Oil emulsions stimulate endoplasmic reticulum (ER) stress signaling pathway, which upregulates inflammatory cytokines, like interleukin-6 (IL-6), which activate and potentiate immune cells. (C) DNA stimulates signal transduction cascade, which upregulates transcription of inflammatory genes. (D) Aluminum causes inflammation and stimulates a release of ATP and DNA from injured cells, which activate DCs and macrophages. (E) Aluminum also stimulates inflammasomes. (F) Gene Adjuvants are immunostimulatory genes added to the plasmid for transcription within the cell (or can be delivered as separate plasmids alongside the vaccine), which are transcribed with the antigenic gene and work to potentiate immune cell response. Created with BioRender.com.

Figure 6.
Mechanism of vaccine delivery and intended site of vaccine delivery. All vaccines ultimately stimulate antigen presenting cells (APCs) which drain to lymph nodes to further activate the adaptive immune response. Epidermal, intradermal and intramuscular (IM) delivery routes rely on different, but related, populations of macrophages for adaptive immune system activation (on the left). Mucosal delivery is enhanced with specific M-cell stimulatory molecules (on the right). Created with BioRender.com.
Figure 6.
Mechanism of vaccine delivery and intended site of vaccine delivery. All vaccines ultimately stimulate antigen presenting cells (APCs) which drain to lymph nodes to further activate the adaptive immune response. Epidermal, intradermal and intramuscular (IM) delivery routes rely on different, but related, populations of macrophages for adaptive immune system activation (on the left). Mucosal delivery is enhanced with specific M-cell stimulatory molecules (on the right). Created with BioRender.com.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kozak, M.; Hu, J. DNA Vaccines: Their Formulations, Engineering and Delivery. Vaccines 2024, 12, 71. https://doi.org/10.3390/vaccines12010071
AMA Style
Kozak M, Hu J. DNA Vaccines: Their Formulations, Engineering and Delivery. Vaccines. 2024; 12(1):71. https://doi.org/10.3390/vaccines12010071
Chicago/Turabian StyleKozak, Michael, and Jiafen Hu. 2024. "DNA Vaccines: Their Formulations, Engineering and Delivery" Vaccines 12, no. 1: 71. https://doi.org/10.3390/vaccines12010071
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.