
Recombinant BCG-LTAK63 Vaccine Candidate for Tuberculosis Induces an Inflammatory Profile in Human Macrophages

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
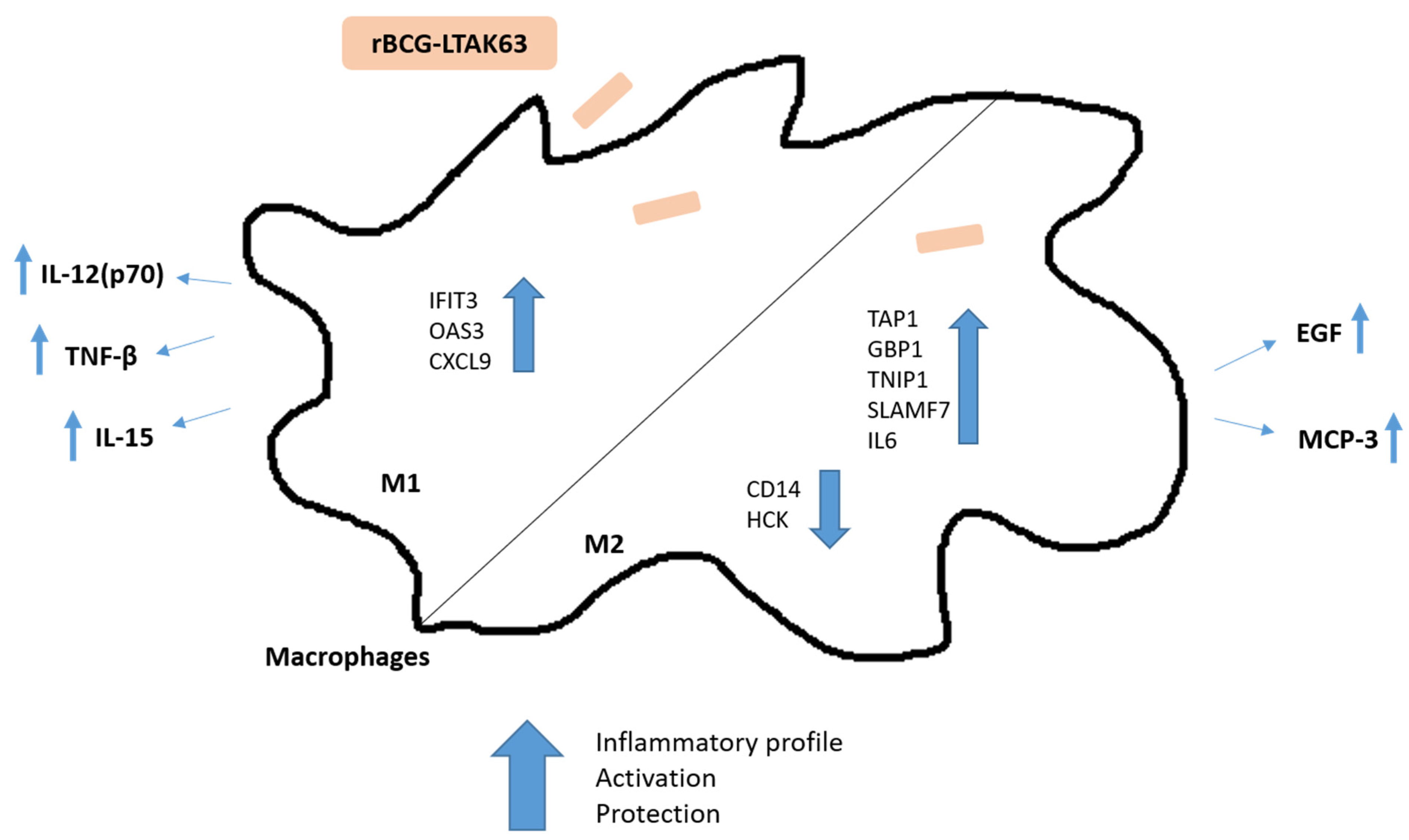
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture of Primary Human-Monocyte-Derived Macrophages
2.2. Bacterial Culture and Macrophage Infection
2.3. Intracellular Mycobacterial Survival Assay
2.4. RNA Extraction, dcRT-MLPA Assay, and Data Analysis
2.5. Cytokine Production
2.6. Statistical Analysis
3. Results
3.1. Intracellular Survival of rBCG-LTAK63 in M1 and M2 Macrophages Is Comparable to BCG
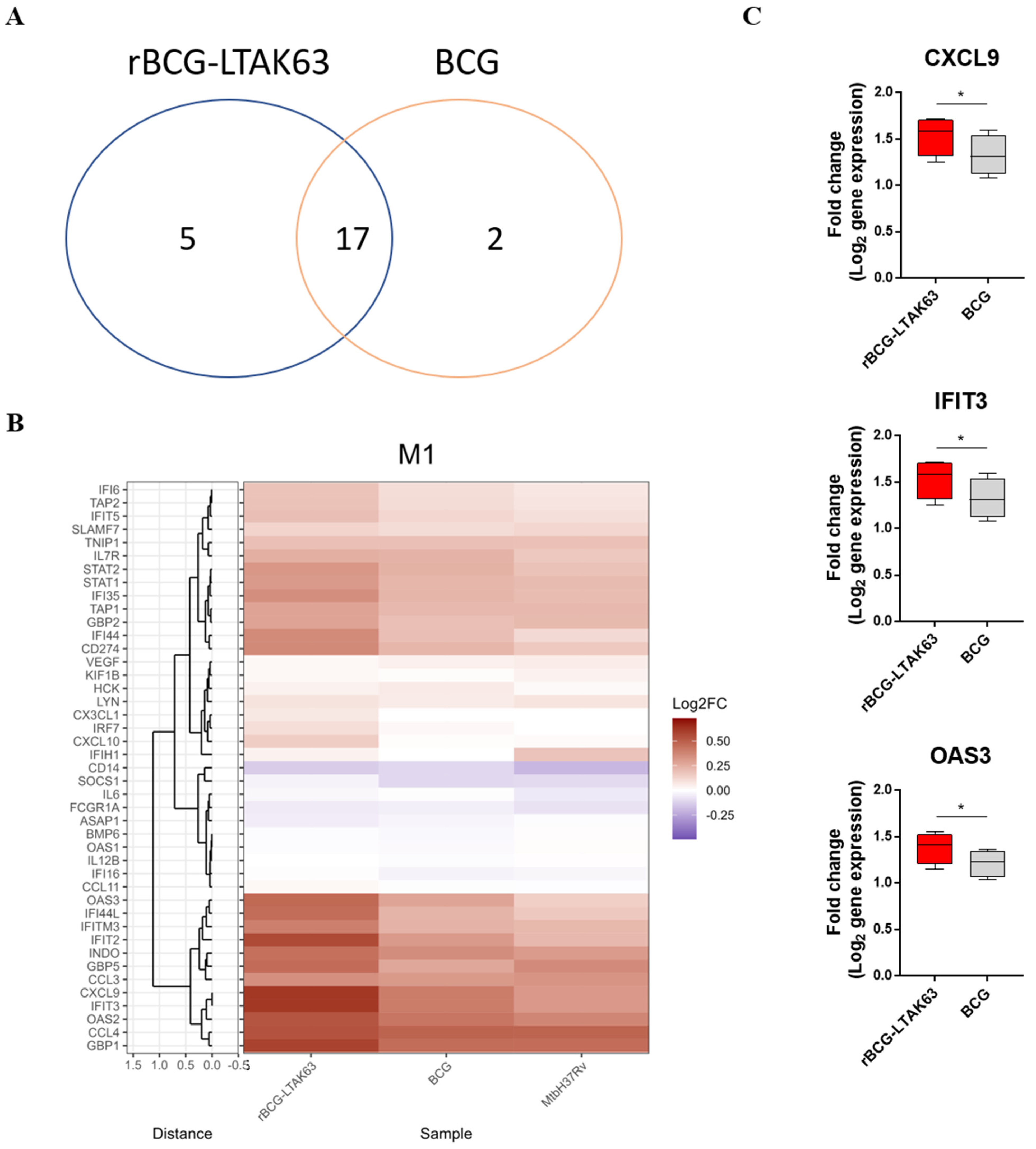
3.2. M1 Macrophages Infected with rBCG-LTAK63 Display Marked Upregulation of Genes Associated with the IFN Signaling Pathway
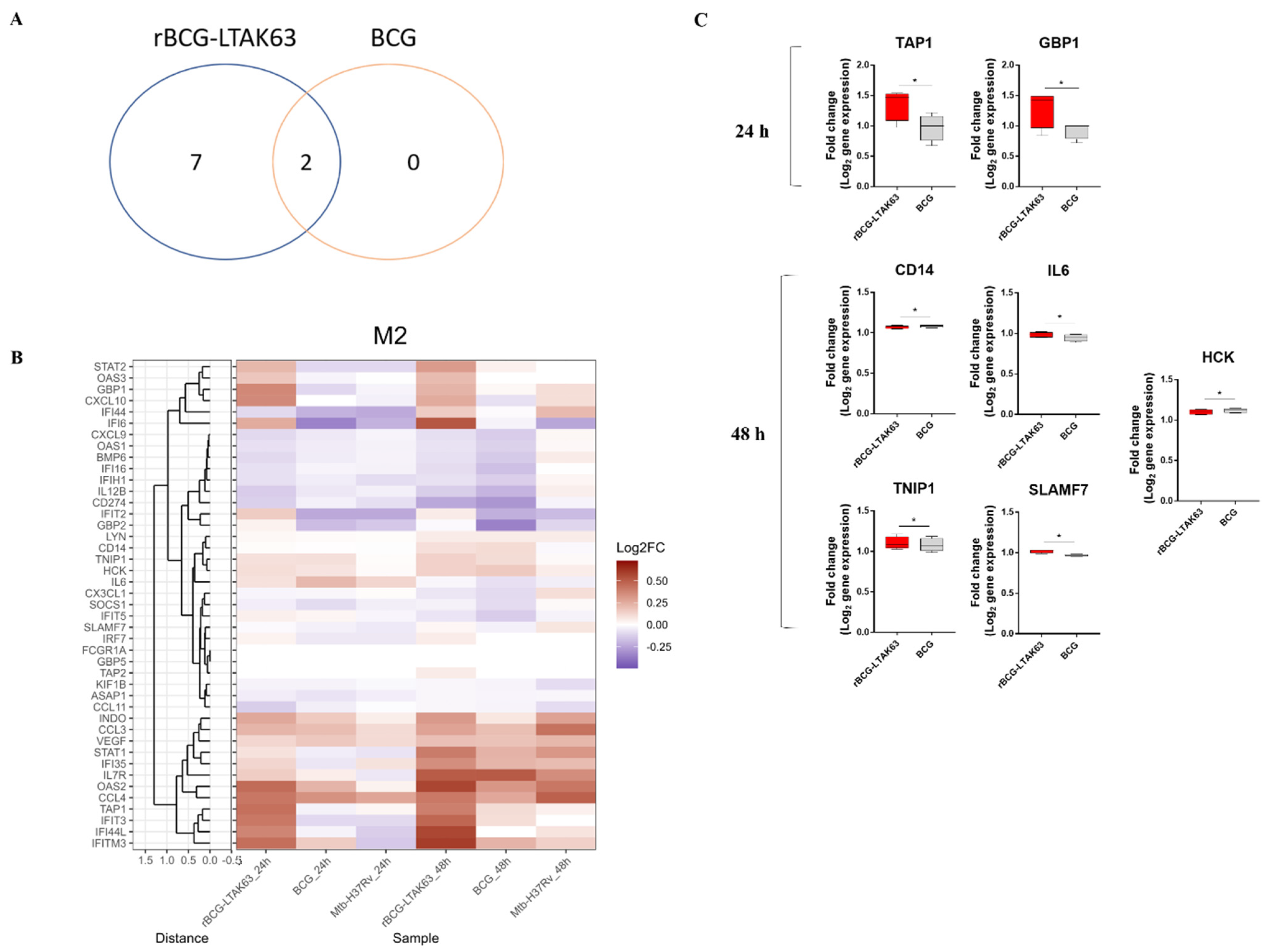
3.3. BCG Induces Downregulation of an Inflammatory Gene Profile in M2 Macrophages, While rBCG-LTAK63 Maintains or Induces Upregulation for 48 h
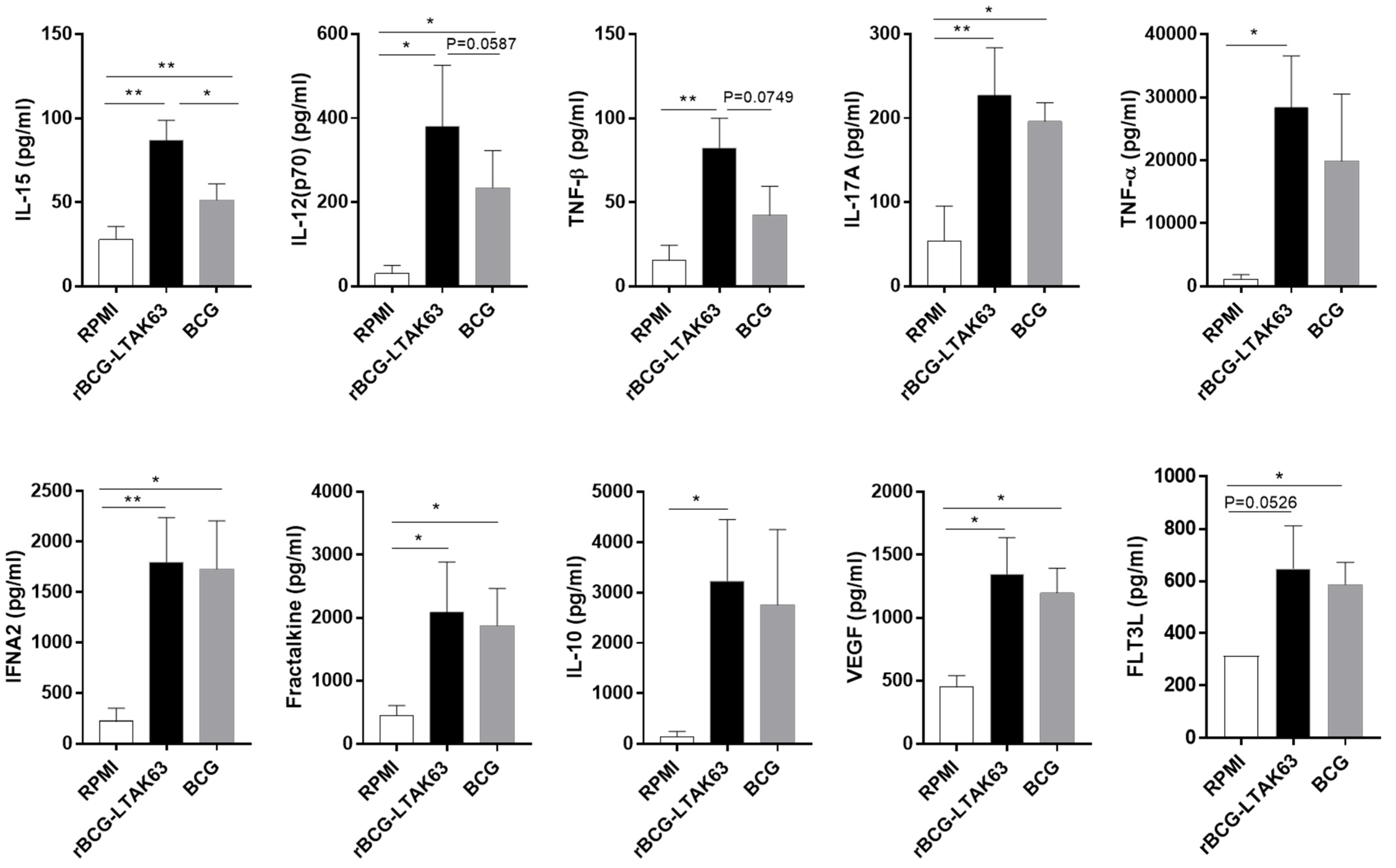
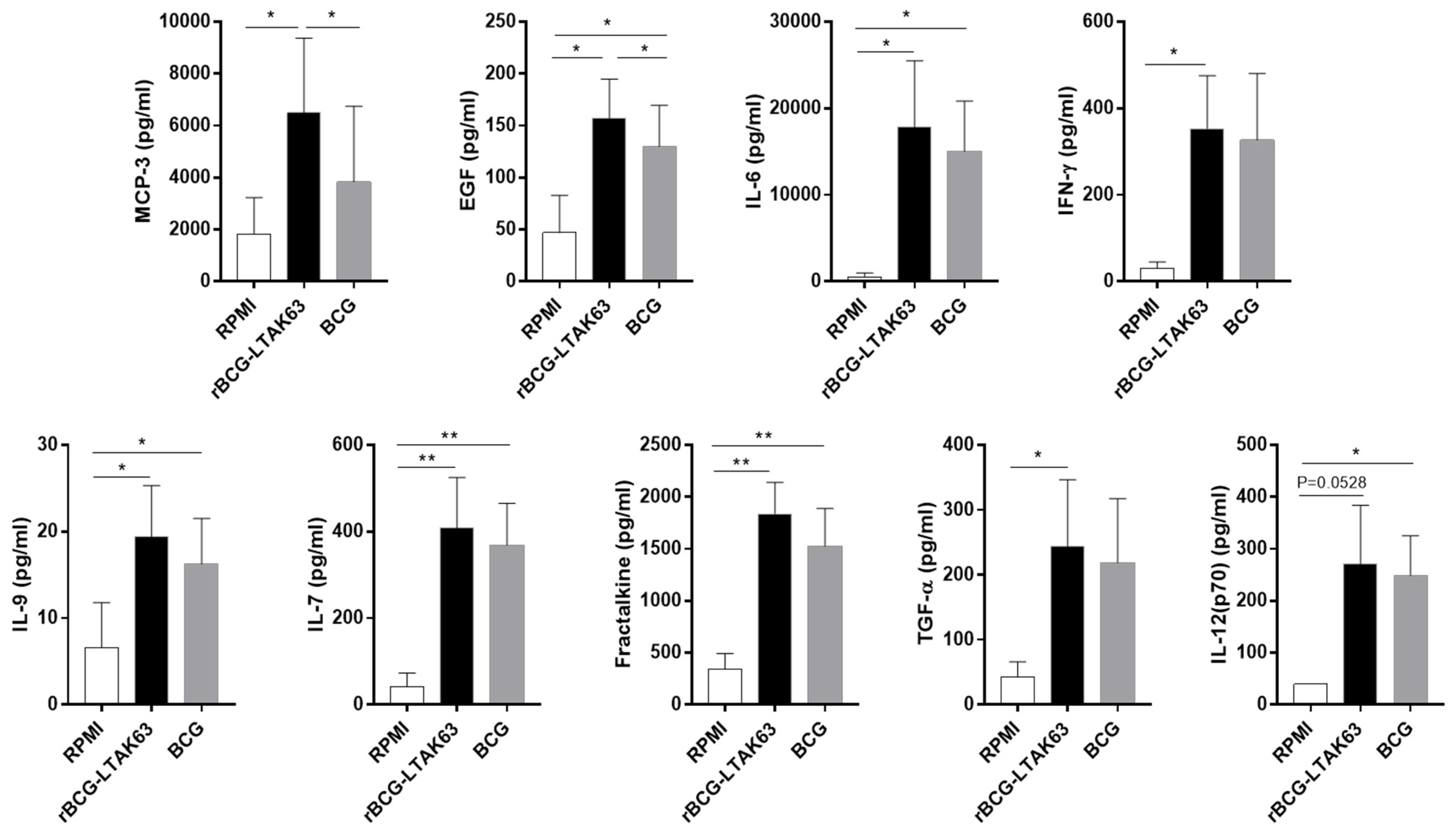
3.4. M1 and M2 Macrophages Infected with rBCG-LTAK63 Display Increased Production of Inflammatory Cytokines
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2021; World Health Organization: Geneva, Switzerland, 2021.
- Kaufmann, S.H.; Hussey, G.; Lambert, P.H. New vaccines for tuberculosis. Lancet 2010, 375, 2110–2119. [Google Scholar] [CrossRef]
- Jamwal, S.V.; Mehrotra, P.; Singh, A.; Siddiqui, Z.; Basu, A.; Rao, K.V. Mycobacterial escape from macrophage phagosomes to the cytoplasm represents an alternate adaptation mechanism. Sci. Rep. 2016, 6, 23089. [Google Scholar] [CrossRef] [PubMed]
- Gengenbacher, M.; Kaufmann, S.H. Mycobacterium tuberculosis: Success through dormancy. FEMS Microbiol. Rev. 2012, 36, 514–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoebe, K.; Janssen, E.; Beutler, B. The interface between innate and adaptive immunity. Nat. Immunol. 2004, 5, 971–974. [Google Scholar] [CrossRef]
- Glass, C.K.; Natoli, G. Molecular control of activation and priming in macrophages. Nat. Immunol. 2016, 17, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Mege, J.L.; Mehraj, V.; Capo, C. Macrophage polarization and bacterial infections. Curr. Opin. Infect. Dis. 2011, 24, 230–234. [Google Scholar] [CrossRef]
- Kelly, B.; O’Neill, L.A. Metabolic reprogramming in macrophages and dendritic cells in innate immunity. Cell Res. 2015, 25, 771–784. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Khader, S.A.; Divangahi, M.; Hanekom, W.; Hill, P.C.; Maeurer, M.; Makar, K.W.; Mayer-Barber, K.D.; Mhlanga, M.M.; Nemes, E.; Schlesinger, L.S.; et al. Targeting innate immunity for tuberculosis vaccination. J. Clin. Investig. 2019, 129, 3482–3491. [Google Scholar] [CrossRef] [Green Version]
- Nieuwenhuizen, N.E.; Kaufmann, S.H.E. Next-Generation Vaccines Based on Bacille Calmette-Guerin. Front. Immunol. 2018, 9, 121. [Google Scholar] [CrossRef]
- Nascimento, I.P.; Rodriguez, D.; Santos, C.C.; Amaral, E.P.; Rofatto, H.K.; Junqueira-Kipnis, A.P.; Goncalves, E.D.C.; D’Imperio-Lima, M.R.; Hirata, M.H.; Silva, C.L.; et al. Recombinant BCG Expressing LTAK63 Adjuvant induces Superior Protection against Mycobacterium tuberculosis. Sci. Rep. 2017, 7, 2109. [Google Scholar] [CrossRef] [PubMed]
- Rappuoli, R.; Pizza, M.; Douce, G.; Dougan, G. Structure and mucosal adjuvanticity of cholera and Escherichia coli heat-labile enterotoxins. Immunol. Today 1999, 20, 493–500. [Google Scholar] [CrossRef]
- Tritto, E.; Muzzi, A.; Pesce, I.; Monaci, E.; Nuti, S.; Galli, G.; Wack, A.; Rappuoli, R.; Hussell, T.; De Gregorio, E. The acquired immune response to the mucosal adjuvant LTK63 imprints the mouse lung with a protective signature. J. Immunol. 2007, 179, 5346–5357. [Google Scholar] [CrossRef] [PubMed]
- Moraes, L.; Trentini, M.M.; Fousteris, D.; Eto, S.F.; Chudzinski-Tavassi, A.M.; Leite, L.C.C.; Kanno, A.I. CRISPR/Cas9 Approach to Generate an Auxotrophic BCG Strain for Unmarked Expression of LTAK63 Adjuvant: A Tuberculosis Vaccine Candidate. Front. Immunol. 2022, 13, 867195. [Google Scholar] [CrossRef]
- Carvalho Dos Santos, C.; Rodriguez, D.; Kanno Issamu, A.; Cezar De Cerqueira Leite, L.; Pereira Nascimento, I. Recombinant BCG expressing the LTAK63 adjuvant induces increased early and long-term immune responses against Mycobacteria. Hum. Vaccines Immunother. 2020, 16, 673–683. [Google Scholar] [CrossRef]
- Verreck, F.A.; de Boer, T.; Langenberg, D.M.; van der Zanden, L.; Ottenhoff, T.H. Phenotypic and functional profiling of human proinflammatory type-1 and anti-inflammatory type-2 macrophages in response to microbial antigens and IFN-gamma- and CD40L-mediated costimulation. J. Leukoc. Biol. 2006, 79, 285–293. [Google Scholar] [CrossRef]
- Korbee, C.J.; Heemskerk, M.T.; Kocev, D.; van Strijen, E.; Rabiee, O.; Franken, K.; Wilson, L.; Savage, N.D.L.; Dzeroski, S.; Haks, M.C.; et al. Combined chemical genetics and data-driven bioinformatics approach identifies receptor tyrosine kinase inhibitors as host-directed antimicrobials. Nat. Commun. 2018, 9, 358. [Google Scholar] [CrossRef] [Green Version]
- Joosten, S.A.; Goeman, J.J.; Sutherland, J.S.; Opmeer, L.; de Boer, K.G.; Jacobsen, M.; Kaufmann, S.H.; Finos, L.; Magis-Escurra, C.; Ota, M.O. Identification of biomarkers for tuberculosis disease using a novel dual-color RT-MLPA assay. Genes Immun. 2012, 13, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Haks, M.C.; Goeman, J.J.; Magis-Escurra, C.; Ottenhoff, T.H. Focused human gene expression profiling using dual-color reverse transcriptase multiplex ligation-dependent probe amplification. Vaccine 2015, 33, 5282–5288. [Google Scholar] [CrossRef] [Green Version]
- Bastos, R.G.; Borsuk, S.; Seixas, F.K.; Dellagostin, O.A. Recombinant Mycobacterium bovis BCG. Vaccine 2009, 27, 6495–6503. [Google Scholar] [CrossRef]
- Sorensen, H.P.; Mortensen, K.K. Soluble expression of recombinant proteins in the cytoplasm of Escherichia coli. Microb. Cell Factories 2005, 4, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Costa, A.C.; Costa-Junior Ade, O.; de Oliveira, F.M.; Nogueira, S.V.; Rosa, J.D.; Resende, D.P.; Kipnis, A.; Junqueira-Kipnis, A.P. A new recombinant BCG vaccine induces specific Th17 and Th1 effector cells with higher protective efficacy against tuberculosis. PLoS ONE 2014, 9, e112848. [Google Scholar] [CrossRef] [PubMed]
- Sadler, A.J.; Williams, B.R. Interferon-inducible antiviral effectors. Nature reviews. Immunology 2008, 8, 559–568. [Google Scholar] [PubMed]
- Diamond, M.S.; Farzan, M. The broad-spectrum antiviral functions of IFIT and IFITM proteins. Nat. Rev. Immunol. 2013, 13, 46–57. [Google Scholar] [CrossRef] [PubMed]
- MacMicking, J.D. Interferon-inducible effector mechanisms in cell-autonomous immunity. Nat. Rev. Immunol. 2012, 12, 367–382. [Google Scholar] [CrossRef]
- Boxx, G.M.; Cheng, G. The Roles of Type I Interferon in Bacterial Infection. Cell Host Microbe 2016, 19, 760–769. [Google Scholar] [CrossRef] [Green Version]
- Kovarik, P.; Castiglia, V.; Ivin, M.; Ebner, F. Type I Interferons in Bacterial Infections: A Balancing Act. Front. Immunol. 2016, 7, 652. [Google Scholar] [CrossRef] [Green Version]
- Novikov, A.; Cardone, M.; Thompson, R.; Shenderov, K.; Kirschman, K.D.; Mayer-Barber, K.D.; Myers, T.G.; Rabin, R.L.; Trinchieri, G.; Sher, A.; et al. Mycobacterium tuberculosis triggers host type I IFN signaling to regulate IL-1beta production in human macrophages. J. Immunol. 2011, 187, 2540–2547. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Banerjee, S.; Wang, Y.; Goldstein, S.A.; Dong, B.; Gaughan, C.; Silverman, R.H.; Weiss, S.R. Activation of RNase L is dependent on OAS3 expression during infection with diverse human viruses. Proc. Natl. Acad. Sci. USA 2016, 113, 2241–2246. [Google Scholar] [CrossRef] [Green Version]
- Toledo Pinto, T.G.; Batista-Silva, L.R.; Medeiros, R.C.A.; Lara, F.A.; Moraes, M.O. Type I Interferons, Autophagy and Host Metabolism in Leprosy. Front. Immunol. 2018, 9, 806. [Google Scholar] [CrossRef] [Green Version]
- Han, J.H.; Suh, C.H.; Jung, J.Y.; Ahn, M.H.; Han, M.H.; Kwon, J.E.; Yim, H.; Kim, H.A. Elevated circulating levels of the interferon-gamma-induced chemokines are associated with disease activity and cutaneous manifestations in adult-onset Still’s disease. Sci. Rep. 2017, 7, 46652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loetscher, M.; Gerber, B.; Loetscher, P.; Jones, S.A.; Piali, L.; Clark-Lewis, I.; Baggiolini, M.; Moser, B. Chemokine receptor specific for IP10 and mig: Structure, function, and expression in activated T-lymphocytes. J. Exp. Med. 1996, 184, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Smit, M.J.; Verdijk, P.; van der Raaij-Helmer, E.M.; Navis, M.; Hensbergen, P.J.; Leurs, R.; Tensen, C.P. CXCR3-mediated chemotaxis of human T cells is regulated by a Gi- and phospholipase C-dependent pathway and not via activation of MEK/p44/p42 MAPK nor Akt/PI-3 kinase. Blood 2003, 102, 1959–1965. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Lu, P.; Xia, Y.; Ding, S.; Fan, Y.; Li, X.; Han, P.; Liu, J.; Tian, D.; Liu, M. CXCL9: Evidence and contradictions for its role in tumor progression. Cancer Med. 2016, 5, 3246–3259. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.D.; Lin, Y.; Moreno, J.R.; Randall, T.D.; Khader, S.A. Profiling early lung immune responses in the mouse model of tuberculosis. PLoS ONE 2011, 6, e16161. [Google Scholar] [CrossRef] [Green Version]
- Vignali, D.A.; Kuchroo, V.K. IL-12 family cytokines: Immunological playmakers. Nat. Immunol. 2012, 13, 722–728. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Samperio, P. Role of interleukin-12 family cytokines in the cellular response to mycobacterial disease. Int. J. Infect. Dis. 2010, 14, e366–e371. [Google Scholar] [CrossRef] [Green Version]
- Cooper, A.M.; Mayer-Barber, K.D.; Sher, A. Role of innate cytokines in mycobacterial infection. Mucosal Immunol. 2011, 4, 252–260. [Google Scholar] [CrossRef]
- Croft, M. The role of TNF superfamily members in T-cell function and diseases. Nature reviews. Immunology 2009, 9, 271–285. [Google Scholar]
- Parameswaran, N.; Patial, S. Tumor necrosis factor-alpha signaling in macrophages. Crit. Rev. Eukaryot. Gene Expr. 2010, 20, 87–103. [Google Scholar] [CrossRef]
- Maeurer, M.J.; Trinder, P.; Hommel, G.; Walter, W.; Freitag, K.; Atkins, D.; Storkel, S. Interleukin-7 or interleukin-15 enhances survival of Mycobacterium tuberculosis-infected mice. Infect. Immun. 2000, 68, 2962–2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruckert, R.; Brandt, K.; Ernst, M.; Marienfeld, K.; Csernok, E.; Metzler, C.; Budagian, V.; Bulanova, E.; Paus, R.; Bulfone-Paus, S. Interleukin-15 stimulates macrophages to activate CD4+ T cells: A role in the pathogenesis of rheumatoid arthritis? Immunology 2009, 126, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Mueller, Y.M.; Petrovas, C.; Bojczuk, P.M.; Dimitriou, I.D.; Beer, B.; Silvera, P.; Villinger, F.; Cairns, J.S.; Gracely, E.J.; Lewis, M.G.; et al. Interleukin-15 increases effector memory CD8+ t cells and NK Cells in simian immunodeficiency virus-infected macaques. J. Virol. 2005, 79, 4877–4885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stonier, S.W.; Ma, L.J.; Castillo, E.F.; Schluns, K.S. Dendritic cells drive memory CD8 T-cell homeostasis via IL-15 transpresentation. Blood 2008, 112, 4546–4554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, S.; Crotta, S.; McCabe, T.M.; Wack, A. Pathogenic potential of interferon alphabeta in acute influenza infection. Nat. Commun. 2014, 5, 3864. [Google Scholar] [CrossRef] [Green Version]
- Eggensperger, S.; Tampe, R. The transporter associated with antigen processing: A key player in adaptive immunity. Biol. Chem. 2015, 396, 1059–1072. [Google Scholar] [CrossRef]
- Lankat-Buttgereit, B.; Tampe, R. The transporter associated with antigen processing: Function and implications in human diseases. Physiol. Rev. 2002, 82, 187–204. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.H.; Shenoy, A.R.; Kumar, P.; Das, R.; Tiwari, S.; MacMicking, J.D. A family of IFN-gamma-inducible 65-kD GTPases protects against bacterial infection. Science 2011, 332, 717–721. [Google Scholar] [CrossRef]
- Li, P.; Jiang, W.; Yu, Q.; Liu, W.; Zhou, P.; Li, J.; Xu, J.; Xu, B.; Wang, F.; Shao, F. Ubiquitination and degradation of GBPs by a Shigella effector to suppress host defence. Nature 2017, 551, 378–383. [Google Scholar] [CrossRef]
- Leavy, O. Innate immunity: GBPs coordinate vesicular trafficking for host defence. Nature reviews. Immunology 2011, 11, 372–373. [Google Scholar]
- Qiu, X.; Guo, H.; Yang, J.; Ji, Y.; Wu, C.S.; Chen, X. Down-regulation of guanylate binding protein 1 causes mitochondrial dysfunction and cellular senescence in macrophages. Sci. Rep. 2018, 8, 1679. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yan, H.; Song, Z.; Chen, F.; Wang, H.; Niu, J.; Shi, X.; Zhang, D.; Zhang, N.; Zhai, Z.; et al. Downregulation of TNIP1 Expression Leads to Increased Proliferation of Human Keratinocytes and Severer Psoriasis-Like Conditions in an Imiquimod-Induced Mouse Model of Dermatitis. PLoS ONE 2015, 10, e0127957. [Google Scholar]
- Gurevich, I.; Zhang, C.; Francis, N.; Aneskievich, B.J. TNIP1, a retinoic acid receptor corepressor and A20-binding inhibitor of NF-kappaB, distributes to both nuclear and cytoplasmic locations. J. Histochem. Cytochem. 2011, 59, 1101–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, A.N.; Mehra, S.; Kaushal, D. Role of interleukin 6 in innate immunity to Mycobacterium tuberculosis infection. J. Infect. Dis. 2013, 207, 1253–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comte, D.; Karampetsou, M.P.; Yoshida, N.; Kis-Toth, K.; Kyttaris, V.C.; Tsokos, G.C. Signaling Lymphocytic Activation Molecule Family Member 7 Engagement Restores Defective Effector CD8+ T Cell Function in Systemic Lupus Erythematosus. Arthritis Rheumatol. 2017, 69, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Malaer, J.D.; Mathew, P.A. CS1 (SLAMF7, CD319) is an effective immunotherapeutic target for multiple myeloma. Am. J. Cancer Res. 2017, 7, 1637–1641. [Google Scholar]
- Jia, T.; Serbina, N.V.; Brandl, K.; Zhong, M.X.; Leiner, I.M.; Charo, I.F.; Pamer, E.G. Additive roles for MCP-1 and MCP-3 in CCR2-mediated recruitment of inflammatory monocytes during Listeria monocytogenes infection. J. Immunol. 2008, 180, 6846–6853. [Google Scholar] [CrossRef] [Green Version]
- Lu, N.; Wang, L.; Cao, H.; Liu, L.; Van Kaer, L.; Washington, M.K.; Rosen, M.J.; Dube, P.E.; Wilson, K.T.; Ren, X.; et al. Activation of the epidermal growth factor receptor in macrophages regulates cytokine production and experimental colitis. J. Immunol. 2014, 192, 1013–1023. [Google Scholar] [CrossRef]
- Blischak, J.D.; Tailleux, L.; Mitrano, A.; Barreiro, L.B.; Gilad, Y. Mycobacterial infection induces a specific human innate immune response. Sci. Rep. 2015, 5, 16882. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos, C.C.; Walburg, K.V.; van Veen, S.; Wilson, L.G.; Trufen, C.E.M.; Nascimento, I.P.; Ottenhoff, T.H.M.; Leite, L.C.C.; Haks, M.C. Recombinant BCG-LTAK63 Vaccine Candidate for Tuberculosis Induces an Inflammatory Profile in Human Macrophages. Vaccines 2022, 10, 831. https://doi.org/10.3390/vaccines10060831
dos Santos CC, Walburg KV, van Veen S, Wilson LG, Trufen CEM, Nascimento IP, Ottenhoff THM, Leite LCC, Haks MC. Recombinant BCG-LTAK63 Vaccine Candidate for Tuberculosis Induces an Inflammatory Profile in Human Macrophages. Vaccines. 2022; 10(6):831. https://doi.org/10.3390/vaccines10060831
Chicago/Turabian Styledos Santos, Carina C., Kimberley V. Walburg, Suzanne van Veen, Louis G. Wilson, Carlos E. M. Trufen, Ivan P. Nascimento, Tom H. M. Ottenhoff, Luciana C. C. Leite, and Mariëlle C. Haks. 2022. "Recombinant BCG-LTAK63 Vaccine Candidate for Tuberculosis Induces an Inflammatory Profile in Human Macrophages" Vaccines 10, no. 6: 831. https://doi.org/10.3390/vaccines10060831