
Detection of HBV Covalently Closed Circular DNA


Abstract
:1. Introduction
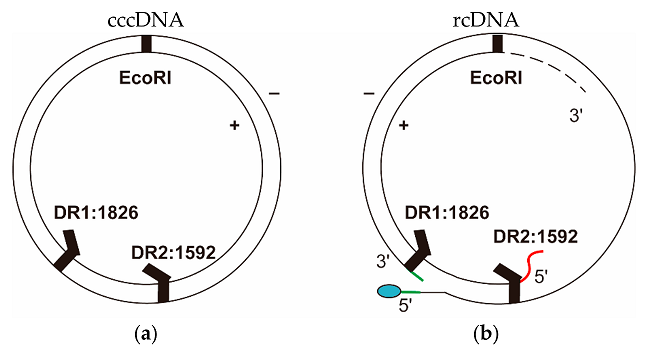
2. Formation and Structure of cccDNA
3. Preparation of cccDNA Samples
4. cccDNA Detection and Quantification
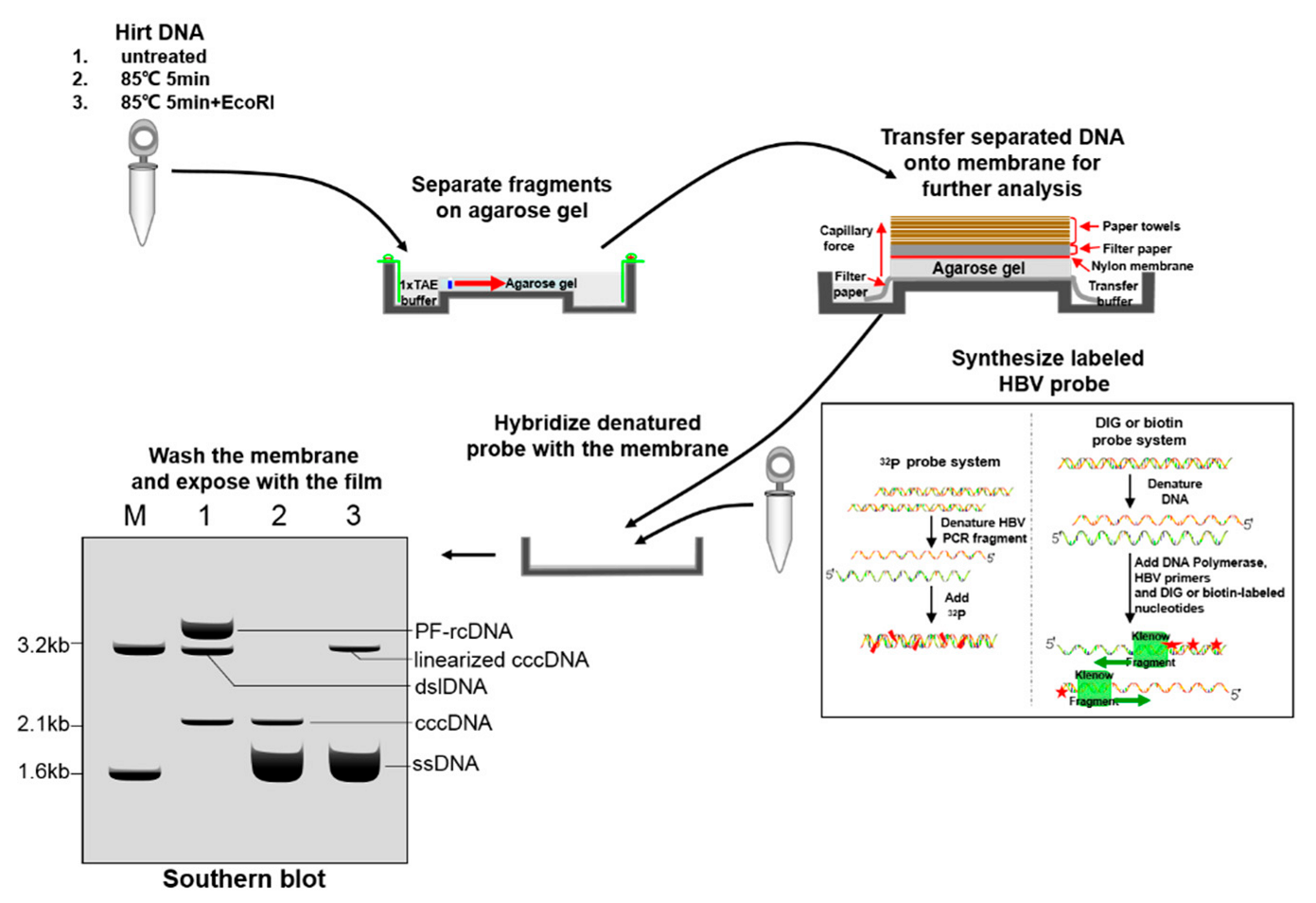
4.1. Southern Blot
4.2. Real-Time qPCR
4.2.1. Conventional qPCR
4.2.2. Competitive qPCR
4.2.3. Semi-Nested and Nested qPCR
4.2.4. Droplet-Digital PCR
4.3. Rolling Circle Amplification qPCR
4.4. Rolling Circle Amplification-In Situ qPCR
4.5. Magnetic Capture Hybridization qPCR
4.6. Invader Assay
4.7. In Situ Hybridization
4.8. Surrogate Markers of cccDNA
5. Conclusions and Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Solà, I.; Comas, D.R.; Wakefeld, V.; Karner, C.; Tsochatzis, E. Guidelines for the Prevention, Care and Treatment of Persons with Chronic Hepatitis B Infection; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Liang, T.J.; Block, T.M.; McMahon, B.J.; Ghany, M.G.; Urban, S.; Guo, J.T.; Locarnini, S.; Zoulim, F.; Chang, K.M.; Lok, A.S. Present and future therapies of hepatitis B: From discovery to cure. Hepatology 2015, 62, 1893–1908. [Google Scholar] [CrossRef] [PubMed]
- Revill, P.; Locarnini, S. Antiviral strategies to eliminate hepatitis B virus covalently closed circular DNA (cccDNA). Curr. Opin. Pharmacol. 2016, 30, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Ward, H.; Tang, L.; Poonia, B.; Kottilil, S. Treatment of hepatitis B virus an update. Future Microbiol. 2016, 11, 1581–1597. [Google Scholar] [CrossRef] [PubMed]
- Lampertico, P.; Agarwal, K.; Berg, T.; Buti, M.; Janssen, H.L.; Papatheodoridis, G.; Zoulim, F.; Tacke, F. EASL 2017 Clinical Practice Guidelines on the management of hepatitis B virus infection. J. Hepatol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Varbobitis, I.; Papatheodoridis, G.V. The assessment of hepatocellular carcinoma risk in patients with chronic hepatitis B under antiviral therapy. Clin. Mol. Hepatol. 2016, 22, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Su, T.H.; Hu, T.H.; Chen, C.Y.; Huang, Y.H.; Chuang, W.L.; Lin, C.C.; Wang, C.C.; Su, W.W.; Chen, M.Y.; Peng, C.Y.; et al. Four-year entecavir therapy reduces hepatocellular carcinoma, cirrhotic events and mortality in chronic hepatitis B patients. Liver Int. 2016, 36, 1755–1764. [Google Scholar] [CrossRef] [PubMed]
- Hoang, J.K.; Yang, H.I.; Le, A.; Nguyen, N.H.; Lin, D.; Vu, V.D.; Chaung, K.; Nguyen, V.; Trinh, H.N.; Li, J.Y.; et al. Lower liver cancer risk with antiviral therapy in chronic hepatitis B patients with normal to minimally elevated ALT and no cirrhosis. Medicine (Baltimore) 2016, 95, e4433. [Google Scholar] [CrossRef] [PubMed]
- Papatheodoridis, G.V.; Chan, H.L.; Hansen, B.E.; Janssen, H.L.; Lampertico, P. Risk of hepatocellular carcinoma in chronic hepatitis B: Assessment and modification with current antiviral therapy. J. Hepatol. 2015, 62, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Durantel, D.; Zoulim, F. New antiviral targets for innovative treatment concepts for hepatitis B virus and hepatitis delta virus. J. Hepatol. 2016, 64, S117–S131. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sze, J.; He, M.L. HBV cccDNA in patient’s sera as an indicator for HBV reactivation and an early signal of liver damage. World J. Gastroenterol. 2004, 10, 82–85. [Google Scholar] [PubMed]
- Zoulim, F. New insight on hepatitis B virus persistence from the study of intrahepatic viral cccDNA. J. Hepatol. 2005, 42, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.J.; Wong, M.L.; Bowden, S.; Liew, C.T.; Hui, A.Y.; Wong, V.W.; Leung, N.W.; Locarnini, S.; Chan, H.L.; et al. Intrahepatic hepatitis B virus covalently closed circular DNA can be a predictor of sustained response to therapy. Gastroenterology 2005, 128, 1890–1897. [Google Scholar] [CrossRef] [PubMed]
- Levrero, M.; Pollicino, T.; Petersen, J.; Belloni, L.; Raimondo, G.; Dandri, M. Control of cccDNA function in hepatitis B virus infection. J. Hepatol. 2009, 51, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.T.; Guo, H. Metabolism and function of hepatitis B virus cccDNA: Implications for the development of cccDNA-targeting antiviral therapeutics. Antivir. Res. 2015, 122, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Nassal, M. HBV cccDNA viral persistence reservoir and key obstacle for a cure of chronic hepatitis B. Gut 2015, 64, 1972–1984. [Google Scholar] [CrossRef] [PubMed]
- Fung, J. HBV therapeutic vaccines and cccDNA inhibitors—Emergence of a cure. Liver Transpl. 2016, 22, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Lucifora, J.; Protzer, U. Attacking hepatitis B virus cccDNA—The holy grail to hepatitis B cure. J. Hepatol. 2016, 64, S41–S48. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.; Thompson, A.J.; Levrero, M. Aiming for cure in HBV and HDV infection. J. Hepatol. 2016, 65, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Mu, D.; Yan, L.; Tang, H.; Liao, Y. A sensitive and accurate quantification method for the detection of hepatitis B virus covalently closed circular DNA by the application of a droplet digital polymerase chain reaction amplification system. Biotechnol. Lett. 2015, 37, 2063–2073. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.N.; Lu, W.; Zheng, Y.; Wang, W.X.; Bai, L.; Chen, L.; Feng, Y.L.; Zhang, Z.Q.; Yuan, Z.H. In situ analysis of intrahepatic virological events in chronic hepatitis B virus infection. J. Clin. Investig. 2016, 126, 1079–1092. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.K.; Seto, W.K.; Cheung, K.S.; Chong, C.K.; Huang, F.Y.; Fung, J.; Lai, C.L.; Yuen, M.F. Hepatitis B virus core-related antigen as a surrogate marker for covalently closed circular DNA. Liver Int. 2016. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Cheng, L.; Murphy, C.M.; Reszka-Blanco, N.J.; Wu, Y.X.; Chi, L.Q.; Hu, J.M.; Su, L.S. Minicircle HBV cccDNA with a Gaussia luciferase reporter for investigating HBV cccDNA biology and developing cccDNA-targeting drugs. Sci. Rep. 2016, 6, 36483. [Google Scholar] [CrossRef] [PubMed]
- Giersch, K.; Allweiss, L.; Volz, T.; Dandri, M.; Lutgehetmann, M. Serum HBV pgRNA as a clinical marker for cccDNA activity. J. Hepatol. 2016, 66, 460–462. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Wang, X.H.; Yan, R.; Mao, R.C.; Liu, Y.J.; Ji, C.H.; Cuconati, A.; Guo, H.T. Establishment of an inducible HBV stable cell line that expresses cccDNA-dependent epitope-tagged HBeAg for screening of cccDNA modulators. Antivir. Res. 2016, 132, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Tajik, Z.; Keyvani, H.; Bokharaei-Salim, F.; Zolfaghari, M.R.; Fakhim, S.; Keshvari, M.; Alavian, S.M. Detection of hepatitis B virus covalently closed circular DNA in the plasma of Iranian HBeAg-negative patients with chronic hepatitis B. Hepat. Mon. 2015, 15, e30790. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Sheng, S.; Nie, B.; Tu, Z. Development of magnetic capture hybridization and quantitative polymerase chain reaction for hepatitis B virus covalently closed circular DNA. Hepat. Mon. 2015, 15, e23729. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.W.; Hu, S.Y.; Xu, C.; Zhao, Y.L.; Xu, D.P.; Zhao, Y.Q.; Zhao, J.M.; Li, Z.B.; Zhang, X.C.; Zhang, H.F.; et al. A novel method for detection of HBVcccDNA in hepatocytes using rolling circle amplification combined with in situ PCR. BMC Infect. Dis. 2014, 14, 608. [Google Scholar] [CrossRef] [PubMed]
- Ogura, N.; Watashi, K.; Noguchi, T.; Wakita, T. Formation of covalently closed circular DNA in Hep38.7-Tet cells, a tetracycline inducible hepatitis B virus expression cell line. Biochem. Biophys. Res. Commun. 2014, 452, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.W.; Han, J.Q.; Zou, Z.S.; Liu, S.H.; Tang, B.; Ren, X.Q.; Li, X.D.; Zhao, Y.L.; Liu, Y.; Zhou, D.Q.; et al. Quantitation of HBV covalently closed circular DNA in micro formalin fixed paraffin-embedded liver tissue using rolling circle amplification in combination with real-time PCR. Clin. Chim. Acta 2011, 412, 1905–1911. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.H.; Li, Z.S.; Dai, J.Y.; Zhu, H.Y.; Yu, J.W.; Lu, S.L. Nested real-time quantitative polymerase chain reaction assay for detection of hepatitis B virus covalently closed circular DNA. Chin. Med. J. 2011, 124, 1513–1516. [Google Scholar] [PubMed]
- Takkenberg, R.B.; Zaaijer, H.L.; Menting, S.; Weegink, C.J.; Terpstra, V.; Cornelissen, M.; Dijkgraaf, M.G.; Jansen, D.L.; Reesink, H.W.; Beld, M.G. Detection of hepatitis B virus covalently closed circular DNA in paraffin-embedded and cryo-preserved liver biopsies of chronic hepatitis B patients. Eur. J. Gastroenterol. Hepatol. 2010, 22, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.T.; Han, T.; Li, Y.; Yang, B.; Wang, Y.J.; Wang, F.M.; Jing, X.; Du, Z. Enhanced specificity of real-time PCR for measurement of hepatitis B virus cccDNA using restriction endonuclease and plasmid-safe ATP-dependent DNase and selective primers. J. Virol. Methods 2010, 169, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Takkenberg, R.B.; Zaaijer, H.L.; Molenkamp, R.; Menting, S.; Terpstra, V.; Weegink, C.J.; Dijkgraaf, M.G.; Jansen, D.L.; Reesink, H.W.; Beld, M.G. Validation of a sensitive and specific real-time PCR for detection and quantitation of hepatitis B virus covalently closed circular DNA in plasma of chronic hepatitis B patients. J. Med. Virol. 2009, 81, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Bourne, E.J.; Dienstag, J.L.; Lopez, V.A.; Sander, T.J.; Longlet, J.M.; Hall, J.G.; Kwiatkowski, R.W.; Wright, T.; Lai, C.J.; Condreay, L.D. Quantitative analysis of HBV cccDNA from clinical specimens: Correlation with clinical and virological response during antiviral therapy. J. Viral. Hepat. 2007, 14, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.L.; Guo, H.T.; Guo, J.T.; Cuconati, A.; Mehta, A.; Block, T.M. Hepatitis B virus e antigen production is dependent upon covalently closed circular (ccc) DNA in HepAD38 cell cultures and may serve as a cccDNA surrogate in antiviral screening assays. Antivir. Res. 2006, 72, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.K.; Yuen, M.F.; Poon, R.T.; Yuen, J.C.; Fung, J.; Lai, C.L. Quantification of hepatitis B virus covalently closed circular DNA in patients with hepatocellular carcinoma. J. Hepatol. 2006, 45, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Mazet-Wagner, A.A.; Baclet, M.C.; Loustaud-Ratti, V.; Denis, F.; Alain, S. Real-time PCR quantitation of hepatitis B virus total DNA and covalently closed circular DNA in peripheral blood mononuclear cells from hepatitis B virus-infected patients. J. Virol. Methods 2006, 138, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Bowden, S.; Jackson, K.; Littlejohn, M.; Locarnini, S. Quantification of HBV covalently closed circular DNA from liver tissue by real-time PCR. Methods Mol. Med. 2004, 95, 41–50. [Google Scholar] [PubMed]
- Wong, D.K.; Yuen, M.F.; Yuan, H.J.; Sum, S.S.; Hui, C.K.; Hall, J.; Lai, C.L. Quantitation of covalently closed circular hepatitis B virus DNA in chronic hepatitis B patients. Hepatology 2004, 40, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Dicaire, A.; Wakil, A.E.; Luscombe, C.; Sacks, S.L. Quantitation of hepatitis B virus (HBV) covalently closed circular DNA (cccDNA) in the liver of HBV-infected patients by LightCycler real-time PCR. J. Virol. Methods 2004, 118, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.B.; Chen, Z.; Ni, W.Q.; Fan, J. A quantitative method to detect HBV cccDNA by chimeric primer and real-time polymerase chain reaction. J. Virol. Methods 2003, 112, 45–52. [Google Scholar]
- He, M.L.; Wu, J.; Chen, Y.; Lin, M.C.; Lau, G.K.; Kung, H.F. A new and sensitive method for the quantification of HBV cccDNA by real-time PCR. Biochem. Biophys. Res. Commun. 2002, 295, 1102–1107. [Google Scholar] [CrossRef]
- Littlejohn, M.; Bowden, S.; Jackson, K. Selective PCR for hepatitis B virus CCC DNA: Validation of real-time assays. Hepatology 2001, 34, 555. [Google Scholar]
- Addison, W.R.; Wong, W.W.; Fischer, K.P.; Tyrrell, D.L. A quantitative competitive PCR assay for the covalently closed circular form of the duck heppatitis B virus. Antivir. Res. 2000, 48, 27–37. [Google Scholar] [CrossRef]
- Kock, J.; Schlicht, H.J. Analysis of the earliest steps of hepadnavirus replication: Genome repair after infectious entry into hepatocytes does not depend on viral polymerase activity. J. Virol. 1993, 67, 4867–4874. [Google Scholar] [PubMed]
- Yan, H.; Zhong, G.C.; Xu, G.W.; He, W.H.; Jing, Z.Y.; Gao, Z.C.; Huang, Y.; Qi, Y.H.; Peng, B.; Wang, H.M. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 1, e00049. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Lempp, F.A.; Mehrle, S.; Nkongolo, S.; Kaufman, C.; Falth, M.; Stindt, J.; Koniger, C.; Nassal, M.; Kubitz, R.; et al. Hepatitis B and D viruses exploit sodium taurocholate co-transporting polypeptide for species-specific entry into hepatocytes. Gastroenterology 2014, 146, 1070–1083. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Hu, J. Formation of hepatitis B virus covalently closed circular DNA: Removal of genome-linked protein. J. Virol. 2007, 81, 6164–6174. [Google Scholar] [CrossRef] [PubMed]
- Bock, C.T.; Schranz, P.; Schroder, C.H.; Zentgraf, H. Hepatitis B virus genome is organized into nucleosomes in the nucleus of the infected cell. Virus Genes 1994, 8, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Newbold, J.E.; Xin, H.; Tencza, M.; Sherman, G.; Dean, J.; Bowden, S.; Locarnini, S. The covalently closed duplex form of the hepadnavirus genome exists in situ as a heterogeneous population of viral minichromosomes. J. Virol. 1995, 69, 3350–3357. [Google Scholar] [PubMed]
- Seeger, C.; Mason, W.S. Molecular biology of hepatitis B virus infection. Virology 2015, 479, 672–686. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J.A.; Litwin, S.; Seeger, C. Mechanism for CCC DNA synthesis in hepadnaviruses. PLoS ONE 2009, 4, e8093. [Google Scholar] [CrossRef] [PubMed]
- Pollicino, T.; Belloni, L.; Raffa, G.; Dediconi, N.; Squardrito, G.; Raimondo, G.; Levrero, M. Hepatitis B virus replication is regulated by the acetylation status of hepatitis B virus cccDNA-bound H3 and H4 histones. Gastroenterology 2006, 130, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.T.; Coates, L.; Aldrich, C.E.; Summers, J.; Mason, W.S. In hepatocytes infected with duck hepatitis B virus, the template for viral RNA synthesis is amplified by an intracellular pathway. Virology 1990, 175, 255–261. [Google Scholar] [CrossRef]
- Kock, J.; Rosler, C.; Zhang, J.J.; Blum, H.M.; Nassal, M.; Thoma, C. Generation of covalently closed circular DNA of hepatitis B viruses via intracellular recycling is regulated in a virus specific manner. PLoS Pathog. 2010, 6, e1001082. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.T.; Jiang, D.; Zhou, T.L.; Cuconati, A.; Block, T.M.; Guo, J.T. Characterization of the intracellular deproteinized relaxed circular DNA of hepatitis B virus: An intermediate of covalently closed circular DNA formation. J. Virol. 2007, 81, 12472–12484. [Google Scholar] [CrossRef] [PubMed]
- Tuttleman, J.S.; Pourcel, C.; Summers, J. Formation of the pool of cccDNA in hepadnavirus-infected cells. Cell 1986, 47, 451–460. [Google Scholar] [CrossRef]
- Guo, H.; Xu, C.; Zhou, T.; Block, T.M.; Guo, J.T. Characterization of the host factors required for hepadnavirus covalently closed circular (ccc) DNA formation. PLoS ONE 2012, 7, e43270. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Summers, J. Illegitimate replication of linear hepadnavirus DNA through nonhomologous. J. Virol. 1995, 69, 4029–4036. [Google Scholar] [PubMed]
- Staprans, S.; Loeb, D.D.; Ganem, D. Mutations affecting hepadnavirus plus-strand DNA synthesis dissociate primer cleavage from translocation and reveal the origin of linear viral DNA. J. Virol. 1991, 65, 1255–1262. [Google Scholar] [PubMed]
- Bock, C.T.; Schwinn, S.; Locarnini, S.; Fyfe, J.; Manns, M.P.; Trautwein, C.; Zentgraf, H. Structural organization of the hepatitis B virus. J. Mol. Biol. 2001, 307, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Werle–Lapostolle, B.; Bowden, S.; Locarnini, S.; Wursthorn, K.; Petersen, J.; Lau, G.K.; Trepo, C.; Marcellin, P.; Goodman, Z.; IV, W.E.; et al. Persistence of cccDNA during the natural history of chronic hepatitis B and decline during adefovir dipivoxil therapy. Gastroenterology 2004, 126, 1750–1758. [Google Scholar] [CrossRef] [PubMed]
- Galibert, F.; Mandart, E.; Fitoussi, F.; Tiollais, P.; Charnay, P. Nucleotide sequence of the hepatitis B virus genome (subtype ayw) cloned in E. coli. Nature 1979, 281, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Mabit, H.; Vons, C.; Dubanchet, S.; Capel, F.; Franco, D.; Petit, M.M. Primary cultured normal human hepatocytes for hepatitis B virus receptor studies. J. Hepatol. 1996, 24, 403–412. [Google Scholar] [CrossRef]
- Sumers, J.; Smith, P.M.; Horwich, A.L. Hepadnavirus envelope proteins regulate covalently closed circular DNA amplification. J. Virol. 1990, 64, 2819–2824. [Google Scholar]
- Xia, Y.C.; Stadler, D.; Ko, C.; Protzer, U. Analyses of HBV cccDNA quantification and modification. In Hepatitis B Virus: Methods and Protocols; Methods in Molecular Biology; Guo, H.T., Cuconati, A., Eds.; Springer: New York, NY, USA, 2017; Volume 1540, pp. 59–72. [Google Scholar]
- Lucifora, J.; Xia, Y.C.; Reisinger, F.; Zhang, K.; Stadler, D.; Cheng, X.M.; Sprinzl, M.F.; Koppensteiner, H.; Makowska, Z.; Volz, T.; et al. Specific and nonhepatotoxic degradation of nuclear hepatitis B virus cccDNA. Science 2014, 343, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Laras, A.; Koskinas, J.; Dimou, E.; Kostamena, A.; Hadziyannis, S.J. Intrahepatic levels and replicative activity of covalently closed circular hepatitis B virus DNA in chronically infected patients. Hepatology 2006, 44, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Hirt, B. Selective extraction of polyoma DNA from infected mouse cell cultures. J. Mol. Biol. 1967, 26, 365–369. [Google Scholar] [CrossRef]
- Chou, Y.C.; Jeng, K.S.; Chen, M.L.; Liu, H.H.; Liu, T.L.; Chen, Y.L.; Liu, Y.C.; Hu, C.P.; Chang, C.M. Evaluation of transcriptional efficiency of hepatitis B virus covalently closed circular DNA by reverse transcription-PCR combined with the restriction enzyme digestion method. J. Virol. 2005, 79, 1813–1823. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.L.; Guo, J.T.; Nunes, F.A.; Molnar-Kimber, K.L.; Wilson, J.M.; Aldrich, C.E.; Saputelli, J.; Litwin, S.; Condreay, L.D.; Seeger, C.; et al. Combination therapy with lamivudine and adenovirus causes transient suppression of chronic woodchuck hepatitis virus infections. J. Virol. 2000, 74, 11754–11763. [Google Scholar] [CrossRef] [PubMed]
- Moraleda, G.; Saputelli, J.; Aldrich, C.E.; Averett, D.; Condreay, L.; Mason, W. Lack of effect of antiviral therapy in nondividing hepatocyte. J. Virol. 1997, 71, 9392–9399. [Google Scholar] [PubMed]
- Arad, U. Modified Hirt procedure for rapid purification of extrachromosomal DNA from mammalian cells. Biotechniques 1998, 24, 760–762. [Google Scholar] [PubMed]
- Birnbiom, H.C.; Doly, J. A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 1979, 7, 1513–1523. [Google Scholar]
- Yang, W.; Mason, W.S.; Summers, J. Covalently closed circular viral DNA formed from two types of linear DNA in woodchuck hepatitis virus-infected liver. J. Virol. 1996, 70, 4567–4575. [Google Scholar] [PubMed]
- Bloom, K.; Ely, A.; Arbuthnot, P. A T7 endonuclease I assay to detect Talen-mediated targeted mutation of HBV cccDNA. In Hepatitis B Virus: Methods and Protocols; Methods in Molecular Biology; Guo, H.T., Cuconati, A., Eds.; Springer: New York, NY, USA, 2017; Volume 1540, pp. 85–95. [Google Scholar]
- Wieland, S.F.; Spangenberg, H.C.; Thimme, R.; Purcell, R.H.; Chisari, F.V. Expansion and contraction of the hepatitis B virus transcriptional template in infected chimpanzees. Proc. Natl. Acad. Sci. USA 2004, 101, 2129–2134. [Google Scholar] [CrossRef] [PubMed]
- Lutze, L.H.; Winegar, R.A. A quick and efficient method for the recovery of plasmid or viral DNA from mammalian cells. Nucleic Acids Res. 1990, 18, 6150. [Google Scholar] [CrossRef] [PubMed]
- Belloni, L.; Pollicino, T.; Nicola, F.D.; Guerrieri, F.; Raffa, G.; Faneiulli, M.; Raimondo, G.; Levrero, M. Nuclear HBx binds the HBV minichromosome and modifies the epigenetic regulation of cccDNA function. Proc. Natl. Acad. Sci. USA 2009, 106, 19975–19979. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, W.; Ogura, N.; Watashi, K.; Wakita, T. Host factor PRPF31 is involved in cccDNA production in HBV-replicating cells. Biochem. Biophys. Res. Commun. 2017, 482, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.W.; Nie, H.; Yan, R.; Guo, J.T.; Block, T.M.; Guo, H.T. A Southern blot assay for detection of hepatitis B virus covalently closed circular DNA from cell cultures. In Antiviral Methods and Protocols; Methods in Molecular Biology; Gong, E.Y., Ed.; Humana Press: Totowa, NJ, USA, 2013; Volume 1030, pp. 151–161. [Google Scholar]
- Chong, C.L.; Chen, M.L.; Wu, Y.C.; Tsai, K.N.; Huang, C.C.; Hu, C.P.; Jeng, K.S.; Chou, Y.C.; Chang, C.M. Dynamics of HBV cccDNA expression and transcription in host factor PRPF31 is indifferent cell growth phase. J. Biomed. Sci. 2011, 18, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kock, J.; Theilmann, L.; Galle, P.; Schlicht, H.J. Hepatitis B virus nucleic acids associated with human peripheral blood mononuclear cells do not originate from replicating virus. Hepatology 1996, 23, 405–413. [Google Scholar] [PubMed]
- Thompson, A.J.; Nguyen, T.; Iser, D.; Ayres, A.; Jackson, K.; Littlejoin, M.; Shavin, J.; Bowden, S.; Gane, E.J.; Abbott, W.; et al. Serum hepatitis B surface antigen and hepatitis B e antigen titers: Disease phase influences correlation with viral load and intrahepatic hepatitis B virus markers. Hepatology 2010, 51, 1933–1944. [Google Scholar] [CrossRef] [PubMed]
- Reaiche, G.Y.; Le Mire, M.F.; Mason, W.S.; Jilbert, A.R. The persistence in the liver of residual duck hepatitis B virus covalently closed circular DNA is not dependent upon new viral DNA synthesis. Virology 2010, 406, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Lenci, I.; Marcuccilli, F.; Tisone, G.; Palolo, D.D.; Tariciotti, L.; Ciotti, M.; Guenci, T.; Perno, C.F.; Angelico, M. Total and covalently closed circular DNA detection in liver tissue of long-term survivors transplanted for HBV-related cirrhosis. Dig. Liver Dis. 2010, 42, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Protzer, U.; Seyfried, S.; Quasdorff, M.; Sass, G.; Svorcova, M.; Webb, D.; Bohne, F.; Hosel, M.; Schirmacher, P.; Tiegs, G. Antiviral activity and hepatoprotection by heme oxygenase-1 in hepatitis B virus infection. Gastroenterology 2007, 133, 1156–1165. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.C.; Stadler, D.; Lucifora, J.; Reisinger, F.; Webb, D.; Hosel, M.; Michler, T.; Wisskirchen, K.; Cheng, X.M.; Zhang, K.; et al. Interferon-γ and tumor necrosis factor-α produced by T cells reduce the HBV persistence form, cccDNA, without cytolysis. Gastroenterology 2016, 150, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Mason, A.L.; Xu, L.; Guo, L.; Kuhns, M.; Perrillo, R.P. Molecular basis for persistent hepatitis B virus infection in the liver after clearance of serum hepatitis B surface antigen. Hepatology 1998, 27, 1736–1742. [Google Scholar] [CrossRef] [PubMed]
- Stoll-Becker, S.; Repp, R.; Glebe, D.; Schaefer, S.; Kreuder, J.; Kann, M.; Lampert, F.; Gerlich, W.H. Transcription of hepatitis B virus in peripheral blood mononuclear cells from persistently infected patients. J. Virol. 1997, 71, 5399–5407. [Google Scholar] [PubMed]
- Yuen, M.F.; Wong, D.K.; Sum, S.S.; Yuan, H.J.; Yuen, J.C.; Chan, A.D.; Wong, B.C.; Lai, C.L. Effect of lamivudine therapy on the serum covalently closed-circular (ccc) DNA of chronic hepatitis B infection. Am. J. Gastroenterol. 2005, 100, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.T.; Chiu, H.T.; Chu, C.M.; Liaw, Y.F. G1 phase dependent Nuclear localization of rcDNA and aphindicolin induced cccDNA. J. Med. Virol. 1998, 55, 42–50. [Google Scholar] [CrossRef]
- Cai, D.W.; Mills, C.; Yu, W.Q.; Yan, R.; Aldrich, C.E.; Saputelli, J.R.; Mason, W.S.; Xu, X.D.; Guo, J.T.; Block, T.M.; et al. Identification of disubstituted sulfonamide compounds as specific inhibitors of hepatitis B virus covalently closed circular DNA formation. Antimicrob. Agents Chemother. 2012, 56, 4277–4288. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shen, T.; Huang, X.B.; Kumar, G.R.; Chen, X.M.; Zeng, Z.Z.; Zhang, R.Y.; Chen, R.; Li, T.; Zhang, T.Y.; et al. Serum hepatitis B virus RNA is encapsidated pregenome RNA that may be associated with persistence of viral infection and rebound. J. Hepatol. 2016, 65, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.Y.; Chen, P.; Hou, X.H.; Xu, W.J.; Wang, D.; Wang, T.Y.; Zhang, L.P.; Zhang, G.; Gao, Z.L.; He, C.Y. The recombined cccDNA produced using minicircle technology mimicked HBV genome in structure and function closely. Sci. Rep. 2016, 6, 25552. [Google Scholar] [CrossRef] [PubMed]
- Osborn, J. A review of radioactive and non-radioactive-based techniques used in life science applications. Part I: Blotting techniques. In Innovations Forum Life Science News 6, Amersham Pharmacia Biotech; Amersham: Little Chalfont, UK, 2000; Volume 4. [Google Scholar]
- Neuhaus-Url, G.; Neuhaus, G. The use of the nonradioactive digoxigenin chemiluminescent technology for plant genomic Southern blot hybridization: A comparison with radioactivity. Transgenic Res. 1993, 2, 115–120. [Google Scholar] [CrossRef]
- Allefs, J.J.; Salentijn, E.M.; Krens, F.A.; Rouwendal, G.J. Optimization of non-radioactive Southern blot hybridization: Single copy detection and reuse of blots. Nucleic Acids Res. 1990, 18, 3099–3100. [Google Scholar] [CrossRef] [PubMed]
- Miotke, L.; Lau, B.T.; Rumma, R.T.; Ji, H.P. High sensitivity detection and quantitation of DNA copy number and single nucleotide variants with single color droplet digital PCR. Anal. Chem. 2014, 86, 2618–2624. [Google Scholar] [CrossRef] [PubMed]
- Morisset, D.; Stebih, D.; Milavec, M.; Gruden, K.; Zel, J. Quantitative analysis of food and feed samples with droplet digital PCR. PLoS ONE 2013, 8, e62583. [Google Scholar] [CrossRef] [PubMed]
- Whale, A.S.; Huggett, J.F.; Cowen, S.; Speris, V.; Shaw, J.; Ellison, S.; Foy, C.A.; Scott, D.J. Comparison of microfluidic digital PCR and conventional quantitative PCR for measuring copy number variation. Nucleic Acids Res. 2012, 40, e82. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.; Dube, S.; Mir, A.; Qin, J.; Sun, G.; Ramakrishnan, R.; Jones, R.C.; Livak, K.J. Taking qPCR to a higher level: Analysis of CNV reveals the power of high throughput qPCR to enhance quantitative resolution. Methods 2010, 50, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Jones, R.C.; Ramakrishnan, R. Studying copy number variations using a nanofluidic platform. Nucleic Acids Res. 2008, 36, e116. [Google Scholar] [CrossRef] [PubMed]
- Margeridon, S.; Carrouee-Durantel, S.; Chemin, I.; Barround, L.; Zoulim, F.; Trepo, C.; Kay, A. Rolling circle amplification, a powerful tool for genetic and functional studies of complete hepatitis B virus genomes from low-level infections and for directly probing covalently closed circular DNA. Antimicrob. Agents Chemother. 2008, 52, 3068–3073. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.M.; Shi, L.; Quina, F.H.; Rosenzweig, Z. Stöber synthesis of monodispersed luminescent silica nanoparticles for bioanalytical assays. Langmuir 2005, 21, 4277–4280. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.K.; Duan, L.; Guo, Z.R.; Mu, Y.D.; Ma, M.; Xu, L.; Zhang, Y.; Gu, N. An improved way to prepare superparamagnetic magnetite-silica core-shell nanoparticles for possible biological application. J. Magn. Magn. Mater. 2005, 285, 65–70. [Google Scholar] [CrossRef]
- Schiestel, T.; Brunner, H.; Tovar, G.E. Controlled surface functionalization of silica nanospheres by covalent conjugation reactions and preparation of high density streptavidin nanoparticles. J. Nanosci. Nanotechnol. 2004, 4, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, R.W.; Lyamichev, V.; Arruda, M.D.; Neri, B. Clinical, genetic, and pharmacogenetic applications of Invader assay. Mol. Diagn. 1999, 4, 353–364. [Google Scholar] [CrossRef]
- Arruda, M.D.; Lyamichev, V.I.; Eis, P.S.; Iszczyszyn, W.; Kwiatkowski, R.W.; Olson, S.M.; Rasmussen, E.B. Invader technology for DNA and RNA analysis: Principles and applications. Expert Rev. Mol. Diagn. 2002, 2, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.B.; Zhu, X.; Yan, L.B.; Du, L.Y.; Liu, C.; Liao, J.; Tang, H. Quantitative intrahepatic HBV cccDNA correlates with histological liver inflammation in chronic hepatitis B virus infection. Int. J. Infect. Dis. 2016, 52, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Meyerhans, A.; Vartanian, J.P.; Wain-Hobson, S. DNA recombination during PCR. Nucleic Acids Res. 1990, 18, 1687–1691. [Google Scholar] [CrossRef] [PubMed]
- Suspene, R.; Thiers, V.; Vartanian, J.P.; Wain-Hobson, S. PCR mediated recombination impacts the analysis of hepatitis B Virus covalently closed circular DNA. Retrovirology 2016, 13, 84. [Google Scholar] [CrossRef] [PubMed]
- Singla, B.; Chakraborti, A.; Sharma, B.K.; Kapil, S.; Chawla, Y.K.; Arora, S.K.; Das, A.; Dhiman, R.K.; Duseja, A. Levels of hepatitis B virus replicative intermediate in serum samples of chronic hepatitis B patients. Mol. Biol. Rep. 2014, 41, 4689–4696. [Google Scholar] [CrossRef] [PubMed]
- Pasquinelli, C.; Melegari, M.; Villa, E.; Scaglioni, P.P.; Seidenari, M.; Mongiardo, N.; Rienzo, B.D.; Manenti, F. Hepatitis B virus infection of peripheral blood mononuclear cells is common in acute and chronic hepatitis. J. Med. Virol. 1990, 31, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Torii, N. Configuration and replication competence of hepatitis B virus DNA in peripheral blood mononuclear cells from chronic hepatitis B patients and patients who have recovered from acute self-limited hepatitis. Hepatol. Res. 2003, 25, 234–243. [Google Scholar] [CrossRef]
- Manicassamy, B.; Manicassamy, S.; Belicha-Villanueva, A.; Pisanelli, G.; Pulendran, B.; Garcia-Sastre, A. Analysis of in vivo dynamics of influenza virus infection in mice using a GFP reporter virus. Proc. Natl. Acad. Sci. USA 2010, 107, 11531–11536. [Google Scholar] [CrossRef] [PubMed]
- Nogales, A.; Rodriguez-Sanchez, I.; Monte, K.; Lenschow, D.J.; Perez, D.R.; Martinez-Sobrido, L. Replication-competent fluorescent-expressing influenza B virus. Virus Res. 2016, 213, 69–81. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methods | Advantages | Disadvantages | Dynamic Range 1 | Limit of Detection 1 | References |
|---|---|---|---|---|---|
| Southern blot | Reliable; reproducible. | Complicated; costly; time-consuming; insensitive; safety concerns. | More than 10 fg DNA | 10 fg DNA (~2 × 106 copies) | [47,48,49,55,56,57,59,65,68,71,72,76,78,81,82,83] |
| Conventional quantitative polymerase chain reaction (qPCR) | Simple; rapid; accurate; economical; sensitive; high-throughput. | Specificity is not absolute when rcDNA is massive. | 2 × 103–2 × 1012 copies/mL | 2.7 × 102 IU/mL (2 × 103 copies/mL) | [11,12,13,26,32,33,34,38,39,41,42,43,44,46,47,48,54,63,68,69,84,85,86,87,88,89] |
| Competitive qPCR | More sensitive than Southern blot; rcDNA and cccDNA can be readily distinguished. | Specificity is not absolute when rcDNA is massive; Southern blot with 32P-labeled probe detection is indispensable. | 2.5–60 ng DNA | 2 × 104 copies | [45] |
| Semi-nested and nested qPCR | Sensitive; specific. | May be contaminated; more risks to amplify unspecific signal. | 3.0 × 102–3.9 × 108 copies/mL | 3.0 × 102 copies/mL | [31,46,90,91] |
| droplet-digital PCR | Super-sensitive; accurate. | Upper detection limit is restricted. | 1–106 copies | 1 copy | [20] |
| Rolling circle amplification qPCR | Practical; sensitive; specific. | Time-consuming; cross-linked proteins could hinder effective amplification. | 102–1010 copies/mL | 102 copies/mL | [30] |
| Rolling circle amplification–in situ qPCR | Sensitive; visible at single-cell resolution. | Diffusion of amplified DNA to neighboring cells; cross-linked proteins could hinder effective amplification. | More than 2 copies/cell | 2 copies/cell | [28] |
| Magnetic capture hybridization qPCR | Enrichment of cccDNA; sensitive; reproducible; specific. | Cannot capture all cccDNA; complicated; costly. | 102–106 IU/mL | 90 IU/mL | [27] |
| Invader assay | Specific; simple; reproducible. | Minimal interference from double-stranded and integrated HBV DNA. | 104–109 copies/mL | 50 copies (104 copies/mL) | [35,37,40,92] |
| In situ hybridization | Specific; visible at single-cell resolution; can distinguish and locate different DNA, RNA and proteins; without diffusion of amplified products. | Complicated probe design. | More than 1 copy | 1 copy under optimal conditions | [21,93] |
| Substituted markers 2 (HBsAg/HBeAg/HBcrAg/pgRNA/GLuc) | Non-invasive; convenient; cost-effective; high-throughput. | Indirect and may not reflect all information about cccDNA. | - | - | [12,22,23,24,25,29,36,63,94,95,96] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhao, J.; Yuan, Q.; Xia, N. Detection of HBV Covalently Closed Circular DNA. Viruses 2017, 9, 139. https://doi.org/10.3390/v9060139
Li X, Zhao J, Yuan Q, Xia N. Detection of HBV Covalently Closed Circular DNA. Viruses. 2017; 9(6):139. https://doi.org/10.3390/v9060139
Chicago/Turabian StyleLi, Xiaoling, Jinghua Zhao, Quan Yuan, and Ningshao Xia. 2017. "Detection of HBV Covalently Closed Circular DNA" Viruses 9, no. 6: 139. https://doi.org/10.3390/v9060139
APA StyleLi, X., Zhao, J., Yuan, Q., & Xia, N. (2017). Detection of HBV Covalently Closed Circular DNA. Viruses, 9(6), 139. https://doi.org/10.3390/v9060139