
Clinical and Laboratory Parameters of Carp Edema Virus Disease: A Case Report
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case description and Fish Sampling
2.2. Blood Sampling
2.3. Pathological and Parasitological Examination; Sampling for Histopathology
2.4. Virus Isolation, PCR and Sequence Analysis
2.5. Haematological and Immunological Parameters
2.6. Statistical Analysis
3. Results
3.1. Virus Isolation, PCR Analysis and Sequencing
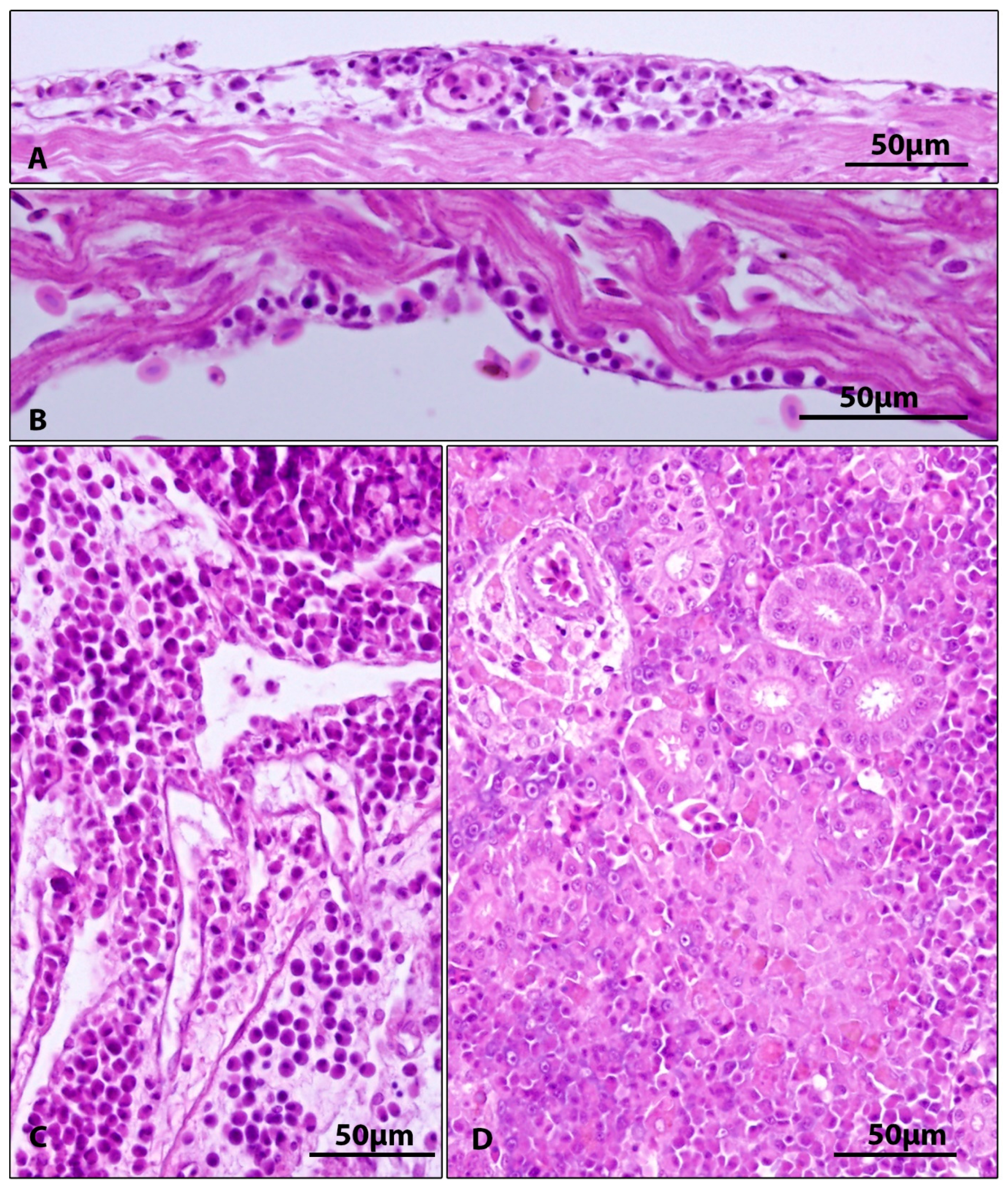
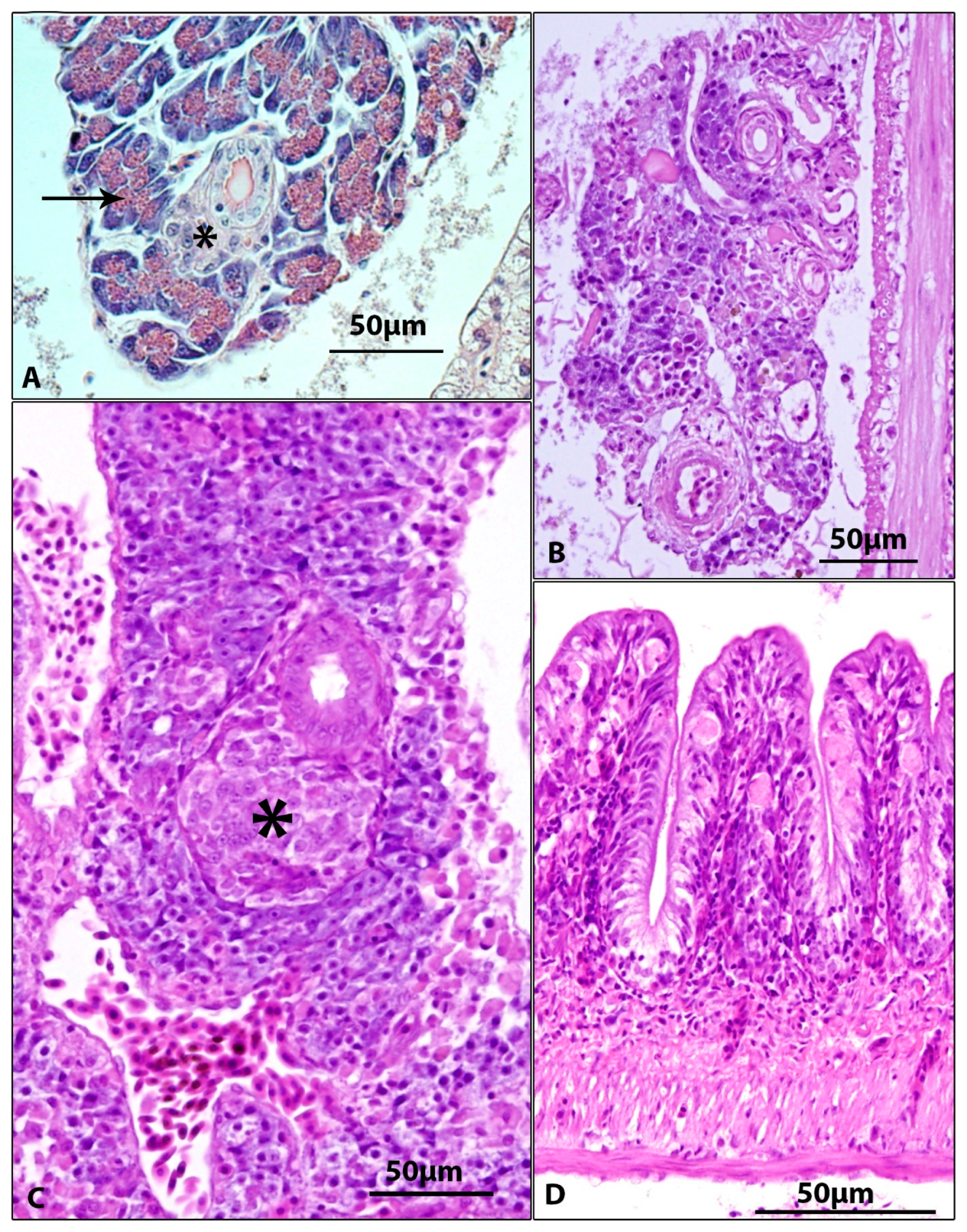
3.2. Gross and Microscopic Pathology, Results of Parasitological and Bacteriological Examination
3.3. Leukocyte Counts, Differential Cell Counts, and Immunological Parameters
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ono, S.I.; Nagai, A.; Sugai, N. A histopathological study on juvenile colorcarp Cyprinus carpio, showing edema. Fish Pathol. 1986, 21, 167–175. [Google Scholar] [CrossRef]
- Amita, K.; Oe, M.; Matoyama, H.; Yamaguchi, N.; Fukuda, H. A survey of koi herpesvirus and carp edema virus in color carp cultured in Niigata Prefecture, Japan. Fish Pathol. 2002, 37, 197–198. [Google Scholar] [CrossRef]
- Lovy, J.; Friend, S.E.; Al-Hussinee, L.; Waltzek, T.B. First report of carp edema virus in the mortality of wild common carp Cyprinus carpio in North America. Dis. Aquat. Org. 2018, 131, 177–186. [Google Scholar] [CrossRef]
- Viadanna, P.; Pilarski, F.; Hesami, S.; Waltzek, T. First report of carp edema virus (CEV) in South American koi. In Proceedings of the 40th East Fish Health Workshop, Charleston, SC, USA, 2–6 March 2015; Volume 12. [Google Scholar]
- Swaminathan, T.R.; Kumar, R.; Dharmaratnam, A.; Basheer, V.S.; Sood, N.; Pradhan, P.K.; Sanil, N.K.; Vijayagopal, P.; Jena, J.K. Emergence of carp edema virus in cultured ornamental koi carp, Cyprinus carpio koi, in India. J. Gen. Virol. 2016, 97, 3392–3399. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, P.; Yang, R.; Chen, J.; Wang, K.; Geng, Y.; Lai, W.; Huang, X.; Chen, D.; Fang, J.; Chen, Z.; et al. First detection of carp edema virus in association with cyprinid herpesvirus 3 in cultured ornamental koi, Cyprinus carpio L., in China. Aquaculture 2018, 490, 162–168. [Google Scholar] [CrossRef]
- Kim, S.W.; Jun, J.W.; Giri, S.S.; Chi, C.; Yun, S.; Kim, H.J.; Kim, S.G.; Kang, J.W.; Park, S.C. First report of carp oedema virus infection of koi (Cyprinus carpio haematopterus) in the Republic of Korea. Transbound. Emerg. Dis. 2018, 65, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Way, K.; Stone, D. Emergence of carp edema virus-like (CEV-like) disease in the UK. CEFAS Finfish News 2013, 15, 32–34. [Google Scholar]
- Haenen, O.; Way, K.; Stone, D.; Engelsma, M. ‘Koi Sleepy Disease’ found for the first time in Koi Carps in the Netherlands. Tijdschr. Voor Diergeneeskd. 2014, 139, 26. [Google Scholar]
- Veselý, T.; Pokorová, D.; Reschová, S.; Piačková, V. Detection of carp edema virus in common carp (Cyprinus carpio) and koi in the Czech Republic. In Proceedings of the 17th International Conference on Diseases of Fish and Shellfish, Las Palmas, Spain, 7–11 September 2015; Abstract and poster; Volume 346. [Google Scholar]
- Lewisch, E.; Gorgoglione, B.; Way, K.; El-Matbouli, M. Carp edema virus/Koi sleepy disease: An emerging disease in Central-East Europe. Transbound. Emerg. Dis. 2015, 62, 6–12. [Google Scholar] [CrossRef]
- Pretto, T.; Abbadi, M.; Panzarin, V.; Quartesan, R.; Manfrin, A.; Toffan, A. Carp edema virus (CEV): First detection in Italy. In Proceedings of the 17th International Conference on Diseases of Fish and Shellfish, Las Palmas, Spain, 7–11 September 2015; Abstract and poster; Volume 343. [Google Scholar]
- Jung-Schroers, V.; Adamek, M.; Teitge, F.; Hellmann, J.; Bergmann, S.M.; Schütze, H.; Kleingeld, D.W.; Way, K.; Stone, D.; Runge, M.; et al. Another potential carp killer? Carp Edema Virus disease in Germany. BMC Vet. Res. 2015, 11, 114. [Google Scholar] [CrossRef] [PubMed]
- Bigarré, L.; Baud, M.; Pallandre, L.; Meunier, E.; Leguay, E. Maladie du sommeil de la carpe: État des lieux des con-naissances et situation épidémiologique en France. Bull. Épidémiologique St. Anim. Aliment. 2016, 76, 12–13. [Google Scholar]
- Bachmann, J.; Keilholz, S. Frühjahrsverluste in der fränkischen Karp-fenteichwirtschaft—Viraler Erreger als Ursache? Erstnachweise der KSD/CEV in Bayern. Fisch Teichwirt 2016, 3, 91–92. [Google Scholar]
- Matějíčková, K.; Pojezdal, L.; Pokorová, D.; Reschová, S.; Piačková, V.; Palíková, M.; Veselý, T.; Papežíková, I. Carp oedema virus disease outbreaks in Czech and Slovak aquaculture. J. Fish Dis. 2020, 43, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Oyamatsu, T.; Matoyama, H.; Yamamoto, K.Y.; Fukuda, H. A trial for the detection of carp edema virus by using polymerase chain reaction. Aquac. Sci. 1997, 45, 247–251. [Google Scholar]
- Miyazaki, T.; Isshiki, T.; Katsuyuki, H. Histopathological and electron microscopy studies on sleepy disease of koi Cyprinus carpio koi in Japan. Dis. Aquat. Org. 2005, 65, 197–207. [Google Scholar] [CrossRef]
- Zhang, X.; Ni, Y.; Ye, J.; Xu, H.; Hou, Y.; Luo, W.; Shen, W. Carp edema virus, an emerging threat to the carp (Cyprinus carpio) industry in China. Aquaculture 2017, 474, 34–39. [Google Scholar] [CrossRef]
- Way, K.; Haenen, O.; Stone, D.; Adamek, M.; Bergmann, S.M.; Bigarré, L.; Diserens, N.; El-Matbouli, M.; Gjessing, M.; Jung-Schroers, V.; et al. Emergence of carp edema virus (CEV) and its significance to European common carp and koi Cyprinus carpio. Dis. Aquat. Org. 2017, 126, 155–166. [Google Scholar] [CrossRef]
- Svobodová, Z.; Pravda, D.; Paláčková, J. Unified Methods of Haematological Examination of Fish; Research Institute of Fish Culture and Hydrobiology: Vodnany, Czech Republic, 1991; pp. 11–31. [Google Scholar]
- Kubala, L.; Lojek, A.; Číž, M.; Vondráček, J.; Dušková, M.; Slavíková, H. Determination of phagocyte activity in whole blood of carp (Cyprinus carpio) by luminol-enhanced chemiluminescence. Vet. Med. 1996, 41, 323–327. [Google Scholar]
- Poisot, T.; Šimková, A.; Hyršl, P.; Morand, S. Interactions between immunocompetence, somatic condition and parasitismin the chub Leuciscus cephalus in early spring. J. Fish Biol. 2009, 75, 1667–1682. [Google Scholar] [CrossRef]
- McEwan, A.D.; Fisher, E.W.; Selman, I.E. Observations on immune globulin levels of neonatal calves and their relationship to disease. J. Comp. Pathol. 1970, 80, 259–265. [Google Scholar] [CrossRef]
- Atosuo, J.; Lehtinen, J.; Vojtek, L.; Lilius, E.M. Escherichia coli K-12 (pEGFPluxABCDEamp): A tool for analysis of bacterial killing by antibacterial agents and human complement activities on a real-time basis. Luminescence 2013, 28, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Buchtíková, S.; Šimková, A.; Rohlenová, K.; Flajšhans, M.; Lojek, A.; Lilius, E.M.; Hyršl, P. The seasonal changes in innate immunity of the common carp (Cyprinus carpio). Aquaculture 2011, 318, 169–175. [Google Scholar] [CrossRef]
- Adamek, M.; Teitge, F.; Baumann, I.; Jung-Schroers, V.; Abd El Rahman, S.; Paley, R.; Piačková, V.; Gela, D.; Kocour, M.; Rakers, S.; et al. Koi sleepy disease as a pathophysiological and immunological consequence of a branchial infection of common carp with carp edema virus. Virulence 2021, 12, 1855–1883. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, Z.; Prajapati, M.; Li, Y. Lymphopenia Caused by Virus Infections and the Mechanisms Beyond. Viruses 2021, 13, 1876. [Google Scholar] [CrossRef] [PubMed]
- Warny, M.; Helby, J.; Nordestgaard, B.G.; Birgens, H.; Bojesen, S.E. Lymphopenia and risk of infection-related death in 98,344 individuals from a prospective Danish population-based study. PLoS Med. 2018, 15, e1002685. [Google Scholar] [CrossRef]
- Davis, A.K.; Maney, D.L.; Maerz, J.C. The use of leukocyte profiles to measure stress in vertebrates: A review for ecologists. Funct. Ecol. 2008, 22, 760–772. [Google Scholar] [CrossRef]
- Witeska, M. Stress in fish-hematological and immunological effects of heavy metals. Electron. J. Ichthyol. 2005, 1, 35–41. [Google Scholar]
- Haugland, G.T.; Jakobsen, R.A.; Vestvik, N.; Ulven, K.; Stokka, L.; Wergeland, H.I. Phagocytosis and Respiratory Burst Activity in Lumpsucker (Cyclopterus lumpus L.) Leucocytes Analysed by Flow Cytometry. PLoS ONE 2012, 7, e47909. [Google Scholar] [CrossRef]
- Hurtado-Nedelec, M.; Makni-Maalej, K.; Gougerot-Pocidalo, M.A.; My-Chan Dang, P.; Jamel El-Benna, J. Assessment of priming of the human neutrophil respiratory burst. Methods Mol. Biol. 2014, 1124, 405–412. [Google Scholar]
- Avtalion, R.R. Temperature effect on antibody production and immunological memory, in carp (Cyprinus carpio) immunized against bovine serum albumin (BSA). Immunology 1969, 17, 927–931. [Google Scholar] [PubMed]
- Le Morvan, C.; Deschaux, P.; Troutaud, D. Effects and mechanisms of environmental temperature on carp (Cyprinus carpio) anti-DNP antibody response and non-specific cytotoxic cell activity: A kinetic study. Dev. Comp. Immunol. 1996, 20, 331–340. [Google Scholar] [CrossRef]
- Adamek, M.; Oschilewski, A.; Wohlsein, P.; Jung-Schroers, V.; Teitge, F.; Dawson, A.; Gela, D.; Piačková, V.; Kocour, M.; Adamek, J.; et al. Experimental infections of different carp strains with the carp edema virus (CEV) give insights into the infection biology of the virus and indicate possible solutions to problems caused by koi sleepy disease (KSD) in carp aquaculture. Vet. Res. 2017, 48, 12. [Google Scholar] [CrossRef] [PubMed]
- Zrnčić, S.; Oraić, D.; Zupičić, I.G.; Pavlinec, Ž.; Brnić, D.; Rogić, Z.A.; Sučec, I.; Steinhagen, D.; Adamek, M. Koi herpesvirus and carp edema virus threaten common carp aquaculture in Croatia. J. Fish Dis. 2020, 43, 1–13. [Google Scholar] [CrossRef]
- Luo, F.; Lian, Z.; Niu, Y.; Lű, A.; Hu, X.; Xie, X.; Sun, J. Molecular characterization of carp edema virus disease: An emerging threat to koi Cyprinus carpio in China. Microb. Pathog. 2020, 149, 104551. [Google Scholar] [CrossRef]
- Marsella, A.; Pretto, T.; Abbadi, M.; Quartesan, R.; Cortinovis, L.; Fiocchi, E.; Manfrin, A.; Toffan, A. Carp edema virus-related mortality in wild adult common carp (Cyprinus carpio) in Italy. J. Fish Dis. 2021, 44, 939–947. [Google Scholar] [CrossRef]
- Casanova, J.L.; Abel, L. Mechanisms of viral inflammation and disease in humans. Science 2021, 374, 1080–1086. [Google Scholar] [CrossRef] [PubMed]
- Magnadóttir, B. Immunological control of fish diseases. Mar. Biotechnol. 2010, 12, 361–379. [Google Scholar] [CrossRef]
- Adamek, M.; Jung-Schroers, V.; Hellmann, J.; Teitge, F. Concentration of carp edema virus (CEV) DNA in koi tissued affected by koi sleepy disease (KSD). Dis. Aquat. Org. 2016, 119, 245–251. [Google Scholar] [CrossRef]
- Pikula, J.; Pojezdal, L.; Papežíková, I.; Minářová, H.; Mikulíková, I.; Banďouchová, H.; Blahová, J.; Bednarska, M.; Mareš, J.; Palíková, M. Carp Edema Virus Infection is Associated with Severe Metabolic Disturbance in Fish. Front. Vet. Sci. 2021, 8, 679970. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Date | Water Temperature | Fish | Pathoanatomical Examination | Parasitological Examination | Bacteriological Examination |
|---|---|---|---|---|---|
| 4 April 2017 (Sampling A) | 12 °C (Tank 1) | Koi carp, diseased (n = 2) | Poor condition (2/2) Skin lesions with loss of scales (2/2) Enophthalmia (2/2) | No parasites | Aeromonas bestiarum (1/2), Aeromonas hydrophila (1/2) |
| 17 April 2017 (Sampling B) | 13 °C (Tank 1) | Koi carp, diseased, CEV+ (n = 5) | Enophthalmia, poor condition (3/5) | No parasites | n. d. |
| Koi carp diseased, CEV− (n = 5) | Haemorrhagic fluid in body cavity (1/5) Bristled scales, poor condition (1/5) | Dactylogyrus sp., 1× (1/5) | |||
| 13 °C (Tank 2) | Koi carp, control (n = 10) | No gross pathology | No parasites |
| Sampling Date | Fish | Sample Type | SVCV (Cell Culture) | SVCV (Nested PCR) | CyHV-3 (Nested PCR) | CEV (Nested PCR) 1st Round/ 2nd Round |
|---|---|---|---|---|---|---|
| 4 April 2017 (Sampling A) | Koi carp, diseased | k, l, s (pooled from 2 fish) | − | − | − | −/− |
| gills (pooled from 2 fish) | − | − | − | +/+ | ||
| 17 April 2017 (Sampling B) | Koi carp, diseased | k, l, s (pooled from each fish), n = 10 | − | − | − | −/− |
| gills, n = 10 | − | − | − | +/+ (5 fish of 10) | ||
| Koi carp, control | gills (pooled from 10 fish) | − | − | − | −/− |
| Koi Carp, Diseased, CEV+ (n = 5) | Koi Carp Diseased, CEV− (n = 5) | Figure | |
|---|---|---|---|
| Gills | Epithelial hyperplasia, fusion of secondary lamellae, cystoid formations between gill lamellae (5/5) | Epithelial hyperplasia, fusion of secondary lamellae, cystoid formations between gill lamellae (5/5) | Figure 2 |
| Pancreas | Absence of pancreatic tissue (2/5) atrophy of pancreas with loss of zymogen granules in exocrine pancreas (3/5) | Absence of pancreatic tissue (1/5) atrophy of pancreas with loss of zymogen granules in exocrine pancreas (4/5) | Figure 4B,C |
| Intestine | Subepithelial infiltrate and presence of spherical formations (probably of macrophage origin) (5/5) | Subepithelial infiltrate and presence of spherical formations (probably of macrophage origin) (3/5) | Figure 4D |
| Heart | Subepicardial and subendocardial infiltrates (2/5) | Subepicardial and subendocardial infiltrates (2/5) | Figure 3A,B |
| Kidney | Proliferation of kidney interstitium, with macrophage aggregation (5/5) | Proliferation of kidney interstitium, with macrophage aggregation (3/5) | Figure 3D |
| Body cavity | Inflammatory infiltration of connective tissue among internal organs (1/5) | Not detected | Figure 3C |
| Control | Diseased CEV+ | Diseased CEV− | |
|---|---|---|---|
| Leukocytes (109·L−1) | 44.0 ± 33.19 a | 42.8 ± 41.88 ab | 19.2 ± 11.58 b |
| Lymphocytes (%) | 91.8 ± 9.67 a | 56.0 ± 37.86 b | 77.6 ± 6.02 b |
| Neutrophils (%) | 7.1 ± 8.96 | 43.6 ± 37.25 | 19.2 ± 5.071 |
| Monocytes (%) | 1.1 ± 1.29 a | 0.40 ± 0.89 b | 3.20 ± 2.59 ab |
| Phagocytes (%) | 8.2 ± 9.67 | 44.0 ± 37.86 | 22.4 ± 6.02 |
| Lymphocytes (109·L−1) | 39.85 ± 29.72 a | 12.79 ± 29 b | 14.97 ± 9.51 b |
| Neutrophils (109·L−1) | 3.78 ± 7.6 | 29.56 ± 42.25 | 3.7 ± 2.24 |
| Monocytes (109·L−1) | 0.37 ± 0.36 a | 0.45 ±1.01 b | 0.53 ± 0.32 ab |
| Phagocytes (109·L−1) | 4.15 ± 7.86 | 30.0 ± 43.2 | 4.23 ± 2.41 |
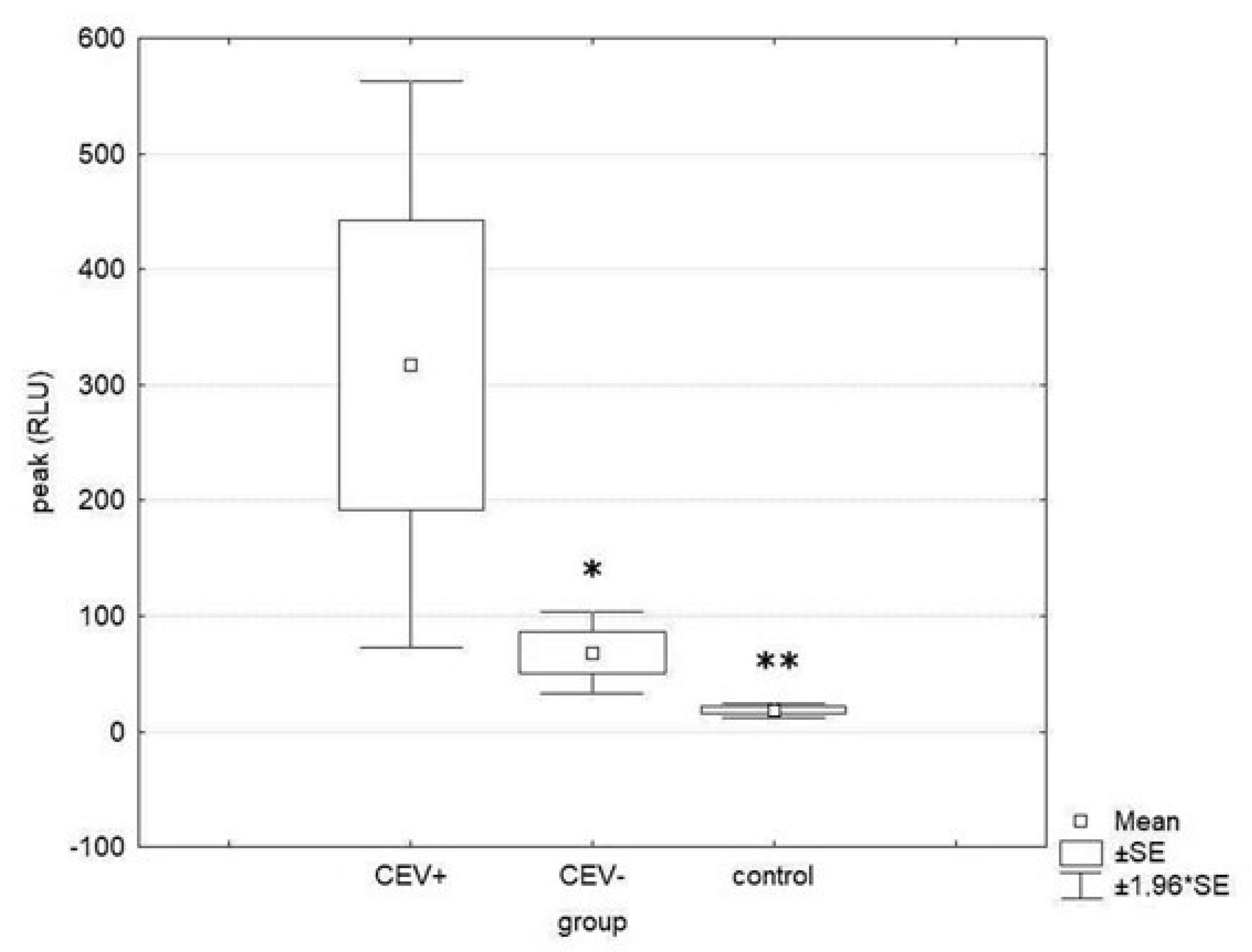
| Phagocytic activity (peak) | 18.3 ± 9.5 a | 317.5 ± 279.7 b | 67.8 ± 40.4 a |
| Phagocytic activity (peak/1000 phagocytes) | 1.59 ± 1.72 | 1.73 ± 1.48 | 1.05 ± 0.90 |
| Lysozyme (mg·mL−1) | 11.02 ± 4.23 | 14.62 ± 4.93 | 11.76 ± 5.75 |
| Complement activity (min) | 46.8 ±27.5 | 49.4 ± 23.7 | 34.3 ± 24.3 |
| Immunoglobulins (mg·mL−1) | 12.3 ± 3.47 | 9.01 ± 2.40 | 11.07 ± 7.74 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papežíková, I.; Piačková, V.; Dyková, I.; Baloch, A.A.; Kroupová, H.K.; Zusková, E.; Pojezdal, Ľ.; Minářová, H.; Syrová, E.; Banďouchová, H.; et al. Clinical and Laboratory Parameters of Carp Edema Virus Disease: A Case Report. Viruses 2023, 15, 1044. https://doi.org/10.3390/v15051044
Papežíková I, Piačková V, Dyková I, Baloch AA, Kroupová HK, Zusková E, Pojezdal Ľ, Minářová H, Syrová E, Banďouchová H, et al. Clinical and Laboratory Parameters of Carp Edema Virus Disease: A Case Report. Viruses. 2023; 15(5):1044. https://doi.org/10.3390/v15051044
Chicago/Turabian StylePapežíková, Ivana, Veronika Piačková, Iva Dyková, Ali Asghar Baloch, Hana Kocour Kroupová, Eliška Zusková, Ľubomír Pojezdal, Hana Minářová, Eva Syrová, Hana Banďouchová, and et al. 2023. "Clinical and Laboratory Parameters of Carp Edema Virus Disease: A Case Report" Viruses 15, no. 5: 1044. https://doi.org/10.3390/v15051044