
Structural Insights into Alphavirus Assembly Revealed by the Cryo-EM Structure of Getah Virus

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Production and Purification of GETV Virions
2.2. Cryo-EM Sample Preparation and Data Collection
2.3. Single Particle Reconstruction
2.4. Model Building and Refinement
2.5. Prediction of Post-Translational Modifications
2.6. Sequence Alignment
3. Results
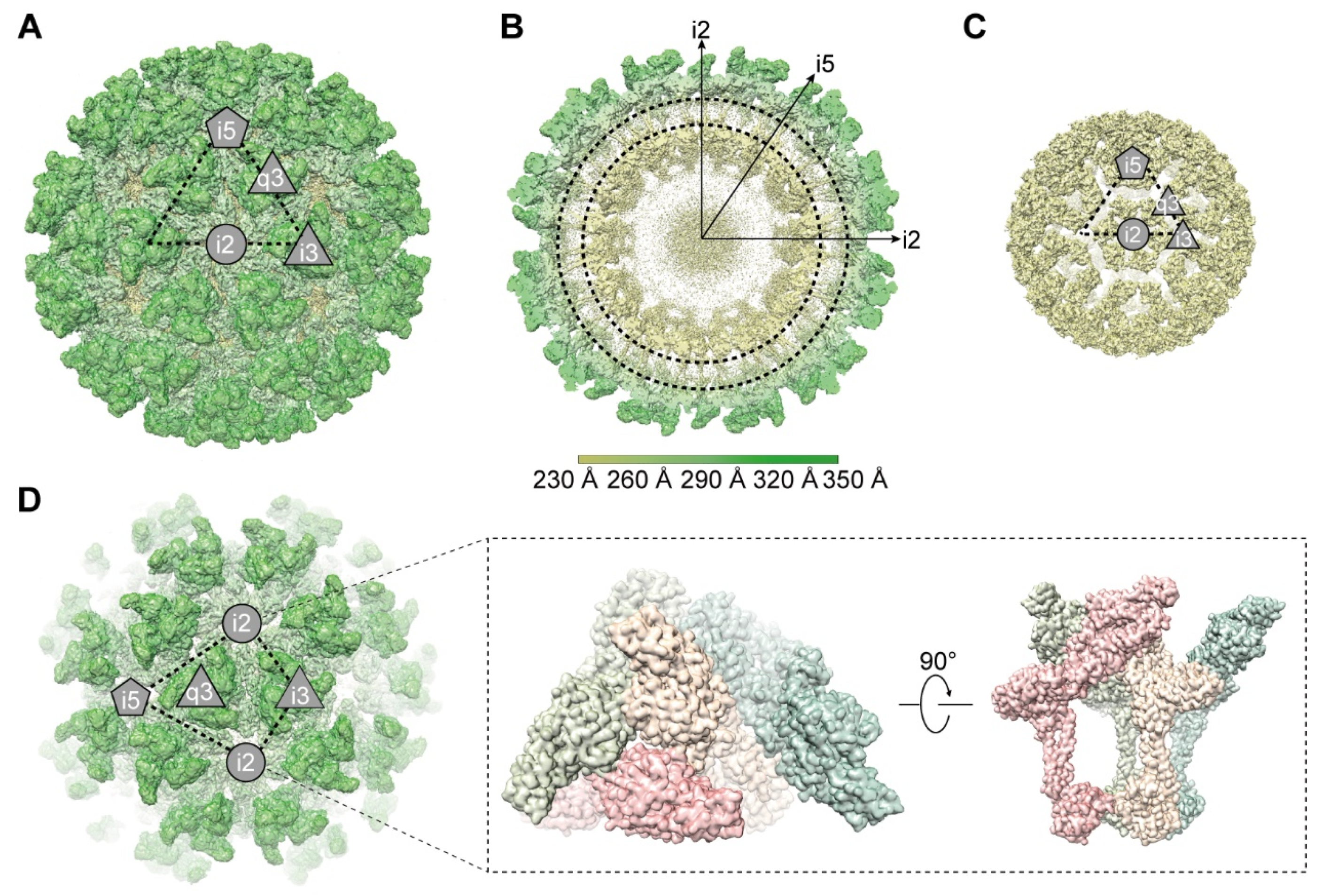
3.1. Overall Cryo-EM Structure of GETV Virions
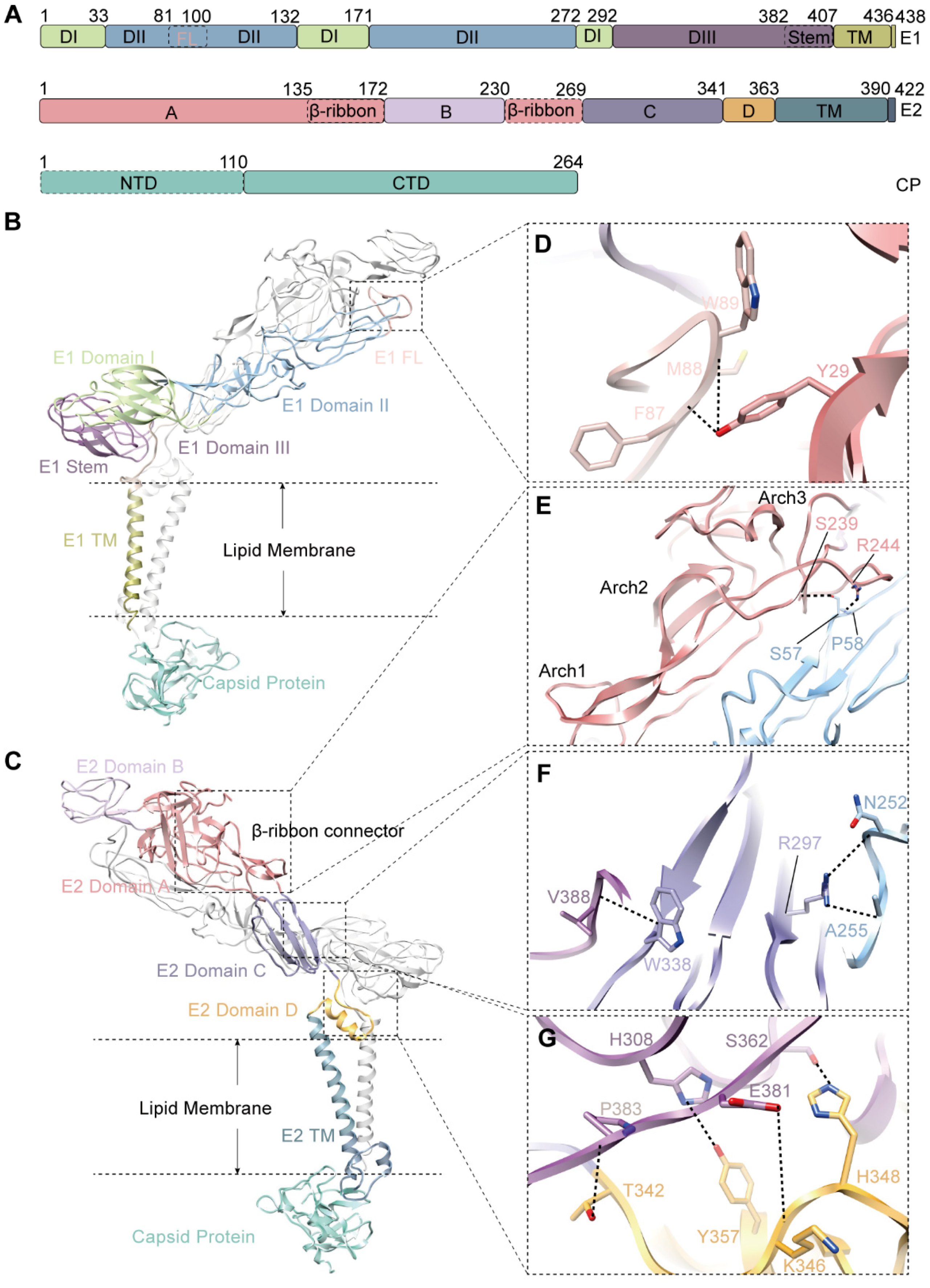
3.2. The Structure of Envelope Glycoproteins
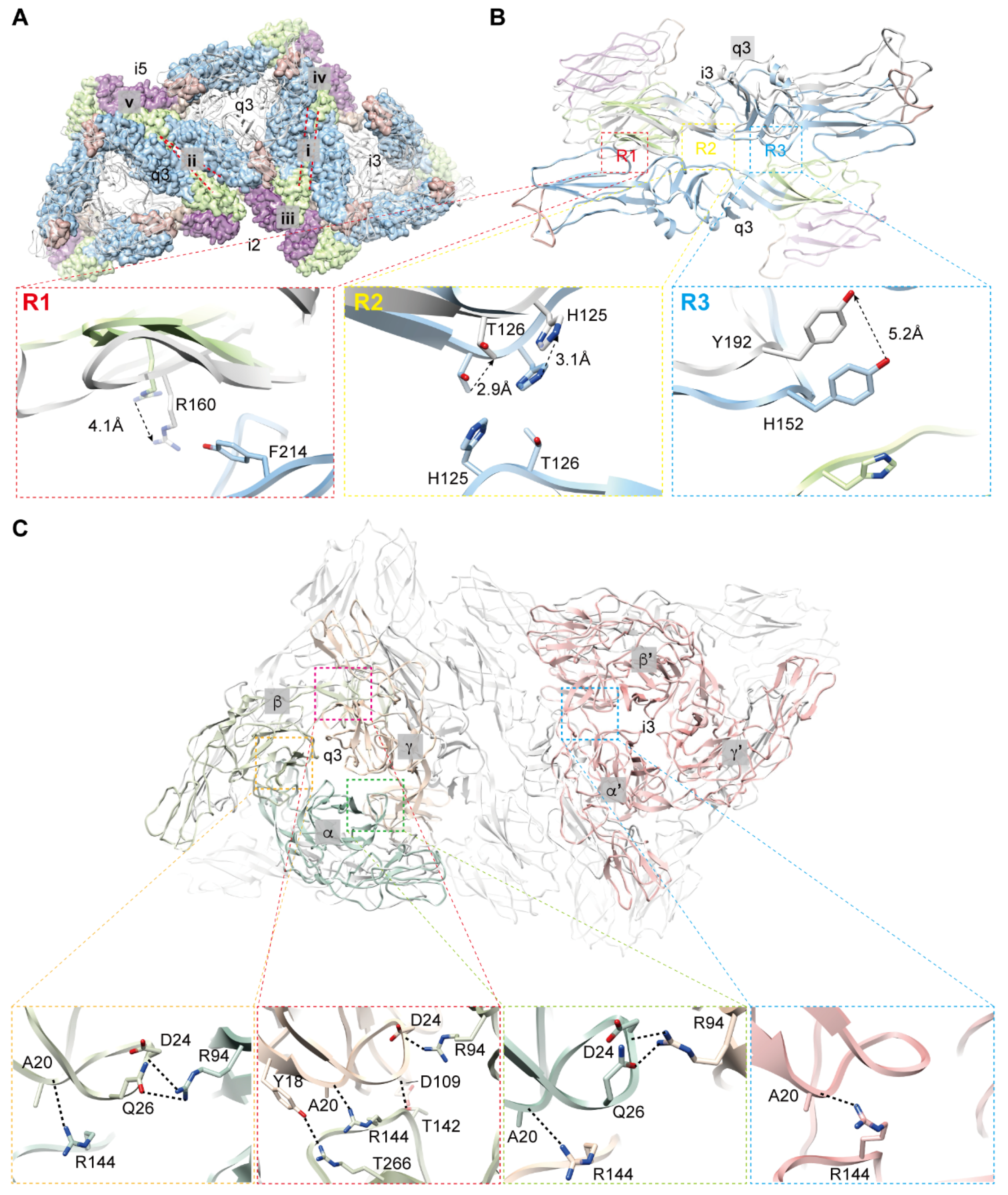
3.3. Interactions among Envelope Glycoproteins in Viral Assembly
3.4. Structure and Assembly of Capsid Protein
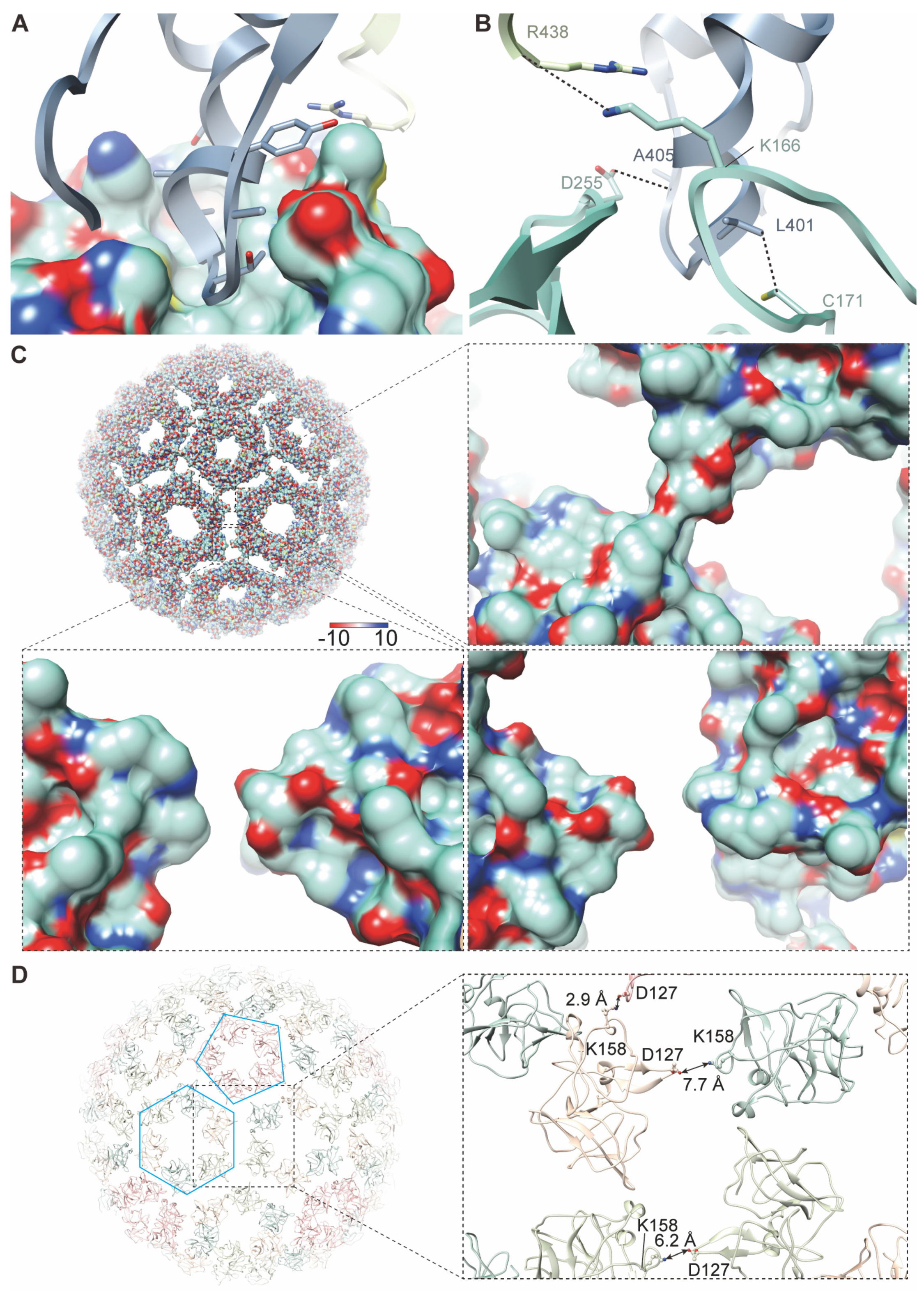
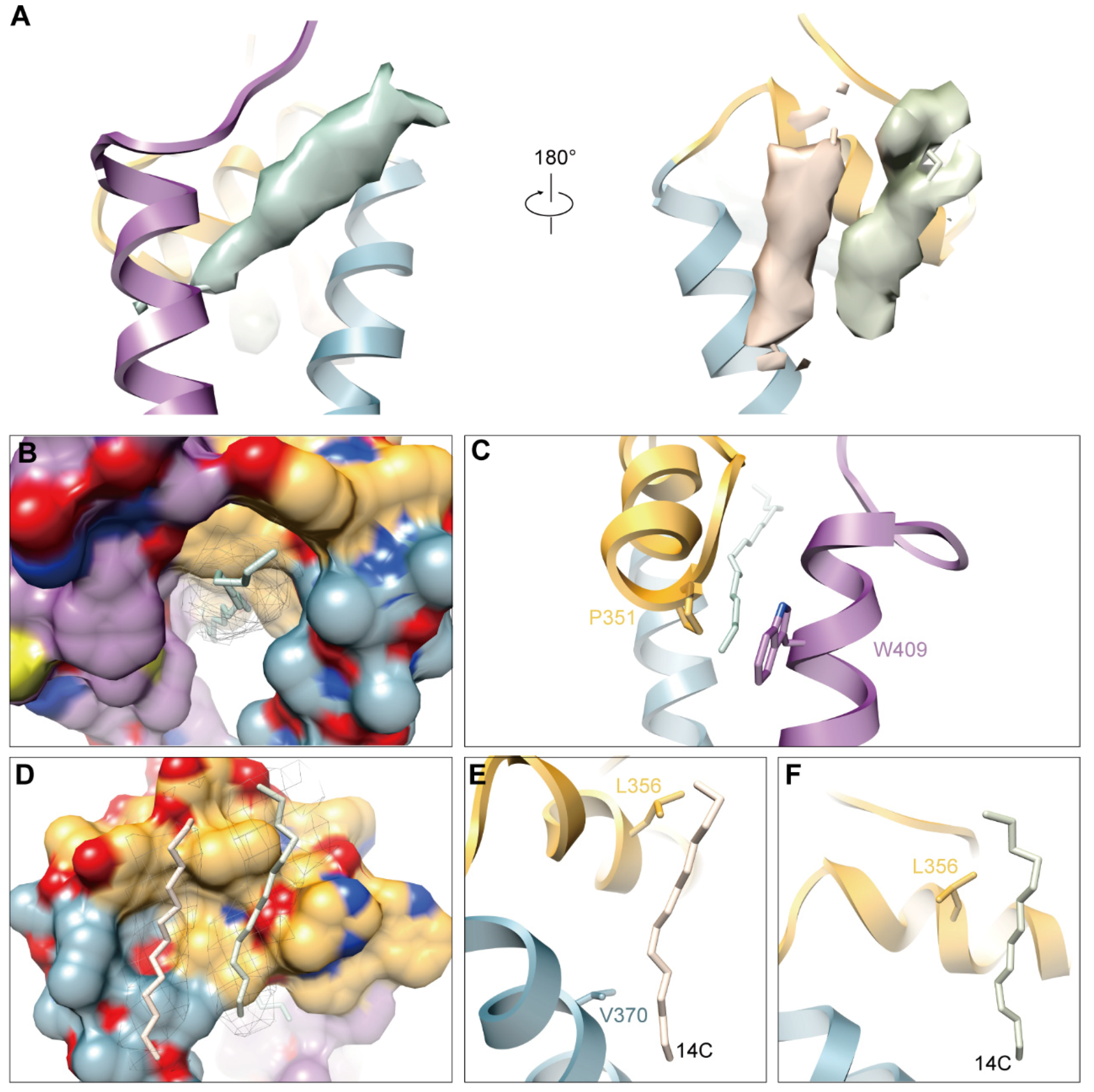
3.5. Hydrophobic Pocket and Newly Discovered Densities in GETV
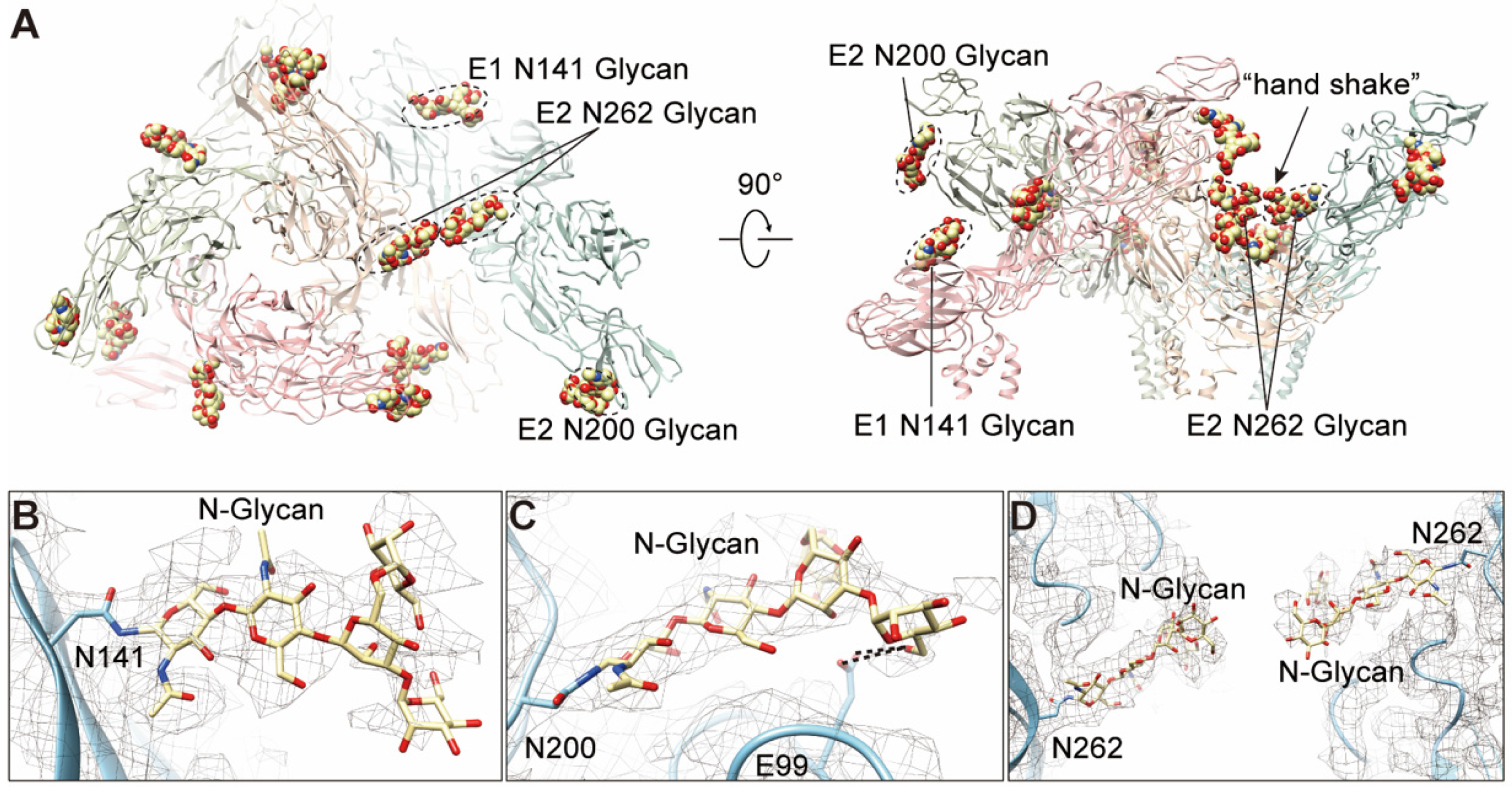
3.6. N-glycosylations in E1 and E2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Esposito, D.L.A.; Fonseca, B. Will Mayaro virus be responsible for the next outbreak of an arthropod-borne virus in Brazil? Braz. J. Infect. Dis. 2017, 21, 540–544. [Google Scholar] [CrossRef] [PubMed]
- Tjaden, N.B.; Cheng, Y.; Beierkuhnlein, C.; Thomas, S.M. Chikungunya Beyond the Tropics: Where and When Do We Expect Disease Transmission in Europe? Viruses 2021, 13, 1024. [Google Scholar] [CrossRef]
- Calisher, C.H. Medically important arboviruses of the United States and Canada. Clin. Microbiol. Rev. 1994, 7, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Kril, V.; Aiqui-Reboul-Paviet, O.; Briant, L.; Amara, A. New Insights into Chikungunya Virus Infection and Pathogenesis. Annu. Rev. Virol. 2021, 8, 327–347. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.J.; Kim, H.C.; Klein, T.A.; Park, J.Y.; Cho, Y.S.; Cho, I.S.; Kyung, S.G.; Yeh, J.Y. Characterization of recent Getah virus isolates from South Korea. Acta Virol. 2012, 56, 265–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherer, W.F.; Funkenbusch, M.; Buescher, E.L.; Izumit. Sagiyama virus, a new group A arthropod-borne virus from Japan. I. Isolation, immunologic classification, and ecologic observations. Am. J. Trop. Med. Hyg. 1962, 11, 255–268. [Google Scholar] [CrossRef]
- Spradbrow, P.B. A survey for arbovirus antibodies in pigs and sheep in Queensland. Aust. Vet. J. 1972, 48, 402–407. [Google Scholar] [CrossRef]
- Li, X.D.; Qiu, F.X.; Yang, H.; Rao, Y.N.; Calisher, C.H. Isolation of Getah virus from mosquitos collected on Hainan Island, China, and results of a serosurvey. Southeast Asian J. Trop. Med. Public Health 1992, 23, 730–734. [Google Scholar]
- Li, Y.Y.; Liu, H.; Fu, S.H.; Li, X.L.; Guo, X.F.; Li, M.H.; Feng, Y.; Chen, W.X.; Wang, L.H.; Lei, W.W.; et al. From discovery to spread: The evolution and phylogeny of Getah virus. Infect. Genet Evol. 2017, 55, 48–55. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, X.; Li, L.X.; Shi, N.; Sun, X.T.; Liu, Q.; Jin, N.Y.; Si, X.K. First isolation and characterization of Getah virus from cattle in northeastern China. BMC Vet. Res. 2019, 15, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, G.; Ou, J.; Ji, J.; Ren, Z.; Hu, X.; Wang, C.; Li, S. Emergence of Getah Virus Infection in Horse with Fever in China, 2018. Front. Microbiol. 2019, 10, 1416. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Li, R.; Hu, Y.; Yang, L.; Zhao, D.; Du, L.; Li, J.; Ge, M.; Yu, X. An outbreak of Getah virus infection among pigs in China, 2017. Transbound Emerg. Dis. 2018, 65, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.; Li, L.X.; Lu, R.G.; Yan, X.J.; Liu, H. Highly Pathogenic Swine Getah Virus in Blue Foxes, Eastern China, 2017. Emerg. Infect. Dis. 2019, 25, 1252–1254. [Google Scholar] [CrossRef]
- Nemoto, M.; Bannai, H.; Tsujimura, K.; Kobayashi, M.; Kikuchi, T.; Yamanaka, T.; Kondo, T. Getah Virus Infection among Racehorses, Japan, 2014. Emerg. Infect. Dis. 2015, 21, 883–885. [Google Scholar] [CrossRef]
- Kawamura, H.; Yago, K.; Narita, M.; Imada, T.; Nishimori, T.; Haritani, M. A fatal case in newborn piglets with Getah virus infection: Pathogenicity of the isolate. Nihon Juigaku Zasshi 1987, 49, 1003–1007. [Google Scholar] [CrossRef]
- Band, A.M.; Maatta, J.; Kaariainen, L.; Kuismanen, E. Inhibition of the membrane fusion machinery prevents exit from the TGN and proteolytic processing by furin. FEBS Lett. 2001, 505, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Hasan, S.S.; Sun, C.; Kim, A.S.; Watanabe, Y.; Chen, C.L.; Klose, T.; Buda, G.; Crispin, M.; Diamond, M.S.; Klimstra, W.B.; et al. Cryo-EM Structures of Eastern Equine Encephalitis Virus Reveal Mechanisms of Virus Disassembly and Antibody Neutralization. Cell Rep. 2018, 25, 3136–3147.e5. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro-Filho, H.V.; Coimbra, L.D.; Cassago, A.; Rocha, R.P.F.; Guerra, J.; de Felicio, R.; Carnieli, C.M.; Leme, L.; Padilha, A.C.; Paes Leme, A.F.; et al. Cryo-EM structure of the mature and infective Mayaro virus at 4.4 A resolution reveals features of arthritogenic alphaviruses. Nat. Commun. 2021, 12, 3038. [Google Scholar] [CrossRef]
- Chen, L.; Wang, M.; Zhu, D.; Sun, Z.; Ma, J.; Wang, J.; Kong, L.; Wang, S.; Liu, Z.; Wei, L.; et al. Implication for alphavirus host-cell entry and assembly indicated by a 3.5A resolution cryo-EM structure. Nat. Commun. 2018, 9, 5326. [Google Scholar] [CrossRef] [Green Version]
- Voss, J.E.; Vaney, M.C.; Duquerroy, S.; Vonrhein, C.; Girard-Blanc, C.; Crublet, E.; Thompson, A.; Bricogne, G.; Rey, F.A. Glycoprotein organization of Chikungunya virus particles revealed by X-ray crystallography. Nature 2010, 468, 709–712. [Google Scholar] [CrossRef] [PubMed]
- Lescar, J.; Roussel, A.; Wien, M.W.; Navaza, J.; Fuller, S.D.; Wengler, G.; Wengler, G.; Rey, F.A. The Fusion glycoprotein shell of Semliki Forest virus: An icosahedral assembly primed for fusogenic activation at endosomal pH. Cell 2001, 105, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Fields, W.; Kielian, M. A key interaction between the alphavirus envelope proteins responsible for initial dimer dissociation during fusion. J. Virol. 2013, 87, 3774–3781. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Kim, A.S.; Kafai, N.M.; Earnest, J.T.; Shah, A.P.; Case, J.B.; Basore, K.; Gilliland, T.C.; Sun, C.; Nelson, C.A.; et al. LDLRAD3 is a receptor for Venezuelan equine encephalitis virus. Nature 2020, 588, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Taylor, A. Arthritogenic Alphavirus Capsid Protein. Life 2021, 11, 230. [Google Scholar] [CrossRef]
- Zhang, R.; Hryc, C.F.; Cong, Y.; Liu, X.; Jakana, J.; Gorchakov, R.; Baker, M.L.; Weaver, S.C.; Chiu, W. 4.4 A cryo-EM structure of an enveloped alphavirus Venezuelan equine encephalitis virus. EMBO J. 2011, 30, 3854–3863. [Google Scholar] [CrossRef] [Green Version]
- Kostyuchenko, V.A.; Jakana, J.; Liu, X.; Haddow, A.D.; Aung, M.; Weaver, S.C.; Chiu, W.; Lok, S.M. The structure of barmah forest virus as revealed by cryo-electron microscopy at a 6-angstrom resolution has detailed transmembrane protein architecture and interactions. J. Virol. 2011, 85, 9327–9333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, M.B.; Weaver, S.C. Structure of the recombinant alphavirus Western equine encephalitis virus revealed by cryoelectron microscopy. J. Virol. 2010, 84, 9775–9782. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.C.; Chen, C.; Rayaprolu, V.; Mukhopadhyay, S.; Zlotnick, A. Self-Assembly of an Alphavirus Core-like Particle Is Distinguished by Strong Intersubunit Association Energy and Structural Defects. ACS Nano 2015, 9, 8898–8906. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Huang, X.; Cheng, J.; Zhu, D.; Zhang, X. High-quality, high-throughput cryo-electron microscopy data collection via beam tilt and astigmatism-free beam-image shift. J. Struct. Biol. 2019, 208, 107396. [Google Scholar] [CrossRef]
- Mastronarde, D.N. Automated electron microscope tomography using robust prediction of specimen movements. J. Struct. Biol. 2005, 152, 36–51. [Google Scholar] [CrossRef]
- Rohou, A.; Grigorieff, N. CTFFIND4: Fast and accurate defocus estimation from electron micrographs. J. Struct. Biol. 2015, 192, 216–221. [Google Scholar] [CrossRef]
- Zheng, S.Q.; Palovcak, E.; Armache, J.P.; Verba, K.A.; Cheng, Y.; Agard, D.A. MotionCor2: Anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 2017, 14, 331–332. [Google Scholar] [CrossRef] [Green Version]
- Scheres, S.H. RELION: Implementation of a Bayesian approach to cryo-EM structure determination. J. Struct. Biol. 2012, 180, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Wang, X.; Fang, Q.; Van Etten, J.L.; Rossmann, M.G.; Rao, Z.; Zhang, X. Pushing the resolution limit by correcting the Ewald sphere effect in single-particle Cryo-EM reconstructions. Nat. Commun. 2018, 9, 1552. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonine, P.V.; Poon, B.K.; Read, R.J.; Sobolev, O.V.; Terwilliger, T.C.; Urzhumtsev, A.; Adams, P.D. Real-space refinement in PHENIX for cryo-EM and crystallography. Acta Crystallogr. D Struct. Biol. 2018, 74, 531–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320-4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kielian, M. Class II virus membrane fusion proteins. Virology 2006, 344, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-San Martin, C.; Sosa, H.; Kielian, M. A stable prefusion intermediate of the alphavirus fusion protein reveals critical features of class II membrane fusion. Cell Host Microbe 2008, 4, 600–608. [Google Scholar] [CrossRef] [Green Version]
- Byrd, E.A.; Kielian, M. An Alphavirus E2 Membrane-Proximal Domain Promotes Envelope Protein Lateral Interactions and Virus Budding. mBio 2017, 8, e01564-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, M.A.; Herrero, L.J.; Jeffery, J.A.L.; Hoehn, M.; Rudd, P.A.; Supramaniam, A.; Kay, B.H.; Ryan, P.A.; Mahalingam, S. Role of envelope N-linked glycosylation in Ross River virus virulence and transmission. J. Gen. Virol. 2016, 97, 1094–1106. [Google Scholar] [CrossRef]
- Li, Y.; Liu, D.; Wang, Y.; Su, W.; Liu, G.; Dong, W. The Importance of Glycans of Viral and Host Proteins in Enveloped Virus Infection. Front. Immunol. 2021, 12, 638573. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.M.; Li, X.H.; Mom, V.; Lu, Z.H.; Liao, X.W.; Han, Y.; Pichoud, C.; Gong, Q.M.; Zhang, D.H.; Zhang, Y.; et al. N-glycosylation mutations within hepatitis B virus surface major hydrophilic region contribute mostly to immune escape. J. Hepatol. 2014, 60, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, C.; Pristatsky, P.; Hoang, V.M.; Casimiro, D.R.; Schwartz, R.M.; Rustandi, R.; Ha, S. Characterization of N-glycosylation profiles from mammalian and insect cell derived chikungunya VLP. J. Chromatogr. B Analyt. Technol. Biomed Life Sci. 2016, 1032, 218–223. [Google Scholar] [CrossRef]
- Knight, R.L.; Schultz, K.L.; Kent, R.J.; Venkatesan, M.; Griffin, D.E. Role of N-linked glycosylation for sindbis virus infection and replication in vertebrate and invertebrate systems. J. Virol. 2009, 83, 5640–5647. [Google Scholar] [CrossRef] [Green Version]
- Basore, K.; Kim, A.S.; Nelson, C.A.; Zhang, R.; Smith, B.K.; Uranga, C.; Vang, L.; Cheng, M.; Gross, M.L.; Smith, J.; et al. Cryo-EM Structure of Chikungunya Virus in Complex with the Mxra8 Receptor. Cell 2019, 177, 1725–1737.e16. [Google Scholar] [CrossRef]
- Parvate, A.D.; Vago, F.; Hasan, S.S.; Lee, J.; Williams, E.P.; Lanman, J.; Jonsson, C.B. A new inactivation method to facilitate cryo-EM of enveloped, RNA viruses requiring high containment: A case study using Venezuelan Equine Encephalitis Virus (VEEV). J. Virol. Methods 2020, 277, 113792. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Sun, Z.; Cui, C.; Wang, S.; Yang, D.; Shi, Z.; Wei, X.; Wang, P.; Sun, W.; Zhu, J.; et al. Structural Insights into Alphavirus Assembly Revealed by the Cryo-EM Structure of Getah Virus. Viruses 2022, 14, 327. https://doi.org/10.3390/v14020327
Wang M, Sun Z, Cui C, Wang S, Yang D, Shi Z, Wei X, Wang P, Sun W, Zhu J, et al. Structural Insights into Alphavirus Assembly Revealed by the Cryo-EM Structure of Getah Virus. Viruses. 2022; 14(2):327. https://doi.org/10.3390/v14020327
Chicago/Turabian StyleWang, Ming, Zhenzhao Sun, Chenxi Cui, Shida Wang, Decheng Yang, Zhibin Shi, Xinyu Wei, Pengfei Wang, Weiyao Sun, Jing Zhu, and et al. 2022. "Structural Insights into Alphavirus Assembly Revealed by the Cryo-EM Structure of Getah Virus" Viruses 14, no. 2: 327. https://doi.org/10.3390/v14020327