
Development of Antibodies against HPV-6 and HPV-11 for the Study of Laryngeal Papilloma

 ,
, 
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Detection of HPV DNA
2.3. Measurement of Viral DNA Load and mRNA Expression in HPV-11-Infected Papilloma by Quantitative Real-Time PCR
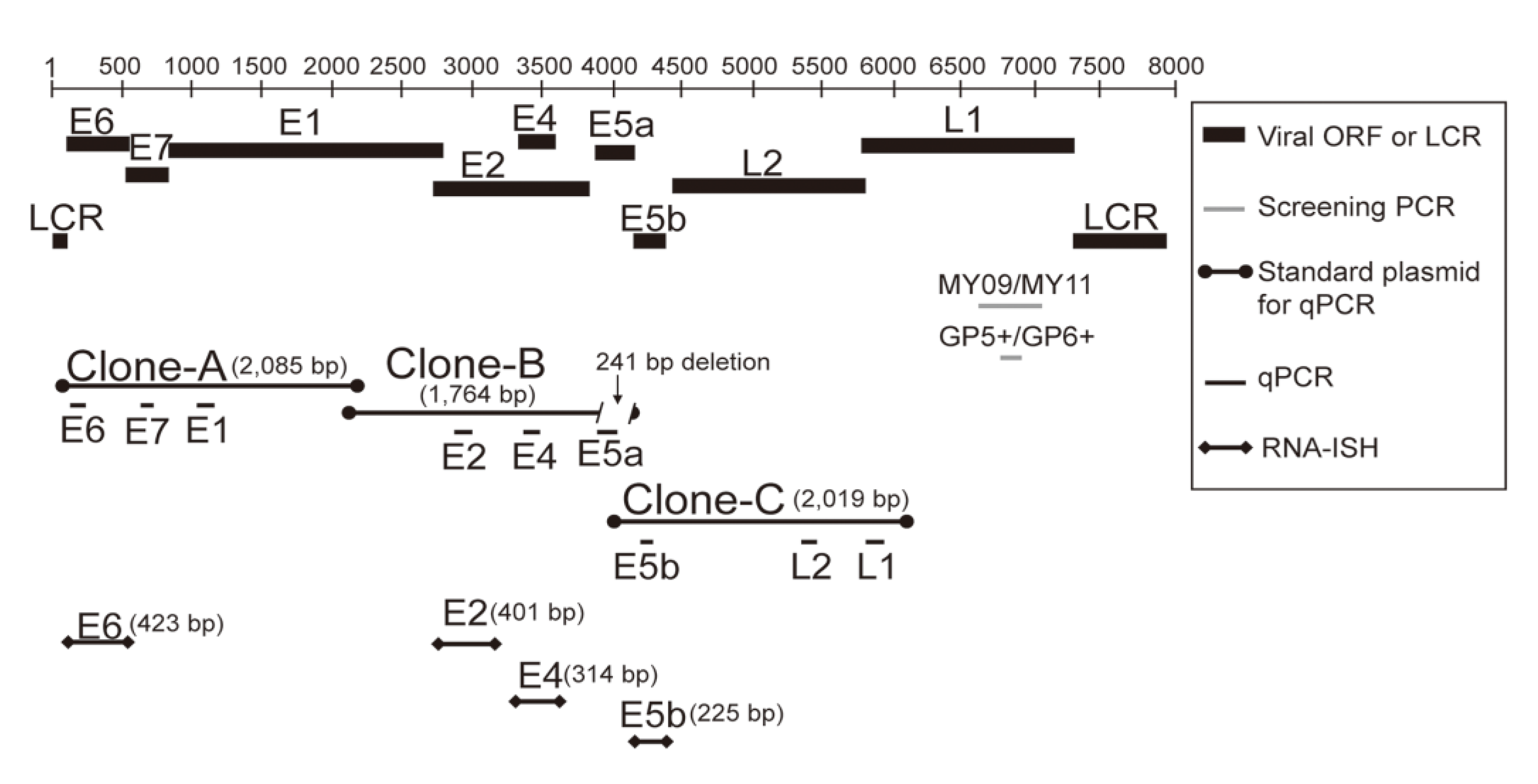
2.4. ISH with HPV DNA Probes
2.5. RNA-ISH with HPV-11 E6, E2, E4, and E5b Digoxigenin RNA Probes
2.6. Generation of an Anti-HPV-11 E1^E4 Polyclonal Antibody
2.6.1. Preparation of the Target Antigen
2.6.2. Preparation of the Polyclonal Antibody against HPV-11 E1^E4
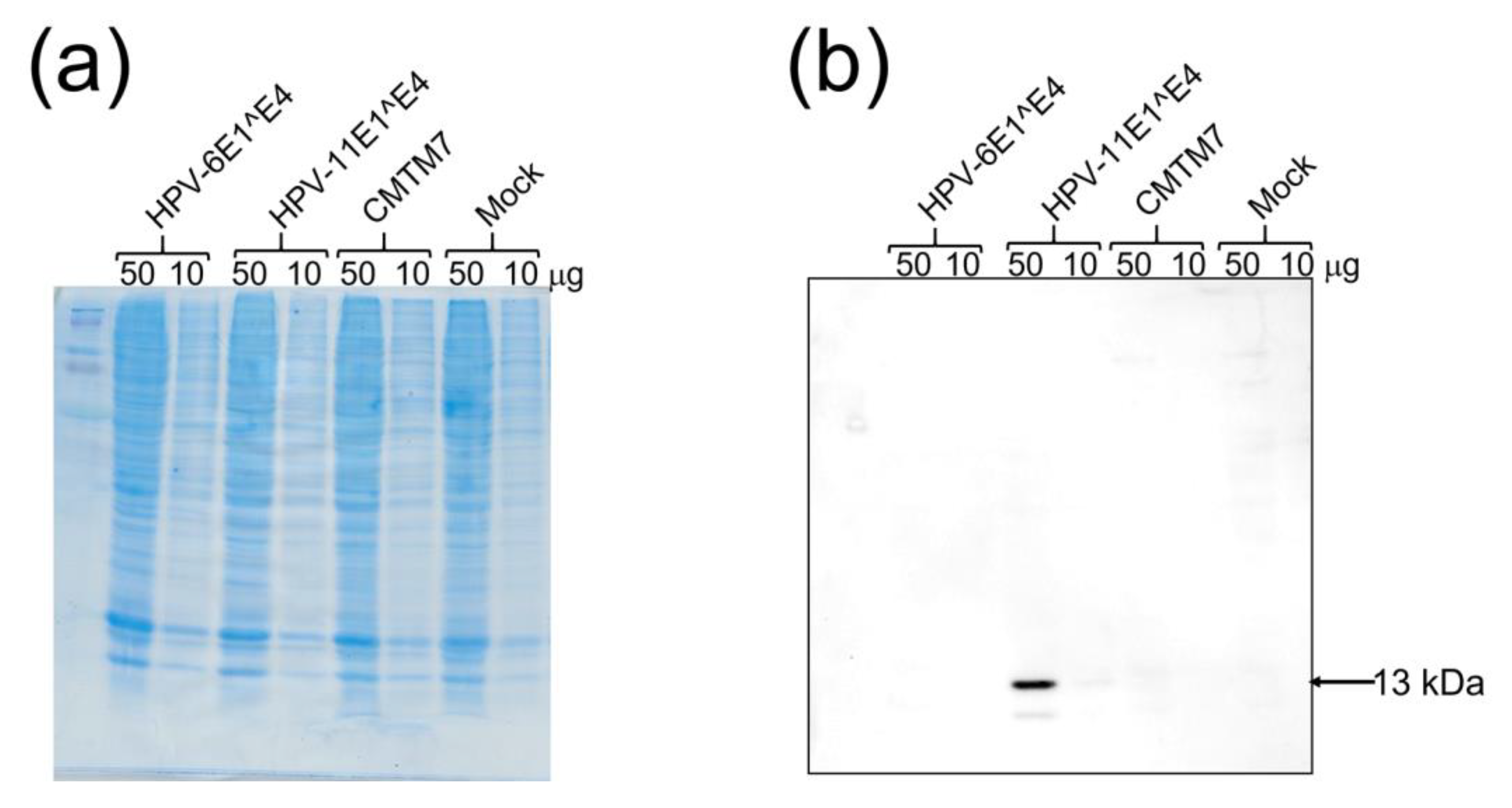
2.7. Western Blotting
2.8. Immunohistochemistry Using the Newly Developed Anti-HPV-11 E1^E4 Antibody and Anti-HPV-6 E1^E4 Antibody
2.9. Statistical Analysis
2.10. Ethics Approval and Informed Consent
3. Results
3.1. HPV-11 DNA Distribution, Subtypes, and Viral Load
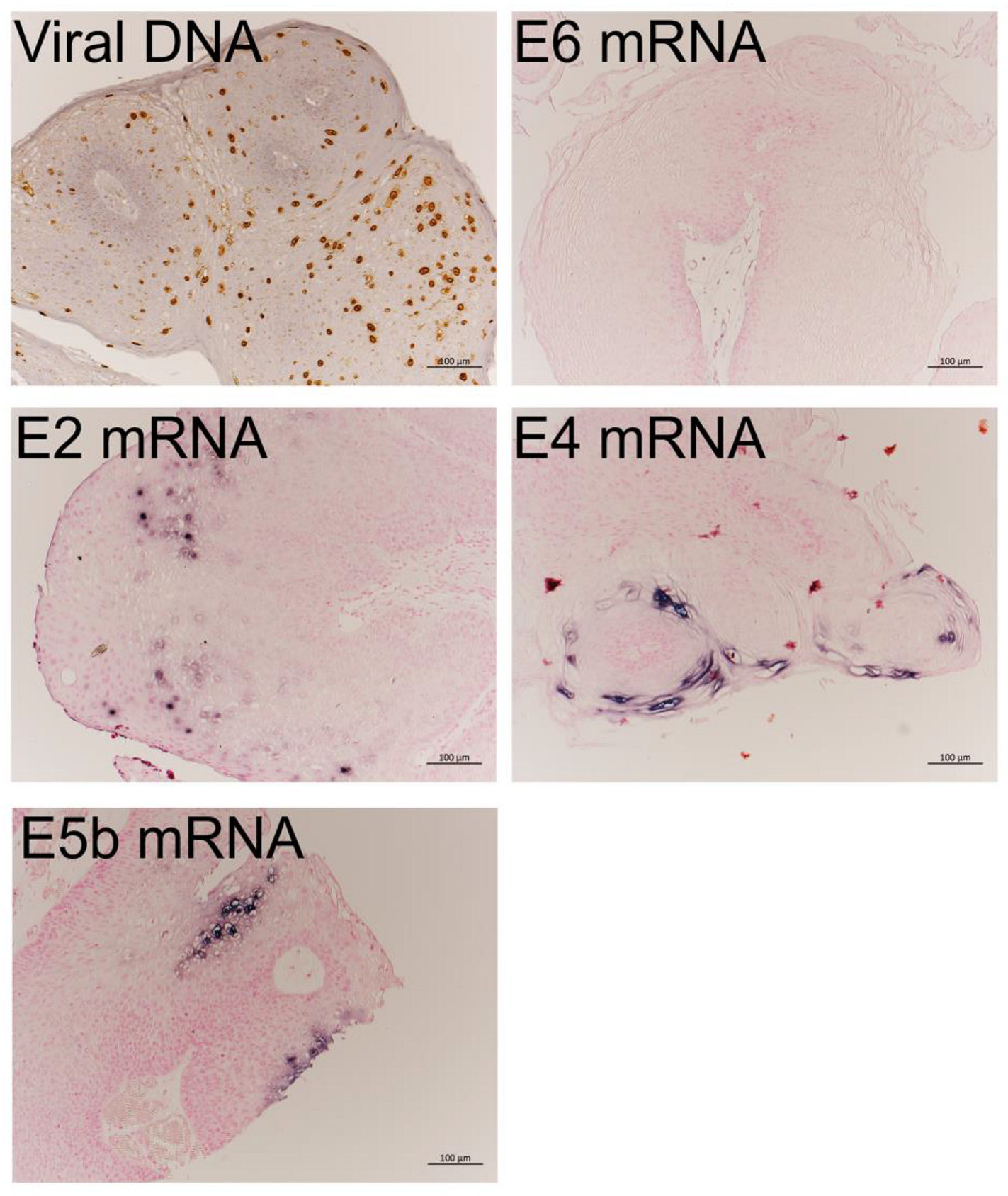
3.2. Expression of HPV-11 mRNAs
3.2.1. Expression Levels of the Nine Viral mRNAs
3.2.2. mRNA Expression in LP Detected by RNA-ISH
3.3. Western Blot Analysis Using the Anti-HPV-11 E1^E4 Antibody
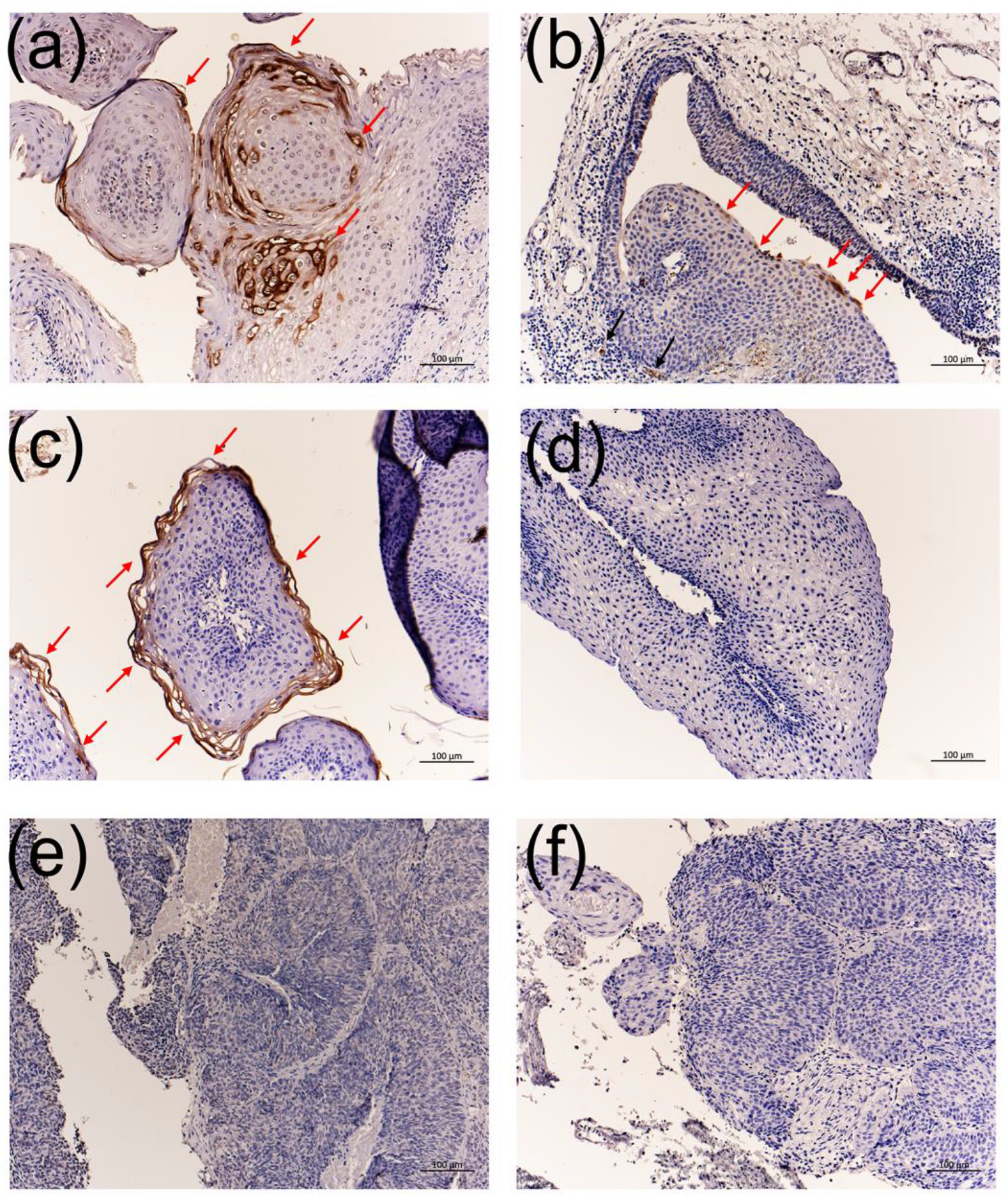
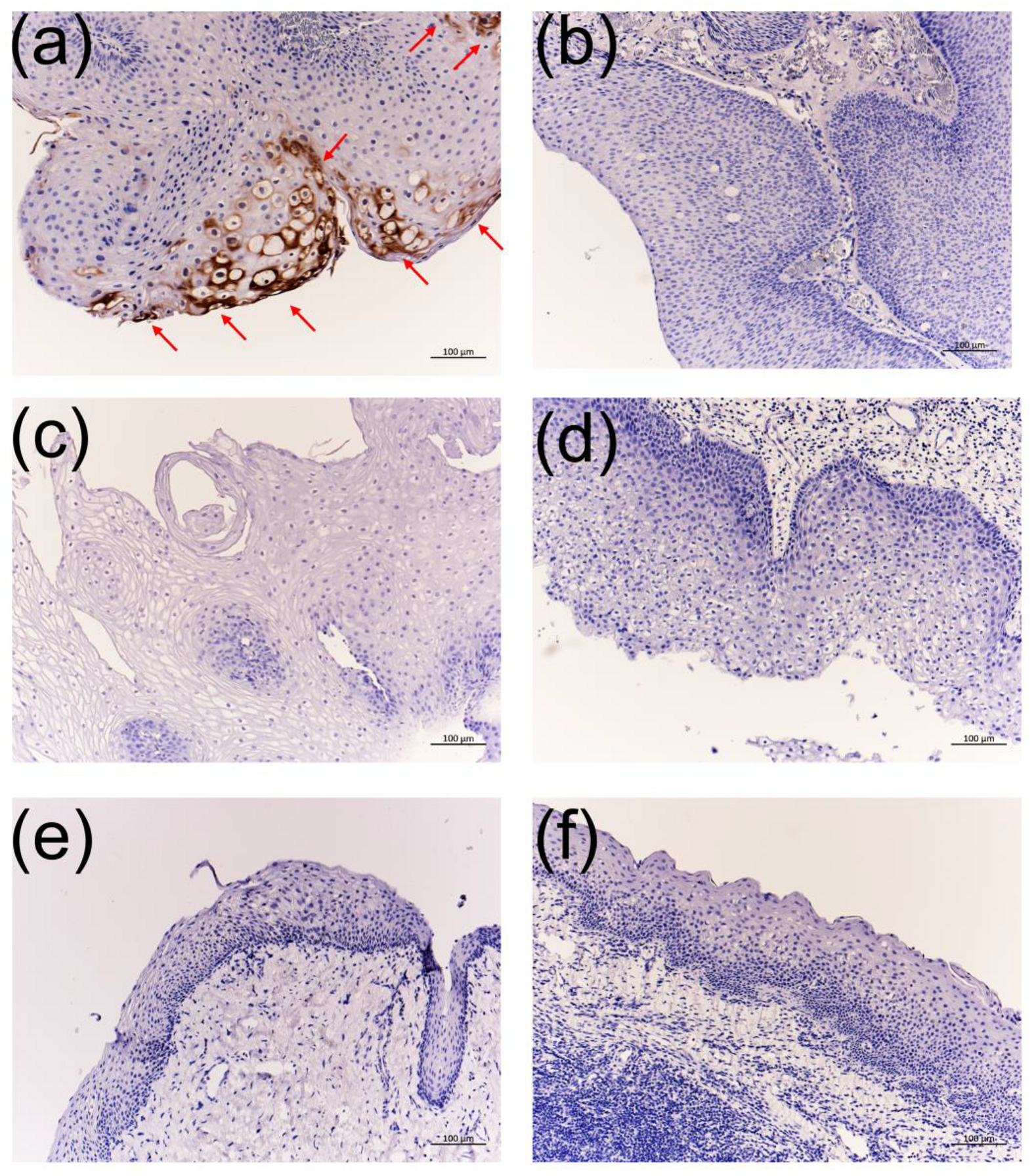
3.4. Immunohistochemistry Using the Anti-HPV-11 and Anti-HPV-6 E1^E4 Antibodies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doorbar, J.; Quint, W.; Banks, L.; Bravo, I.G.; Stoler, M.; Broker, T.R.; Stanley, M.A. The biology and life-cycle of human papillomaviruses. Vaccine 2012, 30, F55–F70. [Google Scholar] [CrossRef] [PubMed]
- Fortes, H.R.; von Ranke, F.M.; Escuissato, D.L.; Araujo Neto, C.A.; Zanetti, G.; Hochhegger, B.; Souza, C.A.; Marchiori, E. Recurrent respiratory papillomatosis: A state-of-the-art review. Respir. Med. 2017, 126, 116–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanazawa, T.; Fukushima, N.; Imayoshi, S.; Nagamoto, T.; Kawada, K.; Nishino, H.; Ichimura, K. Rare case of malignant transformation of recurrent respiratory papillomatosis associated with human papillomavirus type 6 infection and p53 overexpression. SpringerPlus 2013, 2, 153. [Google Scholar] [CrossRef] [Green Version]
- Schraff, S.; Derkay, C.S.; Burke, B.; Lawson, L. American society of pediatric otolaryngology members’ experience with recurrent respiratory papillomatosis and the use of adjuvant therapy. Arch. Otolaryngol. Head Neck Surg. 2004, 130, 1039–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, J.L.; Cohen, S.; Evjen, A.N.; Law, J.H.; Vadivelu, S.; Attia, A.; Schindler, J.S.; Chung, C.H.; Wirth, P.S.; Meijer, C.J.L.M.; et al. Human papillomavirus in early laryngeal carcinoma. Laryngoscope 2009, 119, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Benyo, S.; Keane, A.; Warrick, J.; Choi, K.Y. Hpv-positive oral papillomas in an adolescent-a diagnostic dilemma. Clin. Case Rep. 2021, 9, e04546. [Google Scholar] [CrossRef]
- Amiling, R.; Meites, E.; Querec, T.D.; Stone, L.; Singh, V.; Unger, E.R.; Derkay, C.S.; Markowitz, L.E. Juvenile-onset recurrent respiratory papillomatosis in the united states, epidemiology and hpv types-2015–2020. J. Pediatric Infect. Dis. Soc. 2021, 10, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Woodman, C.B.; Collins, S.I.; Young, L.S. The natural history of cervical hpv infection: Unresolved issues. Nat. Rev. Cancer 2007, 7, 11–22. [Google Scholar] [CrossRef]
- Graham, S.V. The human papillomavirus replication cycle, and its links to cancer progression: A comprehensive review. Clin. Sci. 2017, 131, 2201–2221. [Google Scholar] [CrossRef] [Green Version]
- Egawa, N.; Doorbar, J. The low-risk papillomaviruses. Virus Res. 2017, 231, 119–127. [Google Scholar] [CrossRef]
- Ikegami, T.; Hirakawa, H.; Tsukahara, N.; Murakami, A.; Kise, N.; Kiyuna, A.; Kosugi, T.; Agena, S.; Kinjyo, H.; Hasegawa, N.; et al. Coordinated expression of hpv-6 genes with predominant e4 and e5 expression in laryngeal papilloma. Microorganisms 2021, 9, 520. [Google Scholar] [CrossRef]
- Deng, Z.; Ikegami, T.; Kiyuna, A.; Zhang, C.; Zhang, T.; Matayoshi, S.; Uehara, T.; Maeda, H.; Suzuki, M.; Ganaha, A. Methylation of cpg sites in the upstream regulatory region, physical status and mrna expression of hpv-6 in adult-onset laryngeal papilloma. Oncotarget 2017, 8, 85368–85377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, Y.; Ikegami, T.; Hirakawa, H.; Uehara, T.; Deng, Z.; Agena, S.; Uezato, J.; Kondo, S.; Kiyuna, A.; Maeda, H.; et al. Staging and prognosis of oropharyngeal carcinoma according to the 8th edition of the american joint committee on cancer staging manual in human papillomavirus infection. Eur. Arch. Otorhinolaryngol. 2019, 276, 827–836. [Google Scholar] [CrossRef] [Green Version]
- Kiyuna, A.; Ikegami, T.; Uehara, T.; Hirakawa, H.; Agena, S.; Uezato, J.; Kondo, S.; Yamashita, Y.; Deng, Z.; Maeda, H.; et al. High-risk type human papillomavirus infection and p16 expression in laryngeal cancer. Infect. Agent. Cancer 2019, 14, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikegami, T.; Uehara, T.; Deng, Z.; Kondo, S.; Maeda, H.; Kiyuna, A.; Agena, S.; Hirakawa, H.; Yamashita, Y.; Ganaha, A.; et al. Detection of human papillomavirus in branchial cleft cysts. Oncol. Lett. 2018, 16, 1571–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, H.; Murakami, A.; Yoshida, M.; Muraoka, J.; Wakai, J.; Kenjyou, N.; Ito, Y. Characterization and selection of 3-(1-naphthoyl)-indole derivative-specific alpaca vhh antibodies using a phage display library. Monoclon. Antibodies Immunodiagn. Immunother. 2016, 35, 231–234. [Google Scholar] [CrossRef]
- Derkay, C.S.; Hester, R.P.; Burke, B.; Carron, J.; Lawson, L. Analysis of a staging assessment system for prediction of surgical interval in recurrent respiratory papillomatosis. Int. J. Pediatr. Otorhinolaryngol. 2004, 68, 1493–1498. [Google Scholar] [CrossRef]
- Doorbar, J.; Egawa, N.; Griffin, H.; Kranjec, C.; Murakami, I. Human papillomavirus molecular biology and disease association. Rev. Med. Virol. 2015, 25, 2–23. [Google Scholar] [CrossRef] [Green Version]
- Doorbar, J. The e4 protein; structure, function and patterns of expression. Virology 2013, 445, 80–98. [Google Scholar] [CrossRef] [Green Version]
- DiMaio, D.; Petti, L.M. The e5 proteins. Virology 2013, 445, 99–114. [Google Scholar] [CrossRef]
- Tsai, T.C.; Chen, S.L. The biochemical and biological functions of human papillomavirus type 16 e5 protein. Arch. Virol. 2003, 148, 1445–1453. [Google Scholar] [CrossRef]
- Ilahi, N.E.; Bhatti, A. Impact of hpv e5 on viral life cycle via egfr signaling. Microb. Pathog. 2020, 139, 103923. [Google Scholar] [CrossRef] [PubMed]
- Venuti, A.; Paolini, F.; Nasir, L.; Corteggio, A.; Roperto, S.; Campo, M.; Borzacchiello, G. Papillomavirus e5: The smallest oncoprotein with many functions. Mol. Cancer 2011, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Bravo, I.G.; Alonso, A. Mucosal human papillomaviruses encode four different e5 proteins whose chemistry and phylogeny correlate with malignant or benign growth. J. Virol. 2004, 78, 13613–13626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, A.L.; Caodaglio, A.S.; Sichero, L. Regulation of hpv transcription. Clinics 2018, 73, e486s. [Google Scholar] [CrossRef]
- Steinberg, B.M.; Auborn, K.J.; Brandsma, J.L.; Taichman, L.B. Tissue site-specific enhancer function of the upstream regulatory region of human papillomavirus type 11 in cultured keratinocytes. J. Virol. 1989, 63, 957–960. [Google Scholar] [CrossRef] [Green Version]
- Ottinger, M.; Smith, J.A.; Schweiger, M.R.; Robbins, D.; Powell, M.L.; You, J.; Howley, P.M. Cell-type specific transcriptional activities among different papillomavirus long control regions and their regulation by e2. Virology 2009, 395, 161–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunne, E.F.; Markowitz, L.E.; Saraiya, M.; Stokley, S.; Middleman, A.; Unger, E.R.; Williams, A.; Iskander, J. Cdc grand rounds: Reducing the burden of hpv-associated cancer and disease. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 69–72. [Google Scholar]
- Dartmann, K.; Schwarz, E.; Gissmann, L.; zur Hausen, H. The nucleotide sequence and genome organization of human papilloma virus type 11. Virology 1986, 151, 124–130. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Cases (n = 66) | Anti-HPV-6 E4 IHC | Anti-HPV-11 E4 IHC | |||
|---|---|---|---|---|---|
| Positive | Negative | Positive | Negative | ||
| Laryngeal papilloma | |||||
| HPV-6-positive | 20 | 16 | 4 | 0 | 20 |
| HPV-11-positive | 3 | 0 | 3 | 3 | 0 |
| HPV-negative | 5 | 0 | 5 | 0 | 5 |
| Nasoseptal exophytic papilloma | |||||
| HPV-6-positive | 1 | 0 | 1 | 0 | 1 |
| HPV-11-positive | 1 | 0 | 1 | 1 | 0 |
| HPV-negative | 2 | 0 | 2 | 0 | 2 |
| Paranasal exophytic papilloma (HPV-negative) | 1 | 0 | 1 | 0 | 1 |
| Nasopharyngeal papilloma (HPV-11-positive) | 1 | 0 | 1 | 1 | 0 |
| Oropharyngeal papilloma (HPV-negative) | 1 | 0 | 1 | 0 | 1 |
| Inverted papilloma with SCC (HPV-18-positive) | 1 | 0 | 1 | 0 | 1 |
| Normal vocal cord (HPV-negative) | 5 | 0 | 5 | 0 | 5 |
| Chronic tonsillitis (HPV-negative) | 5 | 0 | 5 | 0 | 5 |
| Laryngeal cancer | |||||
| High-risk HPV-positive 1 | 5 | 0 | 5 | 0 | 5 |
| HPV-negative | 5 | 0 | 5 | 0 | 5 |
| Oropharyngeal cancer | |||||
| High-risk HPV-infected (HPV-16, 33, 35, 56, 67) | 5 | 0 | 5 | 0 | 5 |
| Tongue cancer (HPV-negative) | 5 | 0 | 5 | 0 | 5 |
| Clinicopathological Characteristics | DNA Viral Load | HPV mRNA Expression/β-Actin | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Case | Age (Years) | Sex | Surgery | Number of Tumors | Subsite | Derkay Score | (Copies/ ng DNA) | E6 | E7 | E1 | E2 | E4 | E5a | E5b | L2 | L1 |
| 1 | 44 | M | 1st | multiple | Bil TVC | 5 | 64,192 | 0.11588 | 0.10541 | 0.23529 | 0.39033 | 3.95554 | 0.98977 | 2.33520 | 0.03630 | 0.02664 |
| 2 | 44 | M | 1st | multiple | Bil TVC | 7 | 226,916 | 0.38194 | 1.95009 | 6.36798 | 10.67637 | 56.84702 | 16.06985 | 39.08463 | 0.32797 | 0.11461 |
| 3 | 5 | M | 13th | multiple | E | 5 | 84 | 0.00316 | 0.06096 | 0.01683 | 0.02952 | 0.43326 | 0.20933 | 0.54957 | 0.00101 | 0.00059 |
| 5 | multiple | PW | 3 | 642,111 | 0.02153 | 0.04717 | 0.04567 | 0.09687 | 0.87938 | 0.20339 | 0.49420 | 0.00542 | 0.00482 | |||
| 5 | 14th | multiple | Bil TVC | 4 | 230,366 | 0.03669 | 0.03956 | 0.08475 | 0.13412 | 1.45394 | 0.36106 | 0.85643 | 0.00581 | 0.00167 | ||
| 6 | 15th | multiple | Bil TVC | 5 | 33,153 | 0.02311 | 0.55589 | 0.03616 | 0.12762 | 0.83358 | 0.89123 | 5.44958 | 0.00210 | 0.00170 | ||
| 7 | 16th | multiple | E | 2 | 15,711 | NA | NA | NA | NA | NA | NA | NA | NA | NA | ||
| 7 | multiple | Bil TVC | 4 | 601,070 | 0.04394 | 0.20269 | 0.03255 | 0.15081 | 1.16959 | 0.80580 | 1.18013 | 0.00918 | 0.00594 | |||
| 7 | multiple | PW | 2 | 71,468 | 0.18674 | 0.62598 | 0.11146 | 0.35295 | 4.58308 | 4.86911 | 5.82166 | 0.04036 | 0.06345 | |||
| 4 | 44 | F | 1st | multiple | NP | - | 11,346 | 0.17928 | 0.67253 | 0.08880 | 0.41194 | 3.45012 | 3.11286 | 4.59908 | 0.00536 | 0.00490 |
| 5 | 63 | M | 1st | multiple | NS | - | 246,077 | 0.17783 | 0.77629 | 0.47749 | 0.41261 | 4.80617 | 1.05967 | 2.48394 | 0.13504 | 0.10001 |
| HPV Status of Specimen | No. of Specimens | Anti-HPV-6 E4 IHC | Anti-HPV-11 E4 IHC | ||
|---|---|---|---|---|---|
| Positive | Negative | Positive | Negative | ||
| HPV-6-positive | 21 | 16 | 5 | 0 | 21 |
| HPV-11-positive | 5 | 0 | 5 | 5 | 0 |
| High-risk HPV-positive | 11 | 0 | 11 | 0 | 11 |
| HPV-negative | 29 | 0 | 29 | 0 | 29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikegami, T.; Kise, N.; Kinjyo, H.; Kondo, S.; Suzuki, M.; Tsukahara, N.; Murakami, A.; Kiyuna, A.; Agena, S.; Tanaka, K.; et al. Development of Antibodies against HPV-6 and HPV-11 for the Study of Laryngeal Papilloma. Viruses 2021, 13, 2024. https://doi.org/10.3390/v13102024
Ikegami T, Kise N, Kinjyo H, Kondo S, Suzuki M, Tsukahara N, Murakami A, Kiyuna A, Agena S, Tanaka K, et al. Development of Antibodies against HPV-6 and HPV-11 for the Study of Laryngeal Papilloma. Viruses. 2021; 13(10):2024. https://doi.org/10.3390/v13102024
Chicago/Turabian StyleIkegami, Taro, Norimoto Kise, Hidetoshi Kinjyo, Shunsuke Kondo, Mikio Suzuki, Narutoshi Tsukahara, Akikazu Murakami, Asanori Kiyuna, Shinya Agena, Katsunori Tanaka, and et al. 2021. "Development of Antibodies against HPV-6 and HPV-11 for the Study of Laryngeal Papilloma" Viruses 13, no. 10: 2024. https://doi.org/10.3390/v13102024