
An Investigation of Culicoides (Diptera: Ceratopogonidae) as Potential Vectors of Medically and Veterinary Important Arboviruses in South Africa


Abstract
:1. Introduction
2. Materials and Methods
3. Results
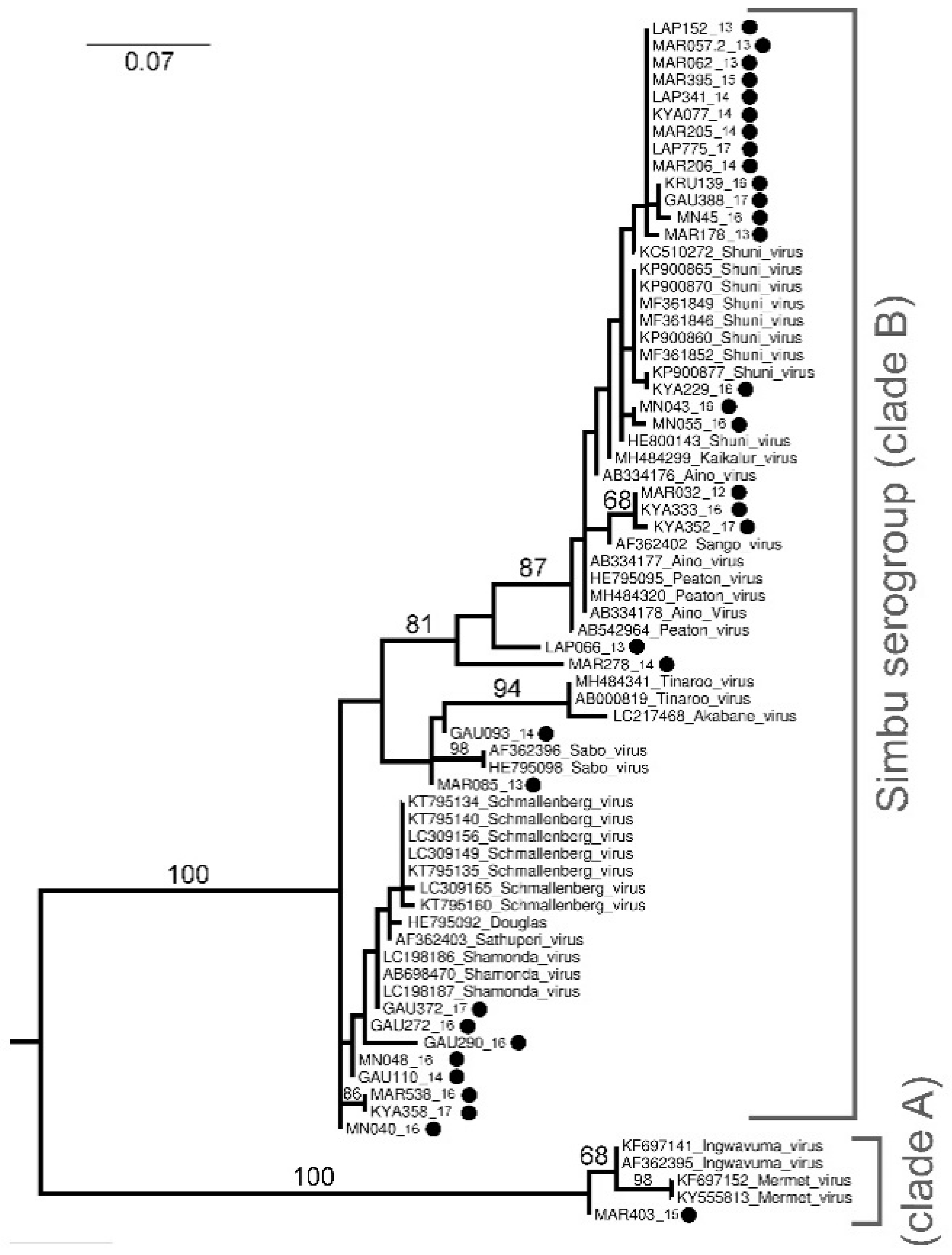
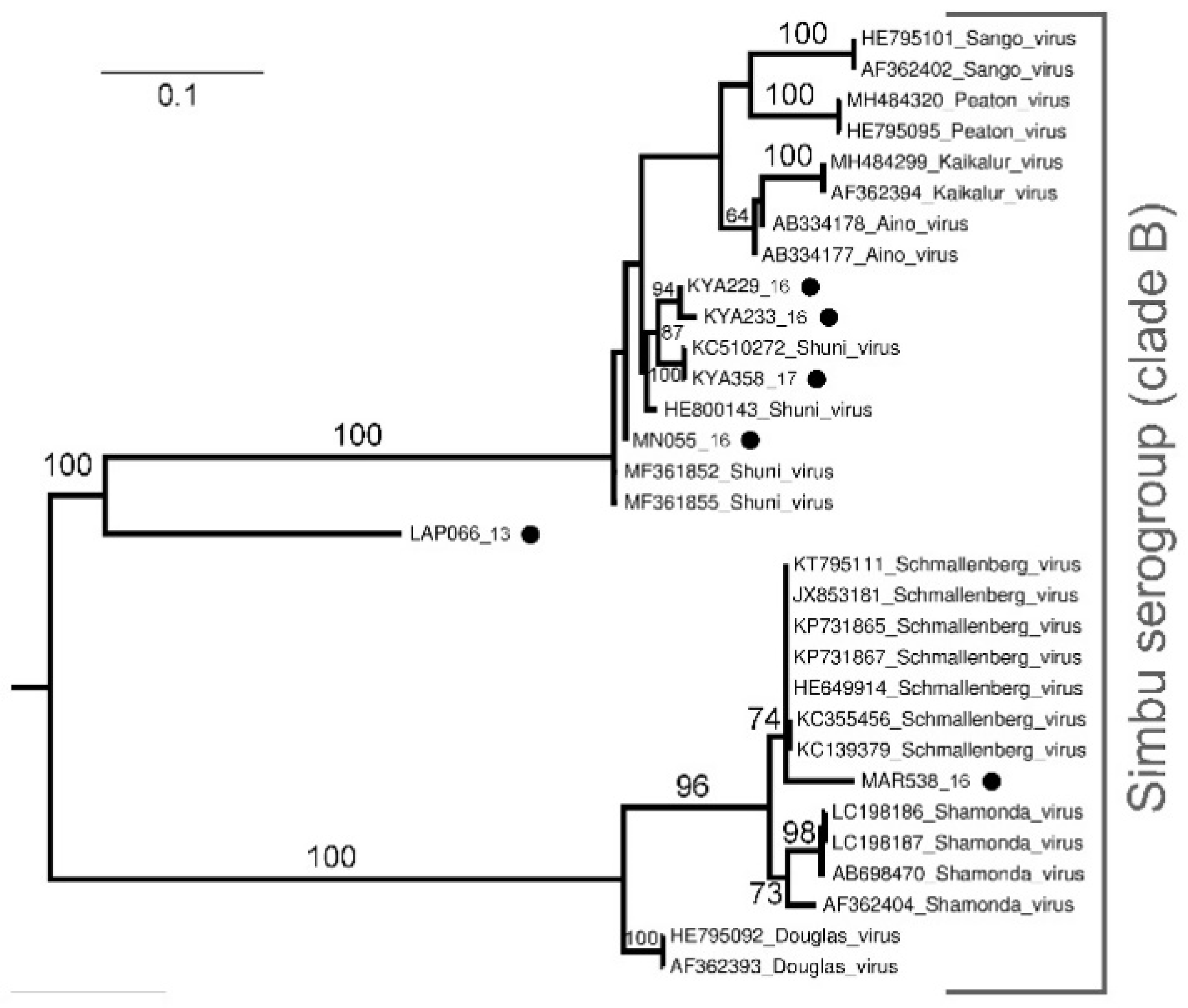
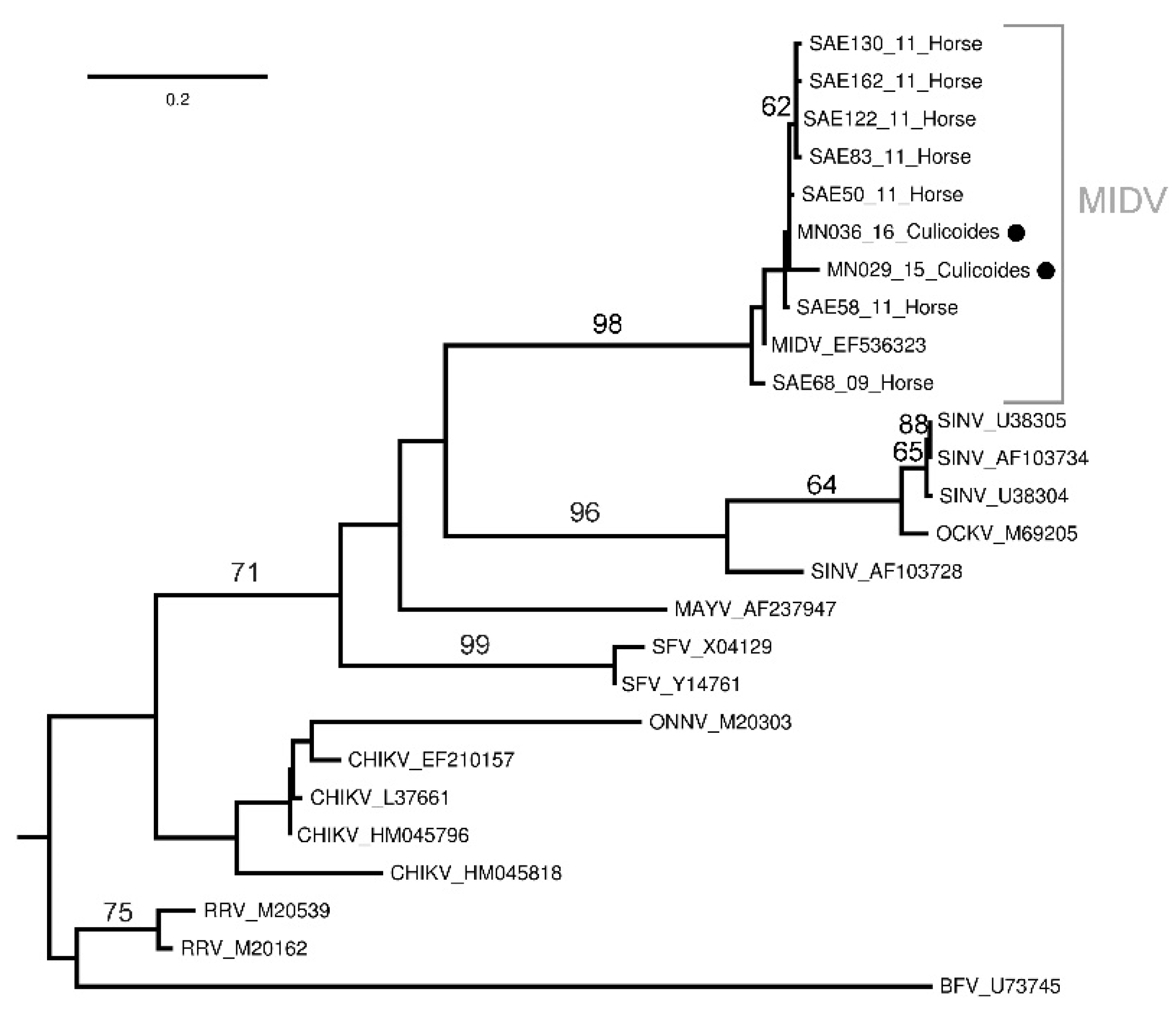
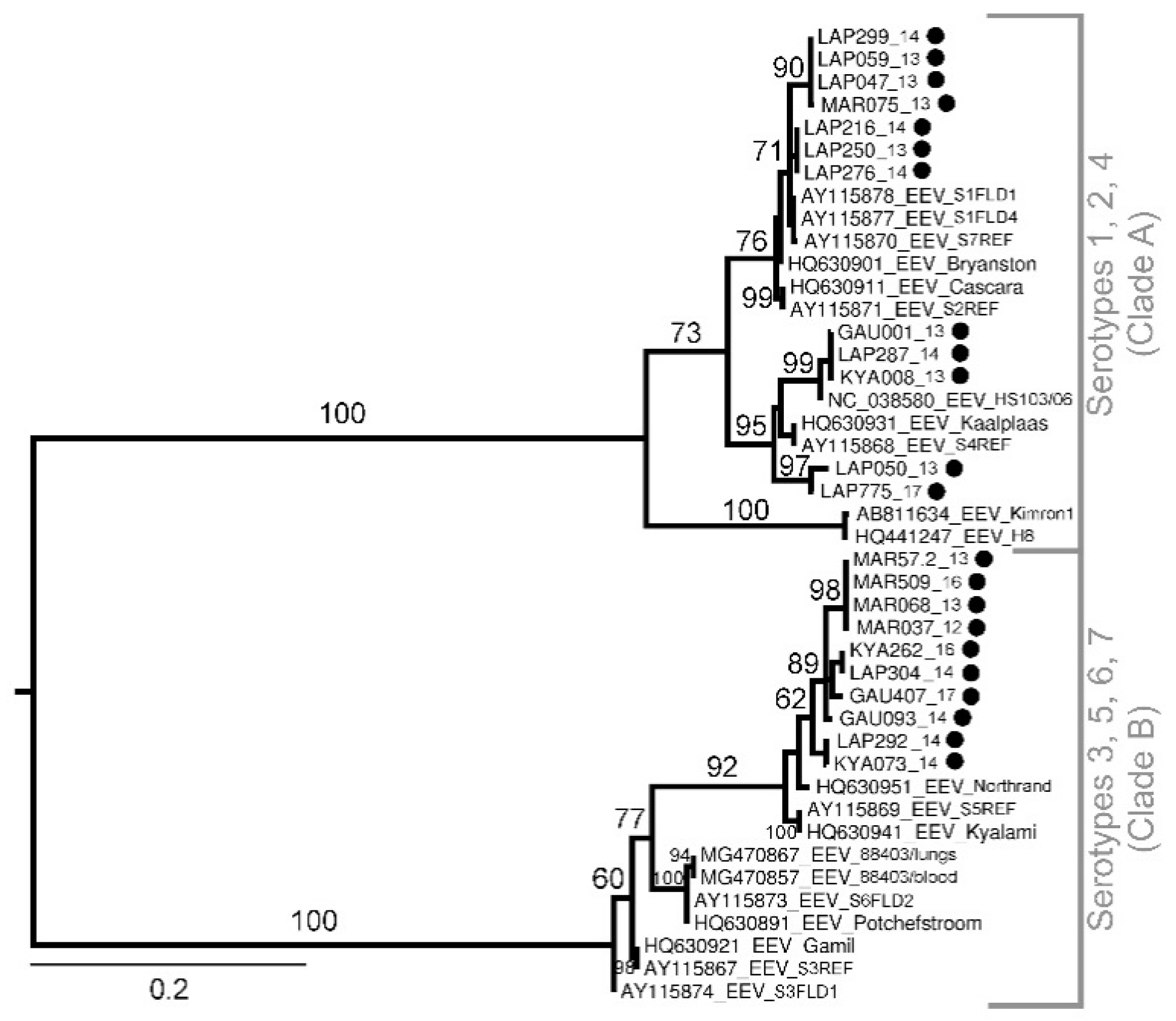
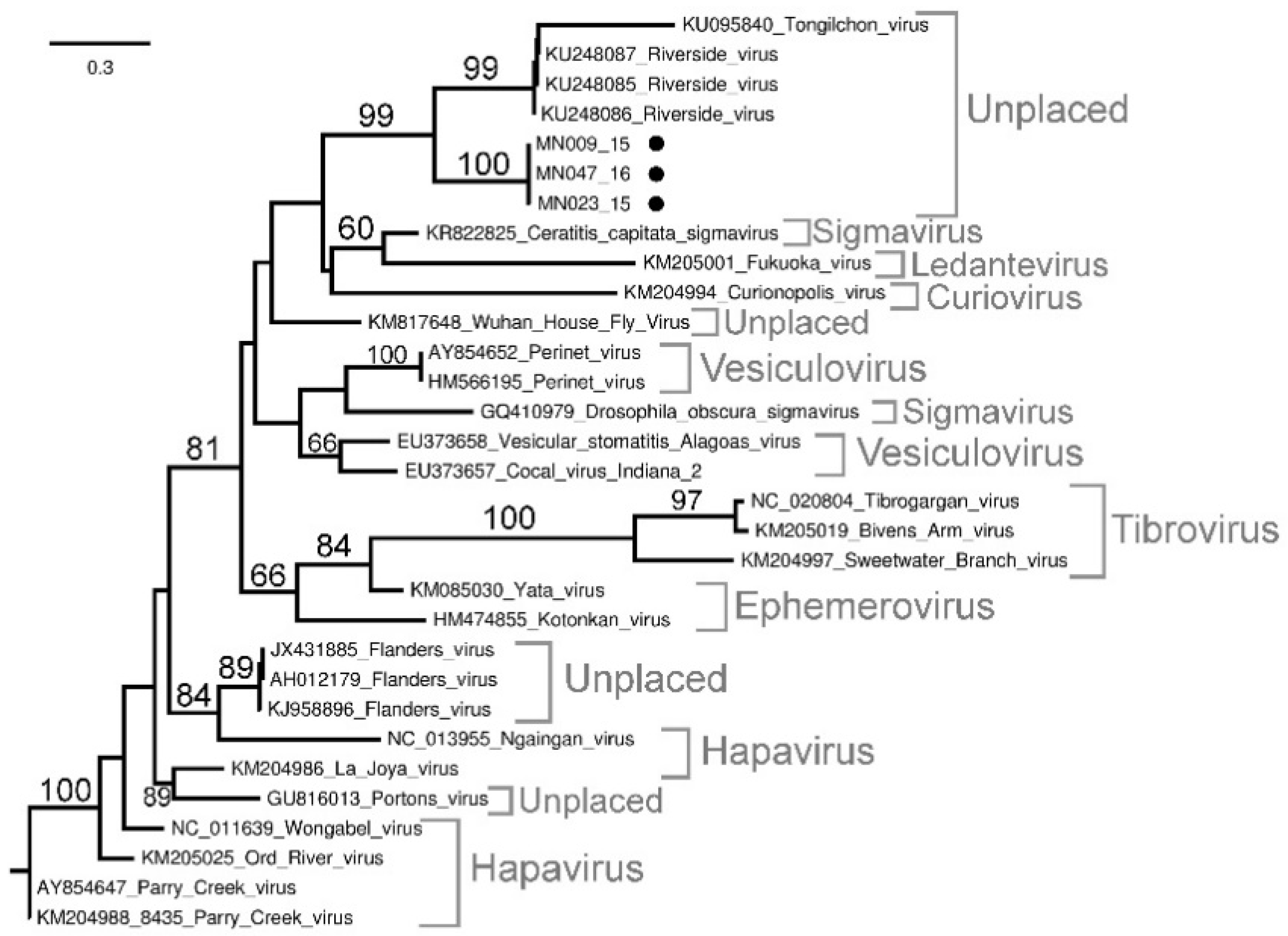
Sequencing and Phylogenetic Analysis of Positive Pools
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Mellor, P.S.; Boorman, J.; Baylis, M. Culicoides biting midges: Their role as arbovirus vectors. Annu. Rev. Entomol. 2000, 45, 307–340. [Google Scholar] [CrossRef]
- Carpenter, S.; Wilson, A.; Mellor, P.S. Culicoides and the emergence of bluetongue virus in northern Europe. Trends Microbiol. 2009, 17, 172–178. [Google Scholar] [CrossRef]
- Hoffmann, B.; Scheuch, M.; Höper, D.; Jungblut, R.; Holsteg, M.; Schirrmeier, H.; Eschbaumer, M.; Goller, K.V.; Wernike, K.; Fischer, M.; et al. Novel Orthobunyavirus in cattle, Europe, 2011. Emerg. Infect. Dis. 2018, 18, 469. [Google Scholar] [CrossRef]
- Grewar, J.D.; Weyer, C.T.; Venter, G.J.; van Helden, L.S.; Burger, P.; Guthrie, A.J.; Coetzee, P.; Labuschagne, K.; Bührmann, G.; Parker, B.J.; et al. A field investigation of an African horse sickness outbreak in the controlled area of South Africa in 2016. Transbound. Emerg. Dis. 2019, 66, 743–751. [Google Scholar] [CrossRef]
- Snyman, J.; Snyman, L.P.; Labuschagne, K.; Vente, G.J.; Venter, M. The utilisation of CytB and COI barcodes for the identification of bloodmeals and Culicoides species (Diptera: Ceratopogonidae) reveals a variety of novel wildlife hosts in South Africa. Acta Trop. 2021, 219, 105913. [Google Scholar] [CrossRef]
- Anderson, D.R.; Spence, L.; Downs, W.G.; Aitken, T.H.G. Oropouche virus: A new human disease agent from Trinidad, West Indies. Am. J. Trop. Med. Hyg. 1961, 10, 574. [Google Scholar] [CrossRef]
- Sabio, I.J.; Mackay, A.J.; Roy, A.; Foil, L.D. Detection of West Nile virus RNA in pools of three species of Ceratopogonids (Diptera: Ceratopogonidae) collected in Louisiana. J. Med. Entomol. 2006, 43, 1020–1022. [Google Scholar]
- Lee, V.H. Isolation of viruses from field populations of Culicoides (Diptera: Ceratopogonidae) in Nigeria. J. Med. Entomol. 1979, 16, 76–79. [Google Scholar] [CrossRef]
- Nevill, E.M.; Erasmus, B.J.; Venter, G.J. A six-year survey of viruses associated with Culicoides biting midges throughout South Africa (Diptera: Ceratopogonidae). In Bluetongue, African Horse Sickness and Related Orbiviruses, Proceedings of the Second International Symposium; Walton, T.E., Osburn, B.L., Eds.; CRC Press: Boca Raton, FL, USA, 1992; pp. 314–319. [Google Scholar]
- Venter, M.; Pretorius, M.; Fuller, J.A.; Botha, E.; Rakgotho, M.; Stivaktas, V.; Weyer, C.; Romito, M.; Williams, J. West Nile virus lineage 2 in horses and other animals with neurologic disease, South Africa, 2008–2015. Emerg. Infect. Dis. 2017, 23, 2060. [Google Scholar] [CrossRef]
- Steyn, J.; Botha, E.; Stivaktas, V.I.; Buss, P.; Beechler, B.R.; Myburgh, J.G.; Steyl, J.; Williams, J.; Venter, M. West Nile virus in wildlife and nonequine domestic animals, South Africa, 2010–2018. Emerg. Infect. Dis. 2019, 25, 2290–2294. [Google Scholar] [CrossRef] [Green Version]
- Steyn, J.; Motlou, P.; van Eeden, C.; Pretorius, M.; Stivaktas, V.I.; Williams, J.; Snyman, L.P.; Buss, P.E.; Beechler, B.; Jolles, A.; et al. Shuni virus in wildlife and nonequine domestic animals, South Africa. Emerg. Infect. Dis. 2020, 26, 1521–1525. [Google Scholar] [CrossRef]
- Steyn, J.; Fourie, I.; Steyl, J.; Williams, J.; Stivaktas, V.; Botha, E.; van Niekerk, S.; Reininghaus, B.; Venter, M. Zoonotic Alphaviruses in fatal and neurologic infections in wildlife and nonequine domestic animals, South Africa. Emerg. Infect. Dis. 2020, 26, 1182–1191. [Google Scholar] [CrossRef]
- Snyman, J.; Koekemoer, O.; Van Schalkwyk, A.; Jansen van Vuren, P.; Snyman, L.P.; Williams, J.; Venter, M. Epidemiology and genomic analysis of equine encephalosis Virus detected in horses with clinical signs in South Africa, 2010–2017. Viruses 2021, 13, 398. [Google Scholar] [CrossRef]
- Motlou, T.P.; Venter, M. Shuni Virus in Cases of Neurologic Disease in Humans, South Africa. Emerg. Infect. Dis. 2021, 27, 565–569. [Google Scholar] [CrossRef]
- Venter, G.J.; Labuschagne, K.; Hermanides, K.G.; Boikanyo, S.N.B.; Majatladi, D.M.; Morey, L. Comparison of the efficiency of five suction light traps under field conditions in South Africa for the collection of Culicoides species. Vet. Parasitol. 2009, 166, 299–307. [Google Scholar] [CrossRef]
- Labuschagne, K. The Culicoides Latreille (Diptera: Ceratopogonidae) Species of South Africa; University of Pretoria: Pretoria South, Africa, 2015. [Google Scholar]
- Van Niekerk, S.; Human, S.; Williams, J.; van Wilpe, E.; Pretorius, M.; Swanepoel, R.; Venter, M. Sindbis and Middelburg old world alphaviruses associated with neurologic disease in horses, South Africa. Emerg. Infect. Dis. 2015, 21, 2225–2229. [Google Scholar] [CrossRef]
- Zaayman, D.; Human, S.; Venter, M. A highly sensitive method for the detection and genotyping of West Nile virus by real-time PCR. J. Virol. Methods. 2009, 157, 155–160. [Google Scholar] [CrossRef]
- Lambert, A.J.; Lanciotti, R.S. Consensus amplification and novel multiplex sequencing method for S segment species identification of 47 viruses of the Orthobunyavirus, Phlebovirus, and Nairovirus genera of the family Bunyaviridae. J. Clin. Microbiol. 2009, 47, 2398. [Google Scholar] [CrossRef] [Green Version]
- Aznar-Lopez, C.; Vazquez-Moron, S.; Marston, D.A.; Juste, J.; Ibáñez, C.; Berciano, J.M.; Salsamendi, E.; Aihartza, J.; Banyard, A.; McElhinney, L.; et al. Detection of rhabdovirus viral RNA in oropharyngeal swabs and ectoparasites of Spanish bats. J. Gen. Virol. 2013, 94, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Bowen, M.D.; Trappier, S.G.; Sanchez, A.J.; Meyer, R.F.; Goldsmith, C.S.; Zaki, S.R.; Dunsterc, L.M.; Peters, C.J.; Ksiazeka, T.G.; Nichola, S.T.; et al. A reassortant bunyavirus isolated from acute hemorrhagic fever cases in Kenya and Somalia. Virology 2001, 291, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Segard, A.; Gardes, L.; Jacquier, E.; Grillet, C.; Mathieu, B.; Rakotoarivony, I.; Setier-Rio, M.-L.; Chavernac, D.; Cêtre-Sossah, C.; Balenghien, T.; et al. Schmallenberg virus in Culicoides Latreille (Diptera: Ceratopogonidae) populations in France during 2011–2012 outbreak. Transbound. Emerg. Dis. 2018, 65, e94–e103. [Google Scholar] [CrossRef]
- Steyn, J.; Venter, G.J.; Labuschagne, K.; Majatladi, D.; Boikanyo, S.N.B.; Lourens, C.; Ebersohn, K.; Venter, E.H. Possible over-wintering of bluetongue virus in Culicoides populations in the Onderstepoort area, Gauteng, South Africa. J. S. Afr. Vet. Assoc. 2016, 87, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Venter, G.J.; Labuschagne, K.; Majatladi, D.; Boikanyo, S.N.B.; Lourens, C.; Ebersohn, K.; Venter, E.H. Culicoides species abundance and potential over-wintering of African horse sickness virus in the Onderstepoort area, Gauteng, South Africa. J. S. Afr. Vet. Assoc. 2014, 85, 6. [Google Scholar] [CrossRef] [Green Version]
- Van Eeden, C.; Williams, J.H.; Gerdes, T.G.H.; van Wilpe, E.; Viljoen, A.; Swanepoel, R.; Venter, M. Shuni virus as cause of neurologic disease in horses. Emerg. Infect. Dis. 2012, 18, 318–321. [Google Scholar] [CrossRef]
- Hirashima, Y.; Kitahara, S.; Kato, T.; Shirafuji, H.; Tanaka, S.; Yanase, T. Congenital malformations of calves infected with Shamonda virus, Southern Japan. Emerg. Infect. Dis. 2017, 23, 993–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Causey, O.R.; Kemp, G.E.; Causey, C.E.; Lee, V.H. Isolations of Simbu group viruses in Ibadan, Nigeria 1964-69, including the new types of Sango, Shamonda, Sabo and Shuni. Ann. Trop. Med. Parasit. 1972, 66, 357–362. [Google Scholar] [CrossRef]
- Behar, A.; Izhaki, O.; Rot, A.; Benor, T.; Yankilevich, M.; Leszkowicz-Mazuz, M.; Brenner, J. Genomic Detection of Schmallenberg Virus, Israel. Emerg. Infect. Dis. 2021, 27, 2197–2200. [Google Scholar] [CrossRef] [PubMed]
- Molini, U.; Dondona, A.C.; Hilbert, R.; Monaco, F. Antibodies against Schmallenberg virus detected in cattle in the Otjozondjupa region, Namibia. J. S. Afr. Vet. Assoc. 2018, 89, 1666. [Google Scholar] [CrossRef] [PubMed]
- Blomström, A.L.; Stenberg, H.; Scharin, I.; Figueiredo, J.; Nhambirre, O.; Abilio, A.P.; Fafetine, J.; Berg, M. Serological screening suggests presence of Schmallenberg virus in cattle, sheep and goat in the Zambezia Province, Mozambique. Transbound. Emerg. Dis. 2014, 61, 289–292. [Google Scholar] [CrossRef] [Green Version]
- Kinney, R.M.; Calisher, C.H. Antigenic relationships among Simbu serogroup (Bunyaviridae) viruses. Am. J. Trop. Med. Hyg. 1981, 30, 1307–1318. [Google Scholar] [CrossRef]
- Mcintosh, B.M.; Mcgillivray, G.M.; Dlckinson, D.B. Ingwavuma virus: An arbovirus isolated in South Africa. S. Afr. Med. J. 1965, 30, 67–70. [Google Scholar]
- Top, F.H.; Kraivapan, C.; Grossman, R.A.; Rozmiarek, H.; Edelman, R.; Gould, D.J. Ingwavuma virus in Thailand. Am. J. Trop. Med. Hyg. 1974, 23, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Converse, J.D.; Tan, R.I.; Rachman, I.T.; Lee, V.H.; Shope, R.E. Ingwavuma Virus (Simbu Group) from Culex and Mansonia Mosquitoes (Diptera: Culicidae) in Indonesia. J. Med. Entomol. 1985, 22, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Ladner, J.T.; Savji, N.; Lofts, L.; Travassos da Rosa, A.; Wiley, M.R.; Gestole, M.C.; Rosen, G.E.; Guzman, H.; Vasconcelos, P.F.C.; Nunes, M.R.T.; et al. Genomic and phylogenetic characterization of viruses included in the Manzanilla and Oropouche species complexes of the genus Orthobunyavirus, family Bunyaviridae. J. Gen. Virol. 2014, 95 Pt 5, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, S.C.; Brault, A.C.; Kang, W.; Holland, J.J. Genetic and fitness changes accompanying adaptation of an arbovirus to vertebrate and invertebrate cells. J. Virol. 1991, 73, 4316. [Google Scholar] [CrossRef] [Green Version]
- Theodoridis, A.; Nevill, E.M.; Els, H.J.; Boshoff, S.T. Viruses isolated from Culicoides midges in South Africa during unsuccessful attempts to isolate bovine ephemeral fever virus. Onderstepoort J. Vet. Res. 1979, 46, 191–198. [Google Scholar]
- Barnard, B.J.H.; Paweska, J.T. Prevalence of antibodies against some equine viruses in zebra (Zebra burchellt) in the Kruger National Park, 1991–1992. Onderstepoort J. Vet. Res. 1993, 60, 175–179. [Google Scholar]
- Hang, J.; Klein, T.A.; Kim, H.C.; Yang, Y.; Jima, D.D.; Richardson, J.H.; Jarman, R.G. Genome sequences of five arboviruses in field-captured mosquitoes in a unique rural environment of South Korea. Genome Announc. 2016, 4, e01644-15. [Google Scholar] [CrossRef] [Green Version]
- Reuter, G.; Boros, Á.; Pál, J.; Kapusinszky, B.; Delwart, E.; Pankovics, P. Detection and genome analysis of a novel rhabdovirus (Riverside virus) from Ochlerotatus sp. mosquitoes in Central Europe. Infect Genet Evol. 2016, 39, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Cybinski, D.H.; Pgard, G. Isolation of a new Rhabdovirus in Australia related to Tibrogargan virus. Aust. J. Biol. Sci. 1986, 39, 225–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drolet, B.S.; Campbell, C.L.; Stuart, M.A.; Wilson, W.C. Vector competence of Culicoides sonorensis (Diptera: Ceratopogonidae) for vesicular stomatitis Virus. J. Med. Entomol. 2005, 42, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Hachiya, M.; Osborne, M.; Stinson, C.; Werner, B.G. Human eastern equine encephalitis in Massachusetts: Predictive indicators from mosquitoes collected at 10 long-term trap sites, 1979-2004. Am. J. Trop. Med. Hyg. 2007, 76, 285–292. [Google Scholar] [CrossRef]
- Bustamante, D.M.; Lord, C.C. Sources of error in the estimation of mosquito infection rates used to assess risk of arbovirus transmission. Am. J. Trop. Med. Hyg. 2010, 82, 1172–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musoke, J.; Hlokwe, T.; Marcotty, T.; Du Plessis, B.J.A.; Michel, A.L. Spillover of Mycobacterium bovis from wildlife to livestock, South Africa. Emerg. Infect. Dis. 2015, 21, 448. [Google Scholar] [CrossRef]
- Venter, G.J.; Nevill, E.M.; Van der Linde, T.C. Geographical distribution and relative abundance of stock-associated Culicoides species (Diptera: Ceratopogonidae) in southern Africa in relation to their potential as viral vectors. Onderstepoort J. Vet. Res. 1996, 63, 25–38. [Google Scholar]
- Steyn, J.; Venter, G.J.; Coetzee, P.; Venter, E.H. The epidemiology of bluetongue virus in Mnisi, South Africa. Am. J. Epidemiol. Inf. Dis. 2015, 3, 95–102. [Google Scholar] [CrossRef]
- Möhlmann, T.W.R.; Oymans, J.; Schreur, P.J.W.; Koenraadt, C.J.M.; Kortekaas, J.; Vogels, C.B.F. Vector competence of biting midges and mosquitoes for Shuni virus. PLoS Negl. Trop. Dis. 2018, 12, e0006993. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Location Code | Province | Animals Near Light Trap | Sampling Period | Trap Nights | No. of Culicoides Collected | Average Catch Size () |
|---|---|---|---|---|---|---|---|
| Hluvukani (Mnisi)/Hlalakahle/Ludlow (Mnisi) | MN | Mpumalanga | Humans, dogs, pigs, goats, cattle, donkeys | 2015/2016–2017 | 45 | 37,612 | 836 |
| Manyeleti/HHWRS (KNP) | MN/KNP | Wildlife, humans | 2015/2016–2017 | 25 | 14,519 | 580 | |
| Lapalala Wilderness | LAP | Limpopo | Wildlife, humans | November 2012–2017 | 142 | 40,555 | 285 |
| Marakele Natural Park | MAR | Wildlife, humans | September 2012–2017 | 157 | 89,087 | 567 | |
| Boschkop | GAU | Gauteng | Horse, human, goats and sheep | 2013–2017 | 88 | 107,863 | 1225 |
| Kyalami | KYA | Horse, humans, goats and sheep | 2013–2017 | 75 | 1,953,680 | 25,782 | |
| Total | 532 | 222,329 | 4179 |
| MIR 95% CI | ||||||||
|---|---|---|---|---|---|---|---|---|
| Site | No. Pools Tested | No. Pools Positive | All Positives | SHUV | Other Orthos | MIDV | EEV | Rhabdo |
| Mnisi | 47 | 10 | 0.4 (0.2–0.7) | 0.2 (0.0–0.3) | 0.1 (0.0–0.2) | - | 0.2 (0.0–0.3) | |
| KNP | 27 | 2 | 0.2 (0.0–0.4) | - | - | 0.1 (0.0–0.4) | - | |
| Boschkop | 64 | 9 | 0.3 (0.1–0.5) | 0.0 (0.0–0.1) | 0.2 (0.0–0.3) | - | 0.1 (0.0–0.2) | - |
| Kyalami | 59 | 9 | 0.3 (0.1–0.5) | 0.1 (0.0–0.3) | 0.1 (0.0–0.2) | - | 0.1 (0.0–0.2) | - |
| Lapalala | 49 | 20 | 0.8 (0.5–1.2) | 0.1 (0.0–0.3) | 0.0 (0.0–01) | - | 0.7 (0.3–1.0) | - |
| Marakele | 85 | 19 | 0.5 (0.2–0.6) | 0.1 (0.0–0.3) | 0.1 (0.0–0.2) | - | 0.2 (0.1–0.3) | - |
| Total | 331 | 69 | 0.4 (0.3–0.5) | 0.1 (0.1–0.2) | 0.1 (0.0–0.1) | 0.01 (0.0–0.03) | 0.2 (0.1–0.2) | 0.02 (0.0–0.05) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snyman, J.; Venter, G.J.; Venter, M. An Investigation of Culicoides (Diptera: Ceratopogonidae) as Potential Vectors of Medically and Veterinary Important Arboviruses in South Africa. Viruses 2021, 13, 1978. https://doi.org/10.3390/v13101978
Snyman J, Venter GJ, Venter M. An Investigation of Culicoides (Diptera: Ceratopogonidae) as Potential Vectors of Medically and Veterinary Important Arboviruses in South Africa. Viruses. 2021; 13(10):1978. https://doi.org/10.3390/v13101978
Chicago/Turabian StyleSnyman, Jumari, Gert J. Venter, and Marietjie Venter. 2021. "An Investigation of Culicoides (Diptera: Ceratopogonidae) as Potential Vectors of Medically and Veterinary Important Arboviruses in South Africa" Viruses 13, no. 10: 1978. https://doi.org/10.3390/v13101978