Herpes Simplex Virus Mistyping due to HSV-1 × HSV-2 Interspecies Recombination in Viral Gene Encoding Glycoprotein B

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Origin and Ethical Considerations
2.2. HSV Typing and Genomic Sequencing
2.3. Sequence Alignment and Recombination Detection
2.4. Data Availability
3. Results
3.1. Conflicting Assay Results Observed for CT_Sample9
3.2. Genome Consensus Sequence for CT_Sample9 Consistent with HSV-2
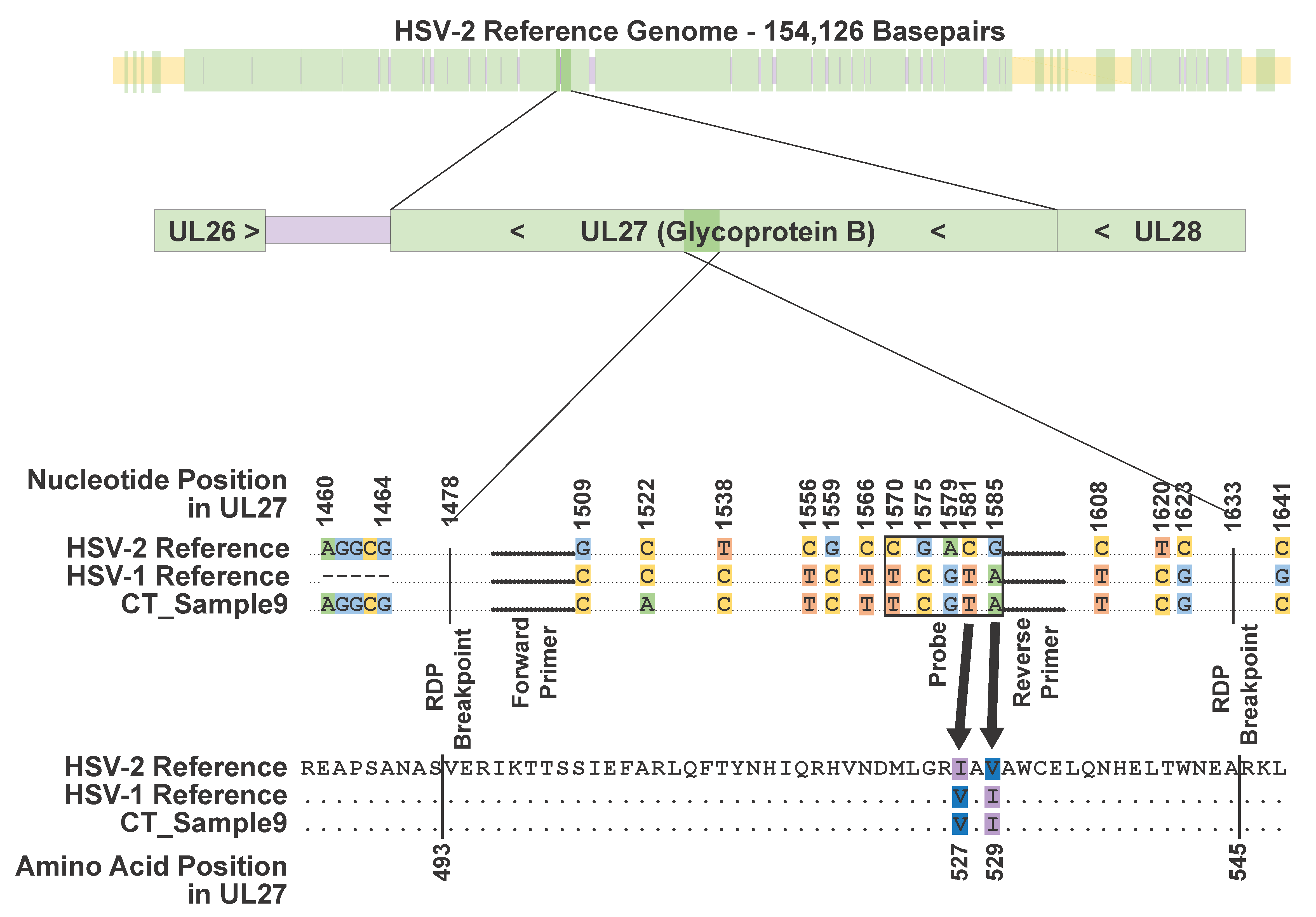
3.3. HSV-1 × HSV-2 Recombination Event Observed in UL27 in CT_Sample9
3.4. Single Nucleotide Variants Also Found in Primer and Probe Sequences for Typing Assay
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Looker, K.J.; Magaret, A.S.; May, M.T.; Turner, K.M.E.; Vickerman, P.; Gottlieb, S.L.; Newman, L.M. Global and Regional Estimates of Prevalent and Incident Herpes Simplex Virus Type 1 Infections in 2012. PLoS ONE 2015, 10, e0140765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Looker, K.J.; Magaret, A.S.; Turner, K.M.E.; Vickerman, P.; Gottlieb, S.L.; Newman, L.M. Global estimates of prevalent and incident herpes simplex virus type 2 infections in 2012. PLoS ONE 2015, 10, e114989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- 2015 STD Treatment Guidelines. Available online: https://www.cdc.gov/std/tg2015/default.htm (accessed on 13 August 2019).
- Miller, J.M.; Binnicker, M.J.; Campbell, S.; Carroll, K.C.; Chapin, K.C.; Gilligan, P.H.; Gonzalez, M.D.; Jerris, R.C.; Kehl, S.C.; Patel, R.; et al. A Guide to Utilization of the Microbiology Laboratory for Diagnosis of Infectious Diseases: 2018 Update by the Infectious Diseases Society of America and the American Society for Microbiologya. Clin. Infect. Dis. 2018, 67, e1–e94. [Google Scholar] [CrossRef] [PubMed]
- US Food and Drug Administration, Nucleic Acid Based Tests, List of Microbial Tests, Herpes Simplex Viruses 2020. Available online: https://www.fda.gov/medical-devices/vitro-diagnostics/nucleic-acid-based-tests (accessed on 7 July 2020).
- Kolb, A.W.; Larsen, I.V.; Cuellar, J.A.; Brandt, C.R. Genomic, phylogenetic, and recombinational characterization of herpes simplex virus 2 strains. J. Virol. 2015, 89, 6427–6434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umene, K.; Yoshida, M.; Fukumaki, Y. Genetic variability in the region encompassing reiteration VII of herpes simplex virus type 1, including deletions and multiplications related to recombination between direct repeats. Springerplus 2015, 4, 200. [Google Scholar] [CrossRef] [Green Version]
- Muylaert, I.; Elias, P. Contributions of nucleotide excision repair, DNA polymerase eta, and homologous recombination to replication of UV-irradiated herpes simplex virus type 1. J. Biol. Chem. 2010, 285, 13761–13768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowden, R.; Sakaoka, H.; Donnelly, P.; Ward, R. High recombination rate in herpes simplex virus type 1 natural populations suggests significant co-infection1. Infect. Genet. Evolut. 2004, 4, 115–123. [Google Scholar] [CrossRef]
- Cavanaugh, N.A.; Urban, M.; Beckman, J.; Spratt, T.E.; Kuchta, R.D. Identifying the features of purine dNTPs that allow accurate and efficient DNA replication by herpes simplex virus I DNA polymerase. Biochemistry 2009, 48, 3554–3564. [Google Scholar] [CrossRef] [Green Version]
- Norberg, P.; Bergström, T.; Rekabdar, E.; Lindh, M.; Liljeqvist, J.-A. Phylogenetic analysis of clinical herpes simplex virus type 1 isolates identified three genetic groups and recombinant viruses. J. Virol. 2004, 78, 10755–10764. [Google Scholar] [CrossRef] [Green Version]
- Szpara, M.L.; Gatherer, D.; Ochoa, A.; Greenbaum, B.; Dolan, A.; Bowden, R.J.; Enquist, L.W.; Legendre, M.; Davison, A.J. Evolution and diversity in human herpes simplex virus genomes. J. Virol. 2014, 88, 1209–1227. [Google Scholar] [CrossRef] [Green Version]
- Norberg, P.; Tyler, S.; Severini, A.; Whitley, R.; Liljeqvist, J.-Å.; Bergström, T. A genome-wide comparative evolutionary analysis of herpes simplex virus type 1 and varicella zoster virus. PLoS ONE 2011, 6, e22527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolb, A.W.; Lee, K.; Larsen, I.; Craven, M.; Brandt, C.R. Quantitative Trait Locus Based Virulence Determinant Mapping of the HSV-1 Genome in Murine Ocular Infection: Genes Involved in Viral Regulatory and Innate Immune Networks Contribute to Virulence. PLoS Pathog. 2016, 12, e1005499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blümel, J.; Gräper, S.; Matz, B. Structure of simian virus 40 DNA replicated by herpes simplex virus type 1. Virology 2000, 276, 445–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, D.E.; Weller, S.K. The role of DNA recombination in herpes simplex virus DNA replication. IUBMB Life 2003, 55, 451–458. [Google Scholar] [CrossRef]
- Weller, S.K.; Sawitzke, J.A. Recombination Promoted by DNA Viruses: Phage λ to Herpes Simplex Virus. Annu. Rev. Microbiol. 2014, 68, 237–258. [Google Scholar] [CrossRef] [Green Version]
- Nimonkar, A.V.; Boehmer, P.E. Reconstitution of recombination-dependent DNA synthesis in herpes simplex virus 1. Proc. Natl. Acad. Sci. USA 2003, 100, 10201–10206. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, A.J.; Mohni, K.N.; Kan, Y.; Hendrickson, E.A.; Stark, J.M.; Weller, S.K. The HSV-1 exonuclease, UL12, stimulates recombination by a single strand annealing mechanism. PLoS Pathog. 2012, 8, e1002862. [Google Scholar] [CrossRef]
- Porter, I.M.; Stow, N.D. Virus particles produced by the herpes simplex virus type 1 alkaline nuclease null mutant ambUL12 contain abnormal genomes. J. Gen. Virol. 2004, 85, 583–591. [Google Scholar] [CrossRef]
- Wertheim, J.O.; Smith, M.D.; Smith, D.M.; Scheffler, K.; Kosakovsky Pond, S.L. Evolutionary origins of human herpes simplex viruses 1 and 2. Mol. Biol. Evol. 2014, 31, 2356–2364. [Google Scholar] [CrossRef] [Green Version]
- Halliburton, I.W. Intertypic recombinants of herpes simplex viruses. J. Gen. Virol. 1980, 48, 1–23. [Google Scholar] [CrossRef]
- Morse, L.S.; Buchman, T.G.; Roizman, B.; Schaffer, P.A. Anatomy of herpes simplex virus DNA. IX. Apparent exclusion of some parental DNA arrangements in the generation of intertypic (HSV-1 X HSV-2) recombinants. J. Virol. 1977, 24, 231–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davison, A.J.; Wilkie, N.M. Inversion of the two segments of the herpes simplex virus genome in intertypic recombinants. J. Gen. Virol. 1983, 64 Pt 1, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, N.M.; Stow, N.D.; Marsden, H.S.; Preston, V.; Cortini, R.; Timbury, M.C.; Subak-Sharpe, J.H. Physical mapping of herpes simplex virus-coded functions and polypeptides by marker rescue and analysis of HSV-1/HSV-2 intertypic recombinants. IARC Sci Publ 1978, 24 Pt 1, 11–31. [Google Scholar]
- Casto, A.M.; Roychoudhury, P.; Xie, H.; Selke, S.; Perchetti, G.A.; Wofford, H.; Huang, M.-L.; Verjans, G.M.G.M.; Gottlieb, G.S.; Wald, A.; et al. Large, stable, contemporary interspecies recombination events in circulating human herpes simplex viruses. J. Infect. Dis. 2019. [Google Scholar] [CrossRef] [Green Version]
- Koelle, D.M.; Norberg, P.; Fitzgibbon, M.P.; Russell, R.M.; Greninger, A.L.; Huang, M.-L.; Stensland, L.; Jing, L.; Magaret, A.S.; Diem, K.; et al. Worldwide circulation of HSV-2 × HSV-1 recombinant strains. Sci. Rep. 2017, 7, 44084. [Google Scholar] [CrossRef] [Green Version]
- Burrel, S.; Boutolleau, D.; Ryu, D.; Agut, H.; Merkel, K.; Leendertz, F.H.; Calvignac-Spencer, S. Ancient Recombination Events between Human Herpes Simplex Viruses. Mol. Biol. Evol. 2017, 34, 1713–1721. [Google Scholar] [CrossRef] [Green Version]
- Corey, L.; Huang, M.-L.; Selke, S.; Wald, A. Differentiation of herpes simplex virus types 1 and 2 in clinical samples by a real-time taqman PCR assay. J. Med. Virol. 2005, 76, 350–355. [Google Scholar] [CrossRef]
- Greninger, A.L.; Roychoudhury, P.; Xie, H.; Casto, A.; Cent, A.; Pepper, G.; Koelle, D.M.; Huang, M.-L.; Wald, A.; Johnston, C.; et al. Ultrasensitive Capture of Human Herpes Simplex Virus Genomes Directly from Clinical Samples Reveals Extraordinarily Limited Evolution in Cell Culture. mSphere 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, R.M.; Lamers, S.L.; Weiner, B.; Ray, S.C.; Colgrove, R.C.; Diaz, F.; Jing, L.; Wang, K.; Saif, S.; Young, S.; et al. Genome Sequencing and Analysis of Geographically Diverse Clinical Isolates of Herpes Simplex Virus 2. J. Virol. 2015, 89, 8219–8232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamers, S.L.; Newman, R.M.; Laeyendecker, O.; Tobian, A.A.R.; Colgrove, R.C.; Ray, S.C.; Koelle, D.M.; Cohen, J.; Knipe, D.M.; Quinn, T.C. Global Diversity within and between Human Herpesvirus 1 and 2 Glycoproteins. J. Virol. 2015, 89, 8206–8218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, C.; Magaret, A.; Roychoudhury, P.; Greninger, A.L.; Cheng, A.; Diem, K.; Fitzgibbon, M.P.; Huang, M.-L.; Selke, S.; Lingappa, J.R.; et al. Highly conserved intragenic HSV-2 sequences: Results from next-generation sequencing of HSV-2 UL and US regions from genital swabs collected from 3 continents. Virology 2017, 510, 90–98. [Google Scholar] [CrossRef]
- Laing, K.J.; Magaret, A.S.; Mueller, D.E.; Zhao, L.; Johnston, C.; De Rosa, S.C.; Koelle, D.M.; Wald, A.; Corey, L. Diversity in CD8(+) T cell function and epitope breadth among persons with genital herpes. J. Clin. Immunol. 2010, 30, 703–722. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Test | Manufacturer | HSV-1 Target | HSV-2 Target | Site of Known HSV-1 x HSV-2 Recombination Event? | ||
|---|---|---|---|---|---|---|
| Gene | Protein Product | Gene | Protein Product | |||
| Aptima | Hologic, Inc. | UL42 | DNA Polymerase Processivity Subunit | UL42 | DNA Polymerase Processivity Subunit | No |
| Aries | Luminex Corporation | Not Available | Not Available | NA | ||
| IsoAmp | BioHelix Corporation | UL27 | Glycoprotein B | UL27 | Glycoprotein B | Yes |
| MultiCode-RTx | EraGen Biosciences, Inc. | UL27 | Glycoprotein B | UL27 | Glycoprotein B | Yes |
| BD ProbeTec | BD Diagnostic Systems | Not Available | Not Available | NA | ||
| Simplexa | Focus Diagnostics | UL30 | DNA Polymerase | UL30 | DNA Polymerase | Yes |
| Amiplivue | Quidel Corporation | Not Available | Not Available | NA | ||
| IMDX | Intelligent Medical | US6 | Glycoprotein D | UL30 | DNA Polymerase | Yes (UL30) |
| Solana | Quidel Corporation | US7 | Glycoprotein I | Intergenic between UL47 and UL48 | Intergenic between VP13/14 and VP16 | Yes (Intergenic between UL47 and UL48) |
| Artus | Qiagen | UL30, US6 | DNA Polymerase, Glycoprotein D | UL30, US6 | DNA Polymerase, Glycoprotein D | Yes (UL30) |
| SeeGene Anyplex | SeeGene | US6 | Glycoprotein D | US6 | Glycoprotein D | No |
| Quidel Molecular Direct | Diagnostic Hybrids | US4 | Glycoprotein G | US4 | Glycoprotein G | No |
| Sentosa | Vela Diagnostics | UL30 | DNA Polymerase | UL30 | DNA Polymerase | Yes |
| Elite MGB Ingenius | Elitechgroup | US6 | Glycoprotein D | US4 | Glycoprotein G | No |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casto, A.M.; Huang, M.-L.W.; Xie, H.; Jerome, K.R.; Wald, A.; Johnston, C.M.; Greninger, A.L. Herpes Simplex Virus Mistyping due to HSV-1 × HSV-2 Interspecies Recombination in Viral Gene Encoding Glycoprotein B. Viruses 2020, 12, 860. https://doi.org/10.3390/v12080860
Casto AM, Huang M-LW, Xie H, Jerome KR, Wald A, Johnston CM, Greninger AL. Herpes Simplex Virus Mistyping due to HSV-1 × HSV-2 Interspecies Recombination in Viral Gene Encoding Glycoprotein B. Viruses. 2020; 12(8):860. https://doi.org/10.3390/v12080860
Chicago/Turabian StyleCasto, Amanda M., Meei-Li W. Huang, Hong Xie, Keith R. Jerome, Anna Wald, Christine M. Johnston, and Alexander L. Greninger. 2020. "Herpes Simplex Virus Mistyping due to HSV-1 × HSV-2 Interspecies Recombination in Viral Gene Encoding Glycoprotein B" Viruses 12, no. 8: 860. https://doi.org/10.3390/v12080860