Long Term Delta-9-tetrahydrocannabinol Administration Inhibits Proinflammatory Responses in Minor Salivary Glands of Chronically Simian Immunodeficieny Virus Infected Rhesus Macaques



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care, Ethics and Experimental Procedures
2.2. Animal Model and Experimental Design
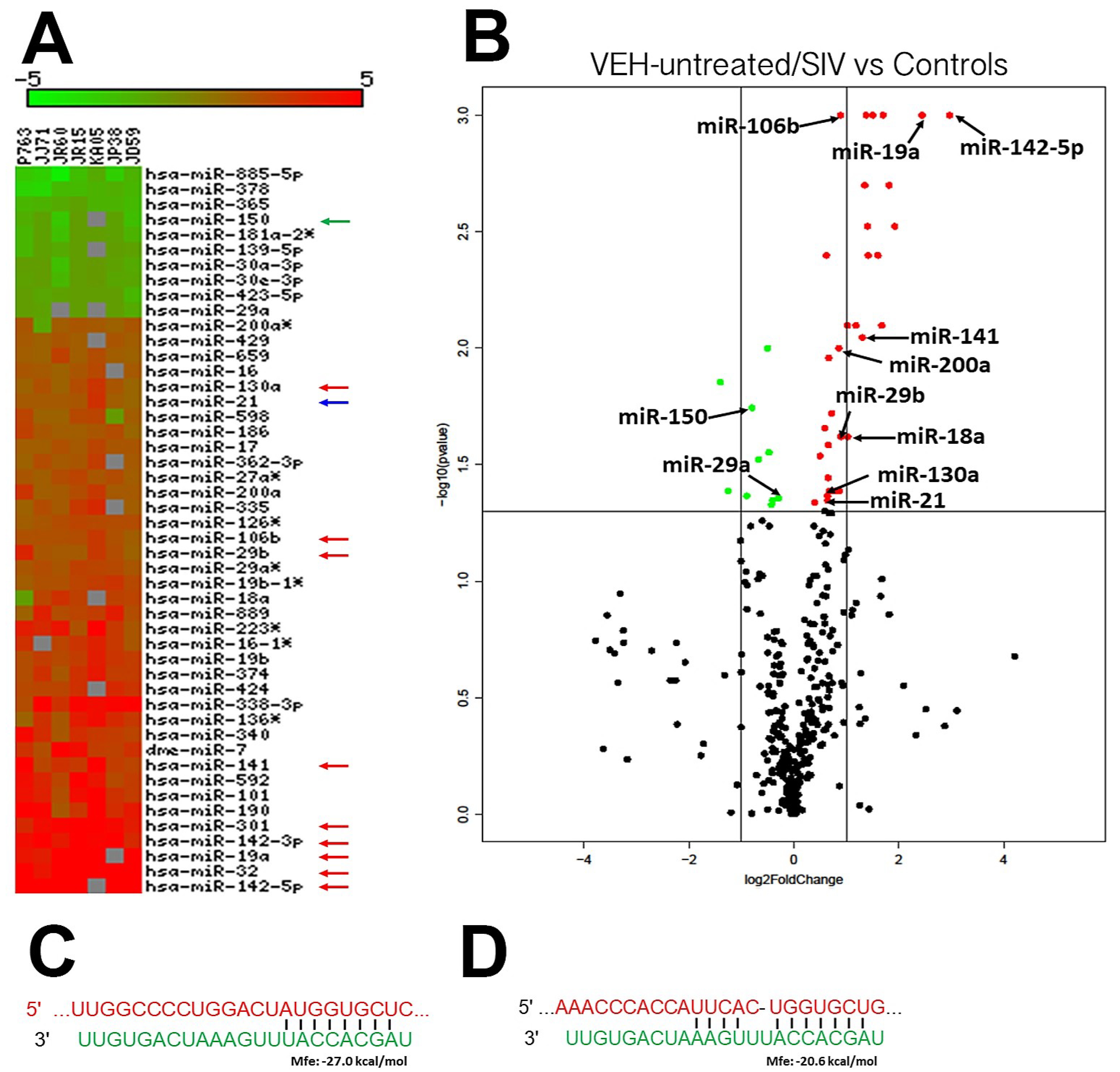
2.3. Global MicroRNA (miRNA) Profiling
2.4. RNA-Seq Library Construction, Clustering and Sequencing
2.5. Cloning of 3′-UTR of AGR2 and TSC22D3 mRNA and Dual-Glo Luciferase Reporter Gene Assay
2.6. Immunofluorescence for Cellular Localization of AGR2, WFDC2 and TSC22D3 in OPM Tissues
2.7. Quantitative Image Analysis of OPM Sections
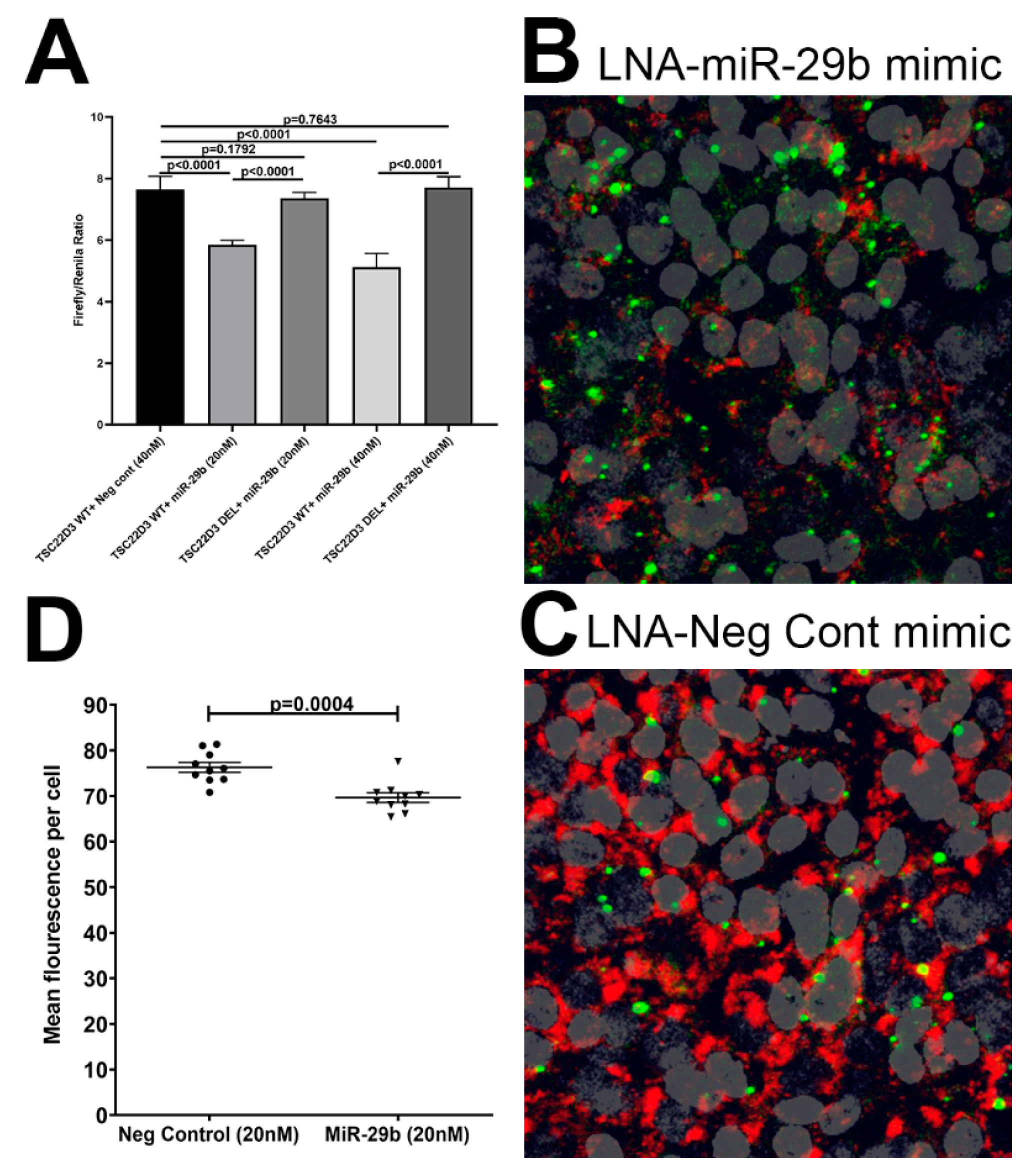
2.8. miR-29b Overexpression Studies
2.9. Quantitation of Mucosal Viral Loads
2.10. Data Analysis and Availability
3. Results
3.1. Plasma Viral Loads, CD4+ and CD8+ T Cell Status and Oral Histopathology
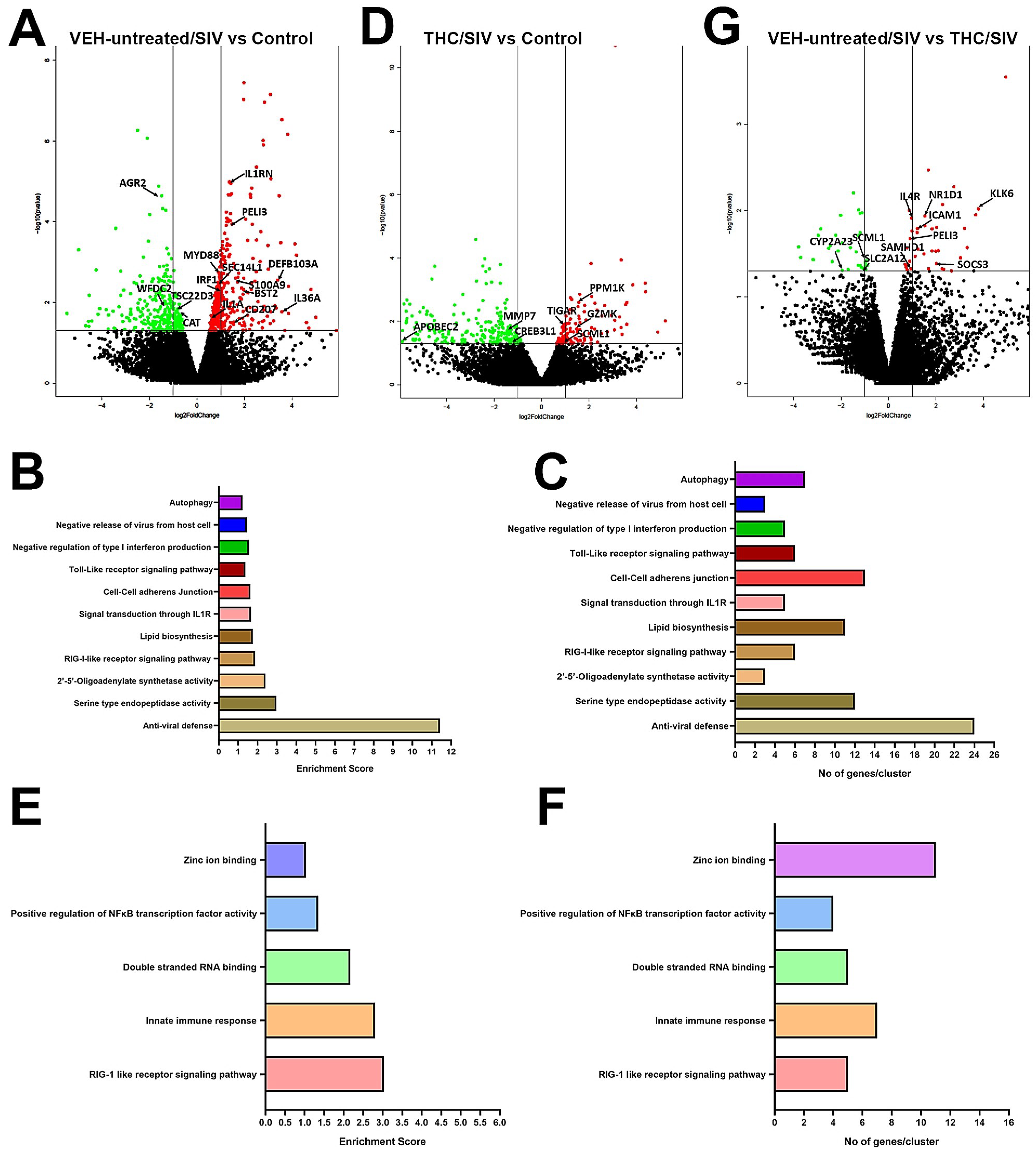
3.2. Genes Associated with Anti-Viral Defense, Interferon Signaling and Toll-Like Receptor Signaling Are Markedly Upregulated in OPM of VEH/SIV but not THC/SIV Rhesus Macaques
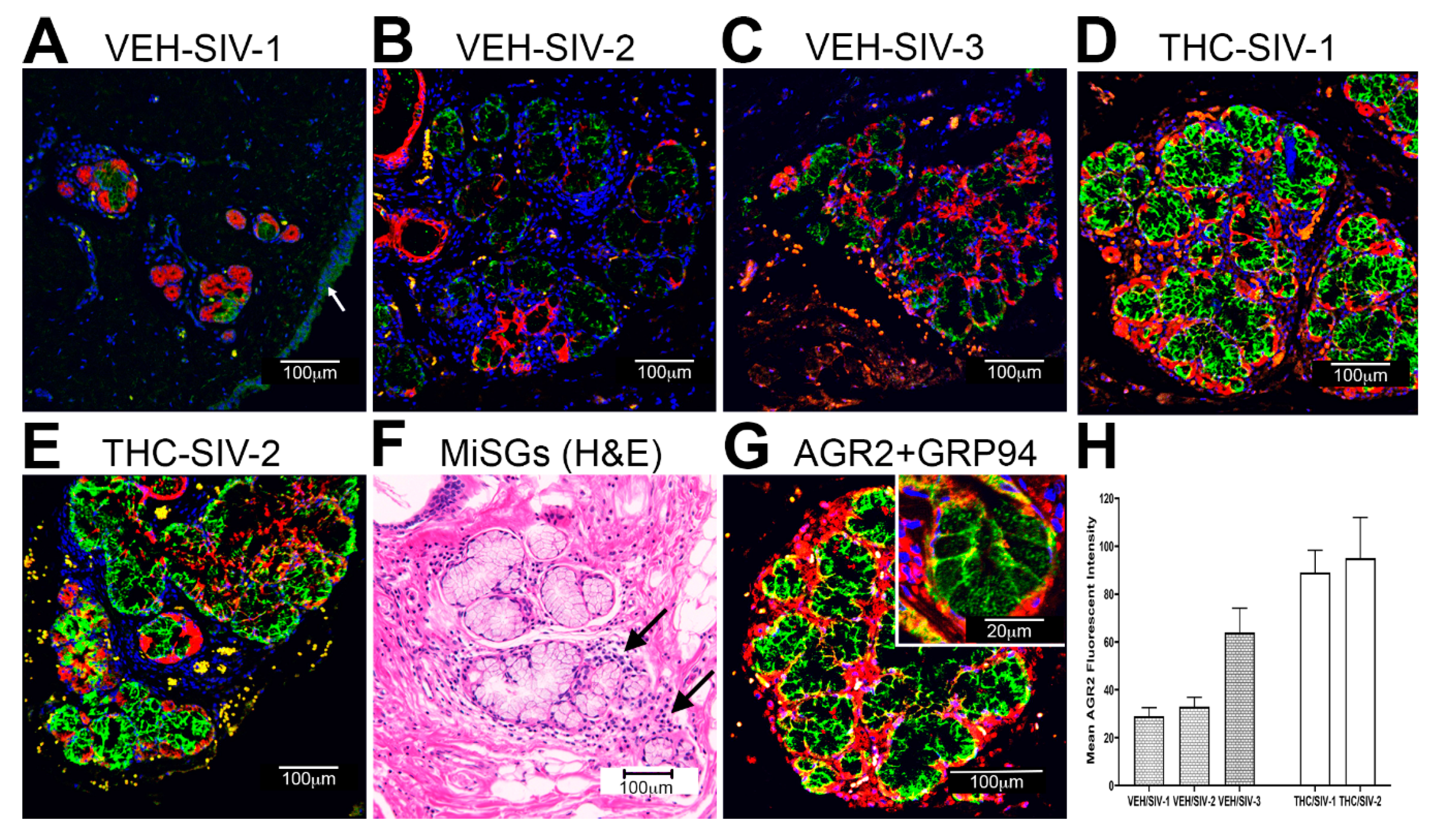
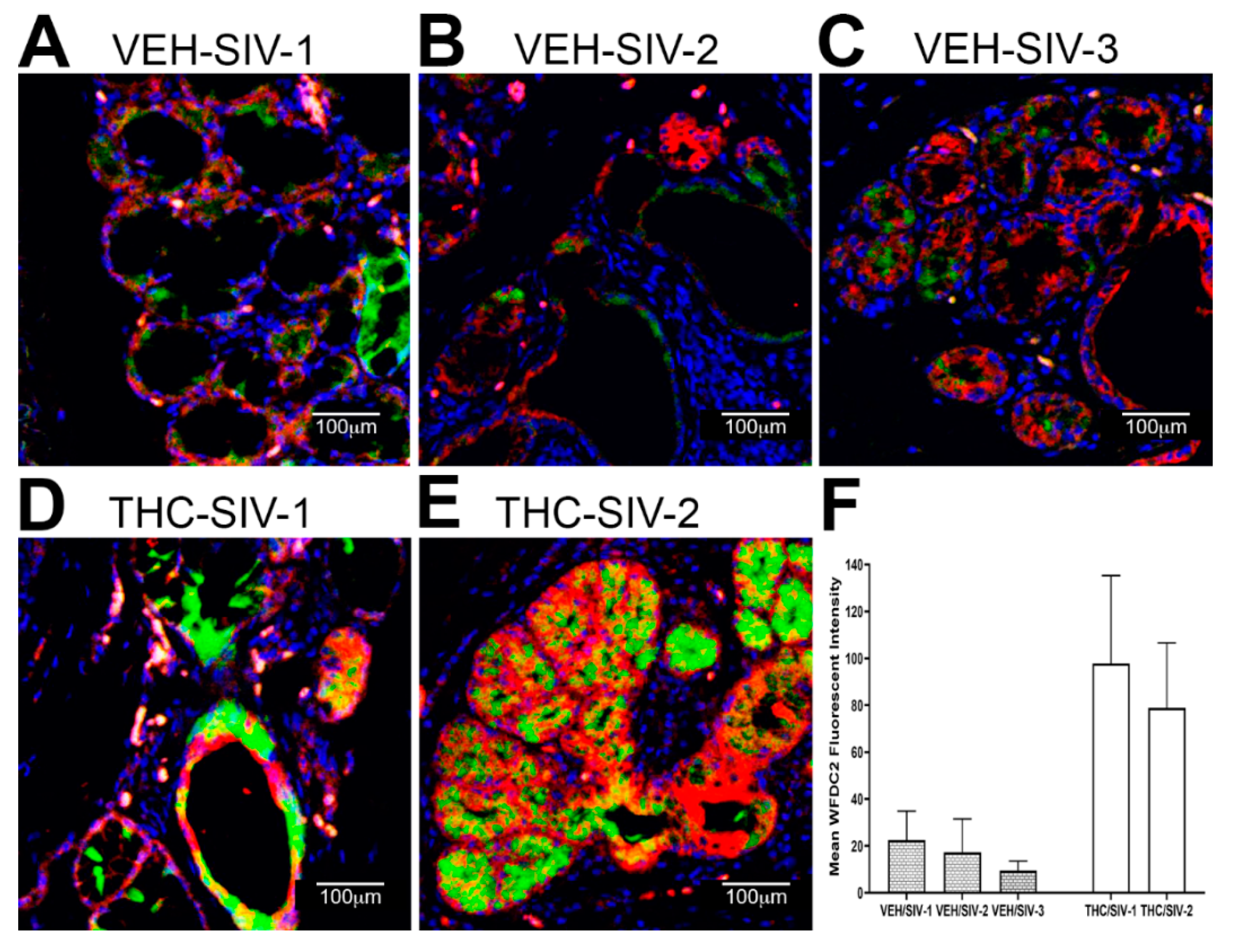
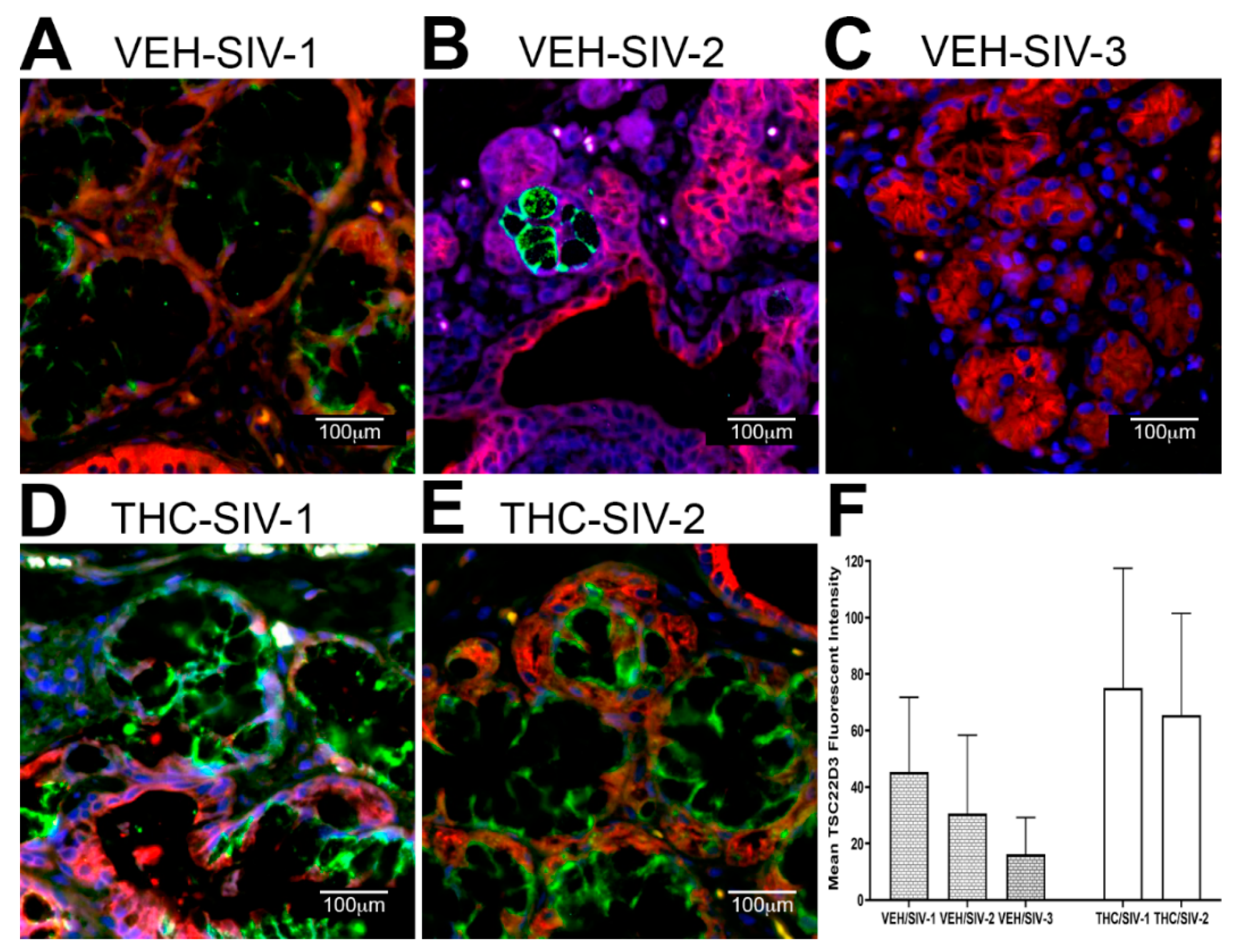
3.3. Endoplasmic Reticulum Stress (ER) regulator Anterior Gradient 2 (AGR2), Epithelial Barrier Enhancing WAP Four-Disulfide Core Domain Protein 2 (WFDC2) and the Anti-Inflammatory TSC22D3 Are Significantly Downregulated in OPM of VEH-Untreated/SIV Rhesus Macaques
3.4. Chronic THC Administration Preserved AGR2, WFDC2 and TSC22D3 Protein Expression in Minor Salivary Glands (MiSGs) of Chronic SIV-Infected Rhesus Macaques
3.5. TSC22D3 Is Post-Transcriptionally Regulated by miR-29b
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lu, F.; Jacobson, R.S. Oral mucosal immunity and HIV/SIV infection. J. Dent. Res. 2007, 86, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Nittayananta, W.; Tao, R.; Jiang, L.; Peng, Y.; Huang, Y. Oral innate immunity in HIV infection in HAART era. J. Oral Pathol. Med. 2015, 45, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentine, J.; Saladyanant, T.; Ramsey, K.; Blake, J.; Morelli, T.; Southerland, J.; Quinlivan, E.B.; Phillips, C.; Nelson, J.; Deparis, K.; et al. Impact of periodontal intervention on local inflammation, periodontitis, and HIV outcomes. Oral Dis. 2016, 22 (Suppl. S1) (Suppl. S1), 87–97. [Google Scholar] [CrossRef] [Green Version]
- Vernon, L.T.; Jayashantha, P.; Chidzonga, M.; Komesu, M.; Nair, R.G.; Johnson, N.W. Comorbidities associated with HIV and antiretroviral therapy (clinical sciences): A workshop report. Oral Dis. 2016, 22, 135–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heron, S.E.; Elahi, S. HIV Infection and Compromised Mucosal Immunity: Oral Manifestations and Systemic Inflammation. Front. Immunol. 2017, 8, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pólvora, T.L.S.; Nobre, Á.V.V.; Tirapelli, C.; Taba, M.; De Macedo, L.D.; Santana, R.C.; Pozzetto, B.; Lourenço, A.G.; Motta, A.C.F. Relationship between human immunodeficiency virus (HIV-1) infection and chronic periodontitis. Expert Rev. Clin. Immunol. 2018, 14, 315–327. [Google Scholar] [CrossRef]
- Annavajhala, M.K.; Khan, S.D.; Sullivan, S.B.; Shah, J.; Pass, L.; Kister, K.; Kunen, H.; Chiang, V.; Monnot, G.C.; Ricupero, C.L.; et al. Oral and Gut Microbial Diversity and Immune Regulation in Patients with HIV on Antiretroviral Therapy. mSphere 2020, 5. [Google Scholar] [CrossRef] [Green Version]
- George, M.; Verhoeven, D.; Sankaran, S.; Glavan, T.; Reay, E.; Dandekar, S. Heightened Cytotoxic Responses and Impaired Biogenesis Contribute to Early Pathogenesis in the Oral Mucosa of Simian Immunodeficiency Virus-Infected Rhesus Macaques. Clin. Vaccine Immunol. 2008, 16, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Saxena, D.; Chen, Z.; Liu, G.; Abrams, W.R.; Phelan, J.A.; Norman, R.G.; Fisch, G.S.; Corby, P.M.; Dewhirst, F.; et al. HIV Infection and Microbial Diversity in Saliva. J. Clin. Microbiol. 2014, 52, 1400–1411. [Google Scholar] [CrossRef] [Green Version]
- Lomeli-Martinez, S.M.; Valentin-Gomez, E.; Varela-Hernandez, J.J.; Alvarez-Zavala, M.; Sanchez-Reyes, K.; Ramos-Solano, M.; Cabrera-Silva, R.I.; Ramirez-Anguiano, V.M.; Lomeli-Martinez, M.A.; Martinez-Salazar, S.Y.; et al. Candida spp. Determination and Th1/Th2 Mixed Cytokine Profile in Oral Samples From HIV+ Patients With Chronic Periodontitis. Front. Immunol. 2019, 10, 1465. [Google Scholar] [CrossRef] [Green Version]
- Nittayananta, W.; Kemapunmanus, M.; Yangngam, S.; Talungchit, S.; Sriplung, H. Expression of oral secretory leukocyte protease inhibitor in HIV-infected subjects with long-term use of antiretroviral therapy. J. Oral Pathol. Med. 2012, 42, 208–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocon, S.; Murphy, C.; Dang, A.T.; Sankaran-Walters, S.; Li, C.-S.; Tarara, R.; Borujerdpur, N.; Dandekar, S.; Paster, B.J.; George, M. Transcription Profiling Reveals Potential Mechanisms of Dysbiosis in the Oral Microbiome of Rhesus Macaques with Chronic Untreated SIV Infection. PLoS ONE 2013, 8, e80863. [Google Scholar] [CrossRef]
- Nittayananta, W.; Amornthatree, K.; Kemapunmanus, M.; Talungchit, S.; Sriplung, H. Expression of oral cytokines in HIV-infected subjects with long-term use of antiretroviral therapy. Oral Dis. 2013, 20, e57–e64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patton, L.L.; Ramirez-Amador, V.; Anaya-Saavedra, G.; Nittayananta, W.; Carrozzo, M.; Ranganathan, K. Urban legends series: Oral manifestations of HIV infection. Oral Dis. 2013, 19, 533–550. [Google Scholar] [CrossRef] [PubMed]
- Emfietzoglou, R.; Pachymanolis, E.; Piperi, C. Impact of Epigenetic alterations in the development of oral diseases. Curr. Med. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, H.; Suzuki, H.I. Systems and Synthetic microRNA Biology: From Biogenesis to Disease Pathogenesis. Int. J. Mol. Sci. 2019, 21, 132. [Google Scholar] [CrossRef] [Green Version]
- Cha, S.; Mona, M.; Lee, K.-E.; Kim, D.H.; Han, K. MicroRNAs in Autoimmune Sjögren’s Syndrome. Genom. Inform. 2018, 16, e19. [Google Scholar] [CrossRef]
- Kulkarni, V.; Uttamani, J.R.; Naqvi, A.R.; Nares, S. microRNAs: Emerging players in oral cancers and inflammatory disorders. Tumor Biol. 2017, 39, 1010428317698379. [Google Scholar] [CrossRef] [Green Version]
- Luan, X.; Zhou, X.; Naqvi, A.; Francis, M.; Foyle, D.; Nares, S.; Diekwisch, T.G. MicroRNAs and immunity in periodontal health and disease. Int. J. Oral Sci. 2018, 10, 24. [Google Scholar] [CrossRef]
- Sannigrahi, M.; Sharma, R.; Panda, N.K.; Khullar, M. Role of non-coding RNAs in head and neck squamous cell carcinoma: A narrative review. Oral Dis. 2017, 24, 1417–1427. [Google Scholar] [CrossRef]
- Chen, S.C.Y.; Constantinides, C.; Kebschull, M.; Papapanou, P.N. MicroRNAs Regulate Cytokine Responses in Gingival Epithelial Cells. Infect. Immun. 2016, 84, 3282–3289. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Xue, N.; Xie, Y.-F.; Zhu, N.-W.; Dong, Y.-Y.; Wei, C.-C.; Deng, J.-Y. The negative feedback regulation of microRNA-146a in human periodontal ligament cells after Porphyromonas gingivalis lipopolysaccharide stimulation. Inflamm. Res. 2015, 64, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Ouhara, K.; Savitri, I.J.; Fujita, T.; Kittaka, M.; Kajiya, M.; Iwata, T.; Miyagawa, T.; Yamakawa, M.; Shiba, H.; Kurihara, H. miR-584 Expressed in Human Gingival Epithelial Cells Is Induced by Porphyromonas gingivalis Stimulation and Regulates Interleukin-8 Production via Lactoferrin Receptor. J. Periodontol. 2014, 85, e198–e204. [Google Scholar] [CrossRef]
- Chandra, L.C.; Kumar, V.; Torben, W.; Vande Stouwe, C.; Winsauer, P.; Amedee, A.; Molina, P.E.; Mohan, M. Chronic administration of Delta9-tetrahydrocannabinol induces intestinal anti-inflammatory microRNA expression during acute simian immunodeficiency virus infection of rhesus macaques. J. Virol. 2015, 89, 1168–1181. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Torben, W.; Mansfield, J.; Alvarez, X.; Stouwe, C.V.; Li, J.; Byrareddy, S.N.; Didier, P.J.; Pahar, B.; Molina, P.E.; et al. Cannabinoid Attenuation of Intestinal Inflammation in Chronic SIV-Infected Rhesus Macaques Involves T Cell Modulation and Differential Expression of Micro-RNAs and Pro-inflammatory Genes. Front. Immunol. 2019, 10, 914. [Google Scholar] [CrossRef] [PubMed]
- Molina, P.E.; Amedee, A.M.; LeCapitaine, N.J.; Zabaleta, J.; Mohan, M.; Winsauer, P.J.; Vande Stouwe, C.; McGoey, R.R.; Auten, M.W.; LaMotte, L.; et al. Modulation of gut-specific mechanisms by chronic delta(9)-tetrahydrocannabinol administration in male rhesus macaques infected with simian immunodeficiency virus: A systems biology analysis. AIDS Res. Hum. Retrovir. 2014, 30, 567–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusthen, S.; Kristoffersen, A.K.; Young, A.; Galtung, H.K.; Petrovski, B.É.; Palm, Ø.; Enersen, M.; Jensen, J.L. Dysbiotic salivary microbiota in dry mouth and primary Sjögren’s syndrome patients. PLoS ONE 2019, 14, e0218319. [Google Scholar] [CrossRef] [PubMed]
- Tsigalou, C.; Stavropoulou, E.; Bezirtzoglou, E. Current Insights in Microbiome Shifts in Sjogren’s Syndrome and Possible Therapeutic Interventions. Front. Immunol. 2018, 9, 1106. [Google Scholar] [CrossRef]
- Van Der Meulen, T.; Harmsen, H.J.M.; Bootsma, H.; Liefers, S.C.; Vila, A.V.; Zhernakova, A.; Fu, J.; Wijmenga, C.; Spijkervet, F.K.L.; Kroese, F.G.M.; et al. Dysbiosis of the buccal mucosa microbiome in primary Sjögren’s syndrome patients. Rheumatology 2018, 57, 2225–2234. [Google Scholar] [CrossRef]
- Molina, P.E.; Winsauer, P.; Zhang, P.; Walker, E.; Birke, L.; Amedee, A.; Stouwe, C.V.; Troxclair, D.; McGoey, R.; Varner, K.; et al. Cannabinoid Administration Attenuates the Progression of Simian Immunodeficiency Virus. AIDS Res. Hum. Retrovir. 2011, 27, 585–592. [Google Scholar] [CrossRef] [Green Version]
- Winsauer, P.; Molina, P.E.; Amedee, A.M.; Filipeanu, C.M.; McGoey, R.R.; Troxclair, D.A.; Walker, E.M.; Birke, L.L.; Stouwe, C.V.; Howard, J.M.; et al. Tolerance to chronic delta-9-tetrahydrocannabinol (Δ⁹-THC) in rhesus macaques infected with simian immunodeficiency virus. Exp. Clin. Psychopharmacol. 2011, 19, 154–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Q.; Liu, L.; Cong, Z.; Wu, X.; Wang, H.; Qin, C.; Molina, P.; Chen, Z. Chronic Δ9-Tetrahydrocannabinol Administration Reduces IgE+B Cells but Unlikely Enhances Pathogenic SIVmac251 Infection in Male Rhesus Macaques of Chinese Origin. J. Neuroimmune Pharmacol. 2016, 11, 584–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Mansfield, J.; Fan, R.; MacLean, A.; Li, J.; Mohan, M. miR-130a and miR-212 Disrupt the Intestinal Epithelial Barrier through Modulation of PPARgamma and Occludin Expression in Chronic Simian Immunodeficiency Virus-Infected Rhesus Macaques. J. Immunol. 2018, 200, 2677–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Torben, W.; Kenway, C.S.; Schiro, F.R.; Mohan, M. Longitudinal Examination of the Intestinal Lamina Propria Cellular Compartment of Simian Immunodeficiency Virus-Infected Rhesus Macaques Provides Broader and Deeper Insights into the Link between Aberrant MicroRNA Expression and Persistent Immune Activation. J. Virol. 2016, 90, 5003–5019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiernasz, N.; Leroi, F.; Chevalier, F.; Cornet, J.; Cardinal, M.; Rohloff, J.; Passerini, D.; Skırnisdóttir, S.; Pilet, M.-F. Salmon Gravlax Biopreservation With Lactic Acid Bacteria: A Polyphasic Approach to Assessing the Impact on Organoleptic Properties, Microbial Ecosystem and Volatilome Composition. Front. Microbiol. 2020, 10, 3103. [Google Scholar] [CrossRef] [PubMed]
- Aboulnasr, F.; Paranjape, G.; Badley, A.D. The TRAIL: TRAILshort Axis in HIV Immunopathology. Crit. Rev. Immunol. 2018, 38, 491–503. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2014, 31, 166–169. [Google Scholar] [CrossRef]
- Anders, S.; McCarthy, D.J.; Chen, Y.; Okoniewski, M.; Smyth, G.K.; Huber, W.; Robinson, M.D. Count-based differential expression analysis of RNA sequencing data using R and Bioconductor. Nat. Protoc. 2013, 8, 1765–1786. [Google Scholar] [CrossRef]
- Ebersole, J.L.; Peyyala, R.; Gonzalez, O.A. Biofilm-induced profiles of immune response gene expression by oral epithelial cells. Mol. Oral Microbiol. 2019, 34. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.B.; Xiong, Y.; Pattabiraman, G.; Manavalan, T.T.; Qiu, F.; Medvedev, A.E. Pellino-3 promotes endotoxin tolerance and acts as a negative regulator of TLR2 and TLR4 signaling. J. Leukoc. Biol. 2015, 98, 963–974. [Google Scholar] [CrossRef] [Green Version]
- Janaka, S.K.; Tavakoli-Tameh, A.; Neidermyer, W.J., Jr.; Serra-Moreno, R.; Hoxie, J.A.; Desrosiers, R.C.; Johnson, R.P.; Lifson, J.D.; Wolinsky, S.M.; Evans, D.T. Polymorphisms in Rhesus Macaque Tetherin Are Associated with Differences in Acute Viremia in Simian Immunodeficiency Virus Deltanef-Infected Animals. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, N.; Schenten, V.; Bueb, J.L.; Tolle, F.; Brechard, S. miRNAs Regulate Cytokine Secretion Induced by Phosphorylated S100A8/A9 in Neutrophils. Int. J. Mol. Sci. 2019, 20, 5699. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.-C.; Xu, W.-D.; Liu, X.-Y.; Liu, X.-Y.; Huang, A.-F.; Su, L.; Liu, X.-Y. Biology of IL-36 Signaling and Its Role in Systemic Inflammatory Diseases. Front. Immunol. 2019, 10, 2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aqrawi, L.A.; Jensen, J.L.; Øijordsbakken, G.; Ruus, A.-K.; Nygård, S.; Holden, M.; Jonsson, R.; Galtung, H.K.; Skarstein, K. Signalling pathways identified in salivary glands from primary Sjögren’s syndrome patients reveal enhanced adipose tissue development. Autoimmunity 2018, 51, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Sparber, F.; Dolowschiak, T.; Mertens, S.; Lauener, L.; Clausen, B.E.; Joller, N.; Stoitzner, P.; Tussiwand, R.; LeibundGut-Landmann, S. Langerin+ DCs regulate innate IL-17 production in the oral mucosa during Candida albicans-mediated infection. PLoS Pathog. 2018, 14, e1007069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiripolsky, J.; Romano, R.A.; Kasperek, E.M.; Yu, G.; Kramer, J.M. Activation of Myd88-Dependent TLRs Mediates Local and Systemic Inflammation in a Mouse Model of Primary Sjogren’s Syndrome. Front. Immunol. 2019, 10, 2963. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, A.; Liu, J.; Martinot, A.J.; Mcmahan, K.; Mercado, N.B.; Peter, L.; Tostanoski, L.H.; Yu, J.; Maliga, Z.; Nekorchuk, M.; et al. SARS-CoV-2 infection protects against rechallenge in rhesus macaques. Science 2020, 4776. [Google Scholar] [CrossRef]
- Han, Q.; Bradley, T.; Williams, W.B.; Cain, D.W.; Montefiori, D.C.; Saunders, K.O.; Parks, R.J.; Edwards, R.W.; Ferrari, G.; Mueller, O.; et al. Neonatal Rhesus Macaques Have Distinct Immune Cell Transcriptional Profiles following HIV Envelope Immunization. Cell Rep. 2020, 30, 1553–1569.e6. [Google Scholar] [CrossRef]
- Yu, P.; Qi, F.; Xu, Y.; Li, F.; Liu, P.; Liu, J.; Bao, L.; Deng, W.; Gao, H.; Xiang, Z.; et al. Age-related rhesus macaque models of COVID-19. Anim. Model. Exp. Med. 2020, 3, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Lynn, T.M.; Molloy, E.L.; Masterson, J.C.; Glynn, S.F.; Costello, R.W.; Avdalovic, M.V.; Schelegle, E.S.; Miller, L.A.; Hyde, D.M.; O’Dea, S. SMAD Signaling in the Airways of Healthy Rhesus Macaques versus Rhesus Macaques with Asthma Highlights a Relationship Between Inflammation and Bone Morphogenetic Proteins. Am. J. Respir. Cell Mol. Biol. 2016, 54, 562–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamperschroer, C.; Goldstein, R.; Schneider, P.A.; Kuang, B.; Eisenbraun, M.D. Utilization of lipopolysaccharide challenge in cynomolgus macaques to assess IL-10 receptor antagonism. J. Immunotoxicol. 2019, 16, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Guo, M.; Rao, Y.; Wang, Y.; Xian, Q.; Yu, Q.; Huang, Z.; Wang, X.; Bao, R.; Yue, J.; et al. Mycobacterium tuberculosis Erdman infection of cynomolgus macaques of Chinese origin. J. Thorac. Dis. 2018, 10, 3609–3621. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Ren, S.; Korge, P.; Choi, J.; Dong, Y.; Weiss, J.; Koehler, C.; Chen, J.-N.; Wang, Y. A novel mitochondrial matrix serine/threonine protein phosphatase regulates the mitochondria permeability transition pore and is essential for cellular survival and development. Genome Res. 2007, 21, 784–796. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhang, D.-M.; Feng, X.; Wang, J.; Qin, Y.-Y.; Zhang, T.; Huang, Q.; Sheng, R.; Chen, Z.; Li, M.; et al. TIGAR inhibits ischemia/reperfusion-induced inflammatory response of astrocytes. Neuropharmacology 2018, 131, 377–388. [Google Scholar] [CrossRef]
- Guo, J.; Jin, D. A genetic screen in Drosophila implicates Sex comb on midleg (Scm) in tissue overgrowth and mechanisms of Scm degradation by Wds. Mech. Dev. 2015, 136, 1–7. [Google Scholar] [CrossRef]
- Herich, S.; Schneider-Hohendorf, T.; Rohlmann, A.; Ghadiri, M.K.; Schulte-Mecklenbeck, A.; Zondler, L.; Janoschka, C.; Ostkamp, P.; Richter, J.; Breuer, J.; et al. Human CCR5high effector memory cells perform CNS parenchymal immune surveillance via GZMK-mediated transendothelial diapedesis. Brain 2019, 142, 3411–3427. [Google Scholar] [CrossRef]
- Maurel, M.; Obacz, J.; Avril, T.; Ding, Y.; Papadodima, O.; Treton, X.; Daniel, F.; Pilalis, E.; Hörberg, J.; Hou, W.; et al. Control of anterior GR adient 2 (AGR 2) dimerization links endoplasmic reticulum proteostasis to inflammation. EMBO Mol. Med. 2019, 11, e10120. [Google Scholar] [CrossRef]
- Park, S.-W.; Zhen, G.; Verhaeghe, C.; Nakagami, Y.; Nguyenvu, L.T.; Barczak, A.J.; Killeen, N.; Erle, D. The protein disulfide isomerase AGR2 is essential for production of intestinal mucus. Proc. Natl. Acad. Sci. USA 2009, 106, 6950–6955. [Google Scholar] [CrossRef] [Green Version]
- Bingle, L.; Cross, S.; High, A.S.; Wallace, W.A.H.; Rassl, D.; Yuan, G.; Hellstrom, I.; Campos, M.; Bingle, C.D. WFDC2 (HE4): A potential role in the innate immunity of the oral cavity and respiratory tract and the development of adenocarcinomas of the lung. Respir. Res. 2006, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Parikh, K.; Antanaviciute, A.; Fawkner-Corbett, D.; Jagielowicz, M.; Aulicino, A.; Lagerholm, C.; Davis, S.; Kinchen, J.; Chen, H.H.; Alham, N.K.; et al. Colonic epithelial cell diversity in health and inflammatory bowel disease. Nature 2019, 567, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Hahn, R.T.; Hoppstädter, J.; Hirschfelder, K.; Hachenthal, N.; Diesel, B.; Kessler, S.M.; Huwer, H.; Kiemer, A.K. Downregulation of the glucocorticoid-induced leucine zipper (GILZ) promotes vascular inflammation. Atherosclerosis 2014, 234, 391–400. [Google Scholar] [CrossRef]
- Ricci, E.; Ronchetti, S.; Gabrielli, E.; Pericolini, E.; Gentili, M.; Roselletti, E.; Vecchiarelli, A.; Riccardi, C.; Ricci, E. GILZ restrains neutrophil activation by inhibiting the MAPK pathway. J. Leukoc. Biol. 2018, 105, 187–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronchetti, S.; Gentili, M.; Ricci, E.; Migliorati, G.; Riccardi, C. Glucocorticoid-Induced Leucine Zipper as a Druggable Target in Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Toczewska, J.; Konopka, T. Activity of enzymatic antioxidants in periodontitis: A systematic overview of the literature. Dent. Med. Probl. 2019, 56, 419–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzeldemir-Akcakanat, E.; Alkan, B.; Sunnetci-Akkoyunlu, D.; Gurel, B.; Balta, V.M.; Kan, B.; Akgun, E.; Yilmaz, E.B.; Baykal, A.T.; Çine, N.; et al. Molecular signatures of chronic periodontitis in gingiva: A genomic and proteomic analysis. J. Periodontol. 2019, 90, 663–673. [Google Scholar] [CrossRef]
- Denard, B.; Seemann, J.; Chen, Q.; Gay, A.; Huang, H.; Chen, Y.; Ye, J. The Membrane-Bound Transcription Factor CREB3L1 Is Activated in Response to Virus Infection to Inhibit Proliferation of Virus-Infected Cells. Cell Host Microbe 2011, 10, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, T.; Marusawa, H.; Endo, Y.; Ueda, Y.; Matsumoto, Y.; Chiba, T. Expression of APOBEC2 is transcriptionally regulated by NF-κB in human hepatocytes. FEBS Lett. 2006, 580, 731–735. [Google Scholar] [CrossRef] [Green Version]
- Sze, A.; Olagnier, D.; Lin, R.; Van Grevenynghe, J.; Hiscott, J. SAMHD1 Host Restriction Factor: A Link with Innate Immune Sensing of Retrovirus Infection. J. Mol. Biol. 2013, 425, 4981–4994. [Google Scholar] [CrossRef]
- Wang, S.; Lin, Y.; Yuan, X.; Li, F.; Guo, L.; Wu, B. REV-ERBalpha integrates colon clock with experimental colitis through regulation of NF-kappaB/NLRP3 axis. Nat. Commun. 2018, 9, 4246. [Google Scholar] [CrossRef] [Green Version]
- Wichnieski, C.; Maheshwari, K.; De Souza, L.C.; Nieves, F.; Tartari, T.; Garlet, G.P.; Carneiro, E.; Letra, A.; Silva, R.M. DNA methylation profiles of immune response-related genes in apical periodontitis. Int. Endod. J. 2018, 52, 5–12. [Google Scholar] [CrossRef]
- Bayani, J.; Diamandis, E. The physiology and pathobiology of human kallikrein-related peptidase 6 (KLK6). Clin. Chem. Lab. Med. 2012, 50, 211–233. [Google Scholar] [CrossRef] [PubMed]
- Seyedmajidi, M.; Shafaee, S.; Bijani, A.; Bagheri, S. VCAM1 and ICAM1 expression in oral lichen planus. Int. J. Mol. Cell. Med. 2013, 2, 34–40. [Google Scholar] [PubMed]
- Papathanasiou, E.; Kantarci, A.; Konstantinidis, A.; Gao, H.; Van Dyke, T.E. SOCS-3 Regulates Alveolar Bone Loss in Experimental Periodontitis. J. Dent. Res. 2016, 95, 1018–1025. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Su, L.; Duan, X.; Chen, X.; Hays, A.; Upadhyayula, S.; Shivde, J.; Wang, H.; Li, Y.; Huang, D.; et al. MicroRNA-21 down-regulates inflammation and inhibits periodontitis. Mol. Immunol. 2018, 101, 608–614. [Google Scholar] [CrossRef]
- Kim, C.; Hu, B.; Jadhav, R.R.; Jin, J.; Zhang, H.; Cavanagh, M.M.; Akondy, R.S.; Ahmed, R.; Weyand, C.M.; Goronzy, J.J. Activation of miR-21-Regulated Pathways in Immune Aging Selects against Signatures Characteristic of Memory T Cells. Cell Rep. 2018, 25, 2148–2162.e5. [Google Scholar] [CrossRef] [Green Version]
- Pentenero, M.; Bowers, L.; Jayasinghe, R.; Cheong, S.C.; Farah, C.S.; Kerr, A.R.; Alevizos, I. World Workshop on Oral Medicine VII: Functional pathways involving differentially expressed lncRNAs in oral squamous cell carcinoma. Oral Dis. 2019, 25, 79–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, B. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- De Campos Parra Flores, B.D.C.T.; Lourenco, S.V.; Damascena, A.S.; Kowaslki, L.P.; Soares, F.A.; Coutinho-Camillo, C.M. Altered expression of apoptosis-regulating miRNAs in salivary gland tumors suggests their involvement in salivary gland tumorigenesis. Virchows Arch. 2016, 470, 291–299. [Google Scholar] [CrossRef]
- Bereshchenko, O.; Migliorati, G.; Bruscoli, S.; Riccardi, C. Glucocorticoid-Induced Leucine Zipper: A Novel Anti-inflammatory Molecule. Front. Pharmacol. 2019, 10, 308. [Google Scholar] [CrossRef] [Green Version]
- Ballegeer, M.; Vandewalle, J.; Eggermont, M.; Van Isterdael, G.; Dejager, L.; De Bus, L.; Decruyenaere, J.; Vandenbroucke, R.E.; Libert, C. Overexpression of Gilz Protects Mice Against Lethal Septic Peritonitis. Shock 2019, 52, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Eliasson, L.; Carlén, A. An update on minor salivary gland secretions. Eur. J. Oral Sci. 2010, 118, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Aframian, D.J.; Keshet, N.; Nadler, C.; Zadik, Y.; Vered, M. Minor salivary glands: Clinical, histological and immunohistochemical features of common and less common pathologies. Acta Histochem. 2019, 121, 151451. [Google Scholar] [CrossRef] [PubMed]
- Meer, S. Human immunodeficiency virus and salivary gland pathology: An update. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2019, 128, 52–59. [Google Scholar] [CrossRef]
- Nittayananta, W.; Chanowanna, N.; Pruphetkaew, N.; Nauntofte, B. Relationship between xerostomia and salivary flow rates in HIV-infected individuals. J. Investig. Clin. Dent. 2013, 4, 164–171. [Google Scholar] [CrossRef]
- Nizamuddin, I.; Koulen, P.; McArthur, C. Contribution of HIV Infection, AIDS, and Antiretroviral Therapy to Exocrine Pathogenesis in Salivary and Lacrimal Glands. Int. J. Mol. Sci. 2018, 19, 2747. [Google Scholar] [CrossRef] [Green Version]
- Manikandan, M.; Deva Magendhra Rao, A.K.; Arunkumar, G.; Manickavasagam, M.; Rajkumar, K.S.; Rajaraman, R.; Munirajan, A.K. Oral squamous cell carcinoma: MicroRNA expression profiling and integrative analyses for elucidation of tumourigenesis mechanism. Mol. Cancer 2016, 15, 28. [Google Scholar] [CrossRef] [Green Version]
- Gallo, A.; Vella, S.; Tuzzolino, F.; Cuscino, N.; Cecchettini, A.; Ferro, F.; Mosca, M.; Alevizos, I.; Bombardieri, S.; Conaldi, P.G.; et al. MicroRNA-mediated Regulation of Mucin-type O-glycosylation Pathway: A Putative Mechanism of Salivary Gland Dysfunction in Sjögren Syndrome. J. Rheumatol. 2019, 46, 1485–1494. [Google Scholar] [CrossRef]
- Thorlacius, G.E.; Wahren-Herlenius, M.; Ronnblom, L. An update on the role of type I interferons in systemic lupus erythematosus and Sjogren’s syndrome. Curr. Opin. Rheumatol. 2018, 30, 471–481. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal ID | SIV Inoculum | Duration of Infection | Plasma Viral Loads 106/mL | OPM Viral Loads 106/mg RNA | Opportunistic Infections |
|---|---|---|---|---|---|
| Chronic SIV-Infected and Vehicle Untreated (Group 1) for microRNA and gene expression studies | |||||
| P763 # | SIVmac251 | 145 | NA | 3 | ND |
| JJ71 # * | SIVmac239 | 148 | NA | 0.3 | ND |
| JR60 # * | SIVmac251 | 180 | NA | 10 | ND |
| JR15 # * | SIVmac251 | 180 | NA | 0.3 | ND |
| KA05 # * | SIVmac251 | 90 | NA | 0.7 | ND |
| JP38 # * | SIVmac251 | 180 | NA | 0.1 | ND |
| JD59 # * | SIVmac251 | 150 | NA | 300 | ND |
| IJ60 * | SIVmac251 | 180 | NA | NA | ND |
| Chronic SIV-Infected and Vehicle treated (Group 2) for Immunofluorescence studies | |||||
| JH47 % | SIVmac251 | 180 | 2 | NA | ND |
| JR36 % | SIVmac251 | 180 | 0.5 | 0.02 | ND |
| JD66 % | SIVmac251 | 180 | 0.04 | 0.5 | ND |
| Chronic SIV-Infected and ∆9-THC treated (Group 3) for Immunofluorescence studies | |||||
| JI45 # % * | SIVmac251 | 180 | 3 | 0.02 | ND |
| JT80 # % * | SIVmac251 | 180 | 1 | 9 | ND |
| IV90 # % * | SIVmac251 | 180 | 0.02 | 0.3 | ND |
| Uninfected Controls (Group 4) for microRNA and gene expression studies | |||||
| JC65 # * | NA | NA | NA | NA | NA |
| GJ01 # * | NA | NA | NA | NA | NA |
| GK11 # * | NA | NA | NA | NA | NA |
| GK22 # * | NA | NA | NA | NA | NA |
| JD95 # * | NA | NA | NA | NA | NA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez, X.; Sestak, K.; Byrareddy, S.N.; Mohan, M. Long Term Delta-9-tetrahydrocannabinol Administration Inhibits Proinflammatory Responses in Minor Salivary Glands of Chronically Simian Immunodeficieny Virus Infected Rhesus Macaques. Viruses 2020, 12, 713. https://doi.org/10.3390/v12070713
Alvarez X, Sestak K, Byrareddy SN, Mohan M. Long Term Delta-9-tetrahydrocannabinol Administration Inhibits Proinflammatory Responses in Minor Salivary Glands of Chronically Simian Immunodeficieny Virus Infected Rhesus Macaques. Viruses. 2020; 12(7):713. https://doi.org/10.3390/v12070713
Chicago/Turabian StyleAlvarez, Xavier, Karol Sestak, Siddappa N. Byrareddy, and Mahesh Mohan. 2020. "Long Term Delta-9-tetrahydrocannabinol Administration Inhibits Proinflammatory Responses in Minor Salivary Glands of Chronically Simian Immunodeficieny Virus Infected Rhesus Macaques" Viruses 12, no. 7: 713. https://doi.org/10.3390/v12070713