Structural Similarities, in Relation with the Cross-Reactivity, of Hymenoptera Allergenic Dipeptidyl Peptidases IV—An Overall Comparison Including a New Dipeptidyl Peptidase IV Sequence from Vespa velutina

 , , , , , , , , and
, , , , , , , , and
Abstract
:1. Introduction
2. Results
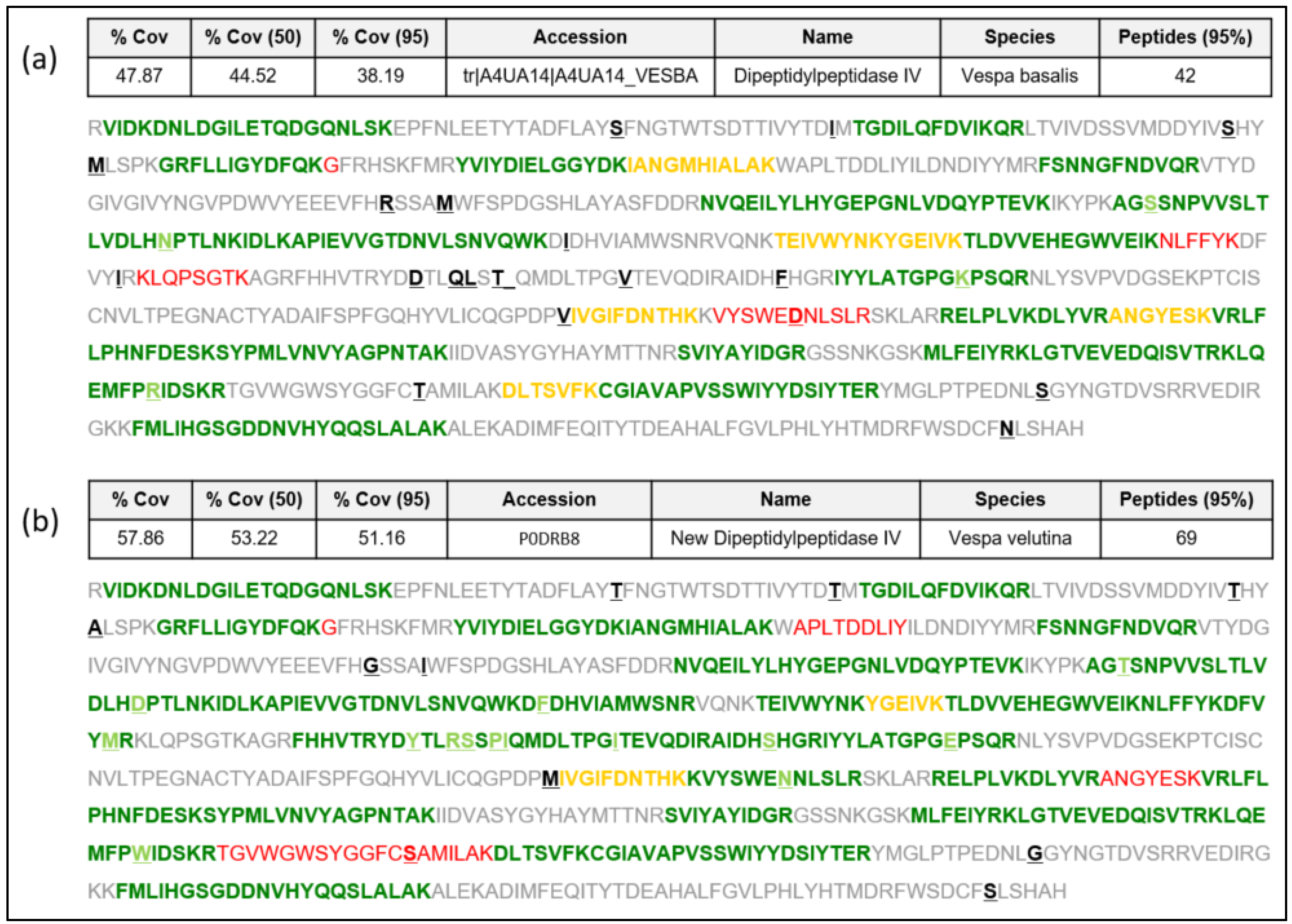
2.1. Structural Similarities in DPPIVS among Hymenoptera Species
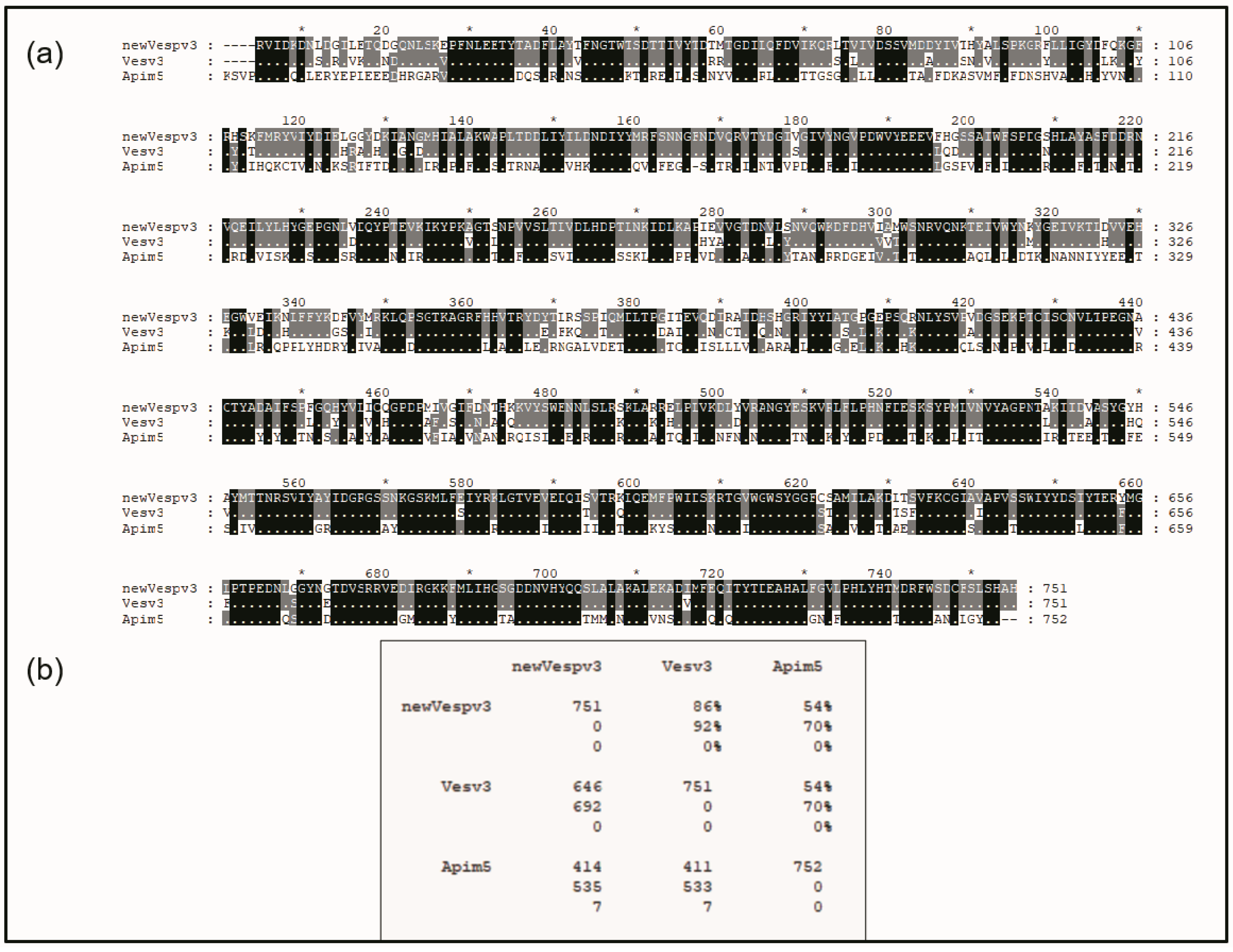
2.1.1. New Complete Vespa velutina Sequence and Comparison with Vespidae and Apidae DPPIVs
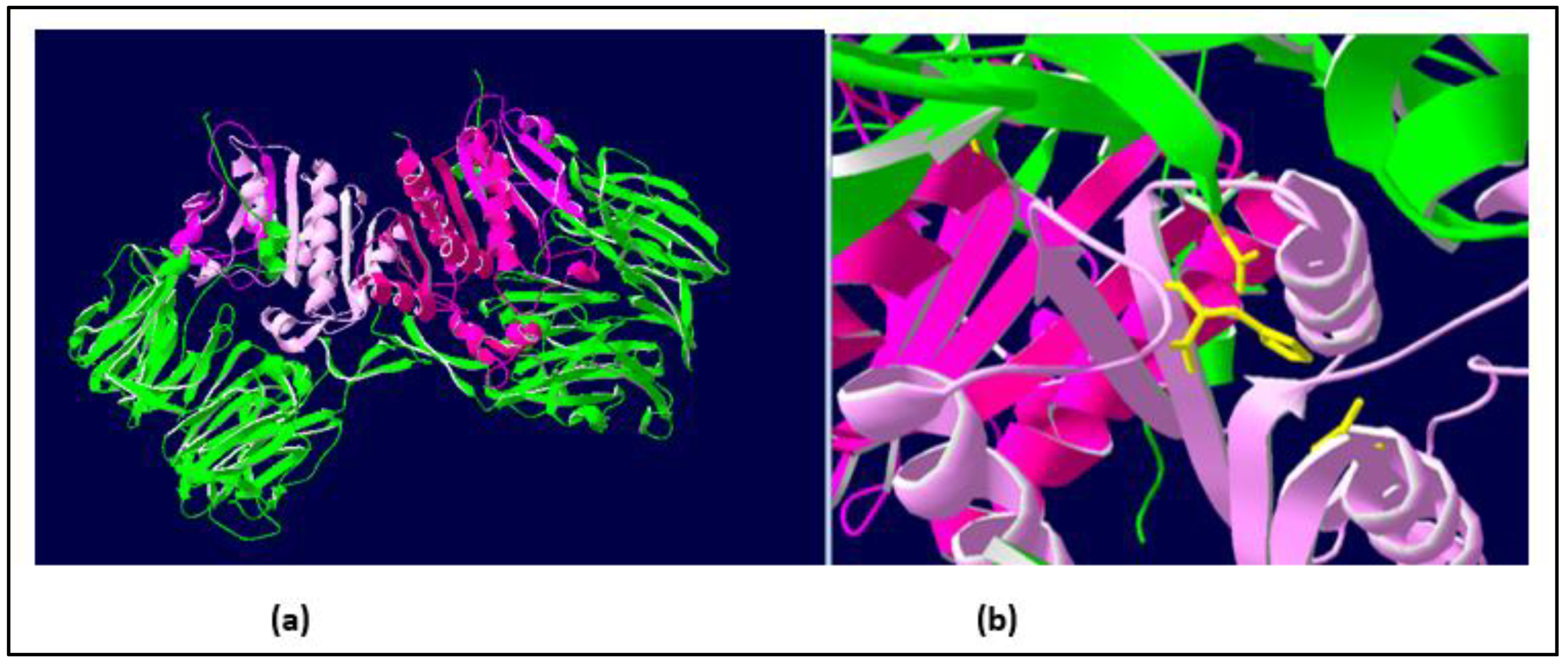
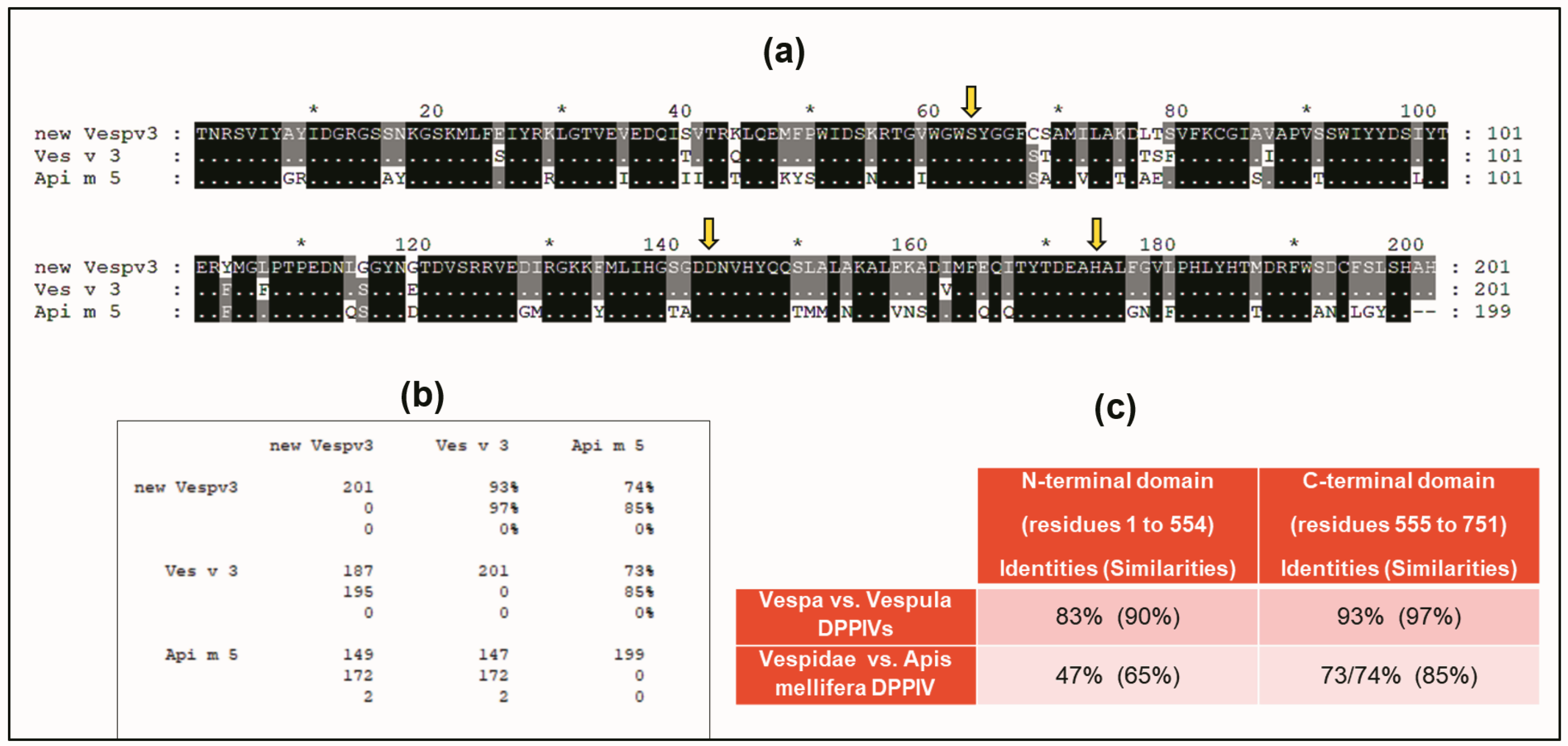
2.1.2. Similarities with Other DPPIV Enzymes: A Conserved Domain of the DPPIVs
3. Clinical Cases of Specific Allergenicity of Hymenoptera DPPIV Components
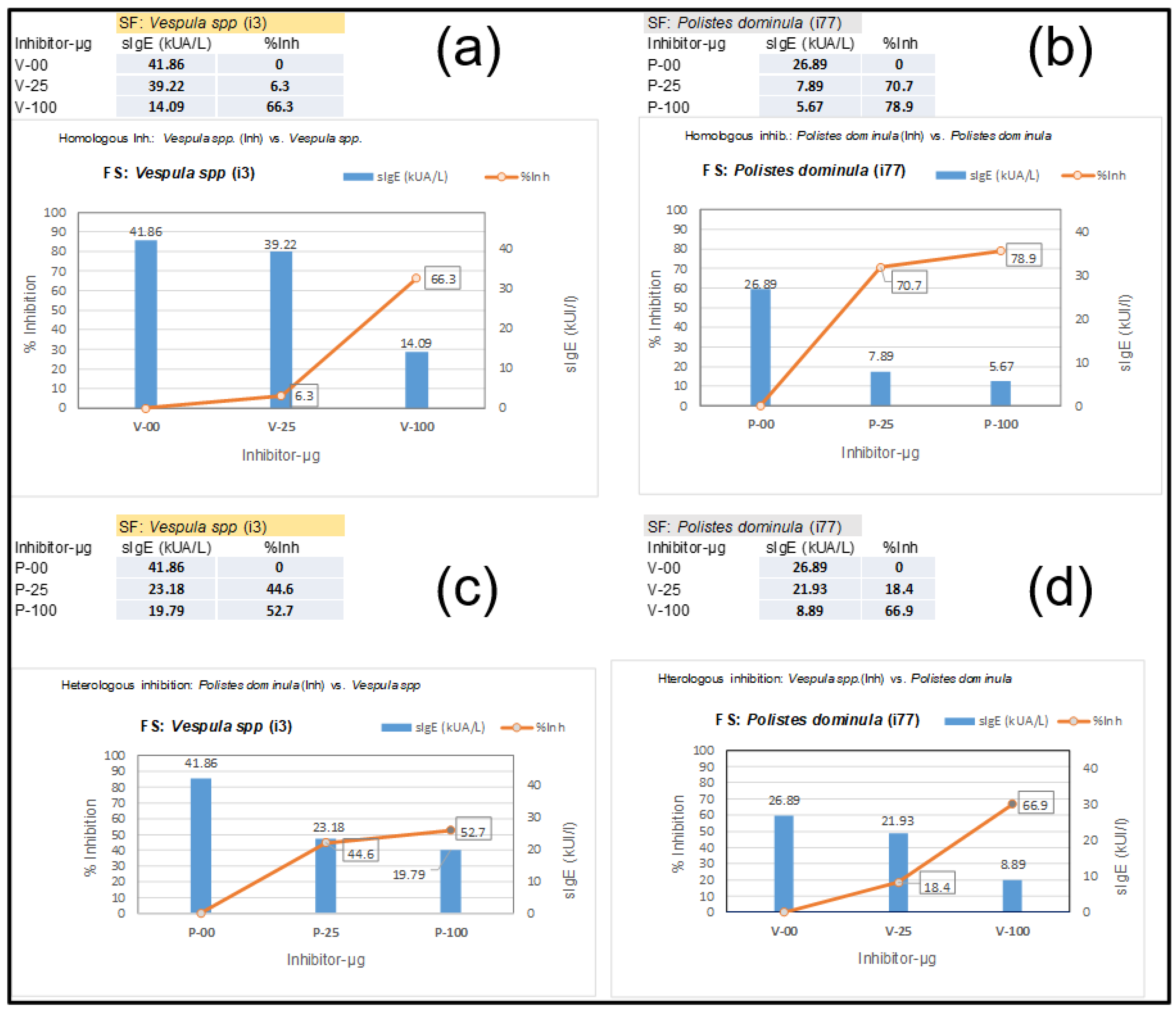
3.1. Ves v 3 Allergenicity Detected by Its Cross-Reactivity with Api m 5 in ImmunoCAP
3.2. Patient Predominantly Sensitized to Api m 5
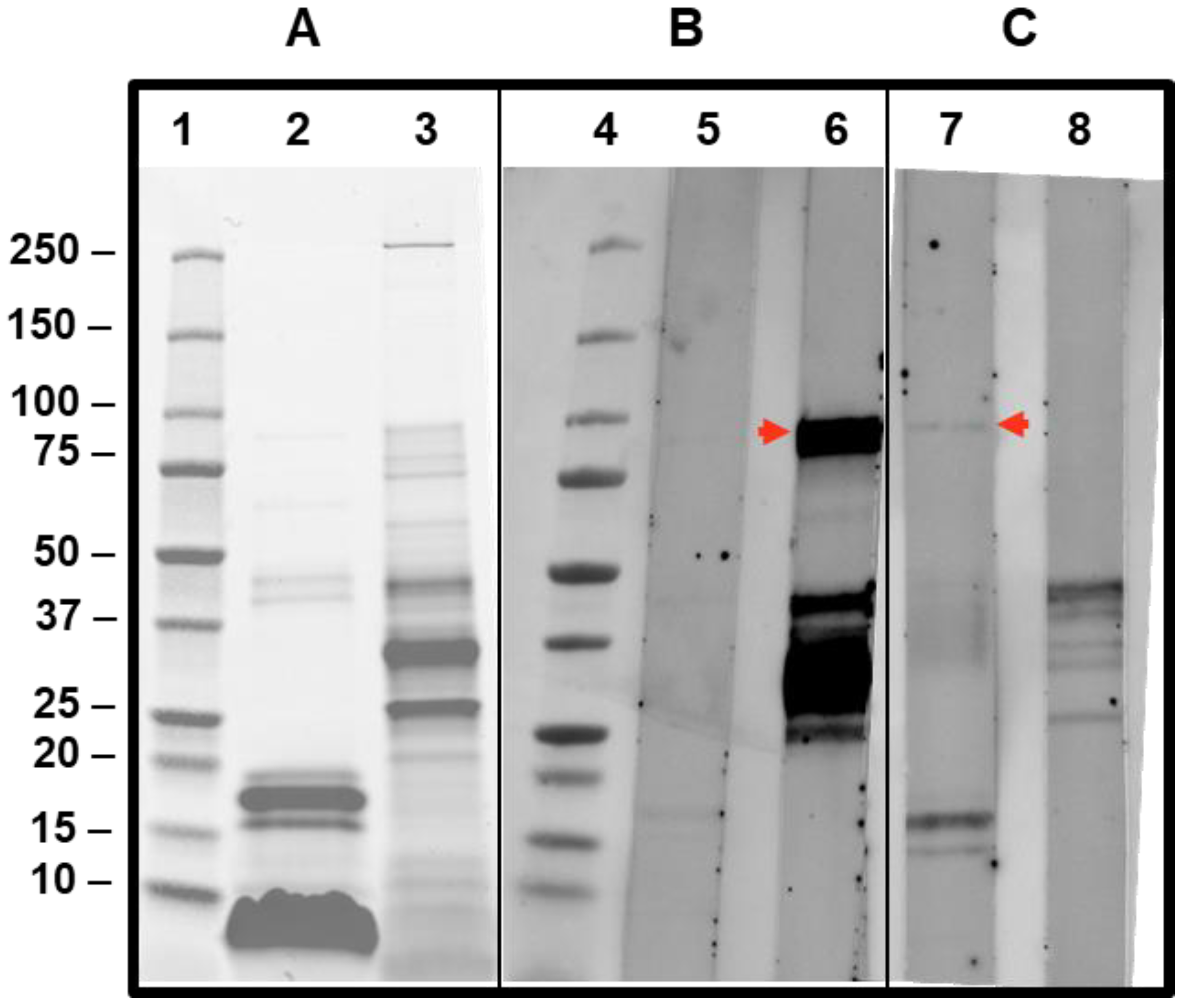
3.3. Immunoblot of Patient’s Sera Showing Positivity to DPPIV Components in Vespula spp. and Apis Mellifera Extract
4. Discussion
5. Material and Methods
5.1. Proteomic Studies
5.1.1. Venom Collection
5.1.2. Mass Spectrometric Analyses
5.2. SDS-PAGE and Immunoblotting
5.3. Measurements on ImmunoCAP and CAP-FEIA Inhibition Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aertgeerts, K.; Ye, S.; Tennant, M.G.; Kraus, M.L.; Rogers, J.; Sang, B.C.; Skene, R.J.; Webb, D.R.; Prasad, G.S. Crystal structure of human dipeptidyl peptidase IV in complex with a decapeptide reveals details on substrate specificity and tetrahedral intermediate formation. Protein Sci. 2004, 13, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Aertgeerts, K.; Ye, S.; Shi, L.; Prasad, S.G.; Witmer, D.; Chi, E.; Sang, B.C.; Wijnands, R.A.; Webb, D.R.; Swanson, R.V. N-linked glycosylation of dipeptidyl peptidase IV (CD26): Effects on enzyme activity, homodimer formation, and adenosine deaminase binding. Protein Sci. 2004, 13, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Ollert, M.; Blank, S. Anaphylaxis to insect venom allergens: Role of molecular diagnostics. Curr. Allergy Asthma Rep. 2015, 15, 26. [Google Scholar] [CrossRef] [PubMed]
- Blank, S.; Seismann, H.; Bockisch, B.; Braren, I.; Cifuentes, L.; McIntyre, M.; Ruhl, D.; Ring, J.; Bredehorst, R.; Ollert, M.W.; et al. Identification, recombinant expression, and characterization of the 100 kDa high molecular weight Hymenoptera venom allergens Api m 5 and Ves v 3. J. Immunol. 2010, 184, 5403–5413. [Google Scholar] [CrossRef]
- Schiener, M.; Hilger, C.; Eberlein, B.; Pascal, M.; Kuehn, A.; Revets, D.; Planchon, S.; Pietsch, G.; Serrano, P.; Moreno-Aguilar, C.; et al. The high molecular weight dipeptidyl peptidase IV Pol d 3 is a major allergen of Polistes dominula venom. Sci. Rep. 2018, 8, 1318. [Google Scholar] [CrossRef]
- Vidal, C.; Armisén, M.; Monsalve, R.; González-Vidal, T.; Lojo, S.; López-Freire, S.; Méndez, P.; Rodríguez, V.; Romero, L.; Galán, A.; et al. Anaphylaxis to Vespa velutina nigrithorax: Pattern of Sensitization for an Emerging Problem in Western Countries. J. Investig. Allergol. Clin. Immunol. 2021, 31, 228–235. [Google Scholar] [CrossRef]
- Pretre, G.; Asturias, J.A.; Lizaso, M.T.; Tabar, A.I. Dipeptidyl peptidase IV of the Vespa velutina nigrithorax venom is recognized as a relevant allergen. Ann. Allergy Asthma Immunol. 2022, 129, 101–105. [Google Scholar] [CrossRef]
- Lee, V.S.Y.; Tu, W.-C.; Jinn, T.-R.; Peng, C.-C.; Lin, L.-J.; Tzen, J.T.C. Molecular cloning of the precursor polypeptide of mastoparan B and its putative processing enzyme, dipeptidyl peptidase IV, from the black-bellied hornet, Vespa basalis. Insect Mol. Biol. 2007, 16, 231–237. [Google Scholar] [CrossRef]
- Bilò, M.B.; Ollert, M.; Blank, S. The role of component-resolved diagnosis in Hymenoptera venom allergy. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 614–622. [Google Scholar] [CrossRef]
- Kohler, J.; Blank, S.; Muller, S.; Bantleon, F.; Frick, M.; Huss-Marp, J.; Lidholm, J.; Spillner, E.; Jakob, T. Component resolution reveals additional major allergens in patients with honeybee venom allergy. J. Allergy. Clin. Immunol. 2014, 133, 1383–1389. [Google Scholar] [CrossRef]
- Arzt, L.; Bokanovic, D.; Schrautzer, C.; Schwarz, I.; Laipold, K.; Aberer, W.; Sturm, G.J. Questionable diagnostic benefit of the commercially available panel of bee venom components. Allergy 2017, 72, 1419–1422. [Google Scholar] [CrossRef] [PubMed]
- Chugo, S.; Lizaso, M.T.; Alvarez, M.J.; Arroabaren, E.; Lizarza, S.; Tabar, A.I. Vespa velutina nigritorax: A New Causative Agent in Anaphylaxis. J. Investig. Allergol. Clin. Immunol. 2015, 25, 231–232. [Google Scholar] [CrossRef] [PubMed]
- Budge, G.E.; Hodgetts, J.; Jones, E.P.; Ostojá-Starzewski, J.C.; Hall, J.; Tomkies, V.; Semmence, N.; Brown, M.; Wakefield, M.; Stainton, K. The invasion, provenance and diversity of Vespa velutina Lepeletier (Hymenoptera: Vespidae) in Great Britain. PLoS ONE 2017, 12, e0185172. [Google Scholar] [CrossRef] [PubMed]
- Pomes, A.; Davies, J.M.; Gadermaier, G.; Hilger, C.; Holzhauser, T.; Lidholm, J.; Lopata, A.L.; Mueller, G.A.; Nandy, A.; Radauer, C.; et al. WHO/IUIS Allergen Nomenclature: Providing a common language. Mol. Immunol. 2018, 100, 3–13. [Google Scholar] [CrossRef]
- King, T.P.; Hoffman, D.; Lowenstein, H.; Marsh, D.G.; Platts-Mills, T.A.; Thomas, W. Allergen nomenclature. WHO/IUIS Allergen Nomenclature Subcommittee. Int. Arch. Allergy Immunol. 1994, 105, 224–233. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Xie, C.; Zhu, B.; Zhu, H.; Liu, S.; Wang, W.; Chen, H.; Ji, Y. Deciphering the venomic transcriptome of killer-wasp Vespa velutina. Sci. Rep. 2015, 5, 9454. [Google Scholar]
- Monsalve, R.I.; Gutiérrez, R.; Hoof, I.; Lombardero, M. Purification and molecular characterization of phospholipase, antigen 5 and hyaluronidases from the venom of the Asian hornet (Vespa velutina). PLoS ONE 2020, 15, e0225672. [Google Scholar] [CrossRef]
- Alonso-Sampedro, M.; Feás, X.; Bravo, S.B.; Chantada-Vázquez, M.P.; Vidal, C. Proteomics of Vespa velutina nigrithorax Venom Sac Queens and Workers: A Quantitative SWATH-MS Analysis. Toxins 2023, 15, 266. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; Hewapathirana, S.; Castillo-Loja-Seisdedos, D.; Kamatchinathan, S.; Kundu, D.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A Hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
- Nicholas, H.B.; Ropelewski, A.J.; Deerfield, D.W. Strategies for multiple sequence alignment. BioTechniques 2002, 32, 572–591. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.F.; Coulson, A.F. Significance of protein sequence similarities. Methods Enzymol. 1990, 183, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-León, B.; Navas, A.; Serrano, P.; Espinazo, M.; Labrador-Horrillo, M.; Monsalve, R.I.; Jurado, A.; Moreno-Aguilar, C. Successful Adaptation of Bee Venom Immunotherapy in a Patient Monosensitized to Api m 10. J. Investig. Allergol. Clin. Immunol. 2020, 30, 296–298. [Google Scholar] [CrossRef]
- Straumann, F.; Bucher, C.; Wuthrich, B. Double sensitization to honeybee and wasp venom: Immunotherapy with one or with both venoms? Value of FEIA inhibition for the identification of the cross-reacting IgE antibodies in double-sensitized patients to honeybee and wasp venom. Int. Arch. Allergy Immunol. 2000, 123, 268–274. [Google Scholar] [CrossRef]
- Suarez-Fueyo, A.; Ramos, T.; Galan, A.; Jimeno, L.; Wurtzen, P.A.; Marin, A.; de Frutos, C.; Blanco, C.; Carrera, A.C.; Barber, D.; et al. Grass tablet sublingual immunotherapy downregulates the TH2 cytokine response followed by regulatory T-cell generation. J. Allergy Clin. Immunol. 2014, 133, 130–138. [Google Scholar] [CrossRef]
- Francis, J.N.; James, L.K.; Paraskevopoulos, G.; Wong, C.; Calderon, M.A.; Durham, S.R.; Till, S.J. Grass pollen immunotherapy: IL-10 induction and suppression of late responses precedes IgG4 inhibitory antibody activity. J. Allergy Clin. Immunol. 2008, 121, 1120–1125. [Google Scholar] [CrossRef]
- DeLuca, K.F.; Mick, J.E.; DeLuca, J.G. Production and purification of recombinant monoclonal antibodies from human cells based on a primary sequence. STAR Protoc. 2022, 3, 101915. [Google Scholar] [CrossRef]
- Barber, D.; Diaz-Perales, A.; Escribese, M.M.; Kleine-Tebbe, J.; Matricardi, P.M.; Ollert, M.; Santos, A.F.; Sastre, J. Molecular allergology and its impact in specific allergy diagnosis and therapy. Allergy 2021, 76, 3642–3658. [Google Scholar] [CrossRef]
- Quercia, O.; Cova, V.; Martini, M.; Cortellini, G.; Murzilli, F.; Bignardi, D.; Cilia, M.; Scarpa, A.; Bilò, M.B. CAP-Inhibition, Molecular Diagnostics, and Total IgE in the Evaluation of Polistes and Vespula Double Sensitization. Int. Arch. Allergy Immunol. 2018, 177, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Caruso, B.; Bonadonna, P.; Severino, M.G.; Manfredi, M.; Dama, A.; Schiappoli, M.; Rizzotti, P.; Senna, G.; Passalacqua, G. Evaluation of the IgE cross-reactions among vespid venoms. A possible approach for the choice of immunotherapy. Allergy 2007, 62, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Blank, S.; Etzold, S.; Darsow, U.; Schiener, M.; Eberlein, B.; Russkamp, D.; Wolf, S.; Graessel, A.; Biedermann, T.; Ollert, M.; et al. Component-resolved evaluation of the content of major allergens in therapeutic extracts for specific immunotherapy of honeybee venom allergy. Hum. Vaccines Immunother. 2017, 13, 2482–2489. [Google Scholar] [CrossRef] [PubMed]
- Bilò, M.B.; Antonicelli, L.; Bonifazi, F. Purified vs. nonpurified venom immunotherapy. Curr. Opin. Allergy Clin. Immunol. 2010, 10, 330–336. [Google Scholar] [CrossRef]
- Perez-Hernandez, D.; Gutiérrez-Vázquez, C.; Jorge, I.; López-Martín, S.; Ursa, A.; Sánchez-Madrid, F.; Vázquez, J.; Yáñez-Mó, M. The intracellular interactome of tetraspanin-enriched microdomains reveals their function as sorting machineries toward exosomes. J. Biol. Chem. 2013, 288, 11649–11661. [Google Scholar] [CrossRef]
- Bonzon-Kulichenko, E.; Pérez-Hernández, D.; Núñez, E.; Martínez-Acedo, P.; Navarro, P.; Trevisan-Herraz, M.; Del Ramos, M.C.; Sierra, S.; Martínez-Martínez, S.; Ruiz-Meana, M.; et al. A robust method for quantitative high-throughput analysis of proteomes by 18O labeling. Mol. Cell. Proteom. 2011, 10, M110.003335. [Google Scholar] [CrossRef]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Jensen, O.N.; Podtelejnikov, A.V.; Neubauer, G.; Mortensen, P.; Mann, M. A strategy for identifying gel-separated proteins in sequence databases by MS alone. Biochem. Soc. Trans. 1996, 24, 893–896. [Google Scholar] [CrossRef]
- Peñas-Martínez, J.; Barrachina, M.N.; Cuenca-Zamora, E.J.; Luengo-Gil, G.; Bravo, S.B.; Caparrós-Pérez, E.; Teruel-Montoya, R.; Eliseo-Blanco, J.; Vicente, V.; García, Á.; et al. Qualitative and Quantitative Comparison of Plasma Exosomes from Neonates and Adults. Int. J. Mol. Sci. 2021, 22, 1926. [Google Scholar] [CrossRef]
- Camino, T.; Lago-Baameiro, N.; Sueiro, A.; Bravo, S.B.; Couto, I.; Santos, F.F.; Baltar, J.; Casanueva, F.F.; Pardo, M. Brown Adipose Tissue Sheds Extracellular Vesicles That Carry Potential Biomarkers of Metabolic and Thermogenesis Activity Which Are Affected by High Fat Diet Intervention. Int. J. Mol. Sci. 2022, 23, 10826. [Google Scholar] [CrossRef]
- Chantada-Vázquez, M.P.; Conde-Amboage, M.; Graña-López, L.; Vázquez-Estévez, S.; Bravo, S.B.; Núñez, C. Circulating Proteins Associated with Response and Resistance to Neoadjuvant Chemotherapy in HER2-Positive Breast Cancer. Cancers 2022, 14, 1087. [Google Scholar] [CrossRef]
- Shilov, I.V.; Seymour, S.L.; Patel, A.A.; Loboda, A.; Tang, W.H.; Keating, S.P.; Hunter, C.L.; Nuwaysir, L.M.; Schaeffer, D.A. The Paragon Algorithm, a next generation search engine that uses sequence temperature values and feature probabilities to identify peptides from tandem mass spectra. Mol. Cell. Proteom. 2007, 6, 1638–1655. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Madrid, F.; Morago, G.; Corbi, A.L.; Carreira, J. Monoclonal antibodies to three distinct epitopes on human IgE: Their use for determination of allergen-specific IgE. J. Immunol. Methods 1984, 73, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Monsalve, R.I.; Vega, A.; Marques, L.; Miranda, A.; Fernandez, J.; Soriano, V.; Cruz, S.; Dominguez-Noche, C.; Sanchez-Morillas, L.; Armisen-Gil, M.; et al. Component-resolved diagnosis of vespid venom-allergic individuals: Phospholipases and antigen 5s are necessary to identify Vespula or Polistes sensitization. Allergy 2012, 67, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Jappe, U.; Raulf-Heimsoth, M.; Hoffmann, M.; Burow, G.; Hubsch-Muller, C.; Enk, A. In vitro hymenoptera venom allergy diagnosis: Improved by screening for cross-reactive carbohydrate determinants and reciprocal inhibition. Allergy 2006, 61, 1220–1229. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sIgE (kUA/L) | IgG4 (mgA/L) | IgE-Total (kU/L) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ves v 1 (i211) | Ves v 5 (i209) | Pol d 1 (h6x*) | Pol d 5 (i210) | Vespula spp. (i3) | Polistes spp. (i4) | Apis mellifera (i1) | rApi m 1 (i208) | rApi m 3 (i215) | rApi m 10 (i217) | rApi m 5 (i216) | Vespula spp. (i3) | t-IgE | |
| Basal serum | 42.7 | 2.14 | 8.49 | 10.5 | 44.8 | 30.0 | 3.58 | 0.0 | 0.0 | 0.01 | 54.9 | Ndet. | 322 |
| Serum (6M) | 5.83 | 0.79 | 1.22 | 4.3 | 13.0 | Ndet. | 4.18 | Ndet. | Ndet. | Ndet. | 27.4 | 11.1 | 281 |
| Serum (12M) | 3.18 | 0.39 | 0.89 | 2.81 | 7.89 | Ndet. | Ndet. | Ndet. | Ndet. | Ndet. | 14.5 | 11.6 | 178 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monsalve, R.I.; Lombardero, M.; Christensen, L.H.; Núñez-Acevedo, B.; González-de-Olano, D.; Sobrino-García, M.; Castillo-Loja, R.M.; Bravo, S.B.; Alonso-Sampedro, M.; Vidal, C. Structural Similarities, in Relation with the Cross-Reactivity, of Hymenoptera Allergenic Dipeptidyl Peptidases IV—An Overall Comparison Including a New Dipeptidyl Peptidase IV Sequence from Vespa velutina. Toxins 2023, 15, 656. https://doi.org/10.3390/toxins15110656
Monsalve RI, Lombardero M, Christensen LH, Núñez-Acevedo B, González-de-Olano D, Sobrino-García M, Castillo-Loja RM, Bravo SB, Alonso-Sampedro M, Vidal C. Structural Similarities, in Relation with the Cross-Reactivity, of Hymenoptera Allergenic Dipeptidyl Peptidases IV—An Overall Comparison Including a New Dipeptidyl Peptidase IV Sequence from Vespa velutina. Toxins. 2023; 15(11):656. https://doi.org/10.3390/toxins15110656
Chicago/Turabian StyleMonsalve, Rafael I., Manuel Lombardero, Lars H. Christensen, Beatriz Núñez-Acevedo, David González-de-Olano, Miriam Sobrino-García, Rosita M. Castillo-Loja, Susana B. Bravo, Manuela Alonso-Sampedro, and Carmen Vidal. 2023. "Structural Similarities, in Relation with the Cross-Reactivity, of Hymenoptera Allergenic Dipeptidyl Peptidases IV—An Overall Comparison Including a New Dipeptidyl Peptidase IV Sequence from Vespa velutina" Toxins 15, no. 11: 656. https://doi.org/10.3390/toxins15110656