
Ecological Niche Modeling of Invasive Macrophyte (Urochloa subquadripara) and Co-Occurrence with South American Natives
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Global Distribution of Urochloa subquadripara, Eichhornia crassipes and Salvinia minima
2.2. CLIMEX
2.3. Parameter Adjustments and Model Validation in the CLIMEX Software
2.3.1. Urochloa Subquadripara
Growth Indices
Stress Parameters
2.3.2. Eichhornia crassipes
Growth Indices
Stress Parameters
2.3.3. Salvinia minima
Growth Indices
Stress Parameters
2.4. Climate Data, Models, and Scenarios
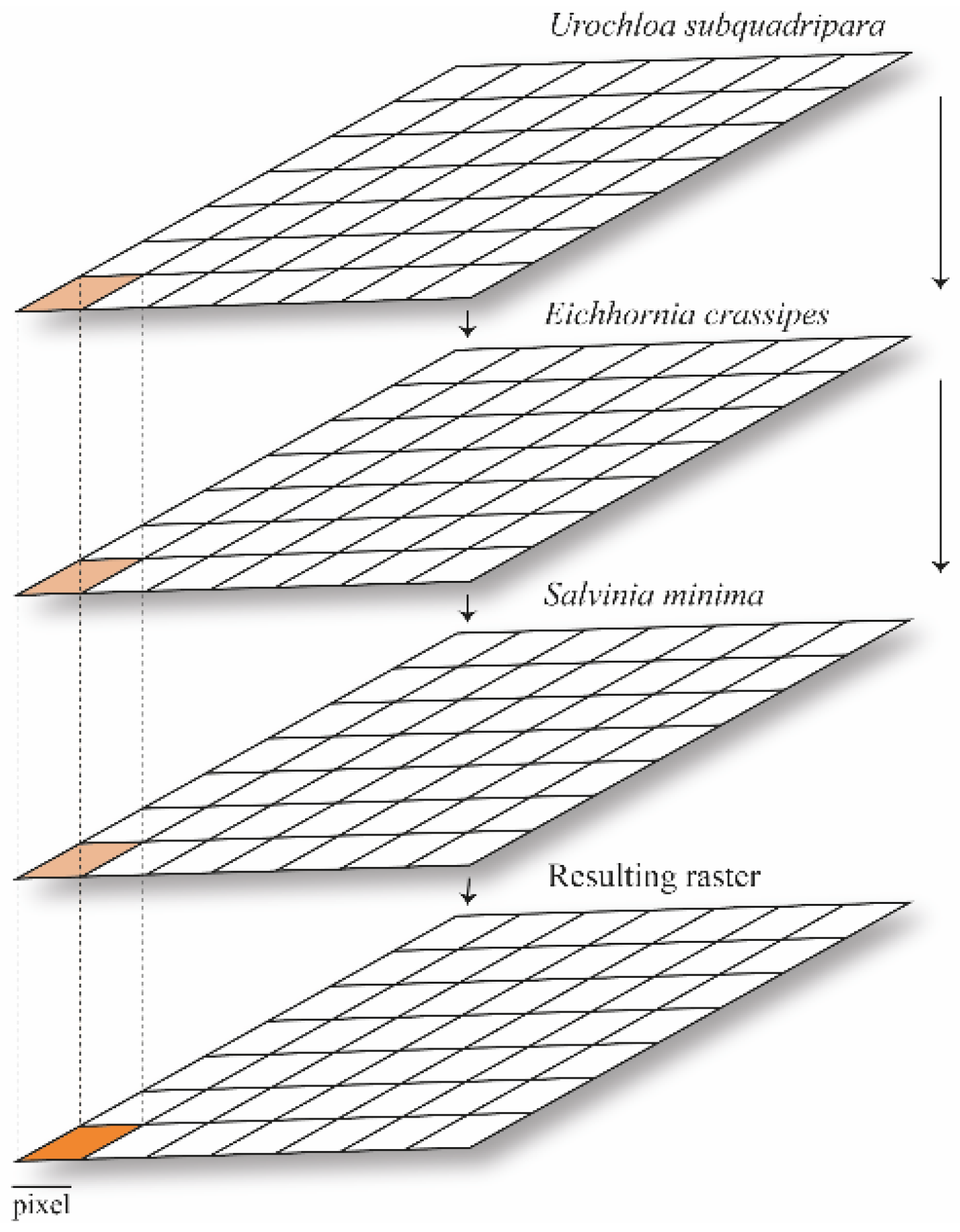
2.5. Multicriteria Decision-Making and Analytical Hierarchy Process
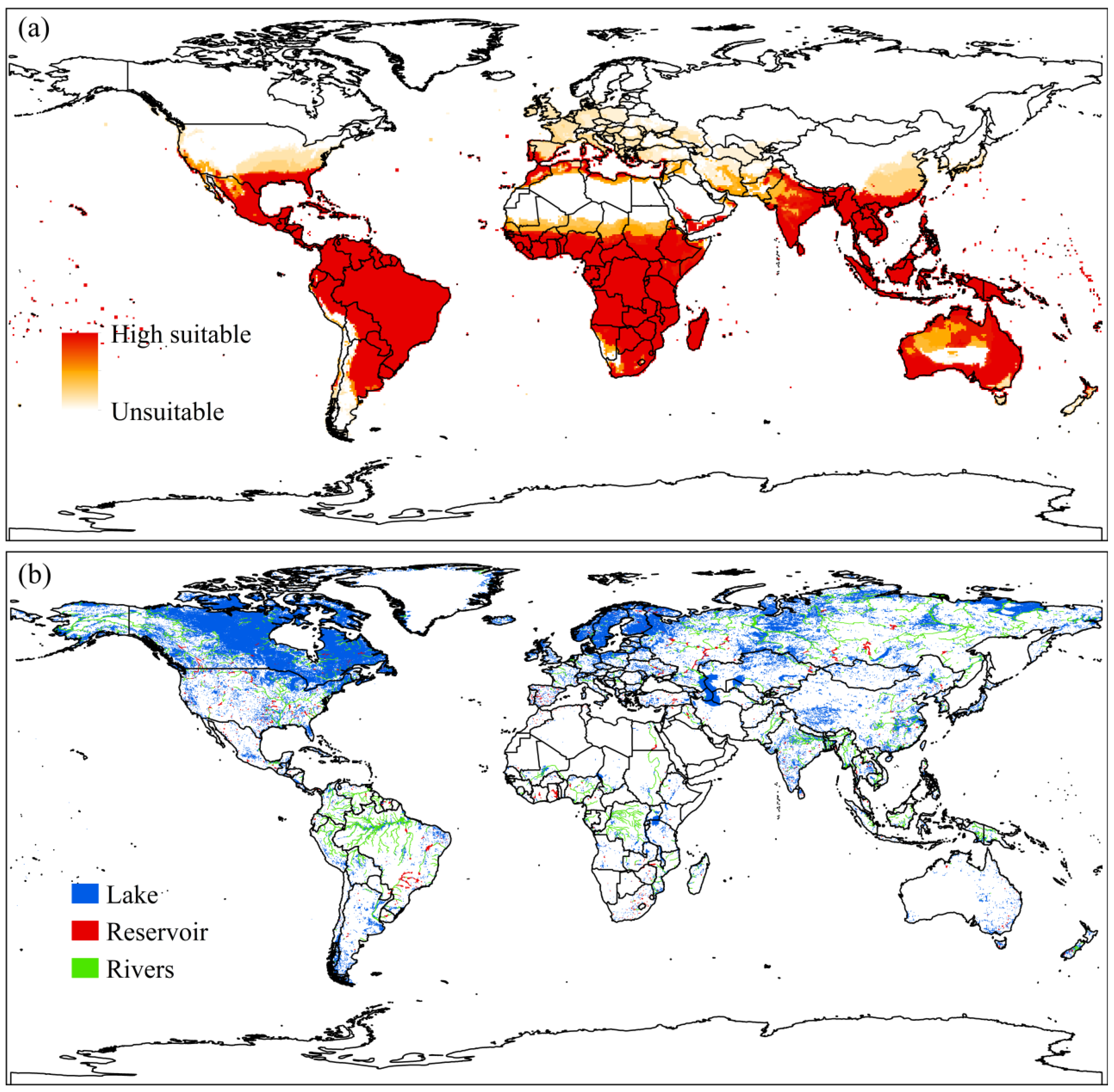
2.6. Global Lakes and Wetlands Database (GLWD)
3. Results
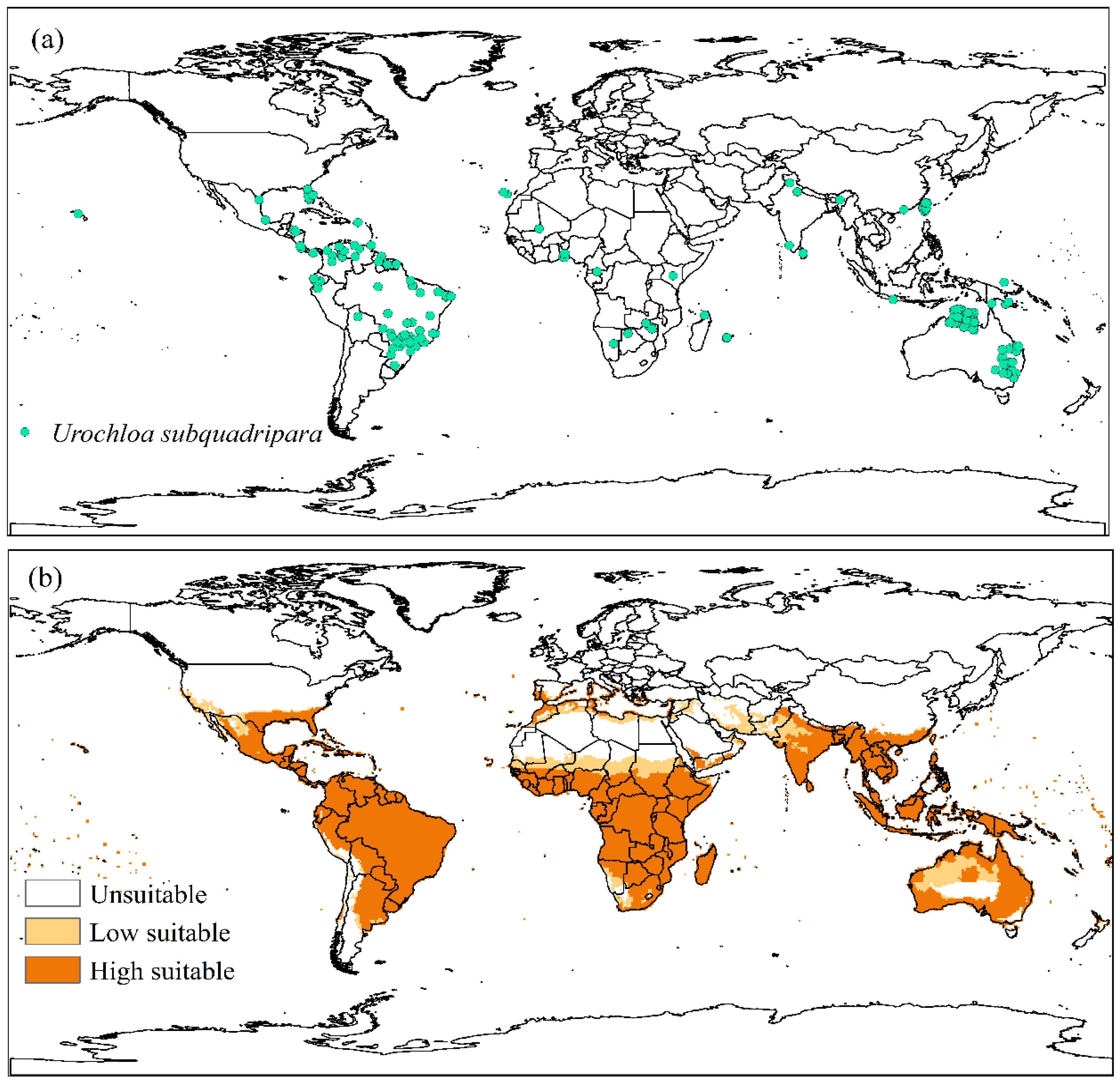
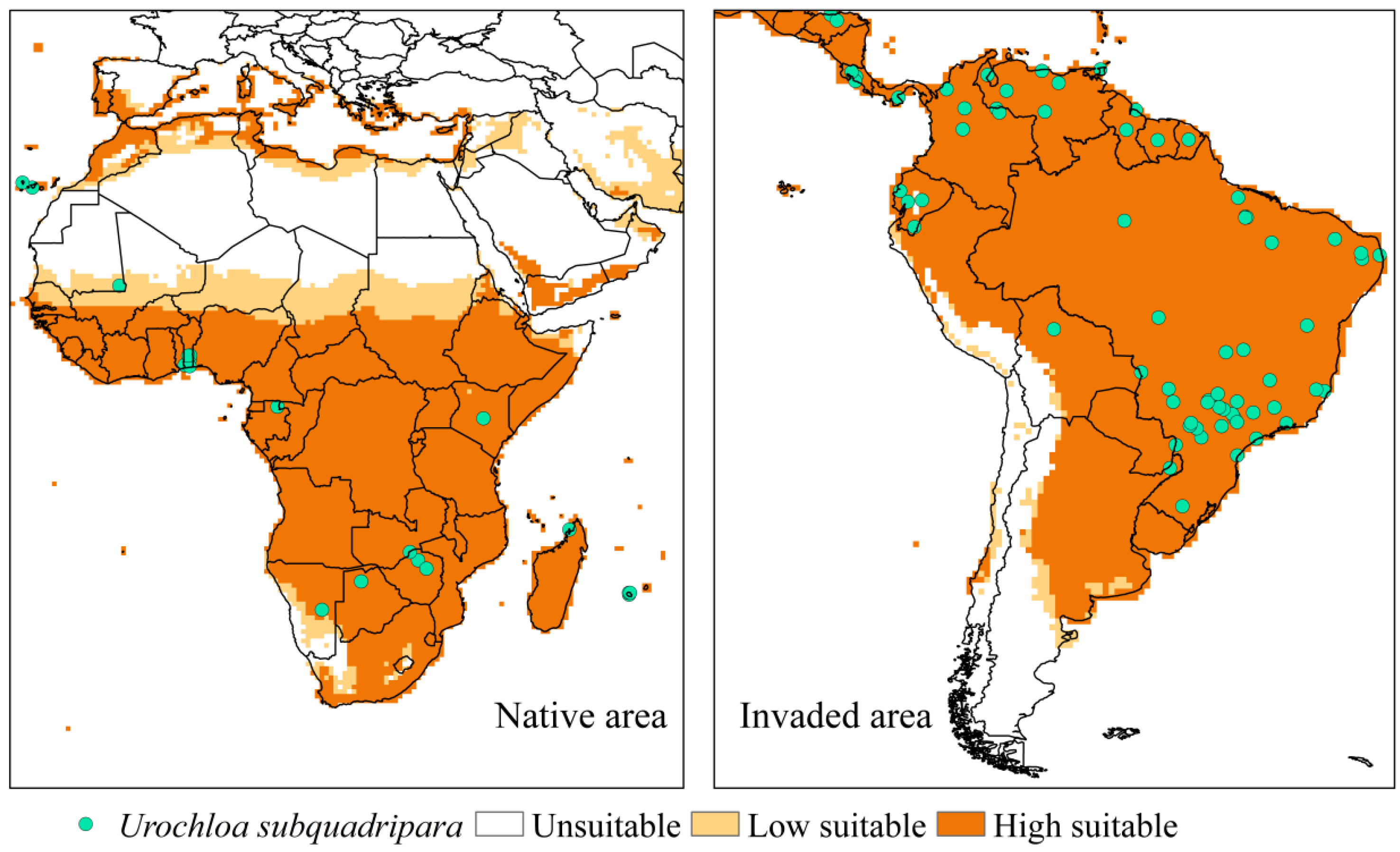
3.1. Invasive Exotic Species (Urochloa subquadripara)
3.2. Native Species
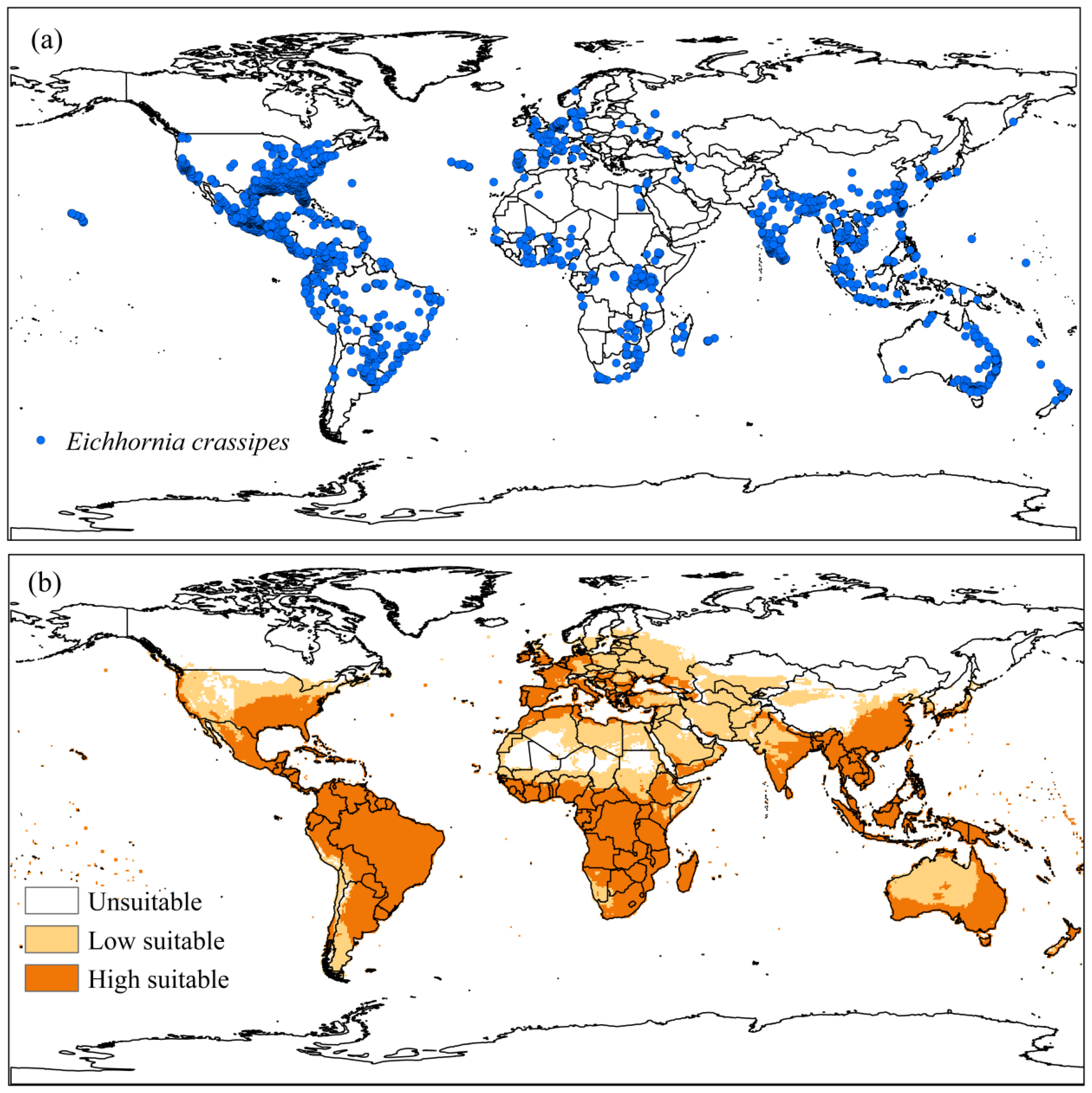
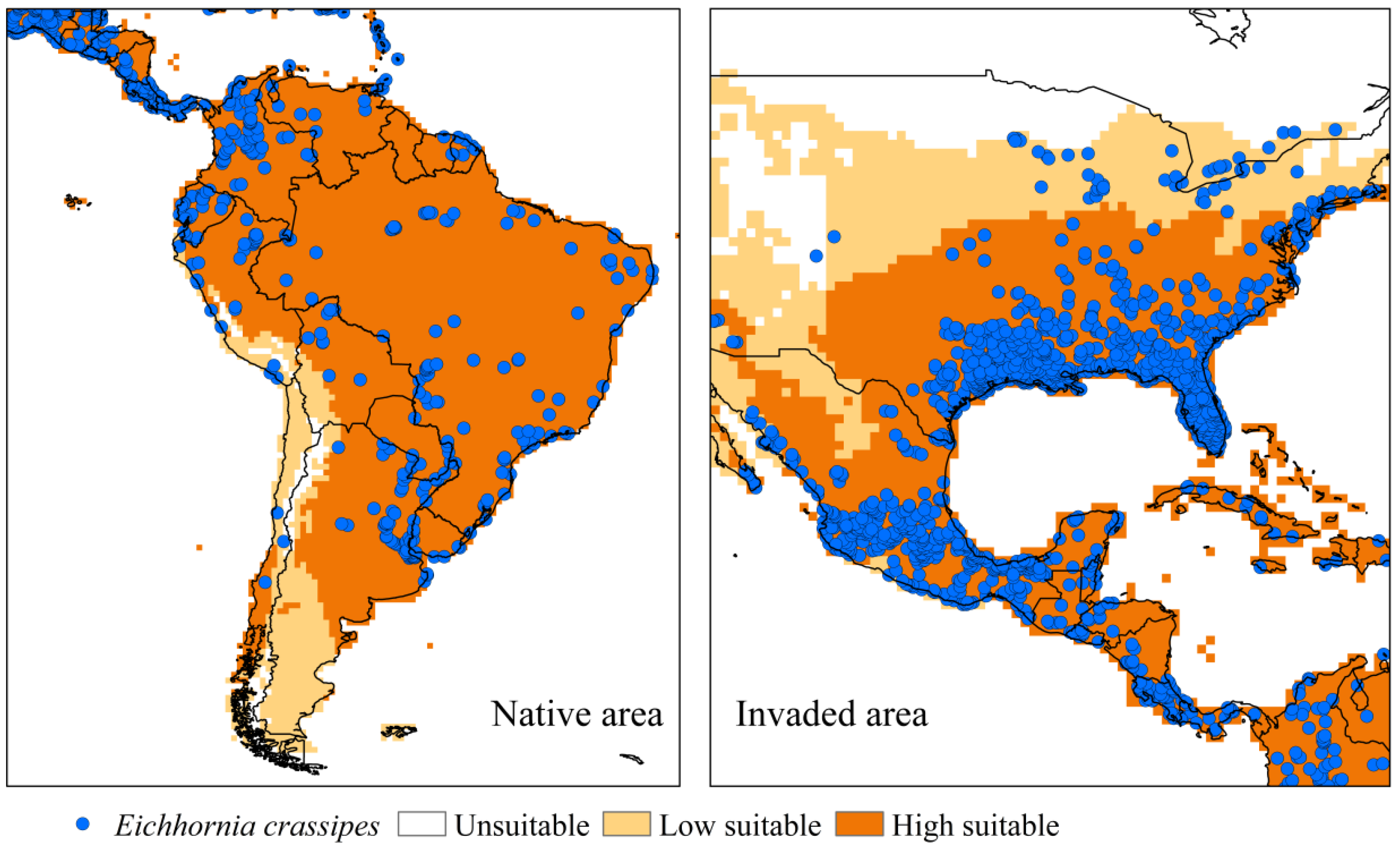
3.2.1. Eichhornia crassipes
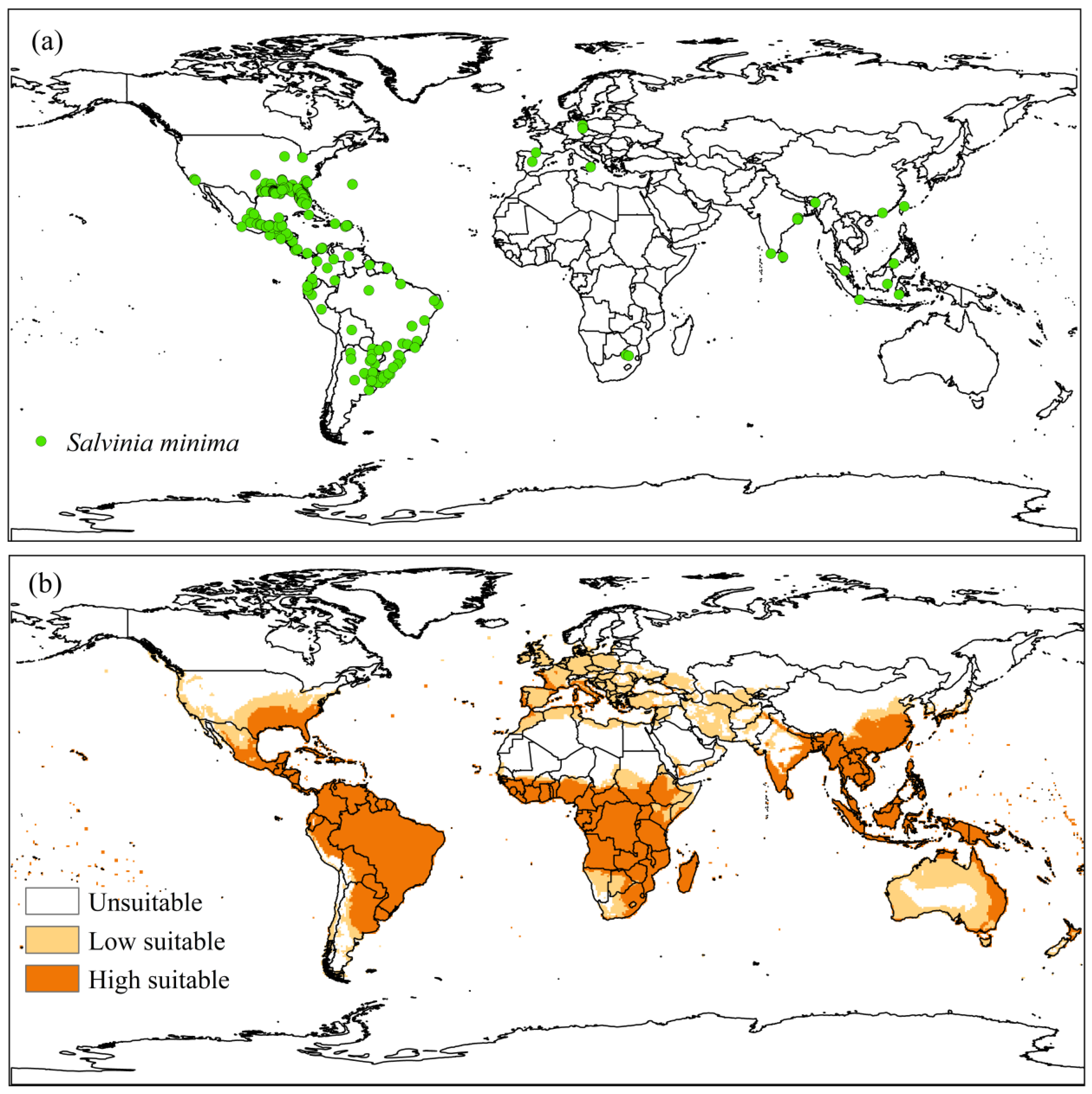
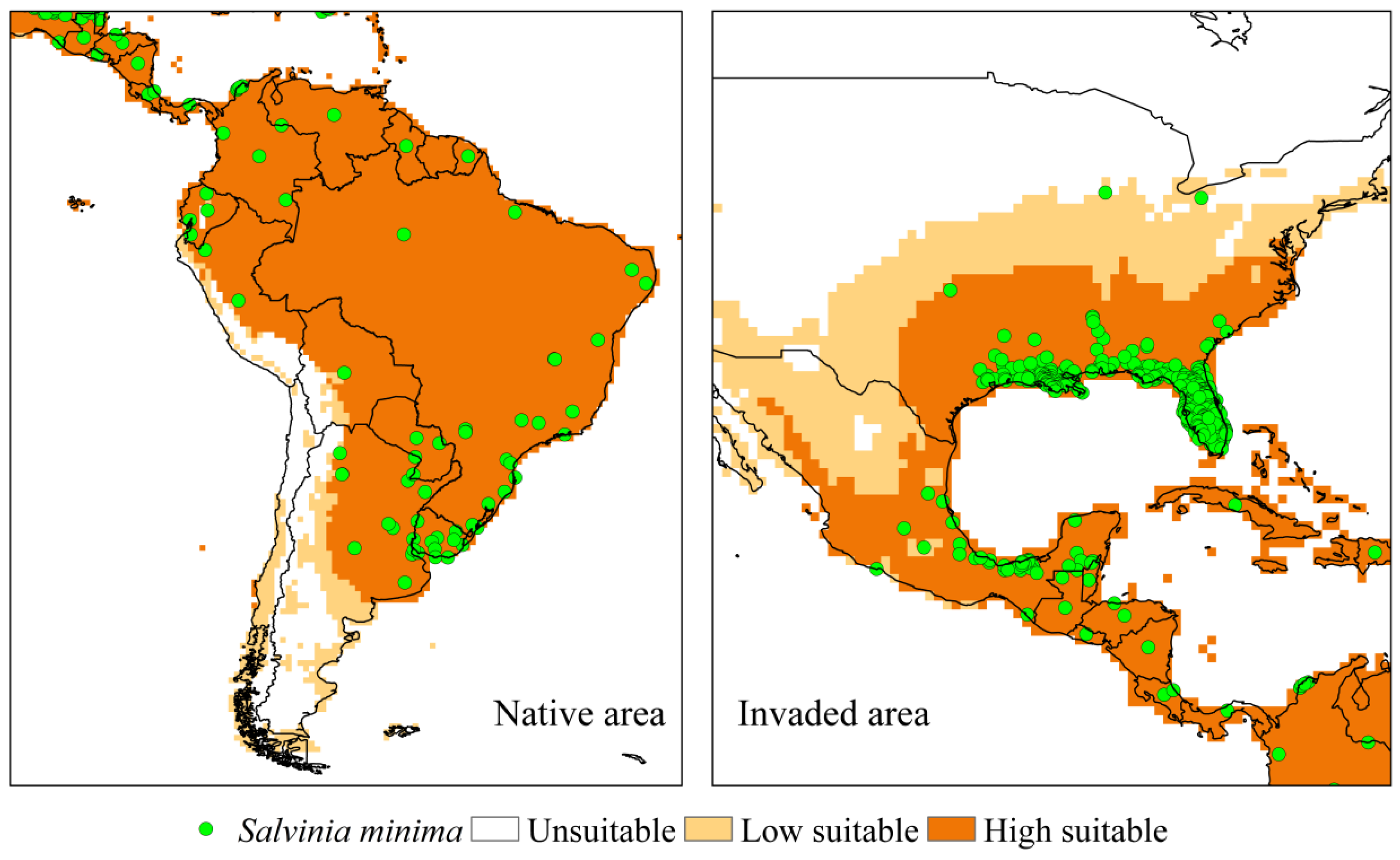
3.2.2. Salvinia minima
3.3. Co-Occurrence of Urochloa subquadripara with Native Species
4. Discussion
4.1. Invasive Exotic Species (Urochloa subquadripara)
4.2. Native Species
4.2.1. Eichhornia crassipes
4.2.2. Salvinia minima
4.3. Co-Occurrence of Urochloa subquadripara with Native Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tasker, S.J.; Foggo, A.; Bilton, D.T. Quantifying the ecological impacts of alien aquatic macrophytes: A global meta-analysis of effects on fish, macroinvertebrate and macrophyte assemblages. Freshw. Biol. 2022, 67, 1847–1860. [Google Scholar] [CrossRef]
- Dudgeon, D. Freshwater Biodiversity; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Merotto, A., Jr.; Gallon, M.; Turra, G.M.; Perez, N.B.; Lamego, F.P.; Cutti, L.; Angonese, P.S. Use of a wiper applicator for the control of the invasive species Eragrostis plana with glyphosate salts. Adv. Weed Sci. 2022, 40. [Google Scholar] [CrossRef] [PubMed]
- Diagne, C.; Leroy, B.; Vaissière, A.-C.; Gozlan, R.E.; Roiz, D.A.; Jarić, I.; Salles, J.-M.; Bradshaw, C.J.A.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Diamante, N.A.; Fabrin, T.M.; Silveira, M.J.; de Oliveira, A.V.; Thomaz, S.M.; Prioli, S.M.A.P.; Prioli, A.J. Molecular analysis of the invasive populations of Urochloa (Poaceae) in a large Neotropical reservoir. Aquat. Bot. 2020, 161, 103183. [Google Scholar] [CrossRef]
- Carniatto, N.; Thomaz, S.M.; Cunha, E.R.; Fugi, R.; Ota, R.R. Effects of an invasive alien Poaceae on aquatic macrophytes and fish communities in a Neotropical reservoir. Biotropica 2013, 45, 747–754. [Google Scholar] [CrossRef]
- Thomaz, S.M.; Carvalho, P.; Mormul, R.P.; Ferreira, F.A.; Silveira, M.J.; Michelan, T.S. TTemporal trends and effects of diversity on occurrence of exotic macrophytes in a large reservoir. Acta Oecologica 2009, 35, 614–620. [Google Scholar] [CrossRef]
- Thomaz, S.M.; Silveira, M.J.; Michelan, T.S. The colonization success of an exotic Poaceae is related to native macrophyte richness, wind disturbance and riparian vegetation. Aquat. Sci. 2012, 74, 809–815. [Google Scholar] [CrossRef]
- Fares, A.L.B.; Nonato, F.A.S.; Michelan, T.S. New records of the invasive macrophyte, Urochloa arrecta extend its range to eastern Brazilian Amazon altered freshwater ecosystems. Acta Amaz. 2020, 50, 133–137. [Google Scholar] [CrossRef]
- Amorim, S.R.; Umetsu, C.A.; Camargo, A.F.M. Effects of a non-native species of Poaceae on aquatic macrophyte community composition: A comparison with a native species. J. Aquat. Plant Manag. 2015, 53, 191–196. [Google Scholar]
- Michelan, T.S.; Thomaz, S.M.; Mormul, R.P.; Carvalho, P. Effects of an exotic invasive macrophyte (tropical signalgrass) on native plant community composition, species richness and functional diversity. Freshw. Biol. 2010, 55, 1315–1326. [Google Scholar] [CrossRef]
- Michelan, T.S.; Dainez Filho, M.S.; Thomaz, S.M. Aquatic macrophyte mats as dispersers of one invasive plant species. Braz. J. Biol. 2017, 78, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Martins, D.; Costa, N.V.; Terra, M.A.; Marchi, S.R. Caracterização da comunidade de plantas aquáticas de dezoito reservatórios pertencentes a cinco bacias hidrográficas do estado de São Paulo. Planta Daninha 2008, 26, 17–32. [Google Scholar] [CrossRef]
- Pott, V.; Pott, A.; Lima, L.; Moreira, S.; Oliveira, A. Aquatic macrophyte diversity of the Pantanal wetland and upper basin. Braz. J. Biol. 2011, 71, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Flora do Brasil. Available online: http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB26027 (accessed on 13 July 2020).
- Domingos, V.D.; Martins, D.; Costa, N.V.; da Costa, A.C.P.R. Initial growth of Brachiaria subquadripara (Trin.) Hitchc. plants under different nutritional conditions. Rev. Bras. Eng. Agrícola Ambient. 2015, 19, 560–566. [Google Scholar] [CrossRef]
- Moraes, I.d.C.; Pereira, W.A.; Nani, T.F.; de Paula, C.M.P.; da Rocha, M.J.; Sobrinho, F.d.S.; Techio, V.H. Karyotype analysis and mode of reproduction of two species of Urochloa P. Beauv. Crop Sci. 2021, 61, 3415–3424. [Google Scholar] [CrossRef]
- Pitelli, R.L.C.M.; Toffaneli, C.M.; Vieira, E.A.; Pitelli, R.A.; Velini, E.D. Dinâmica da comunidade de macrófitas aquáticas no reservatório de Santana, RJ. Planta Daninha 2008, 26, 473–480. [Google Scholar] [CrossRef]
- Michelan, T.S.; Thomaz, S.M.; Bini, L.M. Native macrophyte density and richness affect the invasiveness of a tropical Poaceae species. PLoS ONE 2013, 8, e60004. [Google Scholar] [CrossRef]
- Thomaz, S.M.; Michelan, T.S. Associations between a highly invasive species and native macrophytes differ across spatial scales. Biol. Invasions 2011, 13, 1881–1891. [Google Scholar] [CrossRef]
- Capers, R.S.; Selsky, R.; Bugbee, G.J.; White, J.C. Aquatic plant community invasibility and scale-dependent patterns in native and invasive species richness. Ecology 2007, 88, 3135–3143. [Google Scholar] [CrossRef]
- Fridley, J.D.; Stachowicz, J.J.; Naeem, S.; Sax, D.F.; Seabloom, E.W.; Smith, M.D.; Stohlgren, T.J.; Tilman, D.; Von Holle, B. The invasion paradox: Reconciling pattern and process in species invasions. Ecology 2007, 88, 3–17. [Google Scholar] [CrossRef]
- Stohlgren, T.J.; Jarnevich, C.; Chong, G.W.; Evangelista, P.H. Scale and plant invasions: A theory of biotic acceptance. Preslia 2006, 78, 405–426. [Google Scholar]
- Thomaz, S.M.; Mormul, R.P.; Michelan, T.S. Propagule pressure, invasibility of freshwater ecosystems by macrophytes and their ecological impacts: A review of tropical freshwater ecosystems. Hydrobiologia 2015, 746, 39–59. [Google Scholar] [CrossRef]
- Patel, S. Threats, management and envisaged utilizations of aquatic weed Eichhornia crassipes: An overview. Rev. Environ. Sci. Bio./Technol. 2012, 11, 249–259. [Google Scholar] [CrossRef]
- Sánchez-Galván, G.; Monroy, O.; Gómez, J.; Olguín, E.J. Assessment of the hyperaccumulating lead capacity of Salvinia minima using bioadsorption and intracellular accumulation factors. Water Air Soil Pollut. 2008, 194, 77–90. [Google Scholar] [CrossRef]
- Nichols, P.B.; Couch, J.D.; Al-Hamdani, S.H. Selected physiological responses of Salvinia minima to different chromium concentrations. Aquat. Bot. 2000, 68, 313–319. [Google Scholar] [CrossRef]
- Alahuhta, J.; Lindholm, M.; Baastrup-Spohr, L.; García-Girón, J.; Toivanen, M.; Heino, J.; Murphy, K. Macroecology of macrophytes in the freshwater realm: Patterns, mechanisms and implications. Aquat. Bot. 2021, 168, 103325. [Google Scholar] [CrossRef]
- Lind, L.; Eckstein, R.L.; Relyea, R.A. Direct and indirect effects of climate change on distribution and community composition of macrophytes in lentic systems. Biol. Rev. 2022, 97, 1677–1690. [Google Scholar] [CrossRef]
- Hintz, W.D.; Schuler, M.S.; Borrelli, J.J.; Eichler, L.W.; Stoler, A.B.; Moriarty, V.W.; Ahrens, L.E.; Boylen, C.W.; Nierzwicki-Bauer, S.A.; Relyea, R.A. Concurrent improvement and deterioration of epilimnetic water quality in an oligotrophic lake over 37 years. Limnol. Oceanogr. 2020, 65, 927–938. [Google Scholar] [CrossRef]
- Gubiani, A.; Thomaz, S.M.; Bini, L.M.; Piana, P.A. Metapopulation models predict the temporal response of two macrophytes to drought in a subtropical water reservoir. Ecol. Eng. 2017, 100, 1–7. [Google Scholar] [CrossRef]
- Chaudhry, S.; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: A comprehensive review. Plant Cell Rep. 2021, 41, 1–31. [Google Scholar] [CrossRef]
- Battarbee, R.W.; Grytnes, J.A.; Thompson, R.; Appleby, P.G.; Catalan, J.; Korhola, A.; Birks, H.J.B.; Heegaard, E.; Lami, A. Comparing palaeolimnological and instrumental evidence of climate change for remote mountain lakes over the last 200 years. J. Paleolimnol. 2002, 28, 161–179. [Google Scholar] [CrossRef]
- Santamaría, L. Why are most aquatic plants widely distributed? Dispersal, clonal growth and small-scale heterogeneity in a stressful environment. Acta Oecologica 2002, 23, 137–154. [Google Scholar] [CrossRef]
- Tourinho, L.; Vale, M.M. Choosing among correlative, mechanistic and hybrid models of species’ niche and distribution. Integr. Zool. 2021, 18, 93–109. [Google Scholar] [CrossRef]
- Dandjlessa, J.; Ezin, B.; Zossou, N.; Ahanchede, A. Mapping actual distribution of invasive species siam weed and its abundance across biogeographical zones in Benin (West Africa). Adv. Weed Sci. 2022, 39, e20210081. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.S. Impactos do Aquecimento Global na Distribuição Espaço-Temporal do Tomateiro e do Inseto Praga Neoleucinodes elegantalis; Universidade Federal de Viçosa: Viçosa, Brazil, 2016. [Google Scholar]
- Jae-Min, J.U.; Sung-Hoon, J.U.; Byeon, D.H.; Wang-Hee, L.E. Prediction for potential distribution of yellow crazy ant (Anoplolepis gracilipes) in response to climate change scenario. Korean Biotechnol. Soc. 2016, 4, 313. [Google Scholar]
- Duque, T.S.; da Silva, R.S.; Maciel, J.C.; Silva, D.V.; Fernandes, B.C.C.; Júnior, A.P.B.; dos Santos, J.B. Potential Distribution of and Sensitivity Analysis for Urochloa panicoides Weed Using Modeling: An Implication of Invasion Risk Analysis for China and Europe. Plants 2022, 11, 1761. [Google Scholar] [CrossRef]
- Barroso, G.M.; da Silva, R.S.; Mucida, D.P.; Borges, C.E.; Ferreira, S.R.; dos Santos, J.C.B.; Lins, H.A.; Mendonça, V.; Silva, D.V.; dos Santos, J.B. Spatio-Temporal Distribution of Digitaria insularis: Risk Analysis of Areas with Potential for Selection of Glyphosate-Resistant Biotypes in Eucalyptus Crops in Brazil. Sustainability 2021, 13, 10405. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Brunel, S. Assessing and managing the current and future pest risk from water hyacinth, (Eichhornia crassipes), an invasive aquatic plant threatening the environment and water security. PLoS ONE 2016, 11, e0120054. [Google Scholar] [CrossRef]
- Araújo, F.H.V.; da Silva, A.F.; Ramos, R.S.; Ferreira, S.R.; dos Santos, J.B.; da Silva, R.S.; Shabani, F. Modelling climate suitability for Striga asiatica, a potential invasive weed of cereal crops. Crop Prot. 2022, 160, 106050. [Google Scholar] [CrossRef]
- Ferreira, S.R.; Dos Santos, J.C.B.; Ferreira, A. Análise de risco da invação biologica de Amaranthus palmeri no mundo frente às mudanças climáticas. In Proceedings of the Congresso Técnico Científico da Engenharia e da Agronomia—CONTECC, Virtual, 8 March 2021. [Google Scholar]
- Chen, Y.; Yu, J.; Khan, S. Spatial sensitivity analysis of multi-criteria weights in GIS-based land suitability evaluation. Environ. Model. Softw. 2010, 25, 1582–1591. [Google Scholar] [CrossRef]
- Tavana, M.; Soltanifar, M.; Santos-Arteaga, F.J. Analytical hierarchy process: Revolution and evolution. Ann. Oper. Res. 2021, 326, 879–907. [Google Scholar] [CrossRef]
- GBIF.org. Global Biodiversity Information Facility. Urochloa subquadripara (Trin.) R.D.Webster. Available online: https://www.gbif.org/occurrence/download/0427356-210914110416597 (accessed on 20 August 2022). [CrossRef]
- GBIF.org. Global Biodiversity Information Facility. Eichhornia crassipes (Mart.) Solms. Available online: https://www.gbif.org/occurrence/download/0427889-210914110416597 (accessed on 21 August 2022). [CrossRef]
- GBIF.org. Global Biodiversity Information Facility. Salvinia minima var. minima. Available online: https://www.gbif.org/occurrence/download/0427521-210914110416597 (accessed on 20 August 2022). [CrossRef]
- CABI. Invasive Species Compendium. Salvinia minima var. Minima. Available online: https://www.cabidigitallibrary.org/action/doSearch?AllField=Salvinia+minima (accessed on 20 August 2022).
- CABI. Invasive Species Compendium. Urochloa subquadripara (Trin.) R.D.Webster. Available online: https://www.cabi.org/isc/datasheet/55773 (accessed on 20 August 2022).
- Darren, K.; Gunter, M.; Tania, Y.; Eric, Z.; Neville, H.; Robert, S. Exploring the effects of climate on plants, animals and diseases. CLIMEX Version 2015, 4, 184. [Google Scholar]
- Shelford, V.E. The Ecology of North America; University of Illinois Press: Urbana, IL, USA, 2018; Volume 10, pp. 1237–1243. [Google Scholar]
- Teuton, T.C.; Brecke, B.J.; Unruh, J.B.; MacDonald, G.E.; Miller, G.L.; Ducar, J.T. Factors affecting seed germination of tropical signalgrass (Urochloa subquadripara). Weed Sci. 2004, 52, 376–381. [Google Scholar] [CrossRef]
- Nahdi, M.S.; Darsikin, D. Distribusi dan Kemelimpahan Spesies Tumbuhan Bawah pada Naungan Pinus mercusii, Acacia auriculiformis dan Eucalyptus alba di Hutan Gama Giri Mandiri, Yogyakarta. J. Nat. Indones. 2014, 16, 33–41. Available online: https://natur.ejournal.unri.ac.id/index.php/JN/article/view/2753/2696 (accessed on 13 August 2023). [CrossRef]
- St. Luce, M.; Gouveia, G.G.; Eudoxie, G.D. Comparative effects of food processing liquid slurry and inorganic fertilizers on tanner grass (Brachiaria arrecta) pasture: Grass yield, crude protein and P levels and residual soil N and P. Grass Forage Sci. 2017, 72, 401–413. Available online: https://doi-org.ez36.periodicos.capes.gov.br/10.1111/gfs.12240 (accessed on 13 August 2023). [CrossRef]
- Perozo-Bravo, A.; González, B.; Ortega-Alcalá, J. Efecto de la presión de pastoreo y la suplementación estratégica sobre la composición de la materia seca del pasto tanner (Brachiaria arrecta) antes y después del pastoreo. Rev. Fac. Agron. 2009, 26, 39–58. [Google Scholar]
- Jack, H.; Burke, J.L.; Cranston, L.; Morel, P.C.H.; Knights, M. The mineral content of some tropical forages commonly used in small ruminant production systems in the Caribbean–Part 2. Trop. Agric. 2020, 97, 7862. [Google Scholar]
- Galvão, A.; Silva, J.; Albertino, S.; Monteiro, G.; Calvalcante, D. Levantamento fitossociológico em pastagens de várzea no Estado do Amazonas. Planta Daninha 2011, 29, 69–75. [Google Scholar] [CrossRef]
- Botero, S.; Martínez, S. Análise Produtiva de Dois Modelos de Suplementação de Pastagem Para Mitigar o Impacto da Estação Seca no Gado de Engorda em Montería, Colômbia. Ph.D. Thesis, Zamorano, Escuela Agricola Panamericano, Tegucigalpa, Honduras, 2017. [Google Scholar]
- Evangelista, H.B.; Michelan, T.S.; Gomes, L.C.; Thomaz, S.M. Shade provided by riparian plants and biotic resistance by macrophytes reduce the establishment of an invasive Poaceae. J. Appl. Ecol. 2017, 54, 648–656. Available online: https://doi-org.ez36.periodicos.capes.gov.br/10.1111/1365-2664.12791 (accessed on 13 August 2023). [CrossRef]
- Penfound, W.T.; Earle, T.T. The biology of the water hyacinth. Ecol. Monogr. 1948, 18, 447–472. [Google Scholar] [CrossRef]
- Yan, S.H.; Song, W.; Guo, J.Y. Advances in management and utilization of invasive water hyacinth (Eichhornia crassipes) in aquatic ecosystems—A review. Crit. Rev. Biotechnol. 2017, 37, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Howard, G.W.; Harley, K.L.S. How do floating aquatic weeds affect wetland conservation and development? How can these effects be minimised? Wetl. Ecol. Manag. 1997, 5, 215–225. [Google Scholar] [CrossRef]
- Spencer, D.F.; Ksander, G.G. Seasonal growth of waterhyacinth in the Sacramento/San joaquin Delta, California. J. Aquat. Plant Manage. 2005, 43, 91. [Google Scholar]
- De Lacerda, L.P.; Lange, L.C.; França, M.G.C.; Leão, M.M.D. Growth and differential salinity reduction between Portulaca oleracea and Eichhornia crassipes in experimental hydroponic units. Environ. Technol. 2019, 40, 2267–2275. [Google Scholar] [CrossRef] [PubMed]
- Imchen, T.; Sawant, S.S.; Ezaz, W. Exposure of Eichhornia crassipes (Mart.) Solms to salt water and its implications. Curr. Sci. 2017, 113, 439–443. [Google Scholar] [CrossRef]
- Albano Pérez, E.; Ruiz Téllez, T.; Sánchez Guzmán, J.M. Influence of physico-chemical parameters of the aquatic medium on germination of Eichhornia crassipes seeds. Plant Biol. 2011, 13, 643–648. [Google Scholar] [CrossRef]
- Venter, N.; Cowie, B.W.; Witkowski, E.T.; Snow, G.C.; Byrne, M.J. The amphibious invader: Rooted water hyacinth’s morphological and physiological strategy to survive stranding and drought events. Aquat. Bot. 2017, 143, 41–48. [Google Scholar] [CrossRef]
- GBIF.org. Global Biodiversity Information Facility. Salvinia molesta D.S.Mitch. Available online: https://www.gbif.org/occurrence/download/0010552-230810091245214 (accessed on 13 August 2023). [CrossRef]
- Al-Hamdani, S.H.; Ghazal, J.J. Selected physiological responses of Salvinia minima to various temperatures and light intensities. Am. Fern J. 2009, 99, 155–161. [Google Scholar] [CrossRef]
- Ponce, S.C.; Prado, C.; Pagano, E.; Prado, F.E.; Rosa, M. Effect of solution pH on the dynamic of biosorption of Cr (VI) by living plants of Salvinia minima. Ecol. Eng. 2015, 74, 33–41. [Google Scholar] [CrossRef]
- Prado, C.; Ponce, S.C.; Pagano, E.; Prado, F.E.; Rosa, M. Differential physiological responses of two Salvinia species to hexavalent chromium at a glance. Aquat. Toxicol. 2016, 175, 213–221. [Google Scholar] [CrossRef]
- Room, P.M. Equations relating growth and uptake of nitrogen by Salvinia molesta to temperature and the availability of nitrogen. Aquat. Bot. 1986, 24, 43–59. [Google Scholar] [CrossRef]
- van der Heide, T.; Roijackers, R.M.; van Nes, E.H.; Peeters, E.T. A simple equation for describing the temperature dependent growth of free-floating macrophytes. Aquat. Bot. 2006, 84, 171–175. [Google Scholar] [CrossRef]
- Whiteman, J.B.; Room, P.M. Temperatures lethal to Salvinia molesta Mitchell. Aquat. Bot. 1991, 40, 27–35. [Google Scholar] [CrossRef]
- Oliver, J.D. A Review of the Biology of Giant Salvinia; Liberty University: Lynchburg, VA, USA, 1993. [Google Scholar]
- Paudel, S.; Milleville, A.; Battaglia, L.L. Responses of native and invasive floating aquatic plant communities to salinity and desiccation stress in the southeastern US coastal floodplain forests. Estuaries Coasts 2018, 41, 2331–2339. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Webber, B.L.; Leriche, A.; Ota, N.; Macadam, I.; Bathols, J.; Scott, J.K. CliMond: Global high-resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods Ecol. Evol. 2012, 3, 53–64. [Google Scholar] [CrossRef]
- Hossain, M.S.; Das, N.G. GIS-based multi-criteria evaluation to land suitability modelling for giant prawn (Macrobrachium rosenbergii) farming in Companigonj Upazila of Noakhali, Bangladesh. Comput. Electron. Agric. 2010, 70, 172–186. [Google Scholar] [CrossRef]
- Marinoni, O. Implementation of the analytical hierarchy process with VBA in ArcGIS. Comput. Geosci. 2004, 30, 637–646. Available online: https://doi-org.ez36.periodicos.capes.gov.br/10.1016/j.cageo.2004.03.010 (accessed on 13 August 2023). [CrossRef]
- Ohta, K.; Kobashi, G.; Takano, S.; Kagaya, S.; Yamada, H.; Minakami, H.; Yamamura, E. Analysis of the geographical accessibility of neurosurgical emergency hospitals in Sapporo city using GIS and AHP. Int. J. Geogr. Inf. Sci. 2007, 21, 687–698. [Google Scholar] [CrossRef]
- Marinoni, O.; Higgins, A.; Hajkowicz, S.; Collins, K. The multiple criteria analysis tool (MCAT): A new software tool to support environmental investment decision making. Environ. Model. Softw. 2009, 24, 153–164. [Google Scholar] [CrossRef]
- Saaty, T.L. A scaling method for priorities in hierarchical structures. J. Math. Psychol. 1977, 15, 234–281. [Google Scholar] [CrossRef]
- Saaty, T.L.; Vargas, L.G. Prediction, Projection and Forecasting: Applications of the Analytic Hierarchy Process in Economics, Finance, Politics, Games and Sports; Kluwer Academic Publishers: Boston, MA, USA, 1991. [Google Scholar]
- Kumar, M.; Shaikh, V.R. Site suitability analysis for urban development using GIS based multicriteria evaluation technique. J. Indian Soc. Remote Sens. 2013, 41, 417–424. Available online: https://doi-org.ez36.periodicos.capes.gov.br/10.1007/s12524-012-0221-8 (accessed on 13 August 2023). [CrossRef]
- Lehner, B.; Döll, P. Development and validation of a global database of lakes, reservoirs and wetlands. J. Hydrol. 2004, 296, 1–22. [Google Scholar] [CrossRef]
- Carniatto, N.; Cunha, E.R.; Thomaz, S.M.; Quirino, B.A.; Fugi, R. Feeding of fish inhabiting native and non-native macrophyte stands in a Neotropical reservoir. Hydrobiologia 2020, 847, 1553–1563. [Google Scholar] [CrossRef]
- Morrone, O.; Zuloaga, F.O. Novedades para la flora del nordeste de la Argentina. Hickenia 2006, 3, 9–29. [Google Scholar]
- Gaibor Litardo, J.R. Análisis de Fuentes Nitrogenadas en Los Pastos Brachiaria en el Cantón Babahoyo. Bachelor’s Thesis, Universidad Técnica de Babahoyo, Babahoyo, Ecuador, 2020. [Google Scholar]
- Herrera, A.M.; Martínez, N.; Herrera, P.; Colmenares, O.; Birbe, B. Consumo de heno y producción de leche en vacas doble propósito suplementadas con bloques multinutricionales. Zootec. Trop. 2009, 27, 465–473. [Google Scholar]
- Perozo-Bravo, A.; González, B.; Ortega-Alcalá, J.; Fuenmayor, A.; Pirela, M. Evaluación de la presión de pastoreo en tanner (Urochloa arrecta) y la suplementación estratégica en mautas mestizas en bosque húmedo tropical y suelos ácidos. Rev. Científica 2013, 23, 150–156. [Google Scholar]
- VonBank, J.A.; Casper, A.F.; Pendleton, J.E.; Hagy, H.M. Water hyacinth (Eichhornia crassipes) invasion and establishment in a temperate river system. River Res. Appl. 2018, 34, 1237–1243. [Google Scholar] [CrossRef]
- Gettys, L.A.; Thayer, K.L.; Sigmon, J.W. Phytotoxic Effects of Acetic Acid and d-limonene on Four Aquatic Plants. HortTechnology 2022, 32, 110–118. [Google Scholar] [CrossRef]
- Parys, K.A.; Johnson, S.J. Biological control of common Salvinia (Salvinia minima) in Louisiana using Cyrtobagous salviniae (Coleoptera: Curculionidae). Fla. Entomol. 2013, 96, 10–18. [Google Scholar] [CrossRef]
- Rowe, C.A.; Hauber, D.P.; Wolf, P.G. Genomic variation of introduced Salvinia minima in southeastern United States. Aquat. Bot. 2018, 151, 38–42. [Google Scholar] [CrossRef]
- Jacono, C.C.; Davern, T.R.; Center, T.D. The adventive status of Salvinia minima and S. molesta in the southern United States and the related distribution of the weevil Cyrtobagous salviniae. Castanea 2001, 66, 214–226. [Google Scholar]
- Murphy, K.; Efremov, A.; Davidson, T.A.; Molina-Navarro, E.; Fidanza, K.; Betiol, T.C.C.; Chambers, P.; Grimaldo, J.T.; Martins, S.V.; Springuel, I.; et al. World distribution, diversity and endemism of aquatic macrophytes. Aquat. Bot. 2019, 158, 103127. [Google Scholar] [CrossRef]
- Köppen, W.; Geiger, R. Klimate der Erde; Verlag Justus Perthes: Gotha, Germany, 1928. [Google Scholar]
- Fleming, J.P.; Dibble, E.D. Ecological mechanisms of invasion success in aquatic macrophytes. Hydrobiologia 2015, 746, 23–37. [Google Scholar] [CrossRef]
- Leal, R.P.; Silveira, M.J.; Petsch, D.K.; Mormul, R.P.; Thomaz, S.M. The success of an invasive Poaceae explained by drought resilience but not by higher competitive ability. Environ. Exp. Bot. 2022, 194, 104717. [Google Scholar] [CrossRef]
- Bianco, S.; Carvalho, L.B.; Bianco, M.S.; E Yamauchi, A.K.F. Crescimento e nutrição mineral de Urochloa arrecta. Planta Daninha 2015, 33, 33–40. [Google Scholar] [CrossRef]
- Lukács, B.A.; Sramkó, G.; Molnár, A. Plant diversity and conservation value of continental temporary pools. Biol. Conserv. 2013, 158, 393–400. [Google Scholar] [CrossRef]
- Lawlor, D.W. Genetic engineering to improve plant performance under drought: Physiological evaluation of achievements, limitations, and possibilities. J. Exp. Bot. 2013, 64, 83–108. [Google Scholar] [CrossRef]
- Villamagna, A.M.; Murphy, B.R. Ecological and socio-economic impacts of invasive water hyacinth (Eichhornia crassipes): A review. Freshw. Biol. 2010, 55, 282–298. [Google Scholar] [CrossRef]
- EPPO. European and mediterranean plant protection organization organisation europeenne et mediterraneenne pour la protection des plantes. Report of a Pest Risk Analysis: Eichhornia crassipes. OEPP/EPPO Bol. 2008, 38, 441–449. [Google Scholar] [CrossRef]
- Coetzee, J.A. Into Africa: Salvinia minima Baker (Salviniaceae) invades South Africa. BioInvasions Rec. 2022, 11, 1011–1018. [Google Scholar] [CrossRef]
- Tipping, P.W.; Martin, M.R.; Bauer, L.; Pierce, R.M.; Center, T.D. Ecology of common salvinia, Salvinia minima Baker, in southern Florida. Aquat. Bot. 2012, 102, 23–27. [Google Scholar] [CrossRef]
- Long, R.W.; Lakela, O. Flora of Tropical Florida; University of Miami Press: Oxford, OH, USA, 1971. [Google Scholar]
- Aziz, N.I.H.A.; Hanafiah, M.M.; Halim, N.H.; Fidri, P.A.S. Phytoremediation of TSS, NH3-N and COD from Sewage Wastewater by Lemna minor L., Salvinia minima, Ipomea aquatica and Centella asiatica. Appl. Sci. 2020, 10, 5397. [Google Scholar] [CrossRef]
- Prado, C.; Rodríguez-Montelongo, L.; González, J.A.; Pagano, E.A.; Hilal, M.; Prado, F.E. Uptake of chromium by Salvinia minima: Effect on plant growth, leaf respiration and carbohydrate metabolism. J. Hazard. Mater. 2010, 177, 546–553. [Google Scholar] [CrossRef]
- Castro-Longoria, E.; Trejo-Guillén, K.; Vilchis-Nestor, A.; Avalos-Borja, M.; Andrade-Canto, S.; Leal-Alvarado, D.; Santamaría, J. Biosynthesis of lead nanoparticles by the aquatic water fern, Salvinia minima Baker, when exposed to high lead concentration. Colloids Surf. B Biointerfaces 2014, 114, 277–283. [Google Scholar] [CrossRef]
- Iha, D.S.; Bianchini, I., Jr. Phytoremediation of Cd, Ni, Pb and Zn by Salvinia minima. Int. J. Phytoremediation 2015, 17, 929–935. [Google Scholar] [CrossRef]
- Guimarães, F.; Aguiar, R.; Karam, D.; Oliveira, J.; Silva, J.; Santos, C.; Sant’Anna-Santos, B.; Lizieri-Santos, C. Potential of macrophytes for removing atrazine from aqueous solution. Planta Daninha 2011, 29, 1137–1147. [Google Scholar] [CrossRef]
- Shabani, F.; Kumar, L.; Esmaeili, A. Future distributions of Fusarium oxysporum f. spp. in European, Middle Eastern and North African agricultural regions under climate change. Agric. Ecosyst. Environ. 2014, 197, 96–105. [Google Scholar] [CrossRef]
- IPCC 2018. Framing and Context. In: Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty. Available online: https://www.ipcc.ch/sr15/ (accessed on 22 December 2022).
- Hondzo, M.; Stefan, H.G. Three case studies of lake temperature and stratification response to warmer climate. Water Resour. Res. 1991, 27, 1837–1846. [Google Scholar] [CrossRef]
- Alahuhta, J.; Heino, J.; Luoto, M. Climate change and the future distributions of aquatic macrophytes across boreal catchments. J. Biogeogr. 2011, 38, 383–393. [Google Scholar] [CrossRef]
- Chandrasena, N. How will weed management change under climate change? Some perspectives. J. Crop Weed 2009, 5, 95–105. [Google Scholar]
- Maberly, S.C.; O’Donnell, R.A.; Woolway, R.I.; Cutler, M.E.J.; Gong, M.; Jones, I.D.; Merchant, C.J.; Miller, C.A.; Politi, E.; Scott, E.M.; et al. Global lake thermal regions shift under climate change. Nat. Commun. 2020, 11, 1232. [Google Scholar] [CrossRef]
- Bellard, C.; Thuiller, W.; Leroy, B.; Genovesi, P.; Bakkenes, M.; Courchamp, F. Will climate change promote future invasions? Glob. Change Biol. 2013, 19, 3740–3748. [Google Scholar] [CrossRef]
- Lacoul, P.; Freedman, B. Environmental influences on aquatic plants in freshwater ecosystems. Environ. Rev. 2006, 14, 89–136. [Google Scholar] [CrossRef]
- Kundzewicz, Z.W.; Mata, L.J.; Arnell, N.W.; Döll, P.; Jimenez, B.; Miller, K.; Oki, T.; Şen, Z.; Shiklomanov, I. The implications of projected climate change for freshwater resources and their management. Hydrol. Sci. J. 2008, 53, 3–10. [Google Scholar] [CrossRef]
- Johnson, P.T.J.; Olden, J.D.; Vander Zanden, M.J. Dam invaders: Impoudments facilitate biological invasions into freshwaters. Front. Ecol. Environ. 2008, 6, 357–363. [Google Scholar] [CrossRef]
- Ruhi, A.; Catford, J.A.; Cross, W.F.; Escoriza, D.; Olden, J.D. Understanding the nexus between hydrological alteration and biological invasions. In Multiple Stressors in River Ecosystems; Elsevier: Amsterdam, The Netherlands, 2019; pp. 45–64. [Google Scholar]
- Havel, J.E.; Lee, C.E.; Vander Zanden, J.M. Do reservoirs facilitate invasions into landscapes? BioScience 2005, 55, 518–525. [Google Scholar] [CrossRef]
- CEMIG. Macrófitas Aquáticas. Caracterização e Importância em Reservatórios Hidrelétricos. Available online: https://www.cemig.com.br/wp-content/uploads/2021/03/livro-macrofitas-cemig-2021.pdf (accessed on 28 September 2022).
- ANA. Catálogo de Metadados da Agência Nacional de Águas. Available online: https://metadados.snirh.gov.br/geonetwork/srv/api/records/56ac7cb5-aa22-4081-a135-c7fc0938a449 (accessed on 15 November 2022).
- Hill, S.R. Additions to the Bahama flora. SIDA Contrib. Bot. 1976, 6, 321–327. [Google Scholar]
- Vejarano, I. Evaluación de los Datos Reproductivos y Productivos de los Hatos Brahman y Criollo Yacumeño en Estancias Espíritu, Beni, Bolivia. Doctoral Dissertation, Escuela Agrícola Panamericana, Zamorano, Tegucigalpa, Honduras, 2014. [Google Scholar]
- Cerveira, W.R., Jr.; Silva, A.F.; Cruz, C.; Pitelli, R.A.; Martins, D. Herbicides for rooted aquatic macrophytes. Planta Daninha 2019, 37, e019175938. [Google Scholar] [CrossRef]
- Alves, R.M.A.; Albuquerque, M.B.; Barbosa, L.G. Status of the invasion of a Poaceae species in tropical semiarid reservoirs. Planta Daninha 2017, 35, e017153224. [Google Scholar] [CrossRef]
- Santini, R.; de Lima, J.P.; Gratão, P.L.; Camargo, A.F.M. Evaluation of growth and oxidative stress as indicative of salinity tolerance by the invasive tropical aquatic macrophyte tanner grass. Hydrobiologia 2022, 849, 1261–1271. [Google Scholar] [CrossRef]
- Sato, R.Y.; Costa, A.P.L.; Padial, A.A. The invasive tropical tanner grass decreases diversity of the native aquatic macrophyte community at two scales in a subtropical tidal river. Acta Bot. Bras. 2021, 35, 140–150. [Google Scholar] [CrossRef]
- Bottino, F.; Souza, B.P.; Rocha, R.J.S.; Cunha-Santino, M.B.; Bianchini, I., Jr. Characterization of humic substances from five macrophyte species decomposed under different nutrient conditions. Limnetica 2021, 40, 267–278. [Google Scholar] [CrossRef]
- Pitelli, R.L.C.M.; Pitelli-Merenda, A.M.C.M.; Pitelli, R.A.; Siqueira, R.C.; Barbosa, H.O.; Jesus, L. Composição específica e distribuição da comunidade de macrófitas aquáticas no reservatório de aimorés. Planta Daninha 2014, 32, 475–482. [Google Scholar] [CrossRef]
- Manuel, P.P.; Fanor, C.P.; Gerardo, A.T.R. Cadmium and lead toxicity in tanner grass Brachiaria arrecta. Rev. Cienc. Agrícolas 2014, 31, 3–13. [Google Scholar]
- Grajales, H.; Aguirre, N.J.; Palacio-Baena, J.A. Determinação da demanda de oxigênio da vegetação submersa na área de represamento do córrego La Vega (Antioquia), Colômbia. Biol. Updates 2006, 28, 75–85. [Google Scholar]
- Forero-Montaña, J.; Betancur, J.; Cavelier, J. Dieta del capibara Hydrochaeris hydrochaeris (rodentia: Hydrochaeridae) en Caño Limón, Arauca, Colombia. Rev. Biol. Trop. 2003, 51, 571–578. [Google Scholar]
- Montenegro, J.; Barrantes, E.; DiLorenzo, N. Methane emissions by beef cattle consuming hay of varying quality in the dry forest ecosystem of Costa Rica. Livest. Sci. 2016, 193, 45–50. [Google Scholar] [CrossRef]
- Pohl, R.W.; Davidse, G. Chromosome numbers of Costa Rican grasses. Brittonia 1971, 23, 293–324. [Google Scholar] [CrossRef]
- Viejo, C.; Stalin, M. Respuesta del pasto “Tanner”(Brachiaria arrecta) a diferentes niveles de nitrógeno en la zona de Babahoyo. Bachelor’s Thesis, UTB, Babahoyo, Ecuador, 2019. [Google Scholar]
- Gonzàlez, E.E.B.; Sánchez, E.; Jumbo, D.; Ochoa, H.R.C. Gramíneas y leguminosas promisorias para la alimentación del ganado en la Amazonía sur del Ecuador. Rev. Col. Méd. Vet. Estado Lara 2016, 6, 3. [Google Scholar]
- Cepeda Alarcón, L.G. Evaluación de Cuatro Densidades de Siembra de los Pastos Tanner (Brachiaria arrecta) y Janeiro (Eriochloa polystachya) para la Producción Bovina en la Zona Baja Inundable de Babahoyo. Bachelor’s Thesis, UTB, Babahoyo, Ecuador, 2016. [Google Scholar]
- González Velásquez, A.A.; Macías Intriago, C.X. Influencia de Actividades Agrícolas del Cultivo de Brachiaria arrecta y Brachiaria decumbens en la Calidad Físico-Química de Suelos, Comunidad Matapalo. Bachelor’s Thesis, ESPAM, Calceta, Ecuador, 2017. [Google Scholar]
- Manzo Sánchez, E.A. Análisis de la Eficiencia Agronómica del Nitrógeno en los Cultivos Forrajeros en el Ecuador. Bachelor’s Thesis, UTB, Babahoyo, Ecuador, 2020. [Google Scholar]
- Verloove, F. New records in vascular plants alien to Tenerife (Spain, Canary Islands). Biodivers. Data J. 2021, 9, e62878. [Google Scholar] [CrossRef]
- Otto, R.; Verloove, F. Nuevos xenófitos de La Palma (Islas Canarias, España), con énfasis en las especies naturalizadas y (potencialmente) invasoras. Parte 3. Collect. Bot. 2020, 39, e002. [Google Scholar] [CrossRef]
- Myster, R.W. Regeneration filters in post-agricultural fields of Puerto Rico and Ecuador. Plant Ecol. 2004, 172, 199–209. [Google Scholar] [CrossRef]
- Cross, R.B.; McCarty, L.B.; Estes, A.G. Postemergence tropical signalgrass (Urochloa subquadripara) control with nonorganic arsenical herbicides. Weed Technol. 2016, 30, 815–821. [Google Scholar] [CrossRef]
- Pearsaul, D.G.; Leon, R.G.; Sellers, B.A.; Silveira, M.L.; Odero, D.C. Evaluation of verticutting and herbicides for tropical signalgrass (Urochloa subquadripara) control in turf. Weed Technol. 2018, 32, 392–397. [Google Scholar] [CrossRef]
- Hatch, S.L. Urochloa subquadripara (Poaceae: Paniceae) new to Texas and a key to Urochloa of Texas. Phytoneuron 2010, 8, 1–4. [Google Scholar]
- Solomon, J.; Cumberbatch, N.; Austin, R.; Gonsalves, J.; Seaforth, E. The production parameters of the Barbados Blackbelly and crossbred sheep in a controlled semi-intensive system. Livest. Res. Rural. Dev. 2006, 18, 1–10. [Google Scholar]
- Miranda, J.L.; Osorio, J.L. Análisis de Gramíneas Tropicales y Simulación de Producción Potencial de Leche. Doctoral Dissertation, Escuela Agrícola Panamericana, Zamorano, Tegucigalpa, Honduras, 2012. [Google Scholar]
- Banerjee, D.K.; Pal, D.C. Some new distributional records in Indian grasses. Nelumbo 1970, 12, 70–72. [Google Scholar]
- Lija, M.; Kumar, M.B. Brachiaria subquadripara (Trin.) Hitch. (Poaceae): A New Record of Kerala. Trends Biosci. 2018, 11, 1371–1372. [Google Scholar]
- Njarui, D.M.; Gatheru, M.; Ghimire, S.R. Agro-morphological characterization of Urochloa grass accessions in Kenya. Trop. Grassl./Forrajes Trop. 2018, 7, 258. [Google Scholar] [CrossRef]
- Veldkamp, J.F. Brachiaria, Urochloa (Gramineae-Paniceae) in Malesia. Blumea Biodivers. Evol. Biogeogr. Plants 1996, 41, 413–437. [Google Scholar]
- González-Arcia, M.N.; Alonso-Díaz, M.Á.; Valles-de la Mora, B.; Castillo-Gallegos, E.; Ku-Vera, J.C. Balance de nitrógeno de toretes alimentados con Cratylia argentea y heno de Brachiaria arrecta. Ecosist. Recur. Agropecuários 2018, 5, 523–536. [Google Scholar]
- Klaassen, E.; Kwembeya, E. A checklist of Namibian indigenous and naturalised plants. Occas. Contrib. 2013, 5, 1–596. [Google Scholar]
- Miller Gil, L.; Fábrega Duque, J. Reuse of Treated Domestic Wastewater by Employing Artificial Wetlands in Panama. Air Soil Water Res. 2022, 15, 11786221221074401. [Google Scholar] [CrossRef]
- Dulshani, L.A.M.N.; Ranaweera, B.; Karunarathne, K.H.M.I. Propagation of Brachiaria subquadripara (Trin.) Hitchc. for establishment in national parks of Sri Lanka. In Proceedings of the 15th Agricultural Research Symposium, Kuliyapitiya, Sri Lanka, 28–29 June 2016. [Google Scholar]
- Bello-Faria, J.L.; Mora, R.E.; Herrera, A.M.; Acosta, B.; Chicco, C.F. Protein and energy supplementa-tion of Brahman heifers in the western plains of Venezuela. Proc. West. Sect. Amer. Soc. Anim. Sci. Miles City 2011, 62, 401–404. [Google Scholar]
- Hernández, M.J.; Guenni, O.; Gil, J.L. Acumulación de biomasa e intercepción y uso de luz en dos gramíneas forrajeras bajo un sistema silvopastoril en el estado Yaracuy, Venezuela. Rev. Fac. Agron 2007, 24, 342–347. [Google Scholar]
- Machado, S.A.; Oliveira, A.V.; Fabrin, T.M.C.; Prioli, S.M.A.P.; Prioli, A.J. Molecular characterization of the species Salvinia (Salviniaceae) from the upper Paraná River floodplain. Genet. Mol. Res. 2016, 15, 1–11. [Google Scholar] [CrossRef]
- Miranda, C.V.; Schwartsburd, P.B. Aquatic ferns from Viçosa (MG, Brazil): Salviniales (Filicopsida; Tracheophyta). Braz. J. Bot. 2016, 39, 935–942. [Google Scholar] [CrossRef]
- Coelho, F.F.; Basulto, S.R.; Fávaro, A.; Demetrio, G.R. Sex allocation in aquatic ferns: The role of body size. Plant Species Biol. 2021, 36, 425–435. [Google Scholar] [CrossRef]
- Bianchini, I., Jr.; Peret, A.M.; Cunha-Santino, M.B. A mesocosm study of aerobic mineralization of seven aquatic macrophytes. Aquat Bot. 2006, 85, 163–167. [Google Scholar]
- Flora da Argentina. Available online: http://conosur.floraargentina.edu.ar/species/details/22391 (accessed on 13 July 2020).
- Ronderos, M.M.; Marino, P.I.; Díaz, F.; Estévez, A.L. Biting midges (Diptera: Ceratopogonidae) from Martín García Island, Argentina. Rev. Biol. Trop. 2011, 59, 1183–1194. [Google Scholar] [CrossRef]
- Mahaye, N.; Thwala, M.; Musee, N. Interactions of Coated-Gold Engineered Nanoparticles with Aquatic Higher Plant Salvinia minima Baker. Nanomaterials 2011, 11, 3178. [Google Scholar] [CrossRef] [PubMed]
- Sachan, D.; Ghosh, A.; Das, G. Valorization of aquatic weed Salvinia minima to value-added eco-friendly biosorbent: Preferential removal of dye and heavy metal. Int. J. Environ. Sci. Technol. 2023, 20, 3703–3712. [Google Scholar] [CrossRef]
- Das, M.; Bramhanand, P.S.; Laxminarayana, K.; Roy Chowdhury, S. Effectiveness of common macrophytes for phytoremediation of hexavalent Cr prevalent in chromite mining areas. Int. J. Phytorem. 2022, 24, 787–795. [Google Scholar] [CrossRef]
- Rosati, L.; Fascetti, S.; Romano, V.A.; Potenza, G.; Lapenna, M.R.; Capano, A.; Stinca, A. New chorological data for the Italian vascular flora. Diversity 2020, 12, 22. [Google Scholar] [CrossRef]
- Maciel, S.; de Souza, M.G.C.; Pietrobom, M.R. Licófitas e monilófitas do Bosque Rodrigues Alves Jardim Botânico da Amazônia, município de Belém, estado do Pará, Brasil. Bol. Mus. Para. Emílio Goeldi-Ciên. Nat. 2007, 2, 69–83. [Google Scholar] [CrossRef]
- Leon, B.; Young, K.R. Aquatic plants of Peru: Diversity, distribution and conservation. Biodivers. Conserv. 1996, 5, 1169–1190. [Google Scholar] [CrossRef]
- Miranda, C.V. Salvinia (Salviniaceae) nas Regiões sul e Sudeste do Brasil. Master’s Thesis, Universidade Federal de Viçosa, Viçosa, Brazil, 2017. [Google Scholar]
- Pérez-Consuegra, N.; Cuervo-Gómez, A.; Martínez, C.; Montes, C.; Herrera, F.; Madriñán, S.; Jaramillo, C. Paleogene Salvinia (Salviniaceae) from Colombia and their paleobiogeographic implications. Rev. Palaeobot. Palynol. 2017, 246, 85–108. [Google Scholar] [CrossRef]
- Dharmono, D.; Mahruddin, M.; Irianti, R.; Fajeriadi, H. Aquatic plants as niche for lay eggs and raising juveniles by freshwater fish in three swamp habitats in South Kalimantan, Indonesia. Biodiversitas J. Biol. Divers. 2022, 23, d230341. [Google Scholar] [CrossRef]
- Jaramillo, M.L.C.; Figueroa, V.M.; Alvarado, D.J.D. Especies vegetales vasculares acuáticas de la laguna la tembladera, provincia El Oro, Ecuador. Rev. Metrop. Cienc. Apl. 2020, 3, 36–41. [Google Scholar]
- Rivadeneira, Z.K.A. Use of Eichhornia crassipes, Lemna sp and Salvinia minima Plant Scrubbers in the Decontamination of Wastewater of Livestock Origin, in the Province of Imbabura. Biomed. J. Sci. Tech. Res. 2021, 39, 31313–31319. [Google Scholar] [CrossRef]
- Ayala-Luna, M.; Pernía, B.; Cornejo, X. Determinación de la capacidad de remoción de cadmio por Salvinia auriculata Aubl. en agua contaminada. Rev. Cien. Cienc. Nat. Ambient. 2019, 13, 73–83. [Google Scholar] [CrossRef]
- Najila, N.; Anila, G. Heavy Metal Absorption and Phytoremediation Capacity of Macrophytes of Polachira Wetland of Kollam District, Kerala, India. Res. J. Chem. Environ. 2022, 26, 90–96. [Google Scholar]
- Panda, S.S.; Sahoo, K.; Rana, M.; Rout, N.C.; Dhal, N.K. Antimicrobial activities and phytochemical investigation of some native pteridophytes. Asian J. Pharm. Clin. Res. 2014, 7, 43–45. [Google Scholar]
- Panda, M.; Samal, R.N.; Bhatta, K.S.; Lenka, S.; Patra, H.K.; Nanda, S. Diversity and distribution of vascular macrophytes in Ansupa Lake, Odisha, India. Int. J. Bonorowo Wetl. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Leal-Alvarado, D.A.; Estrella-Maldonado, H.; Sáenz-Carbonell, L.; Ramírez-Prado, J.H.; Zapata-Pérez, O.; Santamaría, J.M. Genes coding for transporters showed a rapid and sharp increase in their expression in response to lead, in the aquatic fern (Salvinia minima Baker). Ecotoxicol. Environ. Saf. 2018, 147, 1056–1064. [Google Scholar] [CrossRef]
- Sota, E.R.; Pazos, L.A. Two cytotypes and a new hybrid in Salvinia seguier. Acta Amazon. 2001, 31, 557–557. [Google Scholar] [CrossRef]
- Dos Santos, V.V.; Leão Barros, I.C.; Moura Júnior, A.M.; Severi, W.; Magalhães, K.M. Samambaias aquáticas da bacia do rio de Contas, Bahia, Brasil. Neotrop. Biol. Conserv. 2014, 9, 42–48. [Google Scholar] [CrossRef]
- Xavier, L.R.C.C.; Araújo, T.O.; Nascimento, P.R.F.; Pereira, S.M.B. Floristic surveys of aquatic macrophytes in reservoirs in the Agreste zone of Pernambuco State, Brazil. Braz. J. Bot. 2012, 35, 313–318. [Google Scholar] [CrossRef]
- Maltchik, L.; Rolon, A.S.; Schott, P. Effects of hydrological variation on the aquatic plant community in a floodplain palustrine wetland of southern Brazil. Limnology 2007, 8, 23–28. [Google Scholar] [CrossRef]
- Goetz, M.N.B.; Dantas, Ê.W.; Barros, I.C.L. Influence of abiotic factors on the composition and abundance of aquatic ferns occurring in the state of Paraíba, Brazil. Aquat. Ecol. 2019, 53, 557–567. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Index | Unit. | US | EC | SM |
|---|---|---|---|---|---|
| Lower temperature threshold | DV0 | °C | 4 | 0.5 | 5 |
| Lower optimum temperature | DV1 | °C | 22 | 25 | 23 |
| Upper optimum temperature | DV2 | °C | 35 | 30 | 30 |
| Upper optimum threshold | DV3 | °C | 39 | 36 | 39 |
| Lower soil moisture threshold | SM0 | -- | 0 | 0 | 0.1 |
| Lower optimum soil moisture | SM1 | -- | 0.1 | 0.1 | 0.2 |
| Upper optimum soil moisture | SM2 | -- | 8 | 8 | 8 |
| Upper soil moisture threshold | SM3 | -- | 10 | 10 | 10 |
| Cold stress temperature threshold | TTCS | °C | 4 | 0.5 | 5 |
| Cold stress temperature rate | THCS | Week−1 | −0.001 | −0.0003 | −0.0003 |
| Cold stress degree-day threshold | DTCS | °C day | 4 | ---- | ---- |
| Cold stress degree-day rate | DHCS | Week−1 | −0.01 | ---- | ---- |
| Heat stress temperature threshold | TTHS | °C | 40 | 37 | 39 |
| Heat stress temperature rate | THHS | week−1 | 0.01 | 0.001 | 0.1 |
| Heat stress threshold | DTHS | °C dia | 39 | ---- | 35 |
| Heat stress degree-day rate | DHHS | week−1 | 0.01 | ---- | 0.1 |
| Dry stress threshold | SMDS | -- | 0.1 | 0.02 | ---- |
| Dry stress rate | HDS | week−1 | 0.005 | −0.005 | |
| Degree days per generation | PPD | °C dia | ---- | 1916 | ---- |
| Criteria | ||
|---|---|---|
| Criterion | Description | |
| Criterion 1 | Ecological niche for U. subquadripara | |
| Criterion 2 | Ecological niche for E. crassipes | |
| Criterion 3 | Ecological niche for S. mínima | |
| Criteria Classes | ||
| Description | Class | Normalized value |
| EI = 0 | 0 | 0 |
| 0 < EI < 30 | 1 | 0.5 |
| EI ≥ 30 | 2 | 1 |
| Criterion 1 | Criterion 2 | Criterion 3 | |
|---|---|---|---|
| Criterion 1 | 1 | 5 | 5 |
| Criterion 2 | 0.2 | 1 | 1 |
| Criterion 3 | 0.2 | 1 | 1 |
| Criterion | Description | Weight * |
|---|---|---|
| Criterion 1 | Ecological niche para U. subquadripara | 0.714 |
| Criterion 2 | Ecological niche para E. crassipes | 0.143 |
| Criterion 3 | Ecological niche para S. minima | 0.143 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duque, T.S.; Souza, I.M.; Mendes, D.S.; da Silva, R.S.; Mucida, D.P.; da Silva, F.D.; Silva, D.V.; dos Santos, J.B. Ecological Niche Modeling of Invasive Macrophyte (Urochloa subquadripara) and Co-Occurrence with South American Natives. Sustainability 2023, 15, 12722. https://doi.org/10.3390/su151712722
Duque TS, Souza IM, Mendes DS, da Silva RS, Mucida DP, da Silva FD, Silva DV, dos Santos JB. Ecological Niche Modeling of Invasive Macrophyte (Urochloa subquadripara) and Co-Occurrence with South American Natives. Sustainability. 2023; 15(17):12722. https://doi.org/10.3390/su151712722
Chicago/Turabian StyleDuque, Tayna Sousa, Iasmim Marcella Souza, Débora Sampaio Mendes, Ricardo Siqueira da Silva, Danielle Piuzana Mucida, Francisca Daniele da Silva, Daniel Valadão Silva, and José Barbosa dos Santos. 2023. "Ecological Niche Modeling of Invasive Macrophyte (Urochloa subquadripara) and Co-Occurrence with South American Natives" Sustainability 15, no. 17: 12722. https://doi.org/10.3390/su151712722