
Physiological and Biochemical Behaviours and Antioxidant Response of Helianthus annuus under Lanthanum and Cerium Stress


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Culture and REEs Treatment
2.2. Fluorescence Measurement
2.3. Photosynthetic Pigments Content
2.4. Endogenous Hydrogen Peroxide Content
2.5. Lipid Peroxidation
2.6. Protein Crude Extract Preparation and Enzymatic Activity Determination
2.7. Determination of REEs Contents
2.8. Statistical Analysis
3. Results
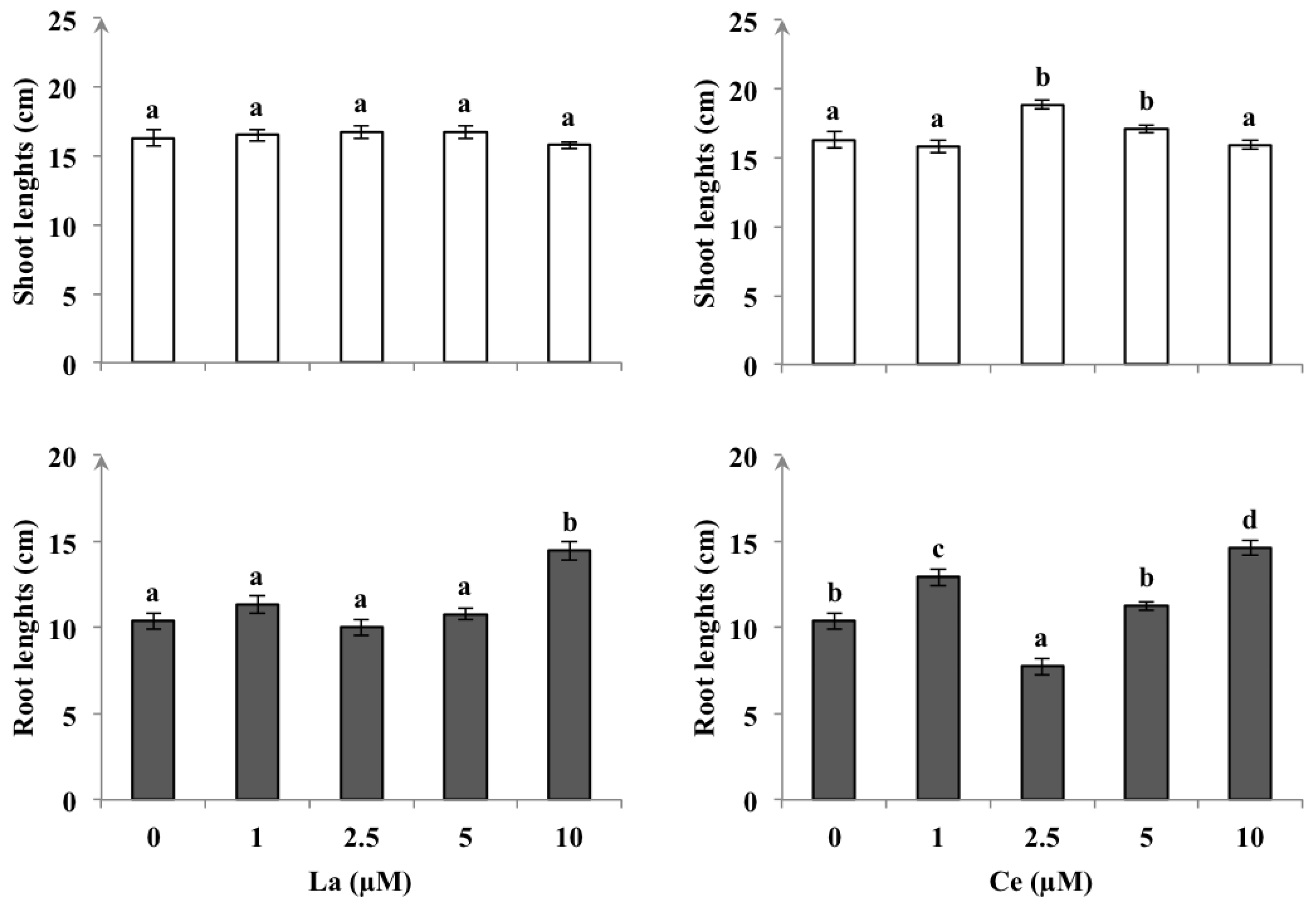
3.1. Plant Growth
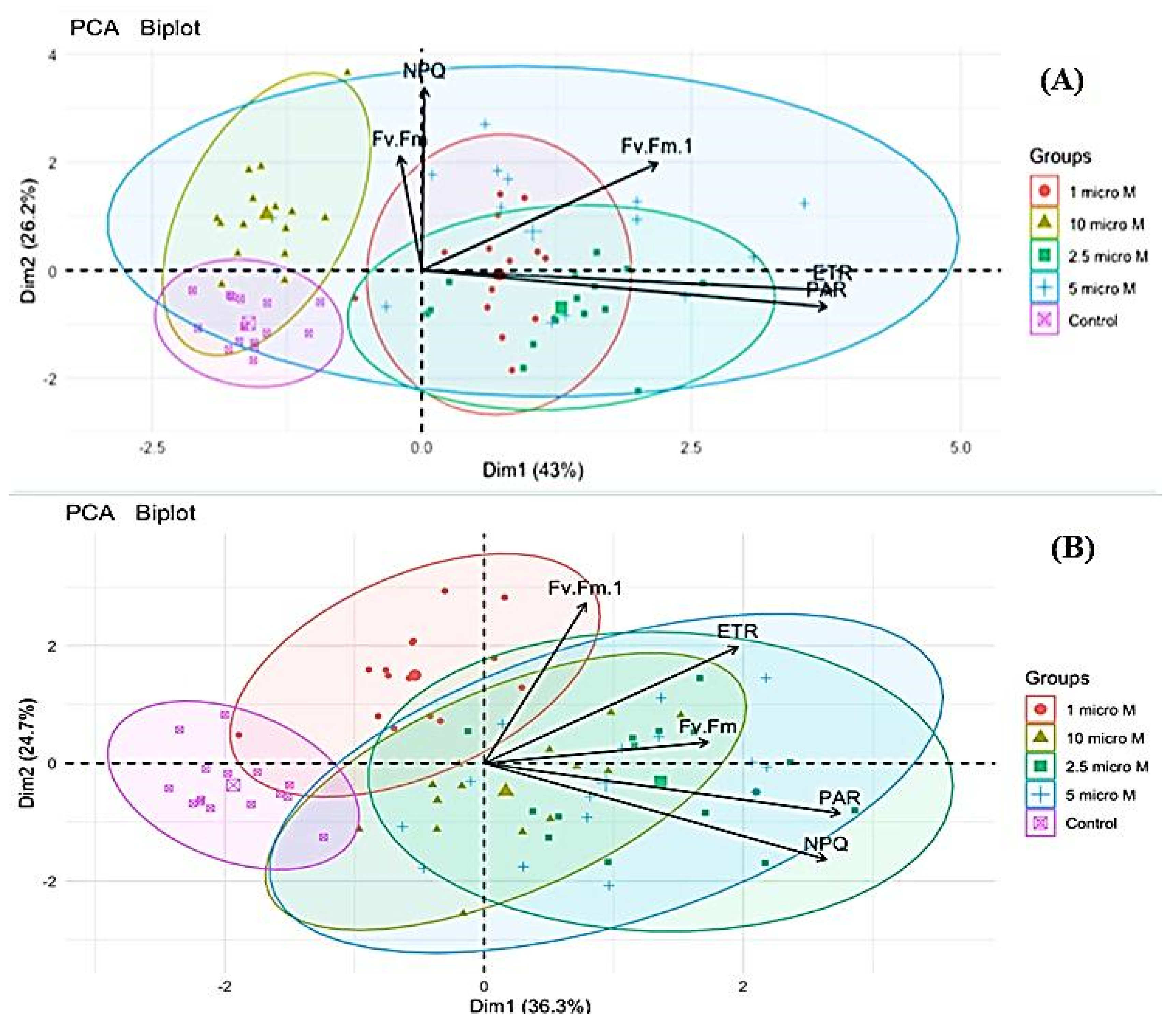
3.2. Chlorophyll Fluorescence Response
Principal Component Analysis
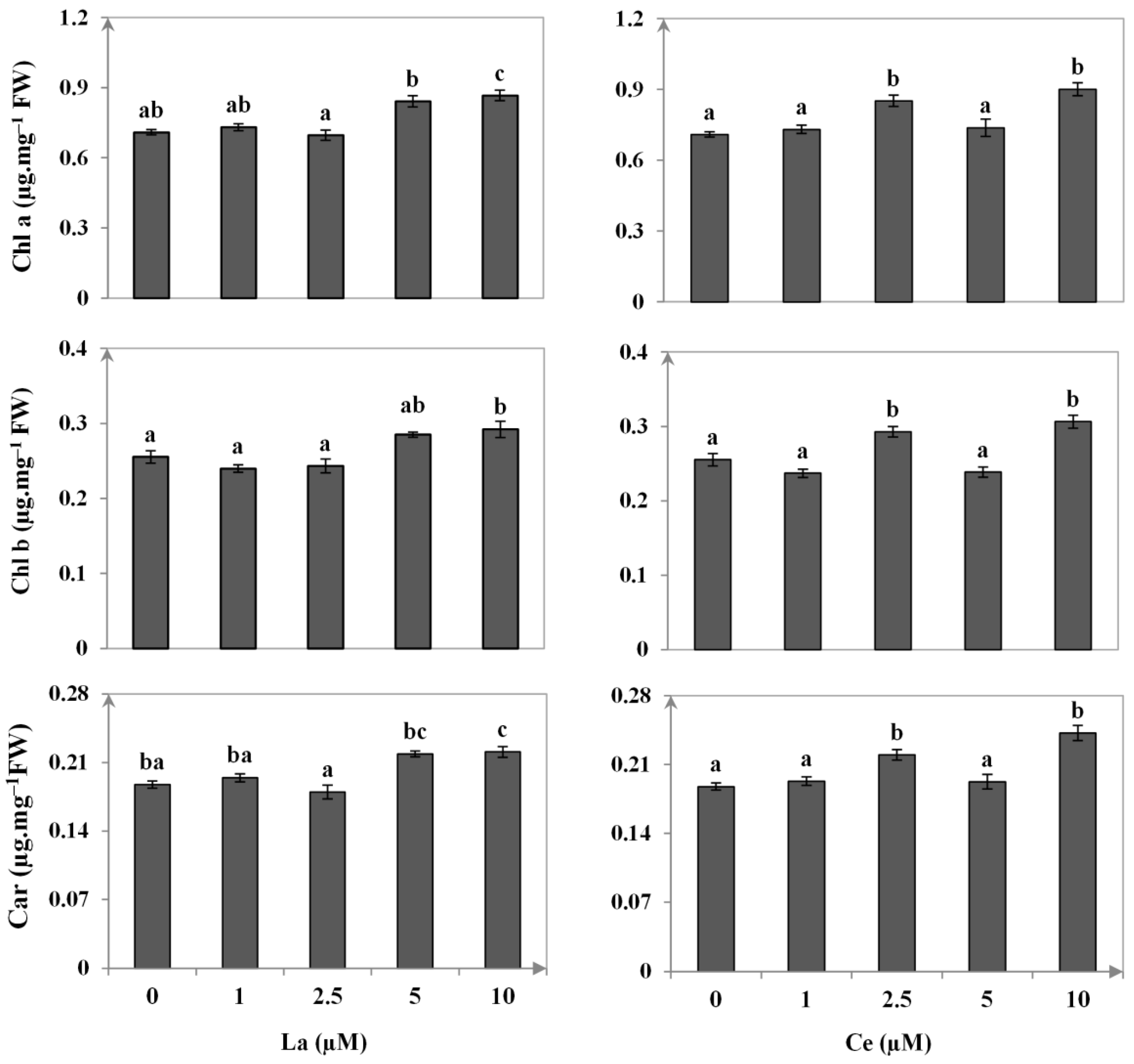
3.3. Photosynthetic Pigments Contents
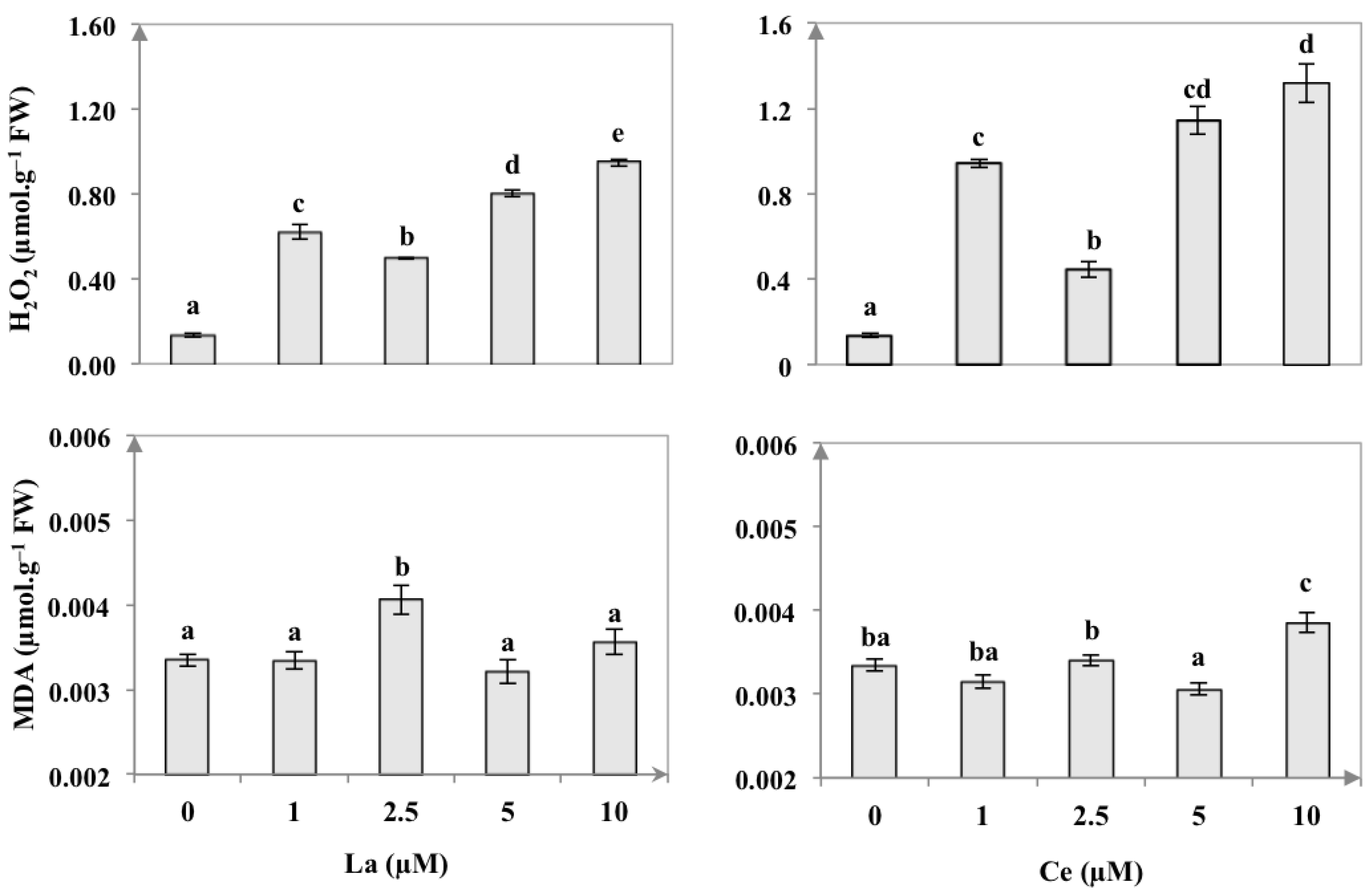
3.4. Endogenous H2O2 Content and Lipid Peroxidation
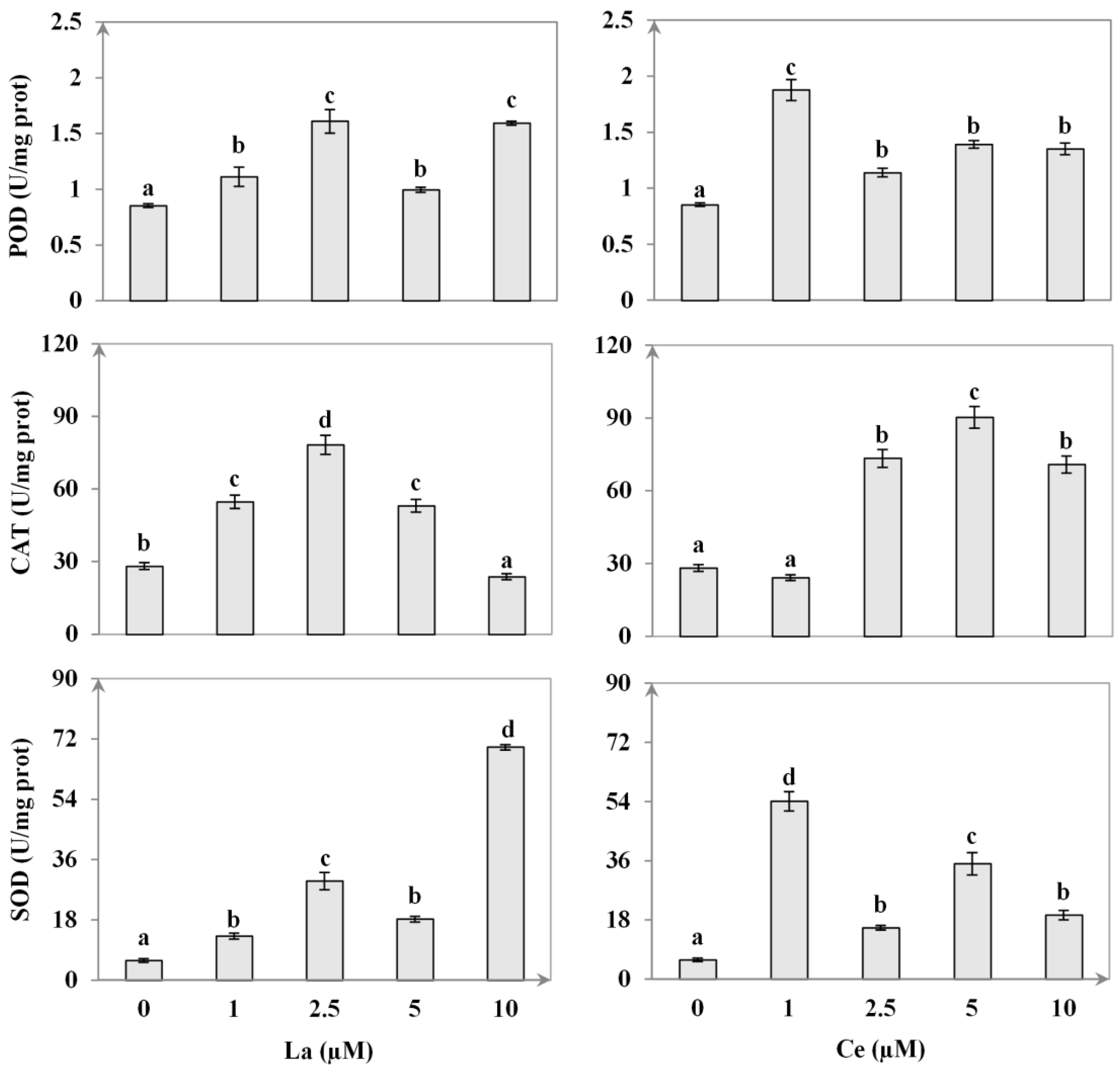
3.5. Enzymatic Activity
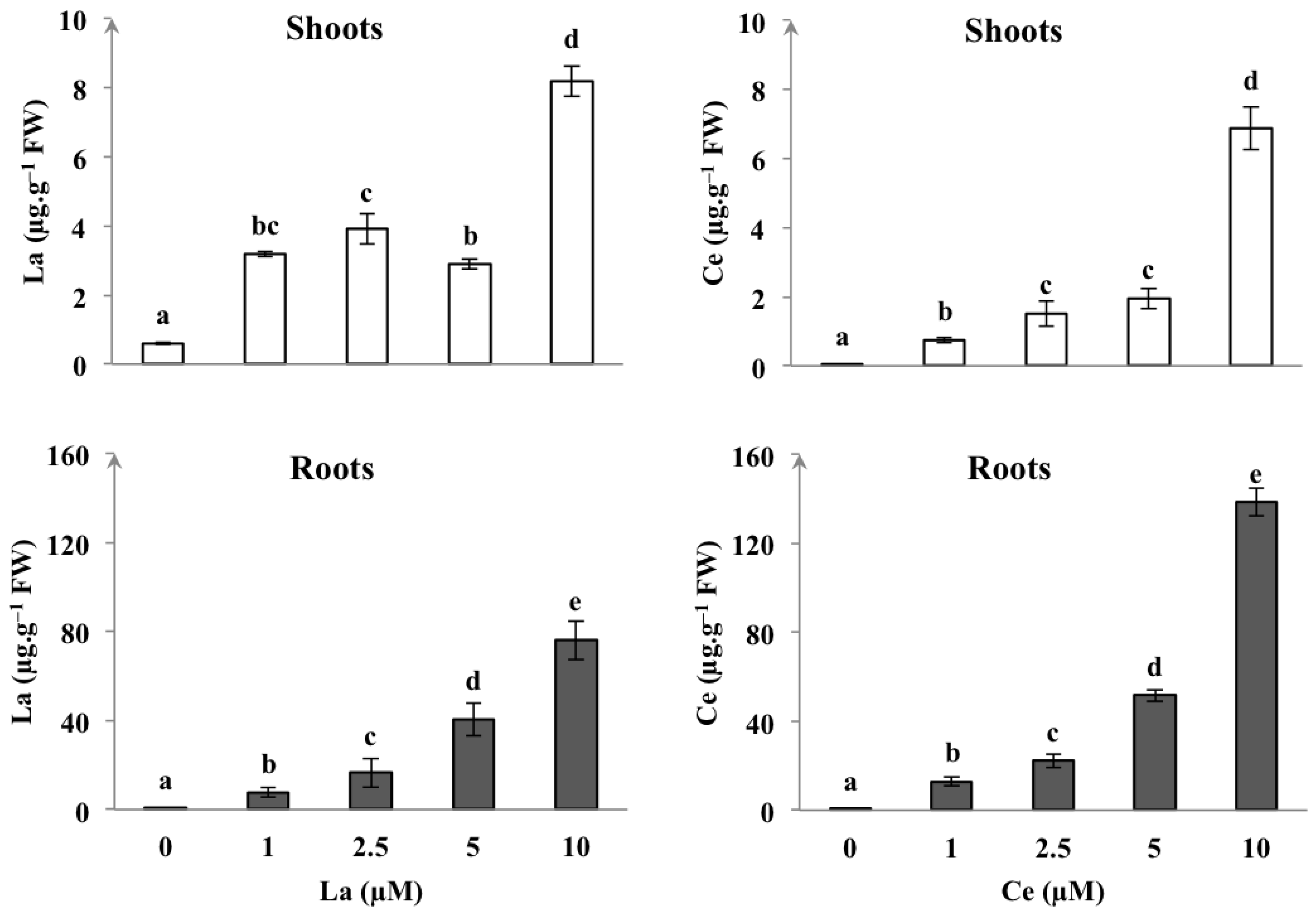
3.6. REEs Contents
4. Discussion
4.1. REEs Influence on Plant Growth
4.2. Chlorophyll Fluorescence and Pigment Content
4.3. Accumulation of H2O2 and MDA
4.4. Antioxidant Enzymes Response
4.5. REEs Accumulation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ippolito, M.P.; Fasciano, C.; d’Aquino, L.; Morgana, M.; Tommasi, F. Responses of antioxidant systems after exposition to rare earths and their role in chilling stress in commosn duckweed (Lemna minor L.): A Defensive Weapon or a Boomerang? Arch. Environ. Contam. Toxicol. 2010, 58, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Grosjean, N.; Jean, M.L.; Berthelot, C.; Chalot, M.; Gross, E.M.; Blaudez, D. Accumulation and fractionation of rare earth elements are conserved traits in the Phytolacca genus. Sci. Rep. 2019, 9, 18458. [Google Scholar] [CrossRef] [Green Version]
- Elbasan, F.; Ozfidan-Konakci, C.; Yildiztugay, E.; Kucukoduk, M. Rare-earth element scandium improves stomatal regulation and enhances salt and drought stress tolerance by up-regulating antioxidant responses of Oryza sativa. Plant Physiol. Biochem. 2020, 152, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Tyler, G. Rare earth elements in soil and plant systems—A review. Plant Soil 2004, 267, 191–206. [Google Scholar] [CrossRef]
- Balaram, V. Rare earth elements: A review of applications, occurrence, exploration, analysis, recycling, and environmental impact. Geosci. Front. 2019, 10, 1285–1303. [Google Scholar] [CrossRef]
- Li, X.X.; Yu, B.; Dong, Y.Y.; Wang, L.S.; Zhang, S.L.; Shangguan, H.Y.; He, Z.H.; Luo, X.M.; Lai, P.F. Lanthanum chloride enhances the photosynthetic characteristics and increases konjac glucomannan contents in Amorphophallus sinensis Belval. Photosynthetica 2020, 58, 165–173. [Google Scholar] [CrossRef]
- Wang, L.; Wang, W.; Zhou, Q.; Huang, X. Combined effects of lanthanum (III) chloride and acid rain on photosynthetic parameters in rice. Chemosphere 2014, 112, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Li, Y.; Wang, L.; Zhou, Q.; Huang, X. Effect of lanthanum (III) on the production of ethylene and reactive oxygen species in soybean seedlings exposed to the enhanced ultraviolet-B radiation. Ecotoxicol. Environ. Saf. 2014, 104, 152–159. [Google Scholar] [CrossRef]
- Duarte, A.C.O.; De Oliveira, C.; Ramos, S.J.; De Castro, E.M.; Siqueira, J.O.; Guilherme, L.R.G. Lanthanum content and effects on growth, gas exchanges, and chlorophyll index in maize plants. Acta Sci. Biol. Sci. 2018, 40, e38469. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Stowers, C.; Rossi, L.; Zhang, W.; Lombardini, L.; Ma, X. Physiological effects of cerium oxide nanoparticles on the photosynthesis and water use efficiency of soybean (Glycine max (L.) Merr.). Environ. Sci. Nano J. 2017, 4, 1086. [Google Scholar] [CrossRef]
- Wang, L.; Li, J.; Zhou, Q.; Yang, G.; Ding, X.L.; Li, X.; Cai, C.X.; Zhang, Z.; Wei, H.Y.; Lu, T.H.; et al. Rare earth elements activate endocytosis in plant cells. Proc. Natl. Acad. Sci. USA 2014, 111, 12936–12941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Wang, X.; Lin, Y.; Chen, Z.; Xu, H.; Wang, L. The effects of cerium on the growth and some antioxydant metabolisms in rice seedlings. Environ. Sci. Pollut. Res. 2012, 19, 3282–3291. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, L.; Li, Y.; Sun, J.; Zhou, Q.; Huang, X. Insight into mechanism of lanthanum (III) induced damage to plant photosynthesis. Ecotoxicol. Environ. Saf. 2016, 127, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Bankaji, I.; Pérez-Clemente, R.M.; Caçador, I.; Sleimi, N. Accumulation potential of Atriplex halimus to zinc and lead combined with NaCl: Effects on physiological parameters and antioxidant enzymes activities. S. Afr. J. Bot. 2019, 123, 51–61. [Google Scholar] [CrossRef]
- D’Aquino, L.; de-Pinto, M.C.; Nardi, L.; Morgana, M.; Tommasi, F. Effect of some light rare earth elements on seed germination, seedling growth and antioxidant metabolism in Triticum durum. Chemosphere 2009, 75, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Ding, Z.; Chen, Y.; Wang, X.; Dai, L. Bioaccumulation of lanthanum and cerium and their effects on the growth of wheat (Triticum aestivum L.) seedlings. Chemosphere 2002, 48, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, L.; Zhou, Q.; Huang, X. Combined effects of simulated acid rain and lanthanum chloride on chloroplast structure and functional elements in rice. Environ. Sci. Pollut. Res. 2016, 23, 8902–8916. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.P.; Shan, C.J.; Wei, A.Z.; Yang, T.; Sa, W.Q.; Feng, B. Leaf senescence and photosynthesis in foxtail millet [Setariaitalica (L.) P. Beauv] varieties exposed to drought conditions. Aust. J. Crop Sci. 2012, 6, 232–237. [Google Scholar]
- Bouslimi, H.; Ferreira, R.; Dridi, N.; Brito, P.; Martins-Dias, S.; Caçador, I.; Sleimi, N. Effects of Barium stress in Brassica juncea and Cakile maritima: The indicator role of some antioxidant enzymes and secondary metabolites. Phyton 2021, 90, 145. [Google Scholar] [CrossRef]
- Bhaduri, A.M.; Fulekar, M.H. Antioxidant enzyme responses of plants to heavy metal stress: Review. Rev. Environ. Sci. Biotechnol. 2012, 11, 55–69. [Google Scholar] [CrossRef]
- Berni, R.; Luyckx, M.; Xu, X.; Legay, S.; Sergeant, K.; Hausman, J.F.; Lutts, S.; Caia, G.; Guerrierod, G. Reactive oxygen species and heavy metal stress in plants: Impact on the cell wall and secondary metabolism. Environ. Exp. Bot. 2018, 161, 98–106. [Google Scholar] [CrossRef]
- Sat, I.G. The effect of heavy metals on peroxidase from Jerusalem artichoke (Helianthus tuberosus L.) tubers. Afr. J. Biotechnol. 2008, 7, 2248–2253. [Google Scholar]
- Nas, F.S.; Ali, M. The effect of lead on plants in terms of growing and biochemical parameters: A review. MOJ Ecol. Environ. Sci. 2018, 3, 265–268. [Google Scholar]
- Kovaříková, M.; Tomášková, I.; Soudek, P. Rare earth elements in plants: Review. Biol. Plant. 2019, 63, 20–32. [Google Scholar] [CrossRef]
- Xie, Y.; Cai, X.; Liu, W.; Tao, G.; Chen, Q.; Zhang, Q. Effects of lanthanum nitrate on growth and chlorophyll fluorescence characteristics of Alternanthera philoxeroides underperchlorate stress. J. Rare Earths 2013, 31, 823. [Google Scholar] [CrossRef]
- Stirbet, A. Govindjee On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient: Short review. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.A.; Duarte, J.G.; Vergine, P.; Antunes, C.D.; Freire, F.; Martins-Dias, S. Phragmites sp. physiological changes in a constructed wetland treating an effluent contaminated with a diazo dye (DR81). Environ. Sci. Pollut. Res. 2014, 21, 9626–9643. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Bouazizi, H.; Jouili, H.; Geitmann, A.; El-Ferjani, E. Copper toxicity in expanding leaves of Phaseolus vulgaris L.: Antioxidant enzyme response and nutrient element uptake. Ecotoxicol. Environ. Saf. 2010, 73, 1304–1308. [Google Scholar] [CrossRef]
- Velikova, V.; Loreto, F. On the relationship between isoprene emission and thermo-tolerance in Phragmites australis leaves exposed to high temperatures and during the recovery from a heat stress. Plant Cell Environ. 2005, 28, 318–327. [Google Scholar] [CrossRef]
- Davis, D.G.; Swanson, H.R. Activity of stress-related enzymes in the perennial weed leafy spurge (Euphorbia esula L.). Environ. Exp. Bot. 2001, 46, 95–108. [Google Scholar] [CrossRef]
- Brito, P.; Ferreira, R.A.; Martins-Dias, S.; Azevedo, O.M.; Caetano, M.; Caçador, I. Cerium uptake, translocation and toxicity in the salt marsh halophyte Halimione portulacoides (L.), Aellen. Chemosphere 2021, 266, 128973. [Google Scholar] [CrossRef]
- Ferreira, R.A.; Martins-Dias, S. Purification of plant complex protein extracts in non-denaturing conditions by in-solution isoelectric focusing. Anal. Biochem. 2016, 509, 100–103. [Google Scholar] [CrossRef]
- Labidi, O.; Vives-Peris, V.; Gómez-Cadenas, A.; Pérez-Clemente, R.M.; Sleimi, N. Assessing of growth, antioxidant enzymes, and phytohormone regulation in Cucurbita pepo under cadmium stress. Food Sci. Nutr. 2021, 9, 2021–2031. [Google Scholar] [CrossRef] [PubMed]
- Sleimi, N.; Bankaji, I.; Kouki, R.; Dridi, N.; Duarte, B.; Caçador, I. Assessment of extraction methods of trace metallic elements in plants: Approval of a common method. Sustainability 2022, 14, 1428. [Google Scholar] [CrossRef]
- Vilela, L.A.F.; Ramos, S.J.; Carneiro, M.A.C.; Faquin, V.; Guilherme, L.R.G.; Siqueira, J.O. Cerium (Ce) and Lanthanum (La) promoted plant growth and mycorrhizal colonization of maize in tropical soil. Aust. J. Crop Sci. 2018, 12, 704–710. [Google Scholar] [CrossRef]
- Huang, G.R.; Wang, L.H.; Zhou, Q. Combined effects of Lanthanum (III) and elevated ultraviolet-B radiation on root growth and ion absorption in soybean seedlings. Environ. Sci. Pollut. Res. 2013, 21, 3621–3633. [Google Scholar] [CrossRef] [PubMed]
- Kotelnikova, A.; Fastovets, I.; Rogova, O.; Volkov, D.S.; Stolbova, V. Toxicity assay of lanthanum and cerium in solutions and soil. Ecotoxicol. Environ. Saf. 2019, 167, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, K.; He, M.; Jiang, C.; Tian, L.; Tian, Y.; Wang, X. Mineral nutrient imbalance, DNA lesion and DNA-protein crosslink involved in growth retardation of Vicia faba L. seedlings exposed to lanthanum ions. J. Environ. Sci. 2012, 24, 214–220. [Google Scholar] [CrossRef]
- Wu, M.; Wang, P.Y.; Sun, L.G.; Zhang, J.J.; Yu, J.; Wang, Y.W.; Chen, G.X. Alleviation of cadmium toxicity by cerium in rice seedlings is related to improved photosynthesis, elevated antioxidant enzymes and decreased oxidative stress. Plant Growth Regul. 2014, 74, 251–260. [Google Scholar] [CrossRef]
- Emmanuel, E.S.C.; Anandkumar, B.; Natesan, M.; Maruthamuthu, S. Efficacy of rare earth elements on the physiological and biochemical characteristics of Zea mays L. Aust. J. Crop Sci. 2010, 4, 289–294. [Google Scholar]
- Sanchez-Zabala, J.; González-Murua, C.; Marino, D. Mild ammonium stress increases chlorophyll content in Arabidopsis thaliana. Plant Signal. Behav. 2015, 10, e991596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caçador, I.; Duarte, B.; Marques, J.C.; Sleimi, N. Carbon mitigation: A salt marsh ecosystem service in times of change. In Halophytes for Food Security in Dry Landss; Khan, M.A., Ozturk, M., Gul, B., Ahmed, M.Z., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2016; pp. 83–110. [Google Scholar] [CrossRef]
- Agathokleous, E.; Feng, Z.Z.; Peñuelas, J. Chlorophyll hormesis: Are chlorophylls major components of stress biology in higher plants? Sci. Total Environ. 2020, 726, 138637. [Google Scholar] [CrossRef]
- Saldaña-Sánchez, W.D.; León-Morales, J.M.; López-Bibiano, Y.; Hernández-Hernández, M.; Langarica-Velázquez, E.C.; García-Morales, S. Effect of V, Se, and Ce on Growth, Photosynthetic Pigments, and Total Phenol Content of Tomato and Pepper Seedlings. J. Soil Sci. Plant Nutr. 2019, 19, 678–688. [Google Scholar] [CrossRef]
- Gao, Y.; Huang, W.; Zhu, L.; Chen, J. Effects of LaCl3 on the growth and photosynthetic characteristics of Fny-infected tobacco seedlings. J. Rare Earths 2012, 30, 725. [Google Scholar] [CrossRef]
- Lambrev, P.H.; Miloslavina, Y.; Jahns, P.; Holzwarth, A.R. On the relationship between non-photochemical quenching and photoprotection of Photosystem II. Biochim. Biophys. Acta 2012, 1817, 760–769. [Google Scholar] [CrossRef] [Green Version]
- Cheeseman, J.M. Hydrogen Peroxide and Plant Stress: A Challenging Relationship. Plant Stress 2007, 1, 4–15. [Google Scholar]
- Zicari, M.A.; d’Aquino, L.; Paradiso, A.; Mastrolitti, S.; Tommasi, F. Effect of cerium on growth and antioxidant metabolism of Lemna minor L. Ecotoxicol. Environ. Saf. 2018, 163, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Wang, W. Antioxidant response of soybean seedlings to joint stress of lanthanum and acid rain. Environ. Sci. Pollut. Res. 2013, 20, 8182–8191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, T.; Lu, Q.; Cai, S.; Chu, W.; Qiu, H.; Xu, T.; Li, F.; Xu, Q. Oxidative effects, nutrients and metabolic changes in aquatic macrophyte, Elodea nuttallii following exposure to lanthanum. Ecotoxicol. Environ. Saf. 2015, 115, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Hossain, M.A.; Teixeira-da-Silva, J.A.; Fujita, M. Plant Response and Tolerance to Abiotic Oxidative Stress: Antioxidant Defense Is a Key Factor. In Crop Stress and Its Management: Perspectives and Strategies; Springer: Dordrecht, The Netherlands, 2012; pp. 261–315. [Google Scholar] [CrossRef]
- Sleimi, N.; Kouki, R.; Hadj-Ammar, M.; Ferreira, R.; Pérez-Clemente, R. Barium effect on germination, plant growth, and antioxidant enzymes in Cucumis sativus L. plants. Food Sci. Nutr. 2021, 9, 2086–2094. [Google Scholar] [CrossRef] [PubMed]
- Malar, S.; Shivendra-Vikram, S.; Favas, P.J.C.; Perumal, V. Lead heavy metal toxicity induced changes on growth and antioxidative enzymes level in water hyacinths [Eichhornia crassipes (Mart.)]. Bot. Stud. 2016, 55, 54. [Google Scholar] [CrossRef] [Green Version]
- Thakur, S.; Singh, L.; Zularisam, A.W.; Sakinah, M.; Din, M.F.M. Lead induced oxidative stress and alteration in the activities of antioxidative enzymes in rice shoots. Plant Biol. 2017, 61, 595–598. [Google Scholar] [CrossRef]
- Nigam, N.; Khare, P.; Yadav, V.; Mishra, D.; Jain, S.; Karak, T.; Panja, S.; Tandon, S. Biochar-mediated sequestration of Pb and Cd leads to enhanced productivity in Mentha arvensis. Ecotoxicol. Environ. Saf. 2019, 172, 411–422. [Google Scholar] [CrossRef]
- Wang, X.; Shi, G.X.; Xu, Q.S.; Xu, B.J.; Zhao, J. Lanthanum and cerium induced oxidative stress in submerged Hydrilla verticillata plants. Russ. J. Plant Physiol. 2007, 54, 693–697. [Google Scholar] [CrossRef]
- Xu, Q.M.; Chen, H. Antioxidant responses of rice seedling to Ce (4+) under hydroponic cultures. Ecotoxicol. Environ. Saf. 2011, 74, 1693–1699. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, P.; Zhang, Z.; He, X.; Li, Y.; Zhang, J.; Zheng, L.; Chu, S.; Yang, K.; Zhao, Y.; et al. Origin of the different phytotoxicity and biotransformation of cerium and lanthanum oxide nanoparticles in cucumber. Nanotoxicology 2015, 9, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Rezaee, A.; Hale, B.; Santos, R.M.; Chiang, Y.W. Accumulation and Toxicity of Lanthanum and Neodymium in Horticultural Plants (Brassica chinensis L. and Helianthus annuus L.). Can. J. Chem. Eng. 2017, 96, 2263–2272. [Google Scholar] [CrossRef]
- Sasmaz, M.; Obek, E.; Sasmaz, A. The accumulation of La, Ce and Y by Lemna minor and Lemna gibba in the Keban gallery water, Elazig Turkey. Water Environ. J. 2018, 32, 75–83. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Fv/Fm | Fv’/Fm’ | NPQ | ETR | PAR | Chl a/Chl b | (Chl a + Chl b)/Car |
|---|---|---|---|---|---|---|---|
| La (µM) | |||||||
| 0 | 0.64 a ± 0.02 | 0.83 a ± 0.01 | 0.73 a ± 0.02 | 0.70 a ± 0.00 | 2.0 a ± 0.00 | 2.80 ± 0.07 | 5.14 ± 0.06 |
| 1 | 0.70 ab ± 0.02 | 0.85 a ± 0.01 | 0.94 b ± 0.06 | 1.35 b ± 0.04 | 3.53 b ± 0.13 | 3.05 ± 0.04 | 5.00 ± 0.05 |
| 2.5 | 0.69 ab ± 0.01 | 0.86 a ± 0.01 | 0.76 a ± 0.04 | 1.45 b ± 0.06 | 4.0 c ± 0.17 | 2.87 ± 0.03 | 5.24 ± 0.06 |
| 5 | 0.71 ab ± 0.02 | 0.88 a ± 0.01 | 1.05 b ± 0.05 | 1.35 b ± 0.09 | 3.53 b ± 0.26 | 2.94 ± 0.08 | 5.12 ± 0.05 |
| 10 | 0.74 b ± 0.02 | 0.84 a ± 0.01 | 1.14 b ± 0.06 | 0.70 a ± 0.00 | 2.0 a ± 0.00 | 2.98 ± 0.03 | 5.24 ± 0.07 |
| Ce (µM) | |||||||
| 0 | 0.64 a ± 0.02 | 0.83 a ± 0.01 | 0.73 a ± 0.02 | 0.70 a ± 0.00 | 2.0 a ± 0.00 | 2.80 ± 0.07 | 5.14 ± 0.06 |
| 1 | 0.72 ab ± 0.02 | 0.88 a ± 0.01 | 0.75 a ± 0.03 | 1.38 c ± 0.06 | 4.0 c ± 0.2 | 3.08 ± 0.04 | 5.01 ± 0.03 |
| 2.5 | 0.79 b ± 0.02 | 0.86 a ± 0.01 | 1.18 b ± 0.04 | 1.19 b ± 0.07 | 2.93 b ± 0.18 | 2.91 ± 0.03 | 5.21 ± 0.06 |
| 5 | 0.70 ab± 0.02 | 0.85 a ± 0.01 | 1.19 b ± 0.04 | 1.12 b ± 0.09 | 2.93 b ± 0.21 | 3.07 ± 0.08 | 5.07 ± 0.09 |
| 10 | 0.73 b ± 0.01 | 0.84 a ± 0.01 | 1.06 b ± 0.03 | 0.97 b ± 0.06 | 2.53 b ± 0.13 | 2.94 ± 0.03 | 4.99 ± 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dridi, N.; Brito, P.; Bouslimi, H.; Ferreira, R.; Martins-Dias, S.; Caçador, I.; Sleimi, N. Physiological and Biochemical Behaviours and Antioxidant Response of Helianthus annuus under Lanthanum and Cerium Stress. Sustainability 2022, 14, 4153. https://doi.org/10.3390/su14074153
Dridi N, Brito P, Bouslimi H, Ferreira R, Martins-Dias S, Caçador I, Sleimi N. Physiological and Biochemical Behaviours and Antioxidant Response of Helianthus annuus under Lanthanum and Cerium Stress. Sustainability. 2022; 14(7):4153. https://doi.org/10.3390/su14074153
Chicago/Turabian StyleDridi, Nesrine, Pedro Brito, Houda Bouslimi, Renata Ferreira, Susete Martins-Dias, Isabel Caçador, and Noomene Sleimi. 2022. "Physiological and Biochemical Behaviours and Antioxidant Response of Helianthus annuus under Lanthanum and Cerium Stress" Sustainability 14, no. 7: 4153. https://doi.org/10.3390/su14074153