
Exploring Local Maize Diversity for Increased Agricultural Sustainability: New Insights into Drought Stress Response and Recovery of Guinea-Bissau Landraces

 ,
,  , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Setup
2.2. Determination of Leaf Relative Water Content (RWC)
2.3. Chlorophyll Fluorescence Analysis
2.4. Photosynthetic Pigments Determination
2.5. Fatty Acids Profiling
2.6. RNA Extraction and cDNA Synthesis
2.7. Real-Time PCR (qPCR)
2.8. Statistical Analysis
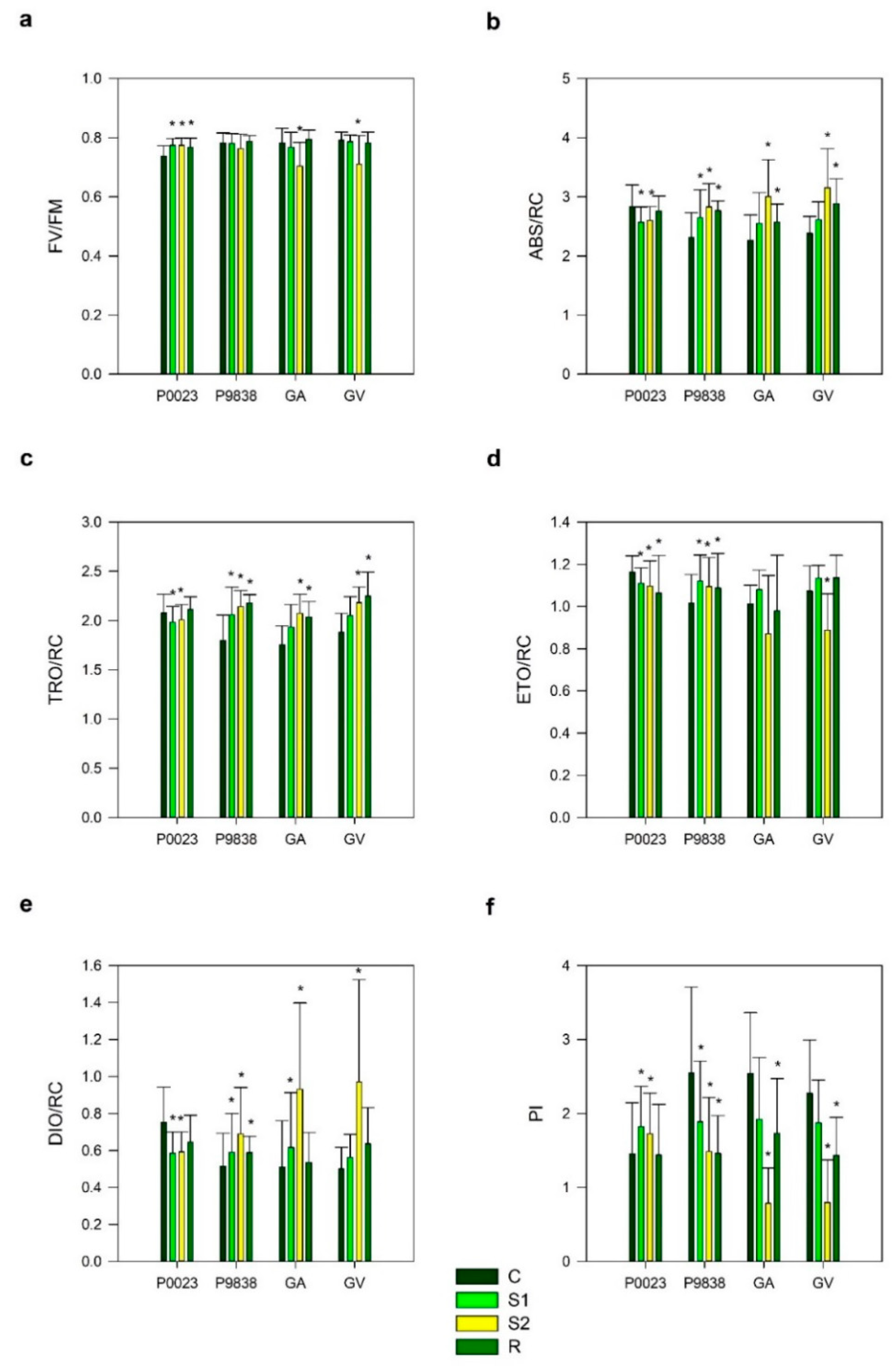
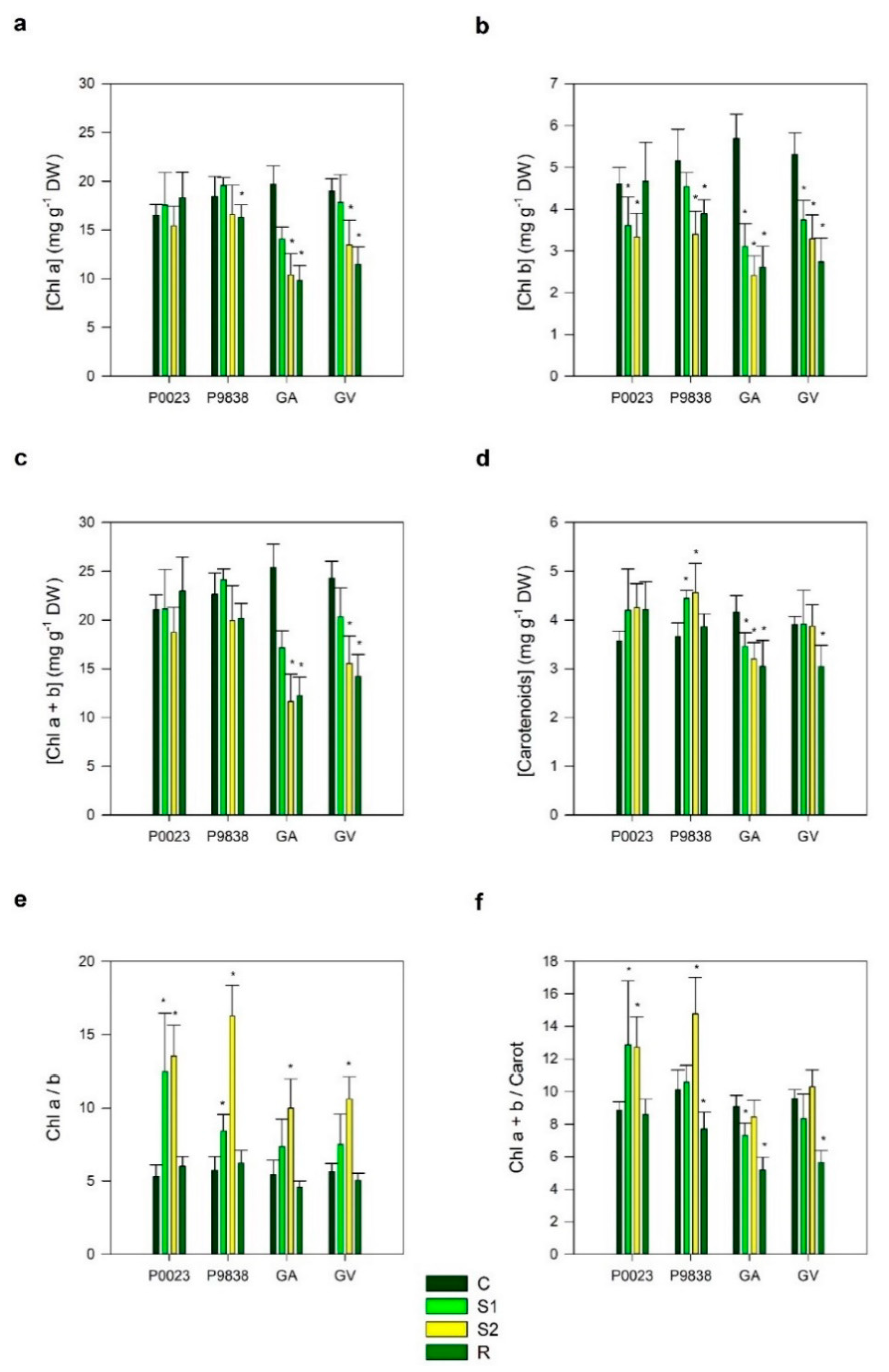
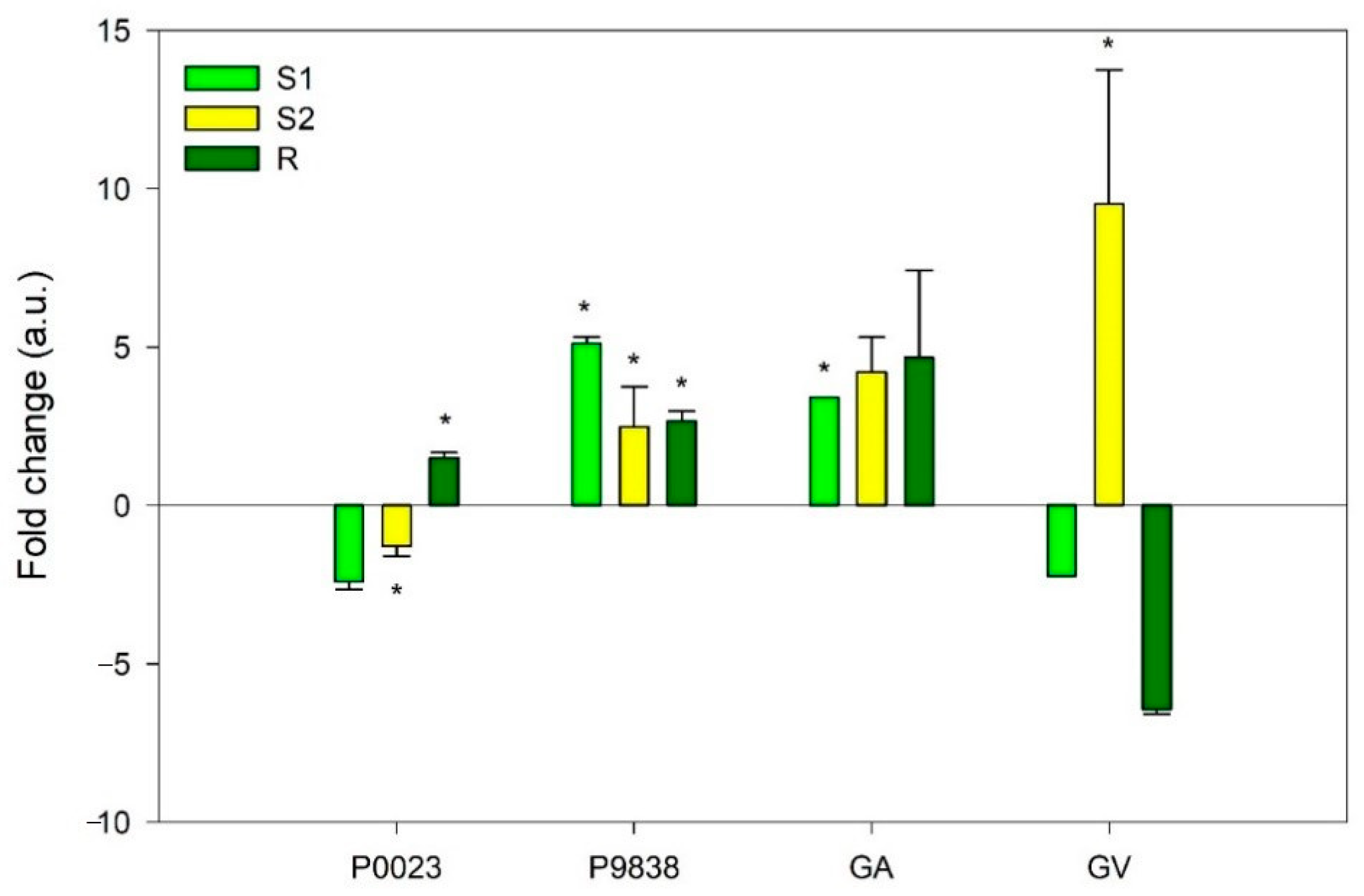
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silva, J.L. O Zea Mays e a Expansão Portuguesa; Instituto de Investigação Científica Tropical: Lisboa, Portugal, 1998; p. 212. [Google Scholar]
- Campos, H.; Cooper, M.; Habben, J.E.; Edmeades, G.O.; Schussler, J.R. Improving drought tolerance in maize: A view from industry. Field Crop. Res. 2004, 90, 19–34. [Google Scholar] [CrossRef]
- Kim, S.G.; Bae, H.H.; Jung, H.J.; Lee, J.-S.; Kim, J.-T.; Go, T.H.; Son, B.-Y.; Baek, S.-B.; Kwon, Y.-U.; Woo, M.-O.; et al. Physiological and protein profiling response to drought stress in KS141, a Korean maize inbred line. J. Crop. Sci. Biotechnol. 2014, 17, 273–280. [Google Scholar] [CrossRef]
- Moola, S. Africa’s Green Revolution Drought Tolerant Maize Scam; ACB Briefing Paper No. 12; African Centre for Biodiversity: Melville, South Africa, 2010; p. 18. Available online: https://www.acbio.org.za/wp-content/uploads/2015/02/ACB-Africa-Drought-Tolerant-Maize-2010.pdf (accessed on 1 February 2021).
- Santpoort, R. The Drivers of Maize Area Expansion in Sub-Saharan Africa. How Policies to Boost Maize Production Overlook the Interests of Smallholder Farmers. Land 2020, 9, 68. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro de Carvalho, M.A.; Bebeli, P.J.; Bettencourt, E.; Costa, G.; Dias, S.; Santos, T.M.; Slaski, J.J. Cereal landraces genetic resources in worldwide GeneBanks. A review. Agron. Sustain. Dev. 2013, 33, 177–203. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Verma, R.P.; Singh, A.; Sharma, H.K.; Devi, G. Barley landraces: Ecological heritage for edaphic stress adaptations and sustainable production. Environ. Sustain. Indic. 2020, 6, 100035. [Google Scholar] [CrossRef]
- Koutsika-Sotiriou, M.; Tsivelikas, A.L.; Gogas, C.; Mylonas, I.G.; Avdikos, I.; Traka-Mavrona, E. Breeding methodology meets sustainable agriculture. Int. J. Plant Breed. Genet. 2013, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Newton, A.C.; Akar, T.; Baresel, J.P.; Bebeli, P.J.; Bettencourt, E.; Bladenopoulos, K.V.; Czembor, J.H.; Fasoula, D.A.; Katsiotis, A.; Koutis, K.; et al. Cereal landraces for sustainable agriculture. A review. Agron. Sustain. Dev. 2010, 30, 237–269. [Google Scholar] [CrossRef] [Green Version]
- Collins, N.C.; Tardieu, F.; Tuberosa, R. Quantitative Trait Loci and Crop Performance under Abiotic Stress: Where Do We Stand? Plant Physiol. 2008, 147, 469–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, M.P.; Ortiz, R. Adapting crops to climate change: A summary. In Climate Change and Crop Production; Reynolds, M.P., Ed.; CABI: Oxfordshire, UK, 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity, and drought stress. Arch Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Li, Y.; Zhou, L.; Xu, Z.; Zhou, G. Maize leaf functional responses to drought episode and rewatering. Agr. Forest Meteorol. 2018, 249, 57–70. [Google Scholar] [CrossRef]
- Paviot, M.C.; Bresnyan, E.W., Jr.; Diaz, A. Guinea Bissau: Unlocking Diversification to Unleash Agriculture Growth; World Bank: Washington, DC, USA, 2019; p. 142. Available online: http://documents1.worldbank.org/curated/en/341991563831364596/pdf/Guinea-Bissau-ASA-Agriculture-sector-Report.pdf (accessed on 1 February 2021).
- Fghire, R.; Anaya, F.; Ali, O.I.; Benlhabib, O.; Ragab, R.; Wahbi, S. Physiological and photosynthetic response of quinoa to drought stress. Chil. J. Agr. Res. 2015, 75, 174–183. [Google Scholar] [CrossRef] [Green Version]
- Opitz, N.; Paschold, A.; Marcon, C.; Malik, W.A.; Lanz, C.; Piepho, H.P.; Hochholdinger, F. Transcriptomic complexity in young maize primary roots in response to low water potentials. BMC Genom. 2014, 15, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challinor, A.; Wheeler, T.; Garforth, C.; Craufurd, P.; Kassm, A. Assessing the vulnerability of food crop systems in Africa to climate change. Clim. Chang. 2007, 83, 381–399. [Google Scholar] [CrossRef]
- Tesfaye, K.; Gbegbelegbe, S.; Cairns, J.E.; Shiferaw, B.; Prasanna, B.M.; Sonder, K.; Boote, K.; Makumbi, D.; Robertson, R. Maize systems under climate change in sub-Saharan Africa: Potential impacts on production and food security. Int. J. Clim. Chang. Str. 2016, 7, 247–271. [Google Scholar] [CrossRef]
- Sylla, M.B.; Nikiema, P.M.; Gibba, P.; Kebe, I.; Klutse, N.A.B. Climate Change over West Africa: Recent Trends and Future Projections. In Adaptation to Climate Change and Variability in Rural West Africa; Yaro, J., Hesselberg, J., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- FAO. Climate-Smart Agriculture Sourcebook; FAO: Rome, Italy, 2017; p. 56. Available online: http://www.fao.org/3/i7994e/i7994e.pdf (accessed on 1 February 2021).
- Efeoǧlu, B.; Ekmekçi, Y.; Çiçek, N. Physiological responses of three maize cultivars to drought stress and recovery. S. Afr. J. Bot. 2009, 75, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Cang, Z.; Jiao, F.; Bai, X.; Zhang, D.; Zhai, R. Influence of drought stress on photosynthetic characteristics and protective enzymes of potato at seedling stage. J. Saudi Soc. Agric. Sci. 2017, 16, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Toumi, I.; Gargouri, M.; Nouairi, I.; Moschou, P.N.; Ben Salem-Fnayou, A.; Mliki, A.; Zarrouk, M.; Ghorbel, A. Water stress induced changes in the leaf lipid composition of four grapevine genotypes with different drought tolerance. Biologia. Plantarum. 2008, 52, 161–164. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Vanderbeld, B.; Wan, J.; Huang, Y. Narrowing down the targets: Towards successful genetic engineering of drought-tolerant crops. Mol. Plant 2010, 3, 469–490. [Google Scholar] [CrossRef]
- Heisey, P.W.; Edmeades, G.O. Maize Production in Drought-Stressed Environments: Technical Options and Research Resource Allocation; Part 1 of CIMMYT 1997/98 World Maize Facts and Trends; CIMMYT: Mexico City, Mexico, 1999; pp. 1–74. Available online: http://hdl.handle.net/10883/759 (accessed on 1 February 2021).
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Mir, R.R.; Zaman-Allah, M.; Sreenivasulu, N.; Trethowan, R.; Varshney, R.K. Integrated genomics, physiology and breeding approaches for improving drought tolerance in crops. Theor. Appl. Genet. 2012, 125, 625–645. [Google Scholar] [CrossRef] [Green Version]
- Colom, M.R.; Vazzana, C. Photosynthesis and PSII functionality of drought-resistant and drought-sensitive weeping lovegrass plants. Environ. Exp. Bot. 2003, 49, 135–144. [Google Scholar] [CrossRef]
- Fleta-Soriano, E.; Díaz, L.; Bonet, E.; Munné-Bosch, S. Melatonin may exert a protective role against drought stress in maize. J. Agron. Crop Sci. 2017, 203, 286–294. [Google Scholar] [CrossRef]
- Lima, A.L.S.; DaMatta, F.M.; Pinheiro, H.A.; Totola, M.R.; Loureiro, M.E. Photochemical responses and oxidative stress in two clones of Coffea canephora under water deficit conditions. Environ. Exp. Bot. 2002, 47, 239–247. [Google Scholar] [CrossRef]
- Liu, F.; Jensen, C.R.; Shahanzari, A.; Andersen, M.N.; Jacobsen, S.E. ABA regulated stomatal control and photosynthetic water use efficiency of potato (Solanum tuberosum L.) during progressive soil drying. Plant Sci. 2005, 168, 831–836. [Google Scholar] [CrossRef]
- Goodarzian-Ghahfarokhi, M.; Mansouri-Far, C.; Saeidi, M.; Abdoli, M. Different physiological and biochemical responses in maize hybrids subjected to drought stress at vegetative and reproductive stages. Acta Biol. Szeged. 2016, 60, 27–37. [Google Scholar]
- Yadav, S.; Sharm, K.D. Molecular and Morphophysiological Analysis of Drought Stress in Plants. In Plant Growth; Rigobelo, E.C., Ed.; IntechOpen: London, UK, 2016; pp. 149–173. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, E.; Hsiao, T.C.; Henderson, D.W. Immediate and subsequent growth responses of maize leaves to changes in water status. Plant Physiol. 1971, 48, 631–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Wang, S.; Cao, B.; Cao, D.; Leng, G.; Li, H.; Yin, L.; Shan, L.; Deng, X. Genotypic variation in growth and physiological response to drought stress and re-watering reveals the critical role of recovery in drought adaptation in maize seedlings. Front. Plant Sci. 2016, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farre, I.; Oijen, M. van, Leffelaar, P.A.; Faci, J.M. Analysis of maize growth for different irrigation strategies in northeastern Spain. Eur. J. Agron. 2000, 12, 225–238. [Google Scholar] [CrossRef]
- Gaffney, J.; Schussler, J.; Löffler, C.; Cai, W.; Paszkiewicz, S.; Messina, C.; Groeteke, J.; Keaschall, J.; Cooper, M. Industry-Scale Evaluation of Maize Hybrids Selected for Increased Yield in Drought-Stress Conditions of the US Corn Belt. Crop Sci. 2015, 55, 1608–1618. [Google Scholar] [CrossRef]
- Catsky, J. Determination of water deficit in disks cut out from leaf blades. Biol. Plantarum. 1960, 2, 76–78. [Google Scholar] [CrossRef]
- Strasser, B.J.; Strasser, R.J. Measuring Fast Fluorescence Transients to Address Environmental Questions: The JIP-test. In Photosynthesis: From Light to Biosphere; Mathis, P., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 977–980. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The Fluorescence Transient as a Tool to Characterize and Screen Photosynthetic Samples. In Probing Photosynthesis: Mechanism, Regulation and Adaptation, 1st ed.; Yunus, M., Pathre, U., Mohanty, P., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 445–483. [Google Scholar]
- Silvestre, S.; Araújo, S.; Vaz Patto, M.C.; Marques da Silva, J. Performance index: An expeditious tool to screen for improved drought resistance in the Lathyrus genus. J. Integr. Plant Biol. 2014, 56, 610–621. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method. Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Feijão, E.; Gameiro, C.; Franzitta, M.; Duarte, B.; Caçador, I.; Cabrita, M.T.; Matos, A.R. Heat wave impacts on the model diatom Phaeodactylum tricornutum: Searching for photochemical and fatty acid biomarkers of thermal stress. Ecol. Indic. 2017, 95, 1026–1037. [Google Scholar] [CrossRef]
- Gameiro, C.; Utkin, A.B.; Cartaxana, P.; Marques da Silva, J.; Matos, A.R. The use of laser induced chlorophyll fluorescence (LIF) as a fast and non-destructive method to investigate water deficit in Arabidopsis. Agri. Water Manag. 2016, 164, 127–136. [Google Scholar] [CrossRef]
- Thirunavukkarasu, N.; Hossain, F.; Arora, K.; Sharma, R.; Shiriga, K.; Mittal, S.; Mohan, S.; Namratha, P.M.; Dogga, S.; Rani, T.S.; et al. Functional mechanisms of drought tolerance in subtropical maize (Zea mays L.) identified using genome-wide association mapping. BMC Genom. 2014, 15, 1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, R.; Maia, M.; Ferreira, A.E.; Silva, A.B.; Freire, A.P.; Cordeiro, C.; Silva, M.S.; Figueiredo, A. Early stage metabolic events associated with the establishment of Vitis vinifera—Plasmopara viticola compatible interaction. Plant Physiol. Biochem. 2019, 137, 1–13. [Google Scholar] [CrossRef]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiango, H.; Figueiredo, A.; Sousa, L.; Sinclair, T.; Marques da Silva, J. Assessing drought tolerance of traditional maize genotypes of Mozambique using chlorophyll fluorescence parameters. S. Afr. J. Bot. 2021, 138, 311–317. [Google Scholar] [CrossRef]
- Cruz de Carvalho, R.; Cunha, A.; Marques da Silva, J. Photosynthesis by six Portuguese maize cultivars during drought stress and recovery. Acta Physiol. Plant. 2011, 33, 359–374. [Google Scholar] [CrossRef] [Green Version]
- Min, H.; Chen, C.; Wei, S.; Shang, X.; Sun, M.; Xia, R.; Liu, X.; Hao, D.; Chen, H.; Xie, Q. Identification of drought tolerant mechanisms in maize seedlings based on transcriptome analysis of recombination inbred lines. Front. Plant Sci. 2016, 7, 1–19. [Google Scholar] [CrossRef]
- Franks, P.J.; Farquhar, G.D. The effect of exogenous abscisic accid on stomatal development, tomatal mechanics, and leaf gas exchange in Tradescantia virginiana. Plant Physiol. 2001, 125, 935–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harb, A.; Krishnan, A.; Ambavaram, M.M.R.; Pereira, A. Molecular and physiological analysis of drought stress in arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiol. 2010, 154, 1254–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques da Silva, J.; Arrabaça, M.C. Photosynthesis in the water stressed C4 grass Setaria sphacelata is mainly limited by stomata with both rapidly and slowly imposed water deficits. Physiol. Plant 2004, 121, 409–420. [Google Scholar] [CrossRef]
- Bjorkman, O.; Demmig, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Epron, D.; Godard, D.; Cornic, G.; Genty, B. Limitation of net CO2 assimilation Rate by internal resistances to CO2 Transfer in the leaves of 2 tree species (Fagus sylvatica L and Castanea sativa Mill). Plant Cell Environ. 1995, 18, 43–51. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Govindjee; Bosa, K.; Kościelniak, J.; Zuk-Gołaszewska, K. Effects of salt stress on photosystem II efficiency and CO2 assimilation of two Syrian barley landraces. Environ. Exp. Bot. 2011, 73, 64–72. [Google Scholar] [CrossRef]
- Correia, P.M.; da Silva, A.B.; Roitsch, T.; Carmo-Silva, E.; Marques da Silva, J. Photoprotection and optimization of sucrose usage contribute to faster recovery of photosynthesis after water deficit at high temperatures in wheat. Physiol. Plant. 2020, 1–14. [Google Scholar] [CrossRef]
- Chandrasekar, V.; Sairam, R.K.; Srivastava, G.C. Physiological and biochemical responses of hexaploid and tetraploid wheat to drought stress. J. Agron. Crop. Sci. 2000, 185, 219–227. [Google Scholar] [CrossRef]
- Aranjuelo, I.; Molero, G.; Erice, G.; Avice, J.C.; Nogués, S. Plant physiology and proteomics reveals the leaf response to drought in alfalfa (Medicago sativa L.). J. Exp. Bot. 2011, 62, 111–123. [Google Scholar] [CrossRef] [Green Version]
- Pintó-Marijuan, M.; Munné-Bosch, S. Photo-oxidative stress markers as a measure of abiotic stress-induced leaf senescence: Advantages and limitations. J. Exp. Bot. 2014, 65, 3845–3857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitajima, K.; Hogan, K.P. Increases of chlorophyll a/b ratios during acclimation of tropical woody seedlings to nitrogen limitation and high light. Plant Cell Environ. 2003, 26, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Upchurch, R.G. Fatty acid unsaturation, mobilization, and regulation in the response of plants to stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Gigon, A.; Matos, A.R.; Laffray, D.; Zuily-Fodil, Y.; Pham-Thi, A.T. Effect of drought stress on lipid metabolism in the leaves of Arabidopsis thaliana (Ecotype Columbia). Ann. Bot. 2004, 94, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.R.; Gigon, A.; Laffray, D.; Pêtres, S.; Zuily-Fodil, Y.; Pham-Thi, A.T. Effects of progressive drought stress on the expression of patatin-like lipid acyl hydrolase genes in Arabidopsis leaves. Physiol. Plant. 2008, 134, 110–120. [Google Scholar] [CrossRef]
- Sebastiana, M.; da Silva, A.B.; Matos, A.R.; Alcântara, A.; Silvestre, S.; Malhó, R. Ectomycorrhizal inoculation with Pisolithus tinctorius reduces stress induced by drought in cork oak. Mycorrhiza 2018, 28, 247–258. [Google Scholar] [CrossRef]
- Dakhma, W.S.; Zarrouk, M.; Cherif, A. Effects of drought-stress on lipids in rape leaves. Phytochemistry 1995, 40, 1383–1386. [Google Scholar] [CrossRef]
- Seiwert, D.; Witt, H.; Ritz, S.; Janshoff, A.; Paulsen, H. The Nonbilayer Lipid MGDG and the Major Light-Harvesting Complex (LHCII) Promote Membrane Stacking in Supported Lipid Bilayers. Biochemistry 2018, 57, 2278–2288. [Google Scholar] [CrossRef]
- Los, D.A.; Murata, N. Membrane fluidity and its roles in the perception of environmental signals. Biochim. Biophys. Acta Biomembr. 2004, 1666, 142–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, B.; Matos, A.R.; Caçador, I. Photobiological and lipidic responses reveal the drought tolerance of Aster tripolium cultivated under severe and moderate drought: Perspectives for arid agriculture in the mediterranean. Plant. Physiol. Biochem. 2020, 154, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Tian, B.; Zhang, F.; Tao, F.; Li, W. Plant adaptation to frequent alterations between high and low temperatures: Remodeling of membrane lipids and maintenance of unsaturation levels. Plant Cell Enviro. 2011, 34, 1431–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, D.; Du, H.; Wang, Z.; Huang, B. Genotypic variation in fatty acid composition and unsaturation levels in bermudagrass associated with leaf dehydration tolerance. J. Am. Soc. Hortic. Sci. 2011, 136, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Loll, B.; Kern, J.; Saenger, W.; Zouni, A.; Biesiadka, J. Towards complete cofactor arrangement in the 3.0 A resolution structure of photosystem II. Nature 2005, 438, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- Camoni, L.; Di Lucente, C.; Pallucca, R.; Visconti, S.; Aducci, P. Binding of phosphatidic acid to 14-3-3 proteins hampers their ability to activate the plant plasma membrane H+-ATPase. IUBMB Life 2012, 64, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cao, B.; Han, N.; Tao, Y.; Zhou, S.F.; Li, W.C.; Fu, F.L. Phospholipase D family and its expression in response to abiotic stress in maize. Plant. Growth. Regul. 2017, 81, 197–207. [Google Scholar] [CrossRef]
- Zhao, J. Phospholipase D and phosphatidic acid in plant defence response: From protein-protein and lipid-protein interactions to hormone signalling. J. Exp. Bot. 2015, 66, 1721–1736. [Google Scholar] [CrossRef] [Green Version]
- An, Z.F.; Li, C.Y.; Zhang, L.X.; Alva, A.K. Role of polyamines and phospholipase D in maize (Zea mays L.) response to drought stress. S. Afr. J. Bot. 2012, 83, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Hayano-Kanashiro, C.; Calderón-Vásquez, C.; Ibarra-Laclette, E.; Herrera-Estrella, L.; Simpson, J. Analysis of gene expression and physiological responses in three Mexican maize landraces under drought stress and recovery irrigation. PLoS ONE 2009, 4, e7531. [Google Scholar] [CrossRef]
- Bellon, M.R.; van Etten, J. Climate Change and On-farm Conservation of Crop Landraces in Centres of Diversity. In Plant Genetic Resources and Climate Change; Jackson, M., Ford-Lloyd, B., Parry, M., Eds.; CAB International: Wallingford, UK, 2014; pp. 137–150. [Google Scholar]
- Ficiciyan, A.; Loos, J.; Sievers-Glotzbach, S.; Tscharntke, T. More than Yield: Ecosystem Services of Traditional versus Modern Crop Varieties Revisited. Sustainability 2018, 10, 2834. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, M.; Feijão, E.; Catarino, L.; Matos, A.R.; Figueiredo, A.; Marques da Silva, J. Exploring Local Maize Diversity for Increased Agricultural Sustainability: New Insights into Drought Stress Response and Recovery of Guinea-Bissau Landraces. Sustainability 2021, 13, 5441. https://doi.org/10.3390/su13105441
Teixeira M, Feijão E, Catarino L, Matos AR, Figueiredo A, Marques da Silva J. Exploring Local Maize Diversity for Increased Agricultural Sustainability: New Insights into Drought Stress Response and Recovery of Guinea-Bissau Landraces. Sustainability. 2021; 13(10):5441. https://doi.org/10.3390/su13105441
Chicago/Turabian StyleTeixeira, Manuela, Eduardo Feijão, Luís Catarino, Ana Rita Matos, Andreia Figueiredo, and Jorge Marques da Silva. 2021. "Exploring Local Maize Diversity for Increased Agricultural Sustainability: New Insights into Drought Stress Response and Recovery of Guinea-Bissau Landraces" Sustainability 13, no. 10: 5441. https://doi.org/10.3390/su13105441