New ISE-Based Apparatus for Na+, K+, Cl−, pH and Transepithelial Potential Difference Real-Time Simultaneous Measurements of Ion Transport across Epithelial Cells Monolayer–Advantages and Pitfalls


Abstract
:1. Introduction
2. Materials and Methods
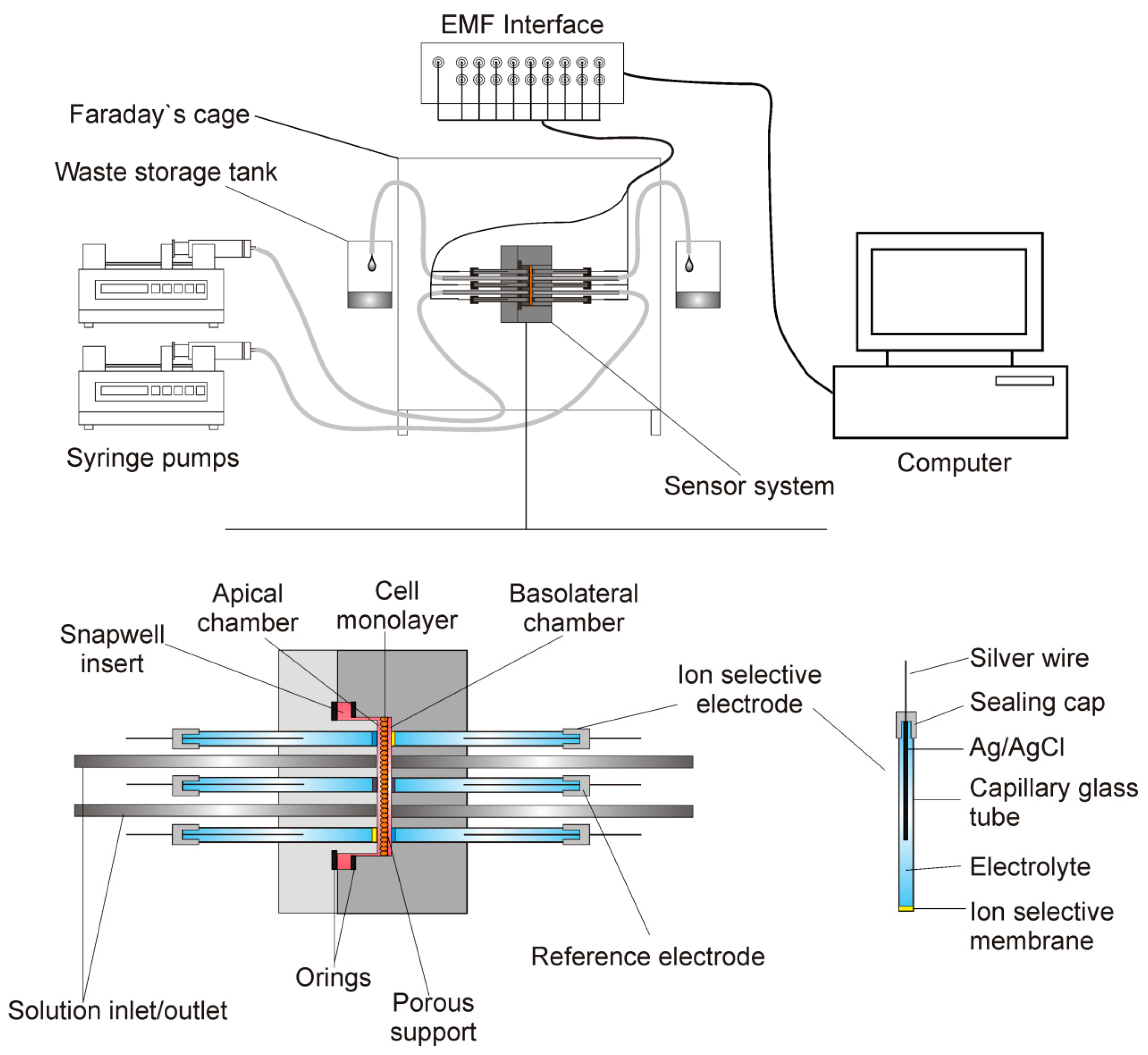
2.1. The Apparatus
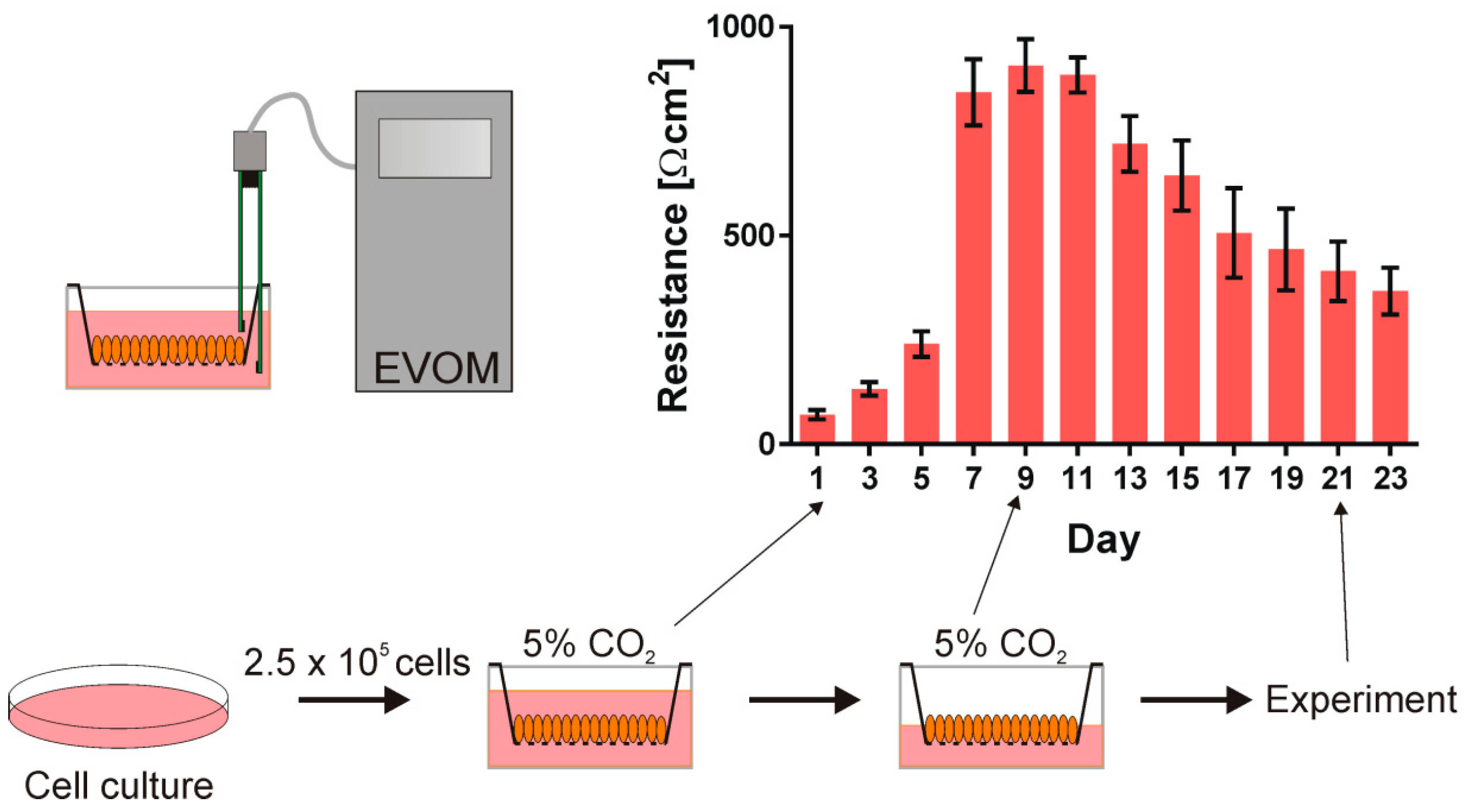
2.2. The Cells
2.3. The Media
2.4. Electrodes
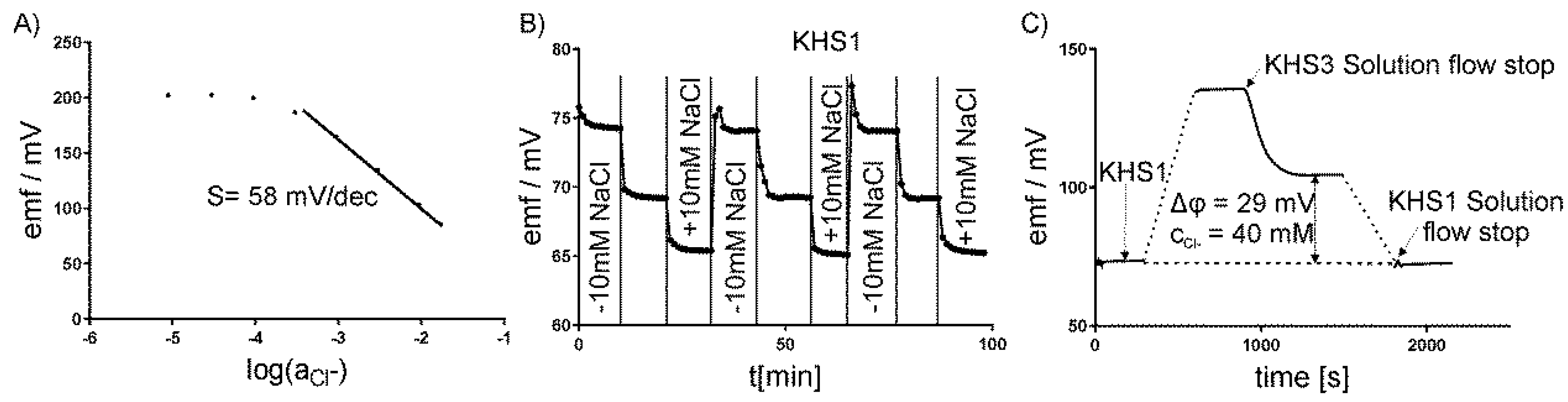
2.5. Calibration of Electrodes
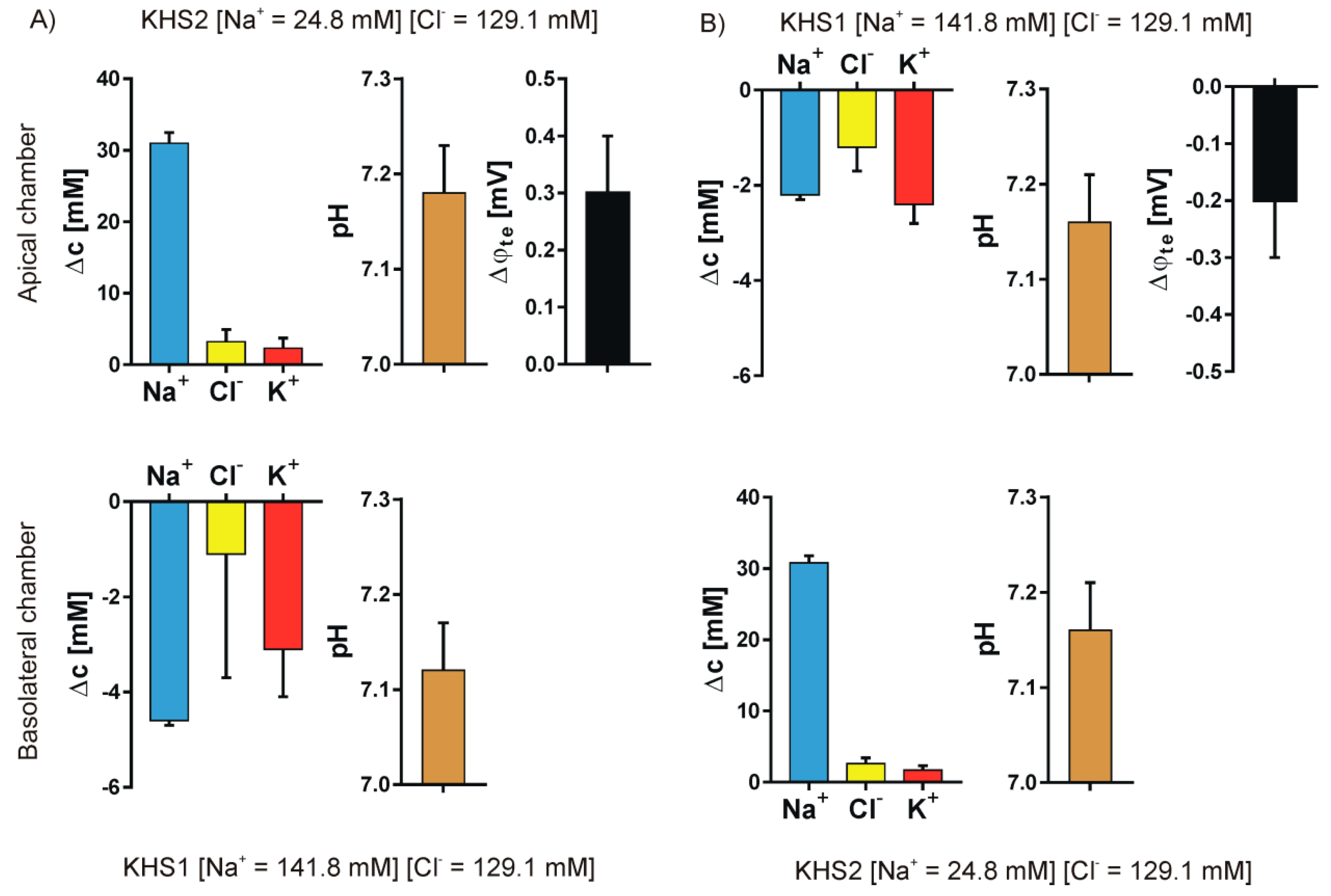
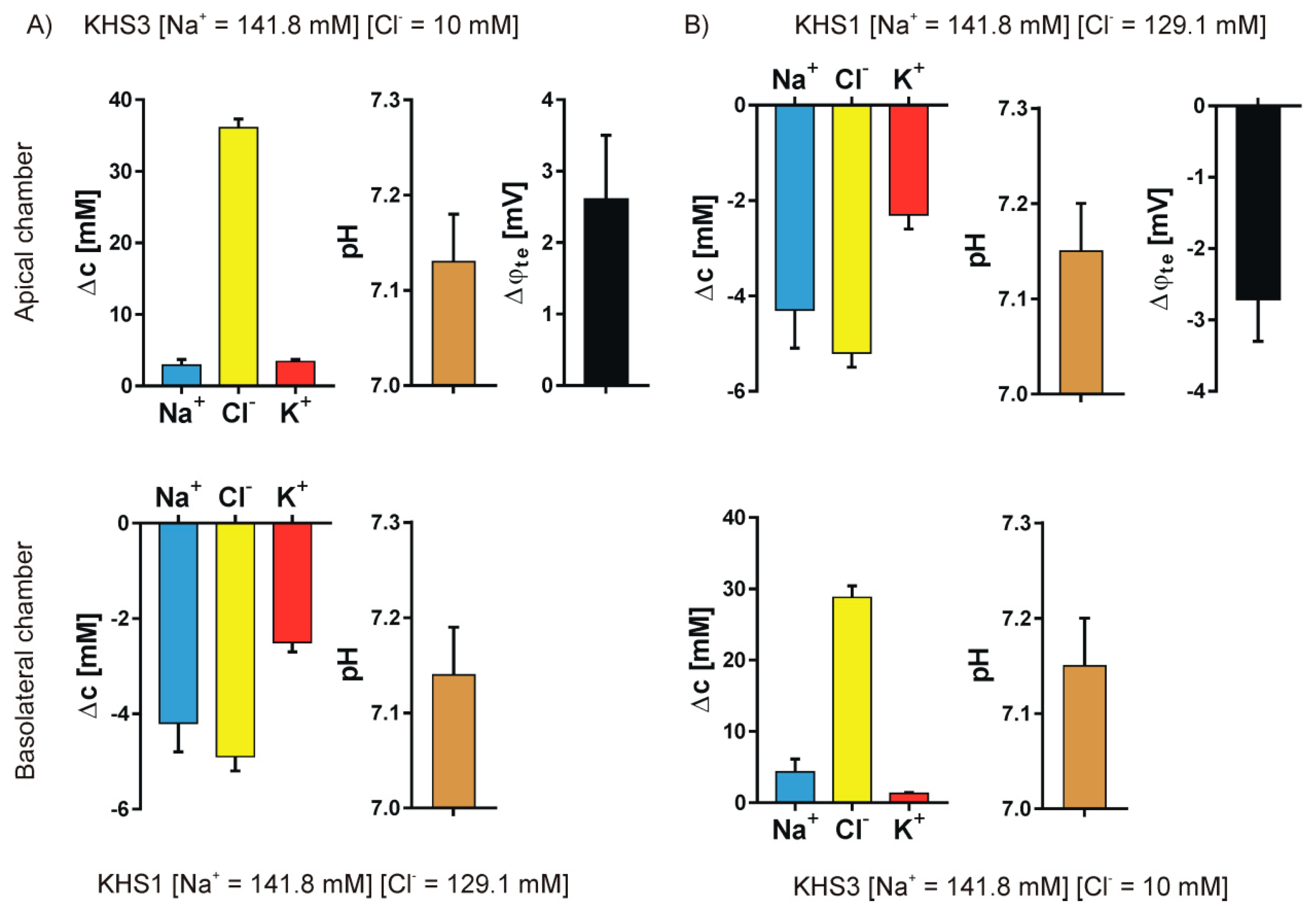
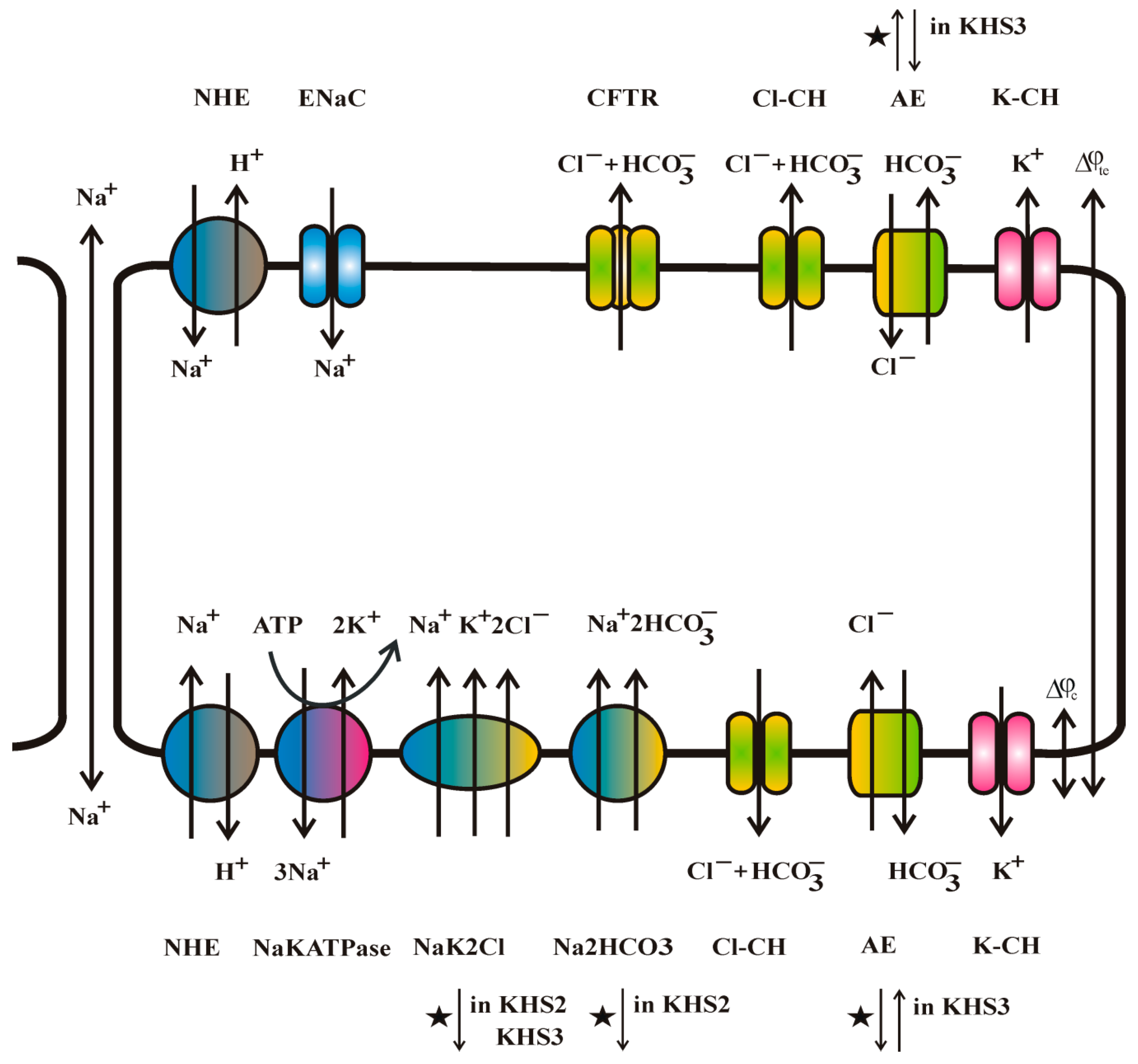
3. Results
4. Discussion
4.1. Medium pH Change Can Be Caused by Different Mechanisms
4.2. Medium Composition Could Affect the Direction of Transport of Biological Molecules
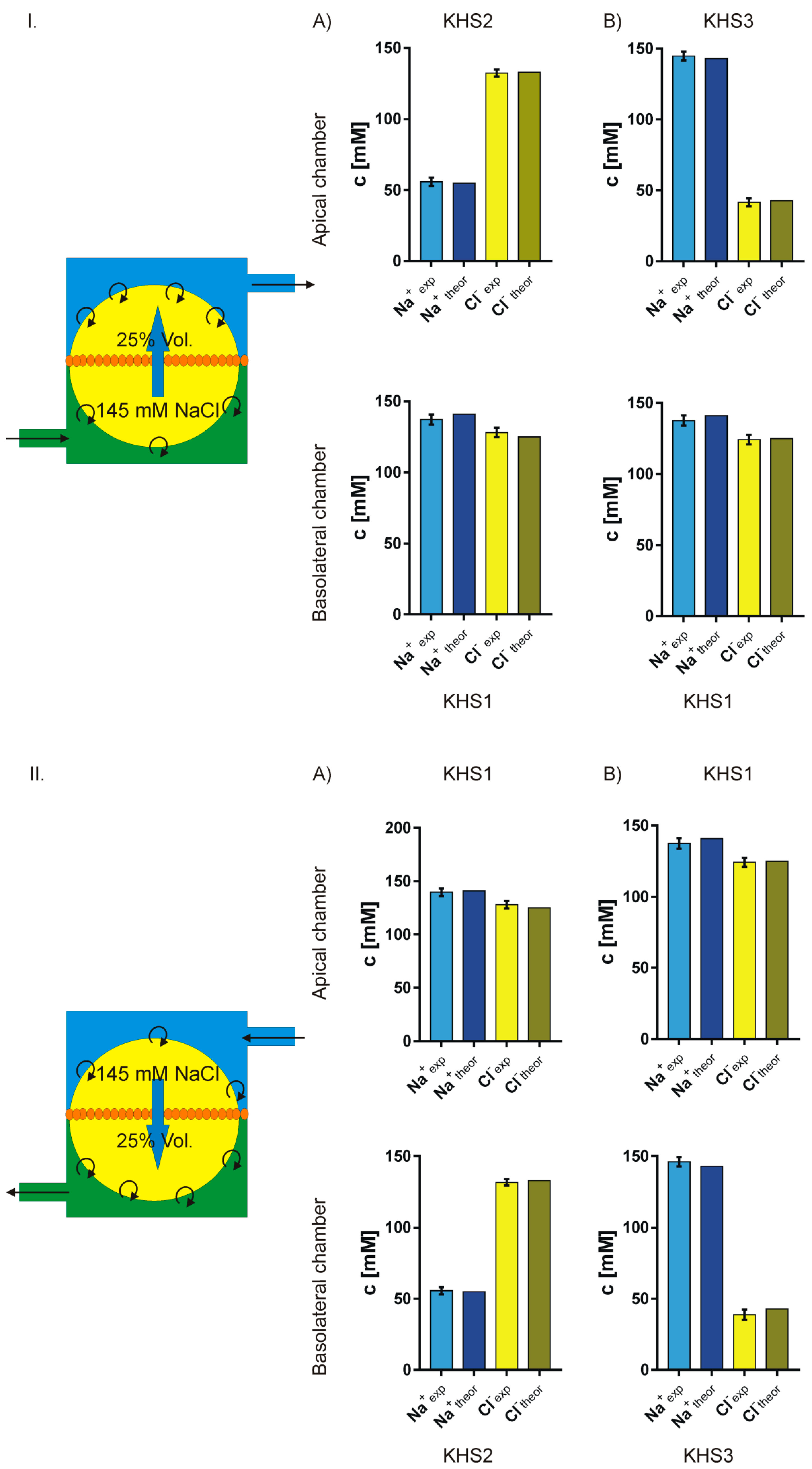
4.3. Isosmotic NaCl Flow Through Epithelial Cell Layer in Both Directions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Webster, M.J.; Tarran, R. Slippery When Wet: Airway Surface Liquid Homeostasis and Mucus Hydration. Cell Vol. Regul. 2018, 81, 293–335. [Google Scholar] [CrossRef]
- Riordan, J.R. The cystic-fibrosis transmembrane conductance regulator. Annu. Rev. Physiol. 1993, 55, 609–630. [Google Scholar] [CrossRef] [PubMed]
- Welsh, M.J.; Tsui, L.C.; Boat, T.F.; Baudet, A.L. Cystic fibrosis. In The Metabolic and Molecular Bases of Inherited Disease; Scriver, C.R., Baudet, A.L., Sly, W.S., Valle, D., Eds.; McGraw Hill: New York, NY, USA, 1995; pp. 3799–3876. [Google Scholar]
- Hollenhorst, M.I.; Richter, K.; Fronius, M. Ion Transport by Pulmonary Epithelia. J. Biomed. Biotechnol. 2011. [Google Scholar] [CrossRef]
- Novak, I. Purinergic signalling in epithelial ion transport: Regulation of secretion and absorption. Acta Physiol. 2011, 202, 501–522. [Google Scholar] [CrossRef]
- Toczylowska-Maminska, R.; Dolowy, K. Ion transporting proteins of human bronchial epithelium. J. Cell. Biochem. 2012, 113, 426–432. [Google Scholar] [CrossRef]
- Mall, M.A.; Galietta, L.J.V. Targeting ion channels in cystic fibrosis. J. Cyst. Fibros. 2015, 14, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Bartoszewski, R.; Matalon, S.; Collawn, J.F. Ion channels of the lung and their role in disease pathogenesis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 313, L859–L872. [Google Scholar] [CrossRef]
- Kunzelmann, K.; Milenkovic, V.M.; Spitzner, M.; Soria, R.B.; Schreiber, R. Calcium-dependent chloride conductance in epithelia: Is there a contribution by Bestrophin? Pflug. Arch. Eur. J. Physiol. 2007, 454, 879–889. [Google Scholar] [CrossRef]
- Bucheimer, R.E.; Linden, J. Purinergic regulation of epithelial transport. J. Physiol. 2004, 555, 311–321. [Google Scholar] [CrossRef]
- Fischer, H.; Widdicombe, J.H. Mechanisms of acid and base secretion by the airway epithelium. J. Membr. Biol. 2006, 211, 139–150. [Google Scholar] [CrossRef]
- Haggie, P.M.; Verkman, A.S. Defective organellar acidification as a cause of cystic fibrosis lung disease: Reexamination of a recurring hypothesis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L859–L867. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.W.; Chen, J.H.; Hughes, L.K.; Li, H.Y.; Sheppard, D.N. The Physiology and Pharmacology of the CFTR Cl-Channel. Chloride Mov. Across Cell. Membr. 2007, 38, 109–143. [Google Scholar] [CrossRef]
- Wei, L.; Vankeerberghen, A.; Cuppens, H.; Eggermont, J.; Cassiman, J.J.; Droogmans, G.; Nilius, B. Interaction between calcium-activated chloride channels and the cystic fibrosis transmembrane conductance regulator. Pflug. Arch. Eur. J. Physiol. 1999, 438, 635–641. [Google Scholar] [CrossRef]
- Gaillard, E.A.; Kota, P.; Gentzsch, M.; Dokholyan, N.V.; Stutts, M.J.; Tarran, R. Regulation of the epithelial Na+ channel and airway surface liquid volume by serine proteases. Pflug. Arch. Eur. J. Physiol. 2010, 460, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardou, O.; Trinh, N.T.N.; Brochiero, E. Molecular diversity and function of K+ channels in airway and alveolar epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L145–L155. [Google Scholar] [CrossRef] [PubMed]
- Ando-Akatsuka, Y.; Abdullaev, I.F.; Lee, E.L.; Okada, Y.; Sabirov, R.Z. Down-regulation of volume-sensitive Cl- channels by CFTR is mediated by the second nucleotide-binding domain. Pflug. Arch. Eur. J. Physiol. 2002, 445, 177–186. [Google Scholar] [CrossRef]
- Zajac, M.; Dolowy, K. Measurement of ion fluxes across epithelia. Prog. Biophys. Mol. Biol. 2017, 127, 1–11. [Google Scholar] [CrossRef]
- Ussing, H.H.; Zerahn, K. Active transport of sodium as the source of electric current in the short-circuited isolated frog skin. Acta Physiol. Scand. 1951, 23, 110–127. [Google Scholar] [CrossRef]
- Hug, M.J.; Clarke, L.L.; Gray, M.A. How to Measure CFTR-Dependent Bicarbonate Transport: From Single Channels to the Intact Epithelium. In Cystic Fibrosis: Diagnosis and Protocols, Volume I: Approaches to Study and Correct Cftr Defects; Humana Press: New York, NY, USA, 2011; pp. 489–509. [Google Scholar] [CrossRef]
- Nair, S.; Kashyap, R.; Laboisse, C.L.; Hopfer, U.; Gratzl, M. Time resolved secretion of chloride from a monolayer of mucin-secreting epithelial cells. Eur. Biophys. J. Biophys. Lett. 2008, 37, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Kashyap, R.; Laboisse, C.; Hopfer, U.; Gratzl, M. Time-resolved release of calcium from an epithelial cell monolayer during mucin secretion. Eur. Biophys. J. Biophys. Lett. 2011, 40, 165–174. [Google Scholar] [CrossRef]
- Toczylowska-Maminska, R.; Lewenstam, A.; Dolowy, K. Multielectrode Bisensor System for Time-Resolved Monitoring of Ion Transport Across an Epithelial Cell Layer. Anal. Chem. 2014, 86, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Zajac, M.; Lewenstam, A.; Dolowy, K. Multi-electrode system for measurement of transmembrane ion-fluxes through living epithelial cells. Bioelectrochemistry 2017, 117, 65–73. [Google Scholar] [CrossRef]
- Anagnostopoulou, P.; Dai, L.; Schatterny, J.; Hirtz, S.; Duerr, J.; Mall, M.A. Allergic airway inflammation induces a pro-secretory epithelial ion transport phenotype in mice. Eur. Respir. J. 2010, 36, 1436–1447. [Google Scholar] [CrossRef] [Green Version]
- Blaisdell, C.J.; Edmonds, R.D.; Wang, X.T.; Guggino, S.; Zeitlin, P.L. pH-regulated chloride secretion in fetal lung epithelia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 278, L1248–L1255. [Google Scholar] [CrossRef]
- Burnett, R.W.; Covington, A.K.; Fogh-Andersen, N.; Kulpmann, W.R.; Lewenstam, A.; Maas, A.H.J.; Muller-Plathe, O.; VanKessel, A.L.; Zijlstra, W.G. Use of ion-selective electrodes for blood-electrolyte analysis. Recommendations for nomenclature, definitions and conventions. Clin. Chem. Lab. Med. 2000, 38, 363–370. [Google Scholar] [CrossRef]
- Lewenstam, A. Routines and Challenges in Clinical Application of Electrochemical Ion-Sensors. Electroanalysis 2014, 26, 1171–1181. [Google Scholar] [CrossRef]
- Parrilla, M.; Cuartero, M.; Padrell-Sánchez, S.; Rajabi, M.; Roxhed, N.; Niklaus, F.; Crespo, G.A. Wearable all-solid-state potentiometric microneedle patch for intradermal potassium detection. Anal. Chem. 2019, 91, 1578–1586. [Google Scholar] [CrossRef] [PubMed]
- Treharne, K.J.; Crawford, R.M.; Mehta, A. CFTR, chloride concentration and cell volume: Could mammalian protein histidine phosphorylation play a latent role? Exp. Physiol. 2006, 91, 131–139. [Google Scholar] [CrossRef]
- Aronson, P.S.; Boron, W.F.; Boulpaep, E.L. Physiology of membranes. In Medical Physiology. A Cellular and Molecular Approach; Boron, W.L., Boulpaep, E.L., Eds.; Elsevier Saunders: Philadelphia, PA, USA, 2005; pp. 50–86. [Google Scholar]
- Baynes, J.W.; Dominiczak, M.H. Medical Biochemistry, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 978-0-7020-7299-4. [Google Scholar]
- Dolowy, K. Ion transport and (selected) ion channels in biological membranes in health and pathology. In Electrochemical Processes in Biological Systems; Wiley-Blackwell: Hoboken, NJ, USA, 2015; pp. 61–82. [Google Scholar]
- Dabrowska, S.; Migdalski, J.; Lewenstam, A. A Breakthrough Application of a Cross-Linked Polystyrene Anion-Exchange Membrane for a Hydrogencarbonate Ion-Selective Electrode. Sensors 2019, 19, 1268. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KHS1 | KHS2 | KHS3 | |
|---|---|---|---|
| [Na+] | 141.8 | 24.8 | 141.8 |
| [Chol+] | 0 | 117.0 | 0 |
| [Cl−] | 129.1 | 129.1 | 10 |
| [Gluc−] | 0 | 0 | 119.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zając, M.; Lewenstam, A.; Stobiecka, M.; Dołowy, K. New ISE-Based Apparatus for Na+, K+, Cl−, pH and Transepithelial Potential Difference Real-Time Simultaneous Measurements of Ion Transport across Epithelial Cells Monolayer–Advantages and Pitfalls. Sensors 2019, 19, 1881. https://doi.org/10.3390/s19081881
Zając M, Lewenstam A, Stobiecka M, Dołowy K. New ISE-Based Apparatus for Na+, K+, Cl−, pH and Transepithelial Potential Difference Real-Time Simultaneous Measurements of Ion Transport across Epithelial Cells Monolayer–Advantages and Pitfalls. Sensors. 2019; 19(8):1881. https://doi.org/10.3390/s19081881
Chicago/Turabian StyleZając, Mirosław, Andrzej Lewenstam, Magdalena Stobiecka, and Krzysztof Dołowy. 2019. "New ISE-Based Apparatus for Na+, K+, Cl−, pH and Transepithelial Potential Difference Real-Time Simultaneous Measurements of Ion Transport across Epithelial Cells Monolayer–Advantages and Pitfalls" Sensors 19, no. 8: 1881. https://doi.org/10.3390/s19081881