
Comparison of UAV and Boat Surveys for Detecting Changes in Breeding Population Dynamics of Sea Turtles


1
School of Biological Sciences, Monash University, Clayton, VIC 3800, Australia
2
Terra Verde Environmental Consulting LLC, San Luis Obispo, CA 93401, USA
*
Author to whom correspondence should be addressed.
Remote Sens. 2021, 13(15), 2857; https://doi.org/10.3390/rs13152857
Submission received: 25 May 2021
/
Revised: 6 July 2021
/
Accepted: 15 July 2021
/
Published: 21 July 2021
(This article belongs to the Special Issue Remote Sensing Applications for Sea Turtle Conservation)
Abstract
:Surveying the breeding population of a given species can be difficult for many logistic reasons. Marine turtles are a challenging taxon for the study of reproductive ecology and breeding strategies, because turtles aggregate off-shore and males remain exclusively at sea. For successful management of sea turtle populations, determining operational sex ratios (OSRs) on a continuing basis is critical for determining long-term population viability, particularly in the context of changing hatchling sex ratios due to temperature-dependent sex determination in a warming climate. To understand how survey technique and stage of the breeding season might influence the ability to detect turtles and determine OSRs, we surveyed the presence and identified the sex of adult male and female green sea turtles (Chelonia mydas) using a boat and small commercial unoccupied/unmanned aerial vehicle (UAV), at the start (October) and peak (December) of a nesting season at an important breeding site at Heron Island, Great Barrier Reef, Australia. The ratio of males to females within the breeding ground detected by both survey methods changed from being male-biased in October to heavily female-biased in December, indicating that most males cease their reproductive effort and depart before the peak of the nesting season. Surveying with a UAV more than doubled the rate of turtles seen per minute of survey effort compared with surveying solely from the boat and allowed surveys to be conducted at times and/or places unsafe or inaccessible for boats. The sex of a slightly greater proportion of turtles seen could not be identified by observers using a UAV versus a boat, although more turtles were detected using the UAV. The departure of many males during the peak of the nesting season is likely due to an increasing biological cost of residency in the area because males encounter fewer receptive females as the season progresses and the limited foraging opportunity is insufficient to support the number of males present. Overall, we found that UAVs are an effective tool for studying important but difficult to observe aspects of sea turtle biology.
1. Introduction
Determining the reproductive mode, mating strategies and reproductive phenology of animals provides fundamental information for understanding the reproductive capacity of a species and its inherent resilience or vulnerability to disturbance in the short or long term. The ratio of breeding males to breeding females in a population, termed the operational sex ratio (OSR), is a critical determinant of population viability and is especially important in endangered populations with environmentally-determined sex, such as sea turtles [1,2,3]. However, OSRs are complex and difficult to measure in sea turtle populations, because adult turtles are usually present at nesting areas only transiently [4] and counting males is logistically problematic due to their completely aquatic life history [3,5]. Surveying breeding turtles is most easily conducted on nesting females as they come onto the beach to nest, and OSRs are generally estimated from offspring sex ratios [6]. As all sea turtles exhibit temperature-dependent sex determination (TSD), there has been increasing concern over the effects that increasingly female-dominated sex ratios will have on population demographics due to warming of nesting environments by climate change, as males become a more limited resource [6,7,8,9]. However, the few studies of adult sea turtle OSRs that have been undertaken indicate that while hatchling sex ratios are typically female-biased, adult OSRs are maintained close to 1:1 by more frequent mating by males than females, illustrating the inherent complexities of OSRs in these species [3,6,10,11]. With projected sand temperatures reaching the limits of survivability [8,9] and the uncertainty inherent in future climate predictions, there is a need for both a greater understanding of the complexities of OSRs in sea turtle populations and adaptive monitoring to understand if management interventions are presently necessary or will become so in the future [12,13]. Continual monitoring of sea turtle populations, especially breeding populations, is therefore critical in order to detect, manage, and anticipate future changes to them [14].
Surveying breeding sea turtle populations at sea has typically been done by human spotters aboard boats or low-flying occupied aircraft [15,16]. Traditional plane surveys are generally highly costly, potentially dangerous and can disturb animals during courtship or mating [17,18]. Boat surveys are cheaper, but may disturb animals on approach, and tidal conditions in coastal breeding areas may make access in boats difficult [19]. Both plane and boat surveys can also have problems when standardising monitoring efforts between repeat surveys, as human spotters may vary in their observational abilities and thus data-collection accuracy [20,21].
A recent solution to some of these problems has been the adoption of unoccupied/unmanned aerial vehicles (UAVs) for ecological surveying, as advances in consumer portable electronic technology have decreased their size, limitations and cost considerably [22,23]. As a result, UAVs have filled an important gap in data acquisition services between boats, occupied aircraft and satellites [24,25,26]. The use of UAVs in wildlife research has expanded to include population counts [27,28,29], habitat mapping and assessment [30,31], radio-tracking [32], physiological and morphometric studies [33,34] and detecting potentially dangerous human-wildlife interactions [35,36]. In several studies, UAVs have matched or outperformed traditional land or water-based estimates of marine wildlife populations [29,37,38,39], and recommended protocols for reporting the methods used facilitate the standardisation of techniques and comparisons between studies [36]. Although not without the possibility of causing some disturbance to animals, UAV survey techniques can reduce the impact of observation relative to other more conventional techniques and therefore may provide more representative data on animal behaviour in a less-invasive manner [39,40]. For the study of sea turtles and other large marine vertebrates, UAVs have become increasingly widespread [41], utilised in studies from population estimates to behavioural monitoring, including the calculation of OSRs in a Mediterranean population of loggerhead turtles, Caretta caretta [11,19]. Making quantitative determinations of absolute turtle abundance at a given location is complex and requires estimating and correcting for the detection probability [41], which varies with a number of factors including local bathymetry, water turbidity and animal arrival and departure at the survey site. However, UAVs have advantages that can provide useful data for detecting relative changes over time. Consequently, for this study, we used UAVs to achieve two main objectives: (1) to determine the OSR of a breeding population of sea turtles at different times during the breeding season, and (2) to compare the effectiveness of boat and UAV survey techniques for the determination of OSR in a sea turtle population.
2. Materials and Methods
2.1. Study Area and Species
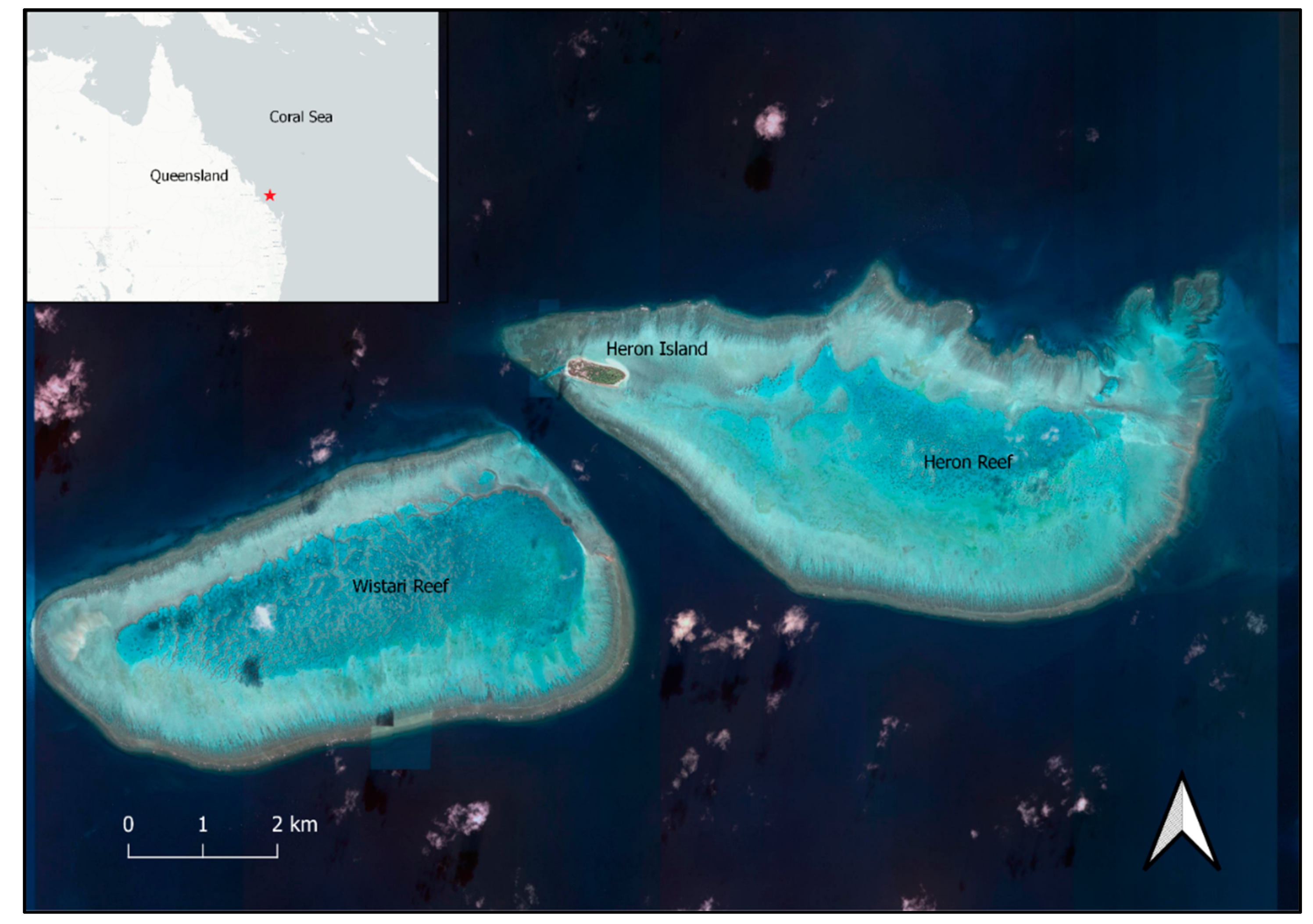
We undertook this study at Heron Reef and the adjacent Wistari Reef, in the southern Great Barrier Reef, Australia, at 23°26′S, 151°54′E. Waters around these reefs reach a depth of approximately 50 m, ascending to a reef crest that is exposed at high tide and encloses a relatively shallow lagoon (Figure 1). Within the lagoon, the water depth reaches a maximum of about 5 m at high tide. Heron Island is located at the western end of Heron Reef and is the nesting site of a large number of green turtles, Chelonia mydas, each austral summer. The Heron and Wistari Reefs do not have nearby seagrass beds which would sustain a large, permanent resident population of turtles, so most adult female and male turtles migrate from distant foraging grounds to this region of the southern Great Barrier Reef [42], where mating takes place and clutches of eggs are deposited on the island during darkness from late October until March, after which most animals disperse from the area [43]. While there are some resident small juvenile animals present year-round [44,45], this age-class remains omnivorous and not reliant on sea grasses [46,47]. Previous surveys in the area have captured and sexed adult animals in what would be considered the non-breeding season [48], although extensive surveys of loggerhead turtles in the Mediterranean have shown that male turtles can arrive at the breeding site almost 4 months in advance of the start of the breeding season and up to 6 months before the start of nesting [49], at a time of year that would result in them being designated as resident non-breeders had they not been observed to arrive. For these reasons, there was the possibility of detecting some non-breeding animals in our study, but the majority of adult-sized animals encountered during the breeding season can be considered to have aggregated for reproduction [50] in a polygamous mating system.
2.2. Survey Techniques
Boat and UAV surveys were undertaken on consecutive days from 17 to 21 October 2016 and from 3 to 5 December 2016. In all cases, the boat survey on a given day was immediately followed by the UAV survey or vice versa, so our two survey techniques were essentially contemporaneous. We launched the UAV from the boat, so were able to undertake both boat and UAV surveys without returning to the island in-between. Boat surveys were conducted using an outboard-powered 5 m rigid-hulled vessel inside the lagoon of the Heron and Wistari Reefs within 2 h either side of the high tide, in order for the water to be deep enough for the vessel to safely enter and exit the lagoon over the reef crest. In addition to the skipper, there were two or three turtle spotters looking for animals from the deck of the boat. Starting at a different location within the lagoon each day, we undertook a total of 540 min active surveying by boat in October 2016 and 190 min active surveying in December 2016. Time spent travelling to and from the survey location was not included in the survey duration. Active surveying was defined as all persons on the vessel visually searching the water around the boat up to a distance of approximately 50 m, looking for the visible shape and/or movement of a turtle in the water. The boat was driven at about 6 knots (approximately 11 km h−1) to maintain a general heading (typically north-south), but we did not follow a predefined transect line, so we describe the survey as being semi-haphazard. When a possible turtle was spotted, the skipper moved the boat closer to the animal in order to determine from the tail whether it was male or female. Mature male turtles have a long tail that extends beyond the rear flippers (Supplementary Video S2), while the tail of females is much shorter and extends only slightly beyond the rear edge of the carapace. If the tail of the turtle could not be clearly seen because of water depth or if the turtle left the area before the sex was unequivocally determined, then it was designated as being of unknown sex. A mating pair was recorded if we observed a male and female turtle copulating at the surface of the water. Each day we began the transect approximately 1 km from the end of the transect the previous day. For reasons of boating safety, we did not repeatedly cross the reef crest during an individual survey, in order to reduce the possibility of striking shallow coral outcroppings on the crest itself. Because we did not capture and mark turtles during the surveys or use some form of in-water photo identification, we could not exclude the possibility that the same animal was seen more than once on a single survey. However, the response of turtles to the boat and their movement rates make it unlikely that this was a common occurrence [5,51].
UAV surveys were conducted using a DJI Phantom 3 Pro quadcopter UAV (approx. 1.3 kg mass, DJI, Shenzhen, China), controlled with the DJI GO app on an Apple iPad 2 (Apple Inc., Cupertino, CA, USA). Our selection of UAV was based on a combination of its size, ease of operation, ability to be safely launched and landed on a small boat at sea, effective range, battery life and low impact on animals. While surveying, we recorded video at FHD 1920 × 780 pixel resolution and 25 frames per second with a UV filter and polarising filter fitted to the UAV’s inbuilt camera. The camera sensor size was 6.1 mm × 4.6 mm, 12.6 megapixels, and the field of view at typical survey altitude was approximately 100 metres horizontally. Video was streamed to the iPad and simultaneously recorded by the UAV’s on-board SD card for later retrieval. We selected FHD resolution as a compromise between the image resolution and the transfer speed and storage capacity of the SD card. The duration of a single flight was typically up to 18 min and we changed batteries between flights so that we could achieve a maximum of up to about 50 min surveying per trip. The UAV was launched from the upstretched, gloved hands of the pilot’s assistant at the front of an anchored boat and at the conclusion of the flight was landed in the same way. No other specialised launching or retrieval equipment was required. We attached small parabolic reflectors to the antennae of the UAV remote control in order to increase the range over which a clear signal could be transmitted between the UAV and remote. A typical UAV survey was undertaken at about 25–30 m elevation, with horizontal velocity of about 4 m s−1 (14.4 km h−1) and with the camera tilted downward at approximately 30 degrees below horizontal (Supplementary Video S1). The UAV pilot monitored the video signal being received on the iPad and looked for objects of turtle size and shape. When one was observed, the pilot tilted the camera to about 90 degrees below horizontal and descended the UAV to about 8–10 m elevation in order to verify the sex, after which the survey was resumed on the original path, altitude and camera tilt. Most UAV surveys followed an approximately linear flight path of two roughly parallel transects, with deviations to approach turtles to sex them as noted above, thus overall we would describe it as semi-haphazard. An example flight path and associated still video images are shown in Figure 2. We surveyed with the UAV for a total of 221 min in October 2016 and 132 min in December 2016. On any given survey day, we undertook the UAV survey immediately before or immediately after the boat survey, moving away from the end of the immediately prior survey by at least 500 m. Thus, because the entry of the boat into the lagoon was constrained to two hours either side of the high tides, our UAV surveys also occurred around this point of the tidal cycle. We could not identify individual animals, so although surveys were undertaken at different locations each day, we could not exclude the possibility that the same animal may have been seen more than once on a single survey and/or on more than one survey.
2.3. Survey Analysis
The number of detected female, male and unknown turtles on each boat survey was recorded and the total number of each in October and December was calculated by summing surveys in the October trips and the December trips respectively. The number of turtles seen in UAV surveys was determined from playback of the recorded video after the conclusion of the trips. Two viewers independently watched the unedited, unprocessed video at normal playback speed using VLC Media Player software (VideoLAN, Paris, France) on a computer monitor and, if necessary, paused the playback to identify the sex of an observed turtle from the length of its tail (Supplementary Video S2). We only considered a turtle as being detected when there was unequivocal movement of the animal relative to its surroundings, or visibility of the outline of the characteristically shaped flippers and carapace. In a few cases, there was a small discrepancy in the count of the two viewers, so a third viewer was used to determine the correct count. The OSR for each survey was determined from the ratio of identified male and female turtles only, and we did not include turtles for which we had been unable to determine sex. We used Two Sample t-tests to determine if there was a significant difference in the OSRs determined from boat and UAV surveys in October compared with December, and to detect differences in the number of turtles seen per minute of survey duration using the boat and UAV techniques. We determined the rate of turtle detections per unit time during surveys rather than per unit distance, because the transects were not straight lines, as noted above; we were interested in comparing the efficiency of the techniques in terms of surveyor effort, and we could not accurately quantify the total area observed in either of the techniques. For all statistical tests, significance was assumed at p < 0.05.
3. Results
Survey Results
Adult green sea turtles of both sexes and some of unknown sex were detected using the boat and UAV survey techniques with a total of 145 turtles encountered in the five boat and five UAV surveys in October 2016, including 24 mating pairs (Supplementary Video S3), and 155 turtles in total in the three boat and three UAV surveys in December 2016, including one mating pair (Table 1). Male and females were often detected at or near the surface while mating or swimming during boat surveys, while turtles could also be relatively easily seen below the surface in UAV imagery (Figure 3). Other turtles were sometimes observed on the video recordings that had not been seen by the UAV pilot at the time of the survey. In these cases, we were sometimes unable to determine the sex of the animal because the UAV’s altitude had been too high to capture sufficient detail, or the movement and/or orientation of the turtle, water clarity, depth or surface chop prevented a clear image being obtained. In order to reduce the incidence of the first limitation, we recommend recording video in UHD resolution or the maximum resolution of the equipment being used, to facilitate being able to zoom into an image on later viewing and sex the animal.
On both the boat surveys and the UAV surveys, the OSR (mean ± SD) in October (boat, 1.55 ± 0.27 males: 1 female, n = 5; UAV, 1.05 ± 0.30 males: 1 female, n = 5) was significantly higher than the mean OSR in December (boat, 0.11 ± 0.016 males: 1 female, n = 3, df = 5, t = 5.88, p = 0.002; UAV, 0.05 ± 0.0001 males: 1 female, n = 3, df = 5, t = 4.49, p = 0.006), thus there was a change from a male-skewed to a highly female-skewed sex ratio. Although a similar total number of turtles was seen in boat surveys (139 turtles) compared with UAV surveys (151 turtles), the UAV recorded on average about twice as many turtles per minute of survey than observers on the boat (UAV, 0.42 ± 0.10 turtles min−1, n = 8; boat, 0.20 ± 0.01 turtles min−1, n = 8, df = 8, t = −1.88, p = 0.049).
4. Discussion
Both boat surveys and UAV surveys appeared to be effective methods for detecting turtles in the breeding aggregation at Heron Island, and we were able to detect many animals at two different points in the breeding season. Our results from both the boat and UAV surveys show that the OSR for this population changed significantly during the breeding season, shifting from being male-skewed at the start of the season to being strongly female-skewed seven to eight weeks later. The apparent departure of males during the peak of the nesting season is consistent with ecological theory surrounding the mating strategy of sea turtles. Laparoscopic examination of the gonads of foraging adult green turtles revealed that male turtles breed more frequently than females over time and varying environmental conditions, such as water temperature affecting foraging opportunities [52]. This has also been shown in leatherback, Dermochelys coriacea [53], and loggerhead sea turtles [6,11]. While some males may maintain year-round residence at a breeding site if foraging opportunities are present, most males migrate annually to breeding sites from remote foraging grounds at the beginning of the breeding season in order to maximize encounters with arriving receptive females [11,54,55]. By being able to store sperm from multiple partners, females maintain high clutch fertility throughout the nesting season without multiple matings, and only a very low rate of multiple paternity of clutches has been reported at Heron Island, indicating that a single successful mating per female is the norm [56]. Total female receptivity declines as the breeding season progresses [57], as most females have already mated [56,58] and there is a diminishing reproductive benefit for breeding males to remain and compete for females at breeding grounds, as successive mating attempts will have progressively lower chances of success [54,55]. If foraging opportunities are limited at the breeding ground, then the decreasing body condition of males further reduces their incentive to remain after the initial mating period. The sex ratio therefore shifts from male-biased to female-biased as the breeding season progresses, with annual variations in timing of this shift linked to changes in biotic and abiotic conditions [11,49,59]. As noted earlier, some non-breeding males may have been detected by our surveys as solitary animals not engaged in mating at the time of detection. However, the more than 90% reduction in the calculated operational sex ratios from the October surveys to the December surveys indicates that a great majority of animals we observed were non-resident breeders. We emphasise that calculating absolute values for the OSR is problematic in areas where some turtles may be resident unless individual detected animals can be categorically determined to be resident or non-resident. Otherwise, the more valuable interpretation lies in the relative OSR values over time as an indicator of the shift in proportional male vs female presence during the breeding season.
During the October surveys when males were more common, the total number of males observed was greater than observed females by more than 1.5:1 on boat surveys compared with being fewer in number than females in the UAV surveys. This difference suggests that boat surveys were potentially male-biased in OSR calculations, likely due to the relative ease of spotting mating pairs and possible attendant males on the surface versus individual male or female turtles travelling below it or resting on the sea floor. In these circumstances, it seems probable that unreceptive, already-mated females may be less frequently spotted from the boat because they are less likely to be seen at the surface with males, and may indeed actively avoid males and spend more time submerged on the bottom of the lagoon, as leatherback turtles have been observed to do [57]. Other authors have noted a similar bias in boat surveys towards more visible animals, with turtles marked with white paint at Raine Island, Australia, being more represented in boat surveys than in UAV or underwater video [60]. In our survey setting, we think that sex ratios determined from UAV surveys may more accurately represent the true ratio of males and females present at the time, because solitary and submerged turtles are more easily seen from above than from water level. It would be useful to undertake a series of UAV surveys simultaneous with boat surveys and along the same transects to observe the same area at the same time. This would allow direct comparison of the number and position in the water column of turtles detected by the two methods, although this would require additional personnel and possibly a larger boat, which might not be feasible in a shallow reef environment. We used the same sampling protocols for both boat and UAV surveys to estimate the OSR, and undertook boat and UAV surveys in the same proportions in October and December, so we think that our results reasonably represent the magnitude of change in the OSR between these two times, although we caution against using absolute values for OSRs determined from boat surveys. A much more extensive survey regime would be necessary to quantify the probability of detection for each survey type and to enable a correction factor to be calculated for each technique. This was beyond the scope of our available time and resources, so we focused on the feasibility and relative performance of each in detecting turtles and estimating changes in the OSR over time. We do not assume that we detected all turtles present in the surveys, but are confident in our comparisons of relative numbers of turtles of different sexes detected by UAVs in a reasonably homogenous environment over a brief time frame, because while the absolute detection probability was not known, it can be assumed to be similar if surveys are conducted in a consistent manner. Accurately determining the functional OSR in sea turtles is very complex [11] because of the simultaneous arrival and departure of both sexes and changing receptivity of females to mating, and it will require repeated sampling over the full course of a breeding season to refine our estimates and accurately account for the subtlety of these changing dynamics. However, while boat and UAV surveys may differ in their probability of detecting males, the OSR values we calculated from each of these survey techniques lie within the range of those reported elsewhere [3,11]. The presence of some resident animals also needs to be accurately quantified and these animals identified to account for any non-breeding adults that might be observed [61]. Shore-based observations from ground level or on some elevated platform may be a simple and cheap way to gain some information on the approximate timing of the arrival of male and female turtles at breeding sites in real time, thereby setting an approximate time frame over which the more involved and expensive at-sea survey techniques could be conducted most informatively. Such shore-based observations have been successfully used to collect informative data on the seasonality of mating and nesting of green turtles at Ascension Island, an important green turtle breeding site in the South Atlantic Ocean [55].
Boat and UAV surveys are unlikely to allow identification of unique individual turtles, raising the potential for repeat counting. However, repeat counting cannot be completely avoided unless animals are uniquely identified in some way, such as by photographic identification while snorkelling [3], or by being captured, marked and released, both of which involve more time and effort by surveyors and greater intervention with the animals in the case of capture and tagging. Schofield et al. [11] concluded that the likelihood of double counting loggerhead turtles at a breeding aggregation on a single transect was low based on breeding turtle movement rates [5], and we think that the same is likely for our field site and location. Nevertheless, we accept that possible double counting is an uncertainty in our data, but we aimed to reduce it by undertaking boat and UAV surveys in different locations each day. Assuming that the probability of recounting an animal with either boat or drone surveys is the same for each method on any given day and that the probability is equal for both sexes, we think that our relative comparisons of OSR over time are valid, as are comparisons of the number of animals spotted per unit time using boat and UAV survey techniques.
We undertook the same number of boat and UAV surveys, but required more survey time to count and accurately identify turtles by boat and thus only counted about half as many turtles per unit time by boat than when using the UAV, similar to other studies comparing these methodologies [11,60]. The wide observation window, aerial viewpoint and increased mobility of a UAV increases the probability of encountering sea turtles by chance compared to traditional boat-based observations [62]. Indeed, a UAV recording we took simultaneously with a boat survey revealed a turtle that was missed by boat observers and that appeared to dive in response to the presence of the boat (Supplementary Video S4). Further, the UAV also allowed surveys to be accomplished across a wider range of times of day, because safe access by boat to the lagoon at Heron Island was limited by daily tide conditions. The UAV overcame this limitation by being able to be launched by boat from outside the reef crest and flown into the lagoon, increasing the total available time for surveying. However, because we matched boat and UAV surveys temporally in order to be able to directly compare their effectiveness at finding turtles and we launched the UAV from the boat, in this study the surveys all occurred when the tide was full or almost full. At Heron Island, it is not feasible to launch UAV surveys of the entire lagoon solely from the island itself because of the limited range of the UAV, so a boat was still required. However, it was not necessary to cross the reef crest only at high tide, so a survey could be undertaken by boat on a falling tide and then by UAV from outside the reef crest on completion of the boat surveys. If no boat survey is being done inside the lagoon, then a UAV can be used at any time during daylight using this method. It would be useful to compare UAV surveys launched from outside the reef crest at all points of the tidal cycle, in order to determine if turtle presence inside the lagoon changes with the tide and whether there is an optimal sampling period as the tide rises and falls. The option to launch from deeper water means that, from a practical perspective, the UAV method offers greater flexibility in timing of surveys, greater safety by reducing the need to drive boats across shallow reef crests, allows easier detection of turtles on the reef crest (Supplementary Video S5), requires less engine run-time because the boat is stationary during the survey, and provides a permanent record of the survey through the recorded video data and associated flight information collected by the control application. A video record of the flight opens up the possibility that automated counting and sexing of turtles may be possible if a sufficiently discriminating computer-based protocol is developed. Further, these benefits are accompanied by a greater encounter rate of animals per unit time, at least in our setting.
Despite the many benefits of using UAVs to survey turtles and other animals, UAVs have their own limitations compared with other survey techniques. While it is relatively easy to observe a moving turtle in shallow water or to distinguish a turtle from surrounding rocks or coral by its shape or movement, it is unlikely that all animals within the survey location will be detected. Thus, for the purposes of determining absolute animal density or abundance, UAVs must be supplemented with some form of capture-mark-recapture method that quantitatively accounts for specific individuals. Nevertheless, the aerial vantage point of a UAV makes detecting an animal in the water easier than from an observer’s point of view on the deck of a small boat. In practical terms of when and where they can and cannot effectively survey, UAV operators must adhere to local regulations governing airspace restrictions, privacy and safety, and ethical considerations must be taken into account for animal and human welfare during data acquisition [40,63,64]. While initial studies on auditory sensitivity and direct observation of behavioural responsiveness have suggested UAVs may elicit little if any reaction from several sea turtle species when flown by trained pilots at altitudes at or above 10 m [62,65], continual reassessment of its effect on study species will be necessary as the technology continues to develop [66]. In regard to image quality, bright sunlight reflecting off the water leads to glares in video, as well as causing glare on the screen the operator uses to fly the UAV [67]. The effect of sun glare can be mitigated through the use of a polarising filter on the UAV camera lens, conducting surveys at times of high sun angle or adjusting the camera angle relative to the water’s surface, while glare can be reduced on the operator’s screen by use of First Person View googles and/or safely controlling the UAV from under a shade structure [60,68,69,70]. Proper flight and survey planning can reduce the issue of turbidity and wind strength during surveys, but these problems may be unavoidable in seasons or locations not suitable for UAV deployment [71]. While several studies have reviewed current best practices for the use of UAVs in the study of marine ecosystems and wildlife (e.g., [64,66]), as with the adoption of any technology for the study of wildlife, continual assessment of the ethical and logistical concerns involved with its use will be necessary as its use expands and the technology develops.
5. Conclusions
Schofield et al. [11] provided the first minimum estimate of male sea turtle numbers in a breeding population using similar UAV technology to this study, and our results have confirmed the utility of UAVs for this purpose. A greater understanding of the biology and demographics of males, such as the changing OSR during the breeding season that we detected, will be important for managing and conserving populations. Building on this need, our study provides further validation that the aforementioned benefits of utilising UAVs to study OSRs in breeding populations of sea turtles outweigh the potential costs, and that UAV surveys continue to be a promising technique in studying this critical but elusive aspect of these species’ biology. The ability to later review UAV footage can lead to an increase in survey success via additional sightings, and manipulation of image quality can better reveal identifying morphometrics, such as the characteristic long tail of male turtles [19,68]. The collection and storage of survey videos also allows this data to have a second-life as public outreach and education materials, which is highly beneficial for effective species management and conservation. Further, trends in UAV technology and its rapid adoption for the study of a variety of wildlife indicate a continual decrease in cost and increase in utility (e.g., increased battery life and methods of data acquisition) in years to come [22,38,72]. While boats and other survey methodologies will remain important for the study and management of wildlife, the UAV continues to prove to be an effective new apparatus in the toolbox of ecologists, managers and conservationists, and provides a superior alternative in many circumstances.
Supplementary Materials
Supplementary materials are publicly accessible at https://doi.org/10.26180/15019743. Supplementary Video S1: Sample video of UAV surveying and point-of-view. Supplementary Video S2: Male turtle seen and sexed on its tail length. Supplementary Video S3: Mating turtles. Supplementary Video S4: UAV-captured video of a boat survey showing a turtle missed by boat observers and its response to the presence of the boat as it passes by. Supplementary Video S5: Turtles aggregating on the shallow water of the reef crest in an area difficult and/or dangerous to survey by boat.
Author Contributions
Conceptualization, R.D.R. and R.S.M.; data curation, R.D.R.; formal analysis, R.D.R. and A.Y.-K.; investigation, R.D.R. and R.S.M.; methodology, R.D.R. and R.S.M.; writing—original draft, R.D.R. and A.Y.-K.; writing—review and editing, R.D.R., R.S.M. and A.Y.-K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data presented in the study are publicly accessible at https://doi.org/10.26180/15019689 (accessed on 20 July 2021).
Acknowledgments
We thank the staff of the University of Queensland Heron Island Research Station for facilities and boats. Thanks to Cassie Speakman and Audrey Prasetya for assistance with video analysis and to undergraduate students of Monash University for assistance with boat and UAV surveys. Work was conducted under animal ethics approval BSCI/2014/08 issued by the Biological Sciences Animal Ethics Committee of Monash University and permit G16/38312.1 issued by the Great Barrier Reef Marine Park Authority. We thank four anonymous reviewers for comments and suggestions that improved the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Berglund, A. The operational sex ratio influences choosiness in a pipefish. Behav. Ecol. 1994, 5, 254–258. [Google Scholar] [CrossRef]
- Steifetten, Ø.; Dale, S. Viability of an endangered population of ortolan buntings: The effect of a skewed operational sex ratio. Biol. Conserv. 2006, 132, 88–97. [Google Scholar] [CrossRef]
- Hays, G.C.; Fossette, S.; Katselidis, K.A.; Schofield, G.; Gravenor, M.B. Breeding periodicity for male sea turtles, operational sex ratios, and implications in the face of climate change. Conserv. Biol. 2010, 24, 1636–1643. [Google Scholar] [CrossRef]
- Reina, R.D.; Mayor, P.A.; Spotila, J.R.; Piedra, R.; Paladino, F.V. Nesting ecology of the leatherback turtle, Dermochelys coriacea, at Parque Nacional Marino Las Baulas, Costa Rica: 1988–1989 to 1999–2000. Copeia 2002, 2002, 653–664. [Google Scholar] [CrossRef]
- Schofield, G.; Hobson, V.J.; Fossette, S.; Lilley, M.K.S.; Katselidis, K.A.; Hays, G.C. BIODIVERSITY RESEARCH: Fidelity to foraging sites, consistency of migration routes and habitat modulation of home range by sea turtles. Divers. Distrib. 2010, 16, 840–853. [Google Scholar] [CrossRef]
- Hays, G.C.; Schofield, G.; Mazaris, A.D. Different male versus female breeding periodicity helps mitigate offspring sex ratio skews in sea turtles. Front. Mar. Sci. 2014, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Jensen, M.P.; Allen, C.D.; Eguchi, T.; Bell, I.P.; LaCasella, E.L.; Hilton, W.A.; Hof, C.A.M.; Dutton, P.H. Environmental warming and feminization of one of the largest sea turtle populations in the world. Curr. Biol. 2018, 28, 154–159.e4. [Google Scholar] [CrossRef] [Green Version]
- Booth, D.T.; Dunstan, A.; Bell, I.; Reina, R.D.; Tedeschi, J. Low male production at the world’s largest green turtle rookery. Mar. Ecol. Prog. Ser. 2020, 653, 181–190. [Google Scholar] [CrossRef]
- Laloe, J.-O.; Tedeschi, J.; Booth, D.T.; Bell, I.; Dunstan, A.; Reina, R.D.; Hays, G.C. Extreme rainfall events and cooling of sea turtle clutches: Implications in the face of climate warming. Ecol. Evol. 2020, 11, 560–565. [Google Scholar] [CrossRef]
- Godley, B.J.; Broderick, A.C.; Downie, J.R.; Glen, F.; Houghton, J.D.; Kirkwood, I.; Reece, S.; Hays, G.C. Thermal conditions in nests of loggerhead turtles: Further evidence suggesting female skewed sex ratios of hatchling production in the Mediterranean. J. Exp. Mar. Biol. Ecol. 2001, 263, 45–63. [Google Scholar] [CrossRef]
- Schofield, G.; Katselidis, K.A.; Lilley, M.K.S.; Reina, R.D.; Hays, G.C. Detecting elusive aspects of wildlife ecology using drones: New insights on the mating dynamics and operational sex ratios of sea turtles. Funct. Ecol. 2017, 31, 2310–2319. [Google Scholar] [CrossRef]
- Fuentes, M.; Hamann, M.; Limpus, C. Past, current and future thermal profiles of green turtle nesting grounds: Implications from climate change. J. Exp. Mar. Biol. Ecol. 2010, 383, 56–64. [Google Scholar] [CrossRef]
- Santidrián Tomillo, P.; Genovart, M.; Paladino, F.V.; Spotila, J.R.; Oro, D. Climate change overruns resilience conferred by temperature-dependent sex determination in sea turtles and threatens their survival. Glob. Chang. Biol. 2015, 21, 2980–2988. [Google Scholar] [CrossRef] [PubMed]
- Hamann, M.; Godfrey, M.H.; Seminoff, J.A.; Arthur, K.; Barata, P.C.R.; Bjorndal, K.A.; Bolten, A.B.; Broderick, A.C.; Campbell, L.M.; Carreras, C.; et al. Global research priorities for sea turtles: Informing management and conservation in the 21st century. Endang. Species Res. 2010, 11, 245–269. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, T.; Gerrodette, T.; Pitman, R.L.; Seminoff, J.A.; Dutton, P.H. At-sea density and abundance estimates of the olive ridley turtle Lepidochelys olivacea in the eastern tropical Pacific. Endang. Species Res. 2007, 3, 191–203. [Google Scholar] [CrossRef]
- Seminoff, J.A.; Eguchi, T.; Carretta, J.; Allen, C.D.; Prosperi, D.; Rangel, R.; Gilpatrick, J.W., Jr.; Forney, K.; Peckham, S.H. Loggerhead sea turtle abundance at a foraging hotspot in the eastern Pacific Ocean: Implications for at-sea conservation. Endang. Species Res. 2014, 24, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Frick, M.G.; Slay, C.K.; Quinn, C.A.; Windham-Reid, A.; Duley, P.A.; Ryder, C.M.; Morse, L.J. Aerial observations of courtship behavior in loggerhead sea turtles (Caretta caretta) from Southeastern Georgia and Northeastern Florida. J. Herpetol. 2000, 34, 153–158. [Google Scholar] [CrossRef]
- Colefax, A.P.; Butcher, P.A.; Kelaher, B.P. The potential for unmanned aerial vehicles (UAVs) to conduct marine fauna surveys in place of manned aircraft. ICES J. Mar. Sci. 2017, 75, 1–8. [Google Scholar] [CrossRef]
- Rees, A.F.; Avens, L.; Ballorain, K.; Bevan, E.; Broderick, A.C.; Carthy, R.; Duclos, G.; Heithaus, M.R.; Johnston, D.; Mangel, J.C.; et al. The potential of unmanned aerial systems for sea turtle research and conservation: A review and future directions. Endang. Species Res. 2018, 35, 81–100. [Google Scholar] [CrossRef] [Green Version]
- Hone, J. On bias, precision and accuracy in wildlife aerial surveys. Wildl. Res. 2008, 35, 253–257. [Google Scholar] [CrossRef]
- Robinson, N.J.; Bigelow, W.; Cuffley, J.; Gary, M.; Hoefer, S.; Mills, S.; Smith, A.; Miguel Blanco, A. Validating the use of drones for monitoring the abundance and behaviour of juvenile green sea turtles in mangrove creeks in The Bahamas. Testudo 2020, 9, 24–35. [Google Scholar]
- Hodgson, J.C.; Baylis, S.M.; Mott, R.; Herrod, A.; Clarke, R.H. Precision wildlife monitoring using unmanned aerial vehicles. Sci. Rep. 2016, 6, 22574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, L.P.; Wich, S.A. Dawn of drone ecology: Low-cost autonomous aerial vehicles for conservation. Trop. Conserv. Sci. 2012, 5, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Angliss, R.P.; Ferguson, M.C.; Hall, P.; Helker, V.; Kennedy, A.; Sformo, T. Comparing manned to unmanned aerial surveys for cetacean monitoring in the Arctic: Methods and operational results. J. Unman. Veh. Syst. 2018, 6, 109–127. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, M.C.; Angliss, R.P.; Kennedy, A.; Lynch, B.; Willoughby, A.; Helker, V.; Brower, A.A.; Clarke, J.T. Performance of manned and unmanned aerial surveys to collect visual data and imagery for estimating arctic cetacean density and associated uncertainty. J. Unman. Veh. Syst. 2018, 6, 128–154. [Google Scholar] [CrossRef]
- Johnston, D.W. Unoccupied aircraft systems in marine science and conservation. Ann. Rev. Mar. Sci. 2019, 11, 439–463. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, R.R.; Holmberg, R.; Dann, P. Looking without landing—Using remote piloted aircraft to monitor fur seal populations without disturbance. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Goebel, M.E.; Perryman, W.L.; Hinke, J.T.; Krause, D.J.; Hann, N.A.; Gardner, S.; LeRoi, D.J. A small unmanned aerial system for estimating abundance and size of Antarctic predators. Polar Biol. 2015, 38, 619–630. [Google Scholar] [CrossRef] [Green Version]
- Fudala, K.; Bialik, R.J. Breeding colony dynamics of Southern elephant seals at Patelnia Point, King George Island, Antarctica. Remote Sens. 2020, 12, 2964. [Google Scholar] [CrossRef]
- Castellanos-Galindo, G.A.; Casella, E.; Mejía-Rentería, J.C.; Rovere, A. Habitat mapping of remote coasts: Evaluating the usefulness of lightweight unmanned aerial vehicles for conservation and monitoring. Biol. Conserv. 2019, 239, 108282. [Google Scholar] [CrossRef]
- Yaney-Keller, A.; Santidrián Tomillo, P.; Marshall, J.M.; Paladino, F.V. Using Unmanned Aerial Systems (UAS) to assay mangrove estuaries on the Pacific coast of Costa Rica. PLoS ONE 2019, 14, e0217310. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, J.A.; Desrochers, A.; Aubry, Y.; Pace, P.; Bird, D.M. A low-cost technique for radio-tracking wildlife using a small standard unmanned aerial vehicle. J. Unman. Veh. Syst. 2017, 5, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Pirotta, V.; Smith, A.; Ostrowski, M.; Russell, D.; Jonsen, I.D.; Grech, A.; Harcourt, R. An economical custom-built drone for assessing whale health. Front. Mar. Sci. 2017, 4. [Google Scholar] [CrossRef]
- Durban, J.W.; Moore, M.J.; Chiang, G.; Hickmott, L.S.; Bocconcelli, A.; Howes, G.; Bahamonde, P.A.; Perryman, W.L.; LeRoi, D.J. Photogrammetry of blue whales with an unmanned hexacopter. Mar. Mamm. Sci. 2016, 32, 1510–1515. [Google Scholar] [CrossRef]
- Butcher, P.A.; Piddocke, T.P.; Colefax, A.P.; Hoade, B.; Peddemors, V.M.; Borg, L.; Cullis, B.R. Beach safety: Can drones provide a platform for sighting sharks? Wildl. Res. 2019, 46, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Barnas, A.F.; Chabot, D.; Hodgson, A.J.; Johnston, D.W.; Bird, D.M.; Ellis-Felege, S.N. A standardized protocol for reporting methods when using drones for wildlife research. J. Unman. Veh. Syst. 2020, 8, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Johnston, D.W.; Dale, J.; Murray, K.T.; Josephson, E.; Newton, E.; Wood, S. Comparing occupied and unoccupied aircraft surveys of wildlife populations: Assessing the gray seal (Halichoerus grypus) breeding colony on Muskeget Island, USA. J. Unman. Veh. Syst. 2017, 5, 178–191. [Google Scholar] [CrossRef]
- Hodgson, J.C.; Mott, R.; Baylis, S.M.; Pham, T.T.; Wotherspoon, S.; Kilpatrick, A.D.; Raja Segaran, R.; Reid, I.; Terauds, A.; Koh, L.P. Drones count wildlife more accurately and precisely than humans. Meth. Ecol. Evol. 2018, 9, 1160–1167. [Google Scholar] [CrossRef] [Green Version]
- Sorrell, K.J.; Clarke, R.H.; Holmberg, R.; McIntosh, R.R. Remotely piloted aircraft improve precision of capture–mark–resight population estimates of Australian fur seals. Ecosphere 2019, 10, e02812. [Google Scholar] [CrossRef] [Green Version]
- Krause, D.J.; Hinke, J.T.; Goebel, M.E.; Perryman, W.L. Drones minimize Antarctic predator responses relative to ground survey methods: An appeal for context in policy advice. Front. Mar. Sci. 2021, 8. [Google Scholar] [CrossRef]
- Schofield, G.; Esteban, N.; Katselidis, K.A.; Hays, G.C. Drones for research on sea turtles and other marine vertebrates—A review. Biol. Conserv. 2019, 238, 108214. [Google Scholar] [CrossRef]
- Read, T.C.; Wantiez, L.; Werry, J.M.; Farman, R.; Petro, G.; Limpus, C.J. Migrations of green turtles (Chelonia mydas) between nesting and foraging grounds across the Coral Sea. PLoS ONE 2014, 9, e100083. [Google Scholar] [CrossRef] [Green Version]
- Limpus, C.J. The green turtle, Chelonia mydas, in Queensland: Breeding males in the southern Great Barrier Reef. Wildl. Res. 1993, 20, 513–523. [Google Scholar] [CrossRef]
- Southwood, A.L.; Reina, R.D.; Jones, V.S.; Speakman, J.R.; Jones, D.R. Seasonal metabolism of juvenile green turtles (Chelonia mydas) at Heron Island, Australia. Can. J. Zool. 2006, 84, 125–135. [Google Scholar] [CrossRef]
- Southwood, A.L.; Reina, R.D.; Jones, V.S.; Jones, D.R. Seasonal diving patterns and body temperatures of juvenile green turtles at Heron Island, Australia. Can. J. Zool. 2003, 81, 1014–1024. [Google Scholar] [CrossRef]
- Amorocho, D.F.; Reina, R.D. Feeding ecology of the East Pacific green sea turtle, Chelonia mydas agassizii, at Gorgona National Park in Colombia. Endang. Species Res. 2007, 3, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Amorocho, D.F.; Reina, R.D. Intake passage time, digesta composition, and digestibility in East Pacific green turtles (Chelonia mydas agassizii) at Gorgona National Park, Colombian Pacific. J. Exp. Mar. Biol. Ecol. 2008, 360, 117–124. [Google Scholar] [CrossRef]
- FitzSimmons, N.N.; Moritz, C.; Moor, S.S. Conservation and dynamics of microsatellite loci over 300 million years of marine turtle evolution. Mol. Biol. Evol. 1995, 12, 432–440. [Google Scholar] [CrossRef] [Green Version]
- Schofield, G.; Scott, R.; Dimadi, A.; Fossette, S.; Katselidis, K.A.; Koutsoubas, D.; Lilley, M.K.S.; Pantis, J.D.; Karagouni, A.D.; Hays, G.C. Evidence-based marine protected area planning for a highly mobile endangered marine vertebrate. Biol. Conserv. 2013, 161, 101–109. [Google Scholar] [CrossRef]
- Jessop, T.S.; FitzSimmons, N.N.; Limpus, C.J.; Whittier, J.M. Interactions between behavior and plasma steroids within the scramble mating system of the promiscuous green turtle, Chelonia mydas. Horm. Behav. 1999, 36, 86–97. [Google Scholar] [CrossRef]
- Schofield, G.; Papafitsoros, K.; Haughey, R.; Katselidis, K. Aerial and underwater surveys reveal temporal variation in cleaning-station use by sea turtles at a temperate breeding area. Mar. Ecol. Prog. Ser. 2017, 575, 153–164. [Google Scholar] [CrossRef]
- Limpus, C.J.; Limpus, D.J.; Arthur, K.E.; Parmenter, C.J. Monitoring Green Turtle Population Dynamics in Shoalwater Bay: 2000–2004; Great Barrier Reef Marine Park Authority: Townsville, Australia, 2005; p. 60.
- James, M.C.; Andrea Ottensmeyer, C.; Myers, R.A. Identification of high-use habitat and threats to leatherback sea turtles in northern waters: New directions for conservation. Ecol. Lett. 2005, 8, 195–201. [Google Scholar] [CrossRef]
- Plotkin, P.T.; Owens, D.W.; Byles, R.A.; Patterson, R. Departure of male olive ridley turtles Lepidochelys olivacea from a nearshore breeding ground. Herpetologica 1996, 52, 1–7. [Google Scholar]
- Godley, B.J.; Broderick, A.C.; Frauenstein, R.; Glen, F.; Hays, G.C. Reproductive seasonality and sexual dimorphism in green turtles. Mar. Ecol. Prog. Ser. 2002, 226, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Fitzsimmons, N.N. Single paternity of clutches and sperm storage in the promiscuous green turtle (Chelonia mydas). Mol. Ecol. 1998, 7, 575–584. [Google Scholar] [CrossRef]
- Reina, R.D.; Abernathy, K.J.; Marshall, G.J.; Spotila, J.R. Respiratory frequency, dive behavior and social interactions of leatherback turtles, Dermochelys coriacea during the inter-nesting interval. J. Exp. Mar. Biol. Ecol. 2005, 316, 1–16. [Google Scholar] [CrossRef]
- Crim, J.L.; Spotila, L.D.; Spotila, J.R.; O’Connor, M.; Reina, R.D.; Williams, C.J.; Paladino, F.V. The leatherback turtle, Dermochelys coriacea, exhibits both polyandry and polygyny. Mol. Ecol. 2002, 11, 2097–2106. [Google Scholar] [CrossRef]
- Baker, J.R. The Evolution of Breeding Seasons; Oxford University Press: London, UK, 1938; pp. 161–177. [Google Scholar]
- Dunstan, A.; Robertson, K.; Fitzpatrick, R.; Pickford, J.; Meager, J. Use of unmanned aerial vehicles (UAVs) for mark-resight nesting population estimation of adult female green sea turtles at Raine Island. PLoS ONE 2020, 15, e0228524. [Google Scholar] [CrossRef]
- Fitzsimmons, N.N. Male Marine Turtles: Gene Flow, Philopatry and Mating Systems of the Green Turtle Chelonia mydas. Ph.D. Thesis, The University of Queensland, St Lucia, QL, Australia, August 1997. [Google Scholar]
- Bevan, E.; Wibbels, T.; Navarro, E.; Rosas, M.; Najera, B.M.Z.; Sarti, L.; Illescas, F.; Montana, J.; Peña, L.J.; Burchfield, P. Using unmanned aerial vehicle (UAV) technology for locating, identifying, and monitoring courtship and mating behavior in the green turtle (Chelonia mydas). Herpetol. Rev. 2016, 47, 27–32. [Google Scholar]
- Clarke, R. Understanding the drone epidemic. Comput. Law Sec. Rev. 2014, 30, 230–246. [Google Scholar] [CrossRef]
- Joyce, K.E.; Duce, S.; Leahy, S.M.; Leon, J.; Maier, S.W. Principles and practice of acquiring drone-based image data in marine environments. Mar. Freshw. Res. 2019, 70, 952–963. [Google Scholar] [CrossRef]
- Bevan, E.; Whiting, S.; Tucker, T.; Guinea, M.; Raith, A.; Douglas, R. Measuring behavioral responses of sea turtles, saltwater crocodiles, and crested terns to drone disturbance to define ethical operating thresholds. PLoS ONE 2018, 13, e0194460. [Google Scholar] [CrossRef] [PubMed]
- Raoult, V.; Colefax, A.P.; Allan, B.M.; Cagnazzi, D.; Castelblanco-Martínez, N.; Ierodiaconou, D.; Johnston, D.W.; Landeo-Yauri, S.; Lyons, M.; Pirotta, V.; et al. Operational protocols for the use of drones in marine animal research. Drones 2020, 4, 64. [Google Scholar] [CrossRef]
- Hodgson, A.; Kelly, N.; Peel, D. Unmanned Aerial Vehicles (UAVs) for Surveying Marine Fauna: A Dugong Case Study. PLoS ONE 2013, 8, e79556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, P.C.; Fleishman, A.B.; Klein, D.J.; McKown, M.W.; Bézy, V.S.; Lohmann, K.J.; Johnston, D.W. A convolutional neural network for detecting sea turtles in drone imagery. Meth. Ecol. Evol. 2019, 10, 345–355. [Google Scholar] [CrossRef]
- Hensel, E.; Wenclawski, S.; Layman, C.A. Using a small, consumer-grade drone to identify and count marine megafauna in shallow habitats. Lat. Am. J. Aquatic. Res. 2018, 46, 1025–1033. [Google Scholar] [CrossRef]
- Barreto, J.; Cajaíba, L.; Teixeira, J.B.; Nascimento, L.; Giacomo, A.; Barcelos, N.; Fettermann, T.; Martins, A. Drone-monitoring: Improving the detectability of threatened marine megafauna. Drones 2021, 5, 14. [Google Scholar] [CrossRef]
- Schaub, J.; Hunt, B.P.V.; Pakhomov, E.A.; Holmes, K.; Lu, Y.; Quayle, L. Using unmanned aerial vehicles (UAVs) to measure jellyfish aggregations. Mar. Ecol. Prog. Ser. 2018, 591, 29–36. [Google Scholar] [CrossRef]
- Fust, P.; Loos, J. Development perspectives for the application of autonomous, unmanned aerial systems (UASs) in wildlife conservation. Biol. Conserv. 2020, 241, 108380. [Google Scholar] [CrossRef]
Figure 1.
Heron Reef and Wistari Reef, southern Great Barrier Reef, Australia, approximately 23°26′S, 151°54′E. Green turtles, Chelonia mydas, aggregate for reproduction in the waters surrounding and within these reefs, with eggs deposited on Heron Island mainly from November to March each year. Map from Google Earth (2021).
Figure 1.
Heron Reef and Wistari Reef, southern Great Barrier Reef, Australia, approximately 23°26′S, 151°54′E. Green turtles, Chelonia mydas, aggregate for reproduction in the waters surrounding and within these reefs, with eggs deposited on Heron Island mainly from November to March each year. Map from Google Earth (2021).

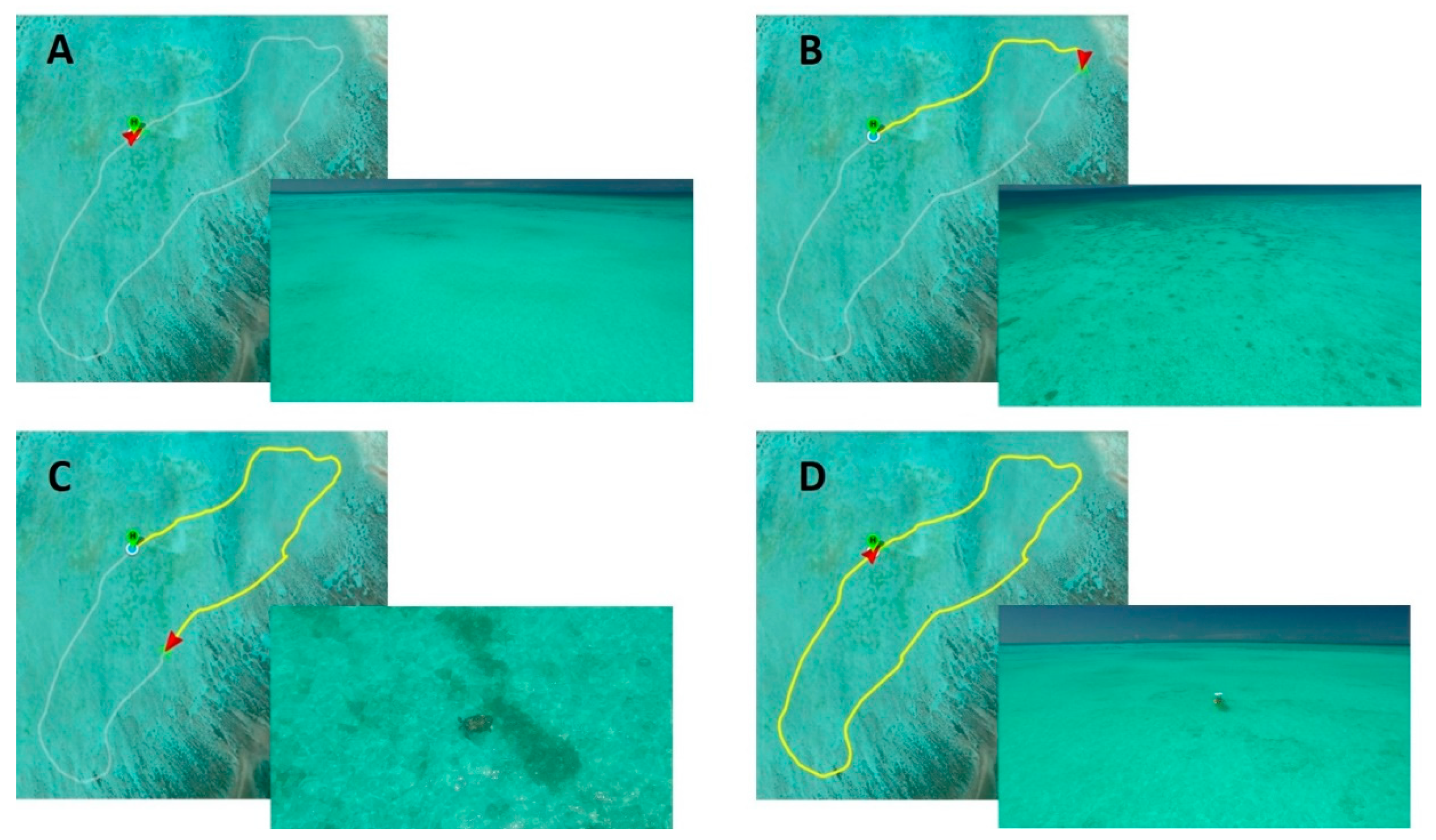
Figure 2.
Example flight path for an unoccupied/unmanned aerial vehicle (UAV) survey with accompanying still images from recorded video. The survey was undertaken in the south-eastern area of Heron Reef, with a total flight distance of about 3500 m and duration of 18 min as recorded by the DJI Go software. Parts (A–D) show the progress of the UAV along the survey transect (in yellow) with accompanying still images from the video recorded at that location on the flight path. Video still image (A) shows a predominately sand-bottomed area of the reef of about 3 m water depth, still image (B) shows sandy-bottom with increasing presence of small coral patches, still image (C) shows a green turtle, Chelonia mydas, observed at approximately 10 m elevation and still image (D) shows the UAV’s return to the survey boat at the end of the transect.
Figure 2.
Example flight path for an unoccupied/unmanned aerial vehicle (UAV) survey with accompanying still images from recorded video. The survey was undertaken in the south-eastern area of Heron Reef, with a total flight distance of about 3500 m and duration of 18 min as recorded by the DJI Go software. Parts (A–D) show the progress of the UAV along the survey transect (in yellow) with accompanying still images from the video recorded at that location on the flight path. Video still image (A) shows a predominately sand-bottomed area of the reef of about 3 m water depth, still image (B) shows sandy-bottom with increasing presence of small coral patches, still image (C) shows a green turtle, Chelonia mydas, observed at approximately 10 m elevation and still image (D) shows the UAV’s return to the survey boat at the end of the transect.

Figure 3.
Mating green turtles, Chelonia mydas, in the Heron Reef lagoon: (left) mounted male and female observed during a boat survey, (right) mounted male and female at upper left, with attendant male below right observed during UAV survey.
Figure 3.
Mating green turtles, Chelonia mydas, in the Heron Reef lagoon: (left) mounted male and female observed during a boat survey, (right) mounted male and female at upper left, with attendant male below right observed during UAV survey.


{kind=link}
{kind=link}
{kind=link}
Table 1.
The number of male, female and unknown sex green sea turtles, Chelonia mydas, observed on boat surveys and UAV surveys in Oct 2016 (n = 5 boat surveys and 5 UAV surveys) and Dec 2016 (n = 3 boat surveys and 3 UAV surveys) surveys on Heron Reef lagoon and Wistari Reef lagoon, Great Barrier Reef, Australia.
Table 1.
The number of male, female and unknown sex green sea turtles, Chelonia mydas, observed on boat surveys and UAV surveys in Oct 2016 (n = 5 boat surveys and 5 UAV surveys) and Dec 2016 (n = 3 boat surveys and 3 UAV surveys) surveys on Heron Reef lagoon and Wistari Reef lagoon, Great Barrier Reef, Australia.
| Boat Surveys | Drone Surveys | |||||
|---|---|---|---|---|---|---|
| Month | Male Turtles | Female Turtles | Unknown Sex | Male Turtles | Female Turtles | Unknown Sex |
| Oct 2016 | 49 | 30 | 11 | 20 | 27 | 8 |
| Dec 2016 | 4 | 44 | 1 | 4 | 83 | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yaney-Keller, A.; San Martin, R.; Reina, R.D. Comparison of UAV and Boat Surveys for Detecting Changes in Breeding Population Dynamics of Sea Turtles. Remote Sens. 2021, 13, 2857. https://doi.org/10.3390/rs13152857
AMA Style
Yaney-Keller A, San Martin R, Reina RD. Comparison of UAV and Boat Surveys for Detecting Changes in Breeding Population Dynamics of Sea Turtles. Remote Sensing. 2021; 13(15):2857. https://doi.org/10.3390/rs13152857
Chicago/Turabian StyleYaney-Keller, Adam, Ricardo San Martin, and Richard D. Reina. 2021. "Comparison of UAV and Boat Surveys for Detecting Changes in Breeding Population Dynamics of Sea Turtles" Remote Sensing 13, no. 15: 2857. https://doi.org/10.3390/rs13152857
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.