How Far Can We Classify Macroalgae Remotely? An Example Using a New Spectral Library of Species from the South West Atlantic (Argentine Patagonia)

Abstract
:
1. Introduction
2. Materials and Methods
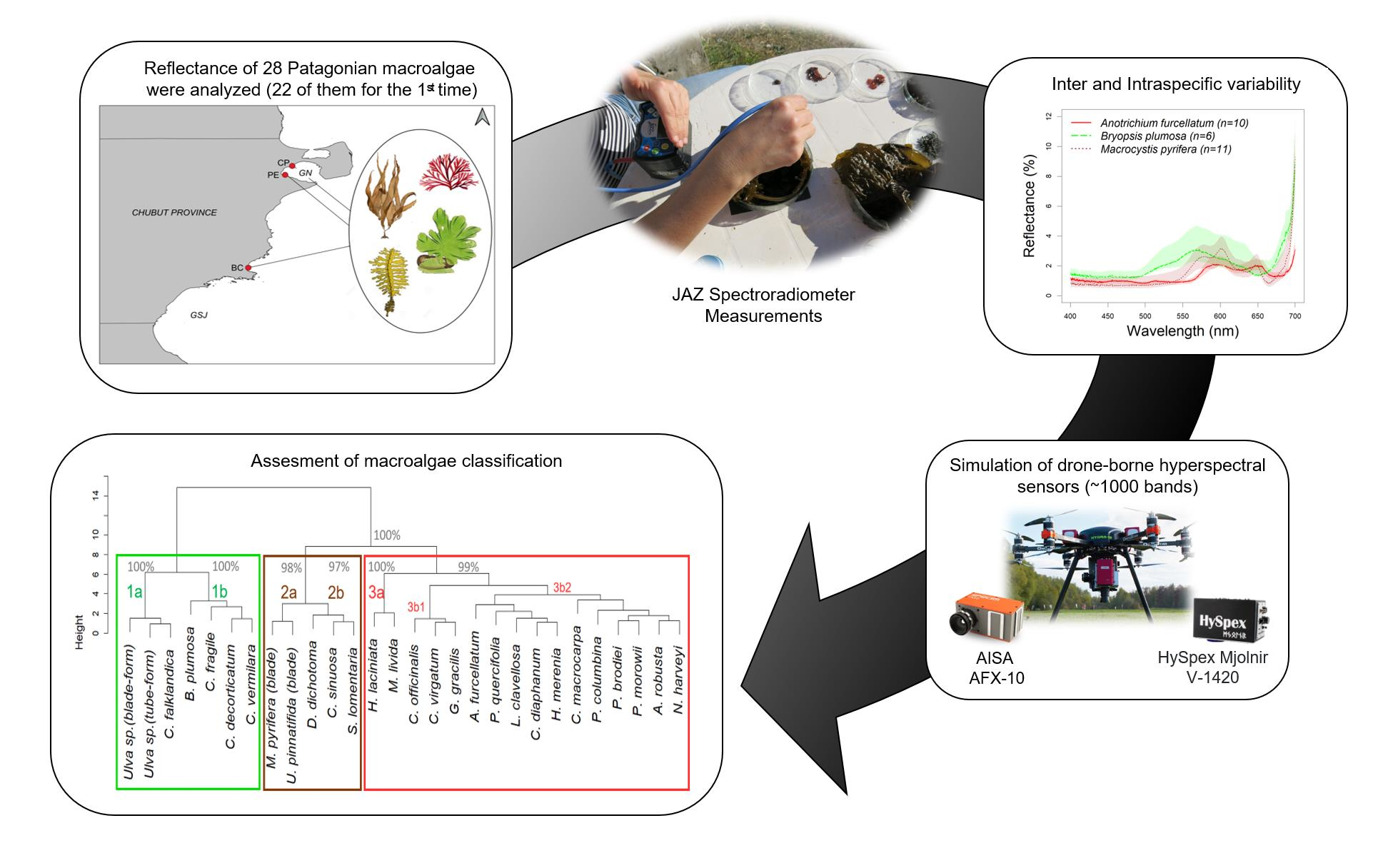
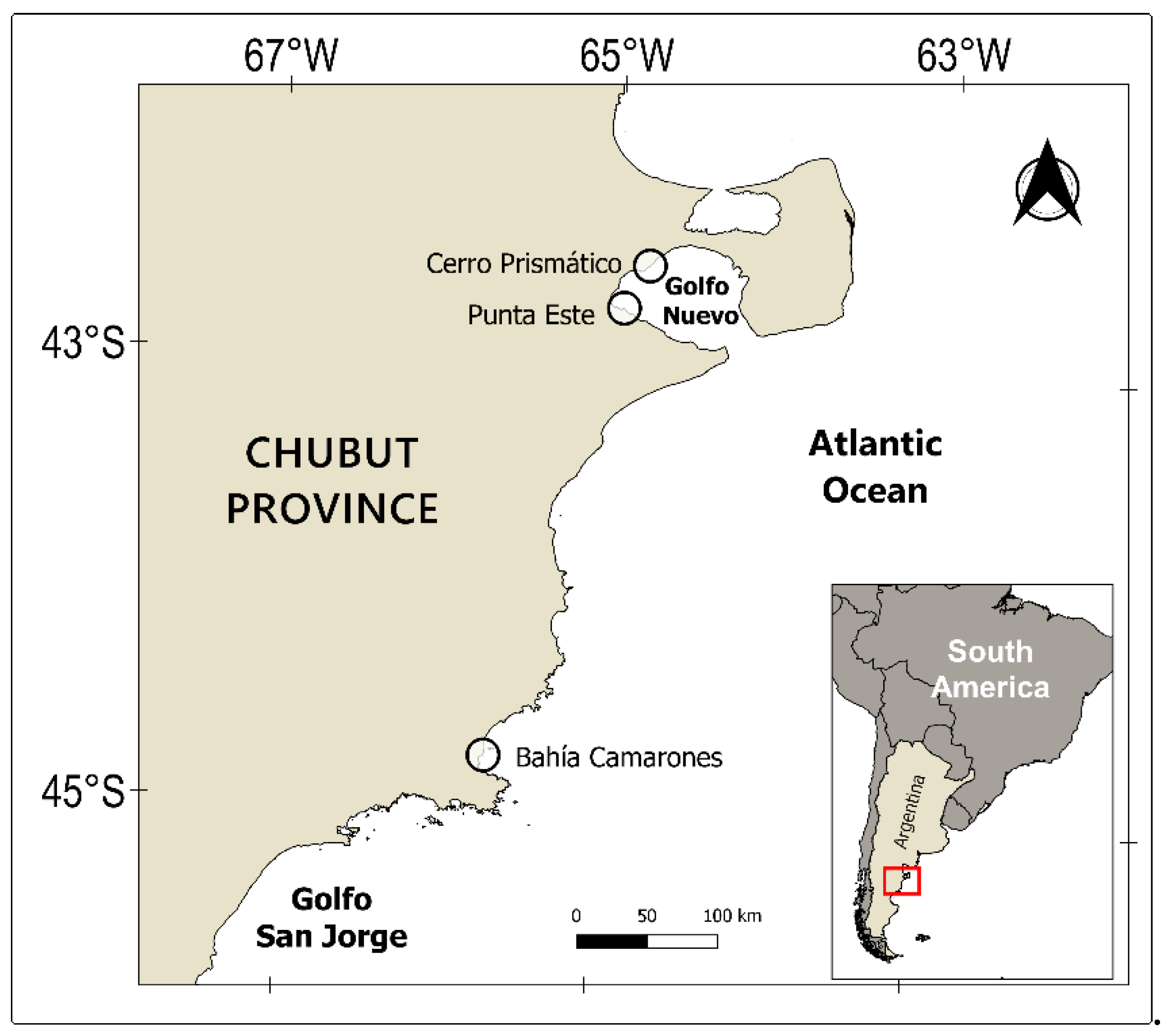
2.1. Study Area
2.2. Sample Collection, Identification and Handling
2.3. Spectral Data Collection
2.4. Data Analysis
2.5. Spectral Classification
2.6. Feature Identification
3. Results and Discussion
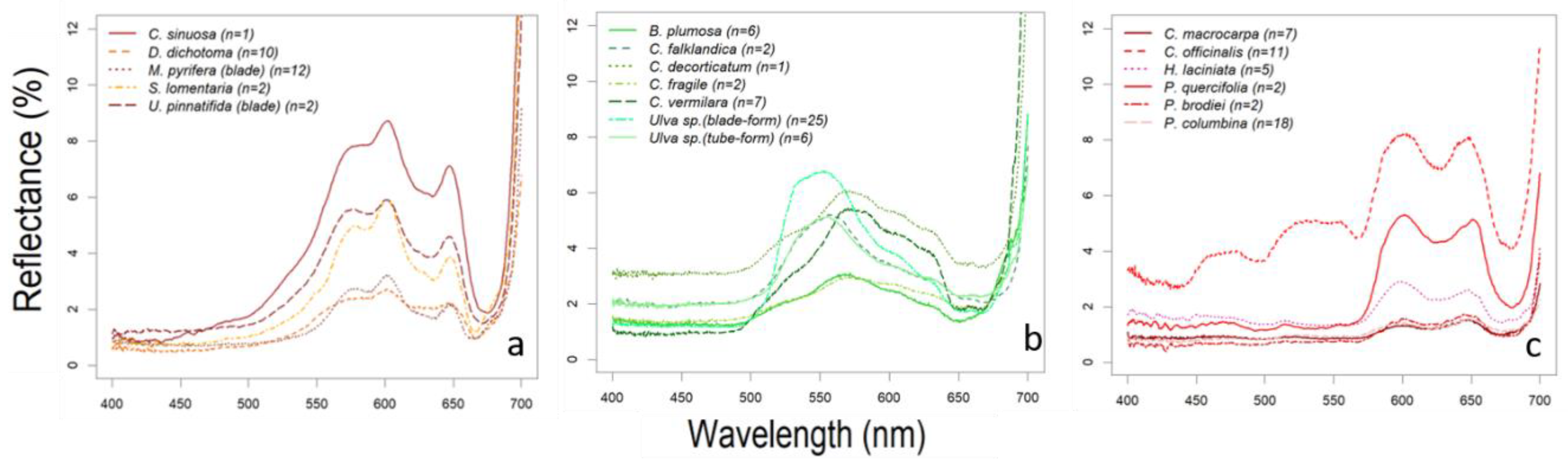
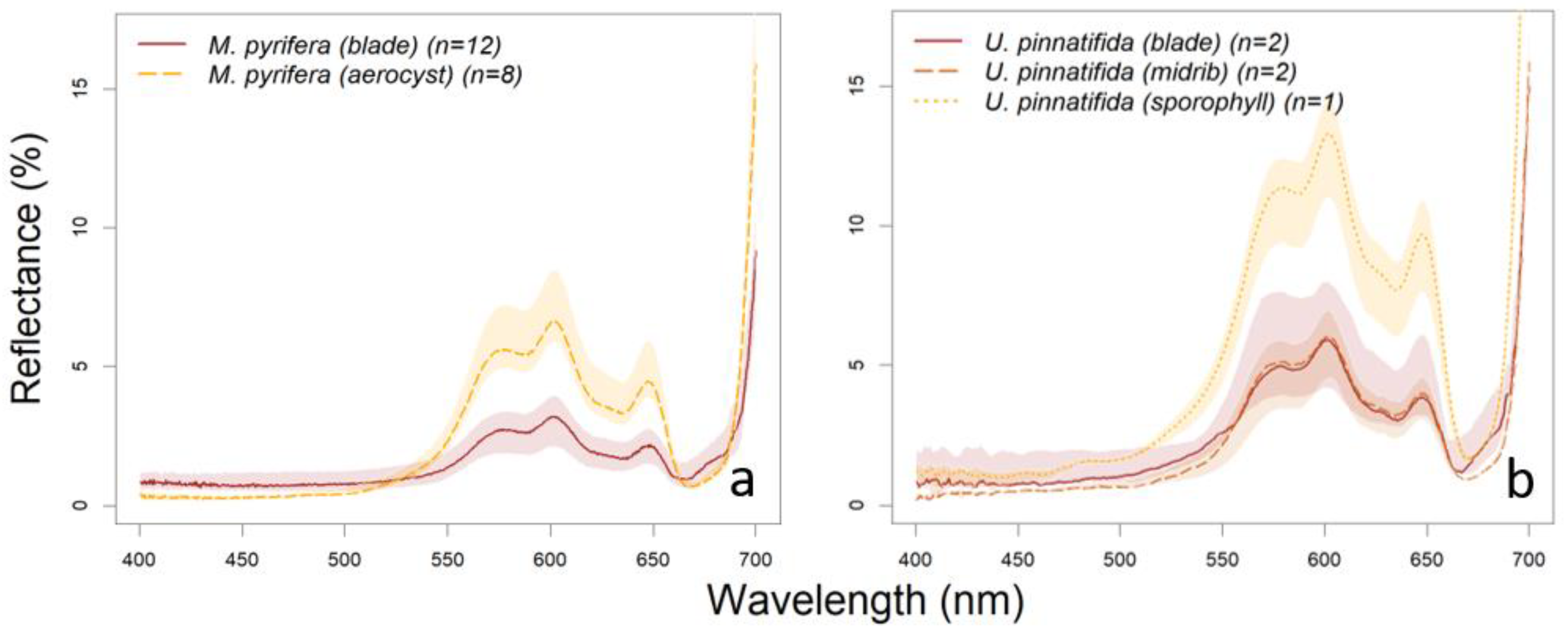
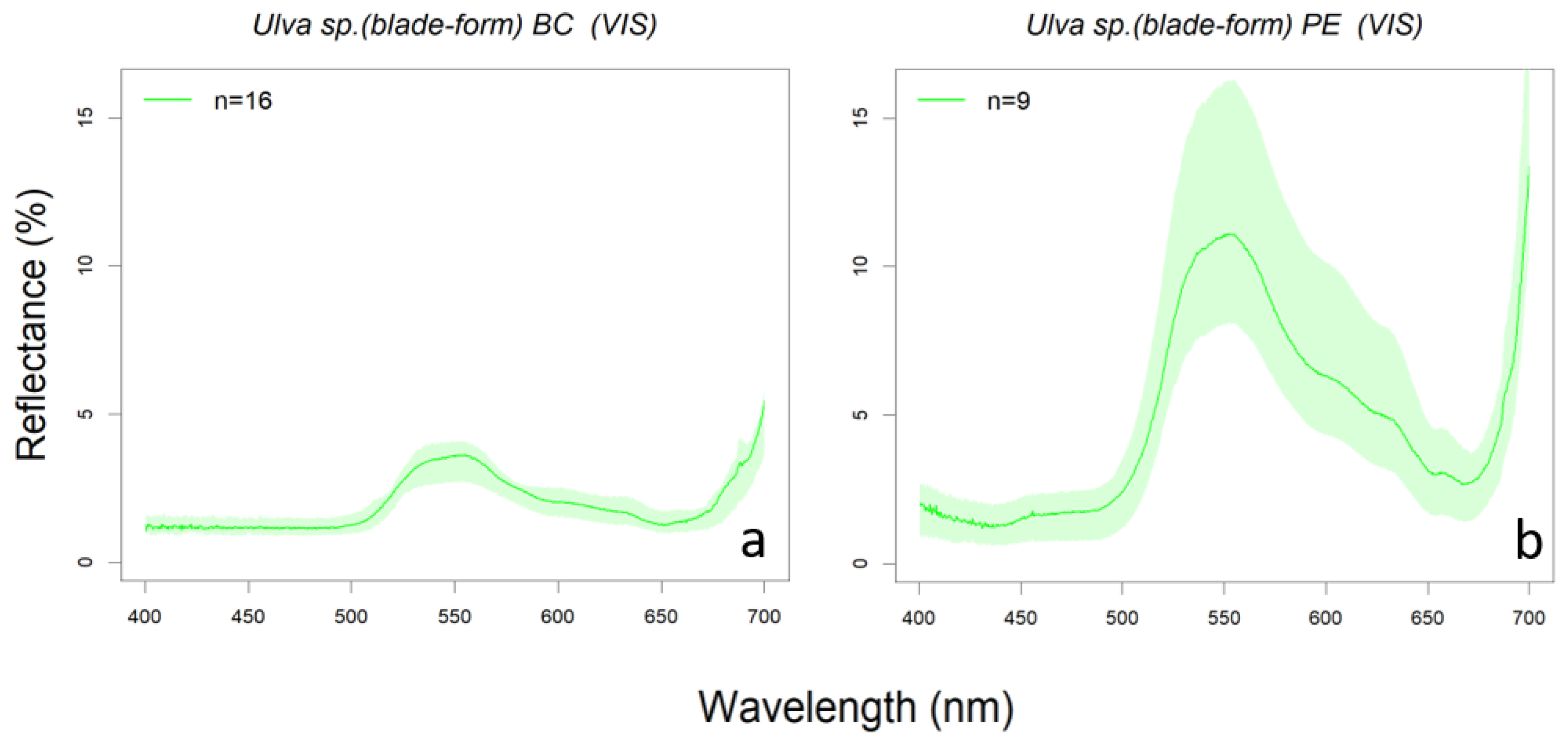
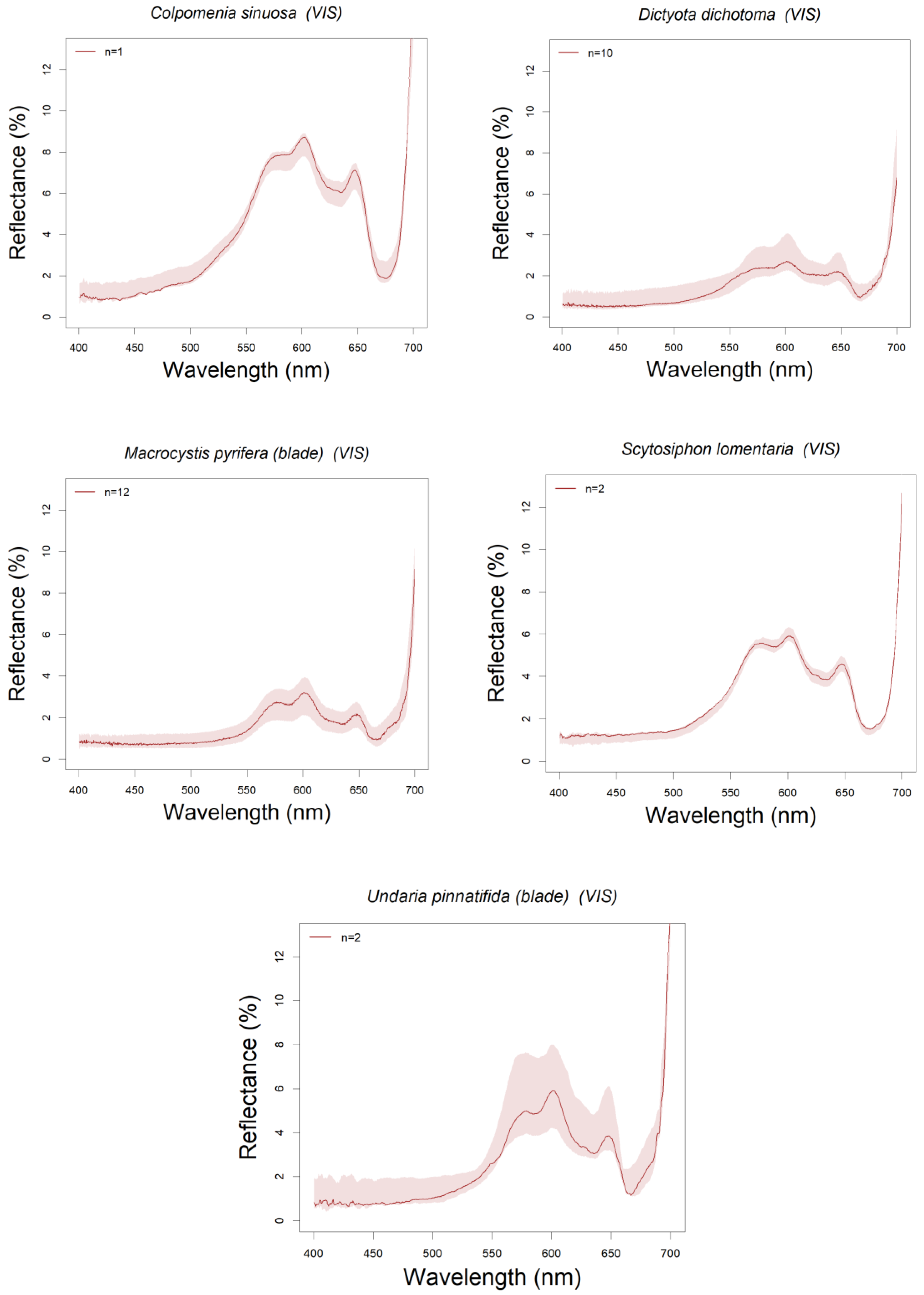
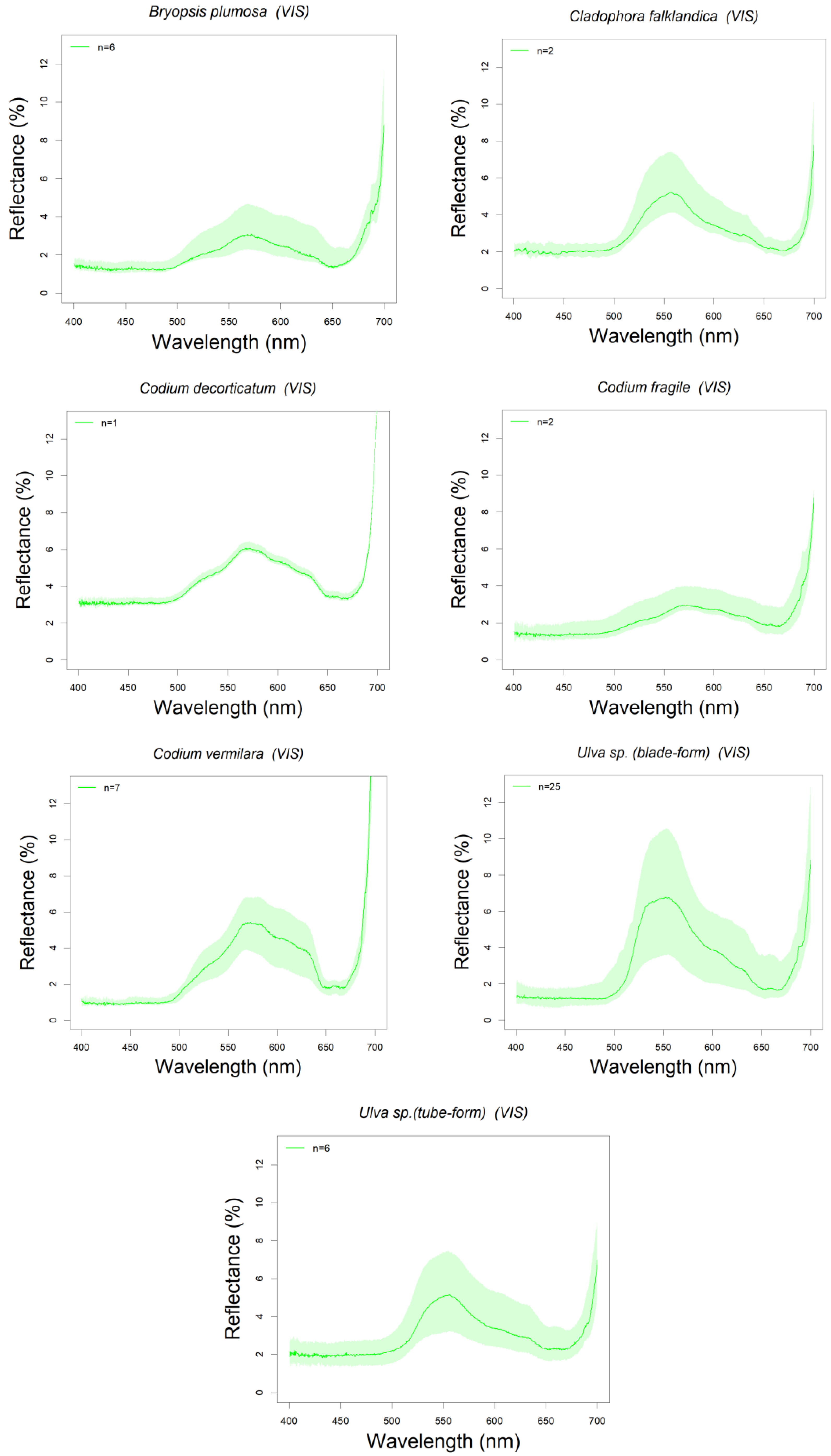
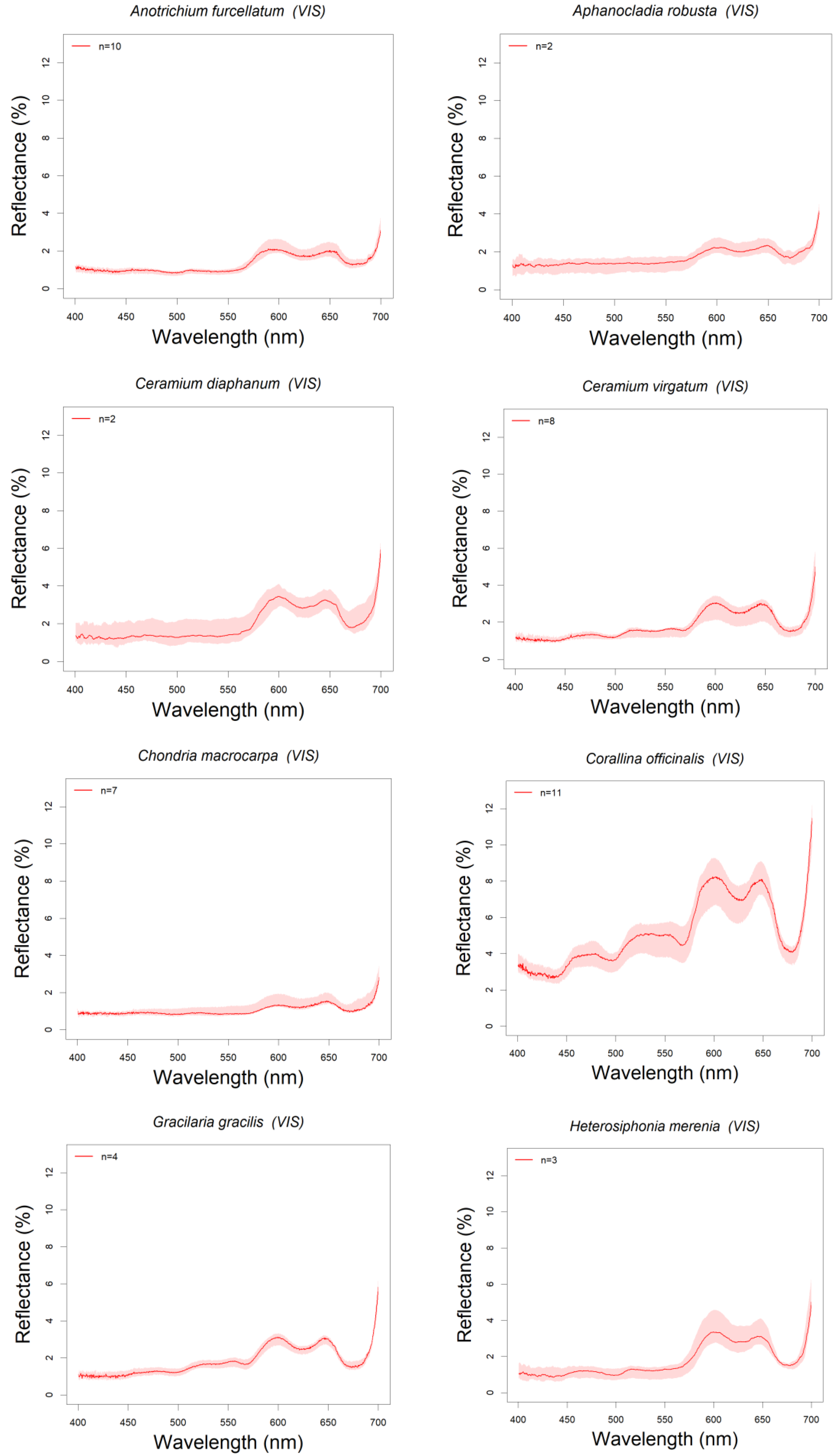
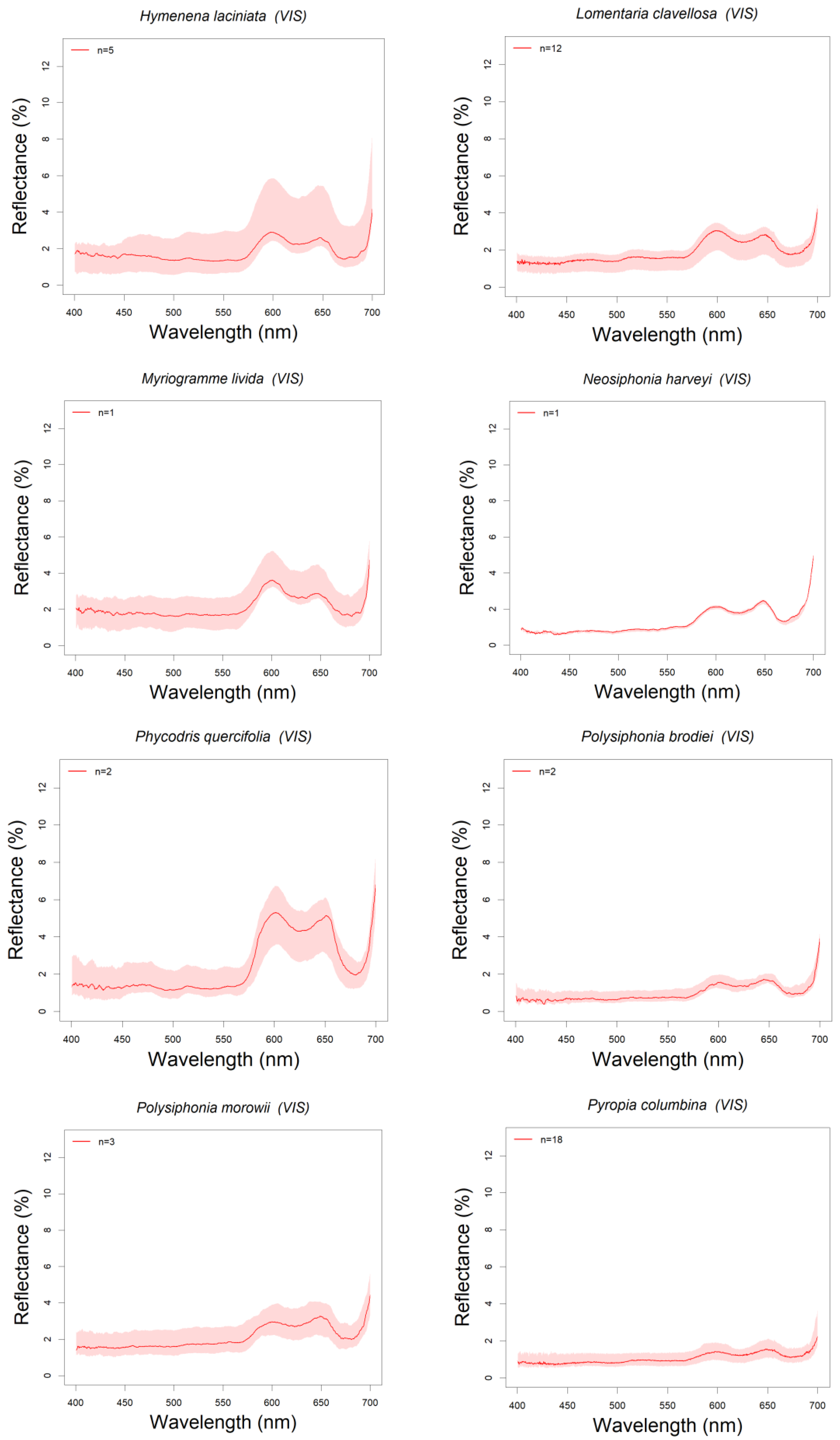
3.1. Library Description
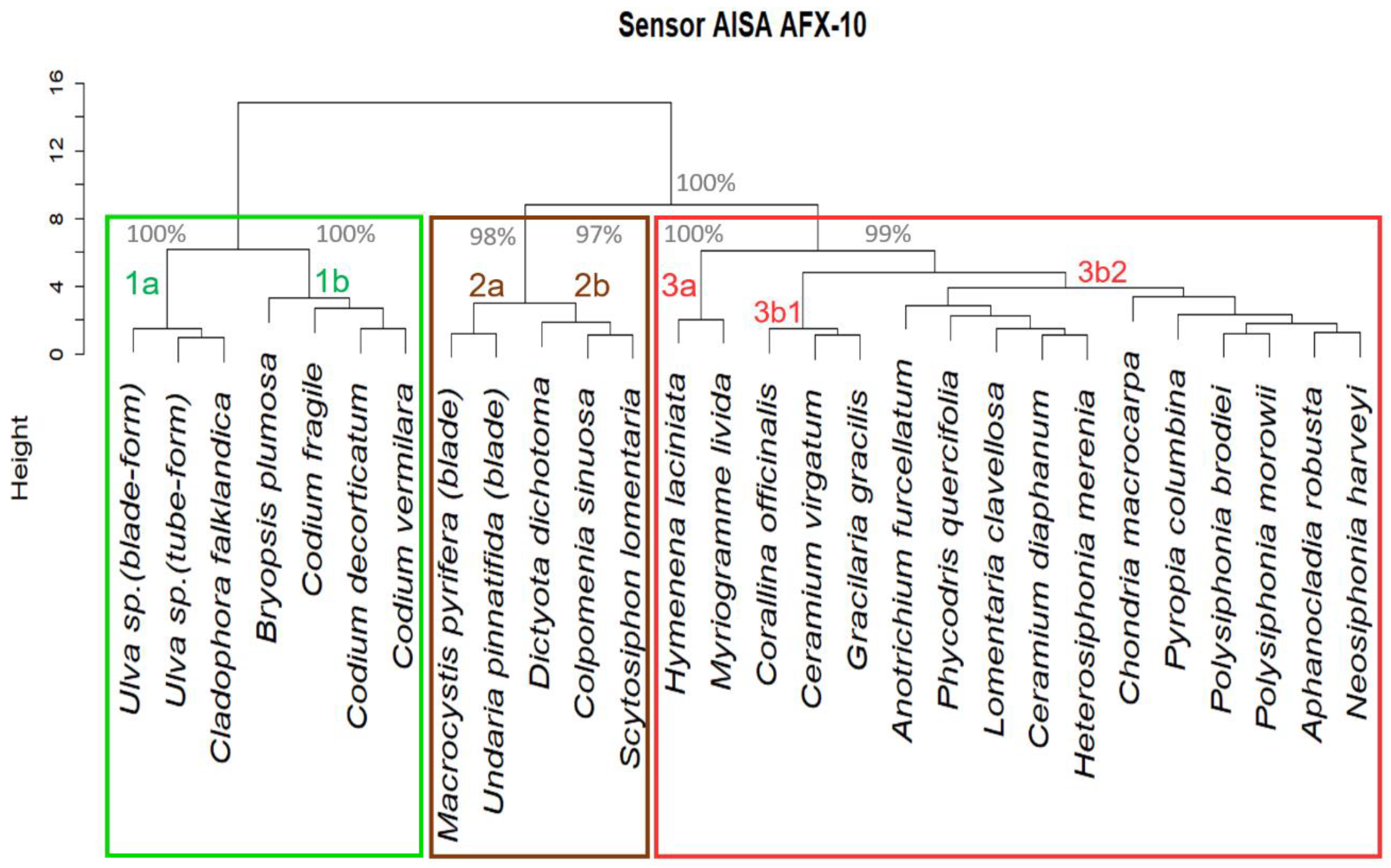
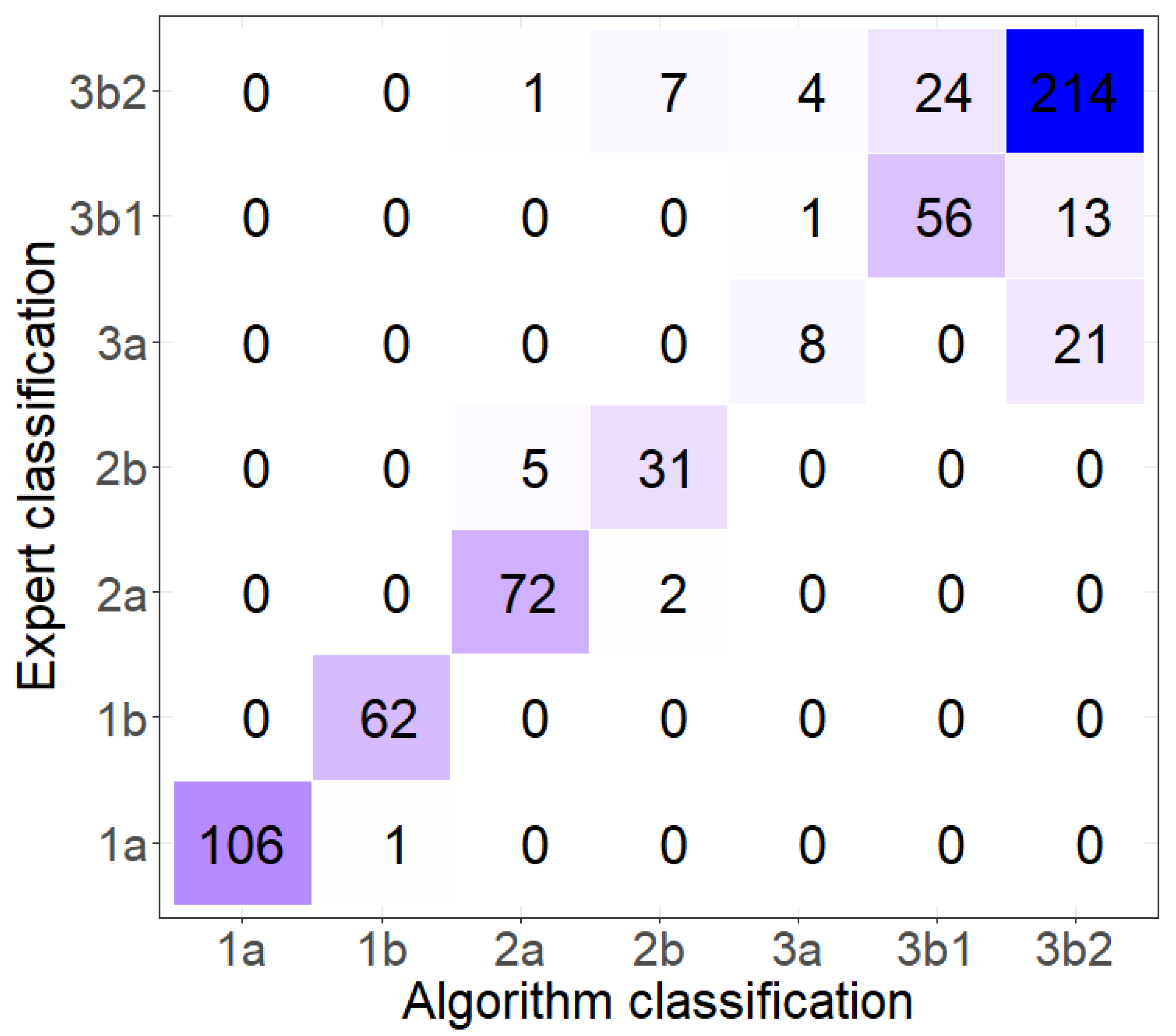
3.2. Algae Group/Classification by Hierarchical Cluster
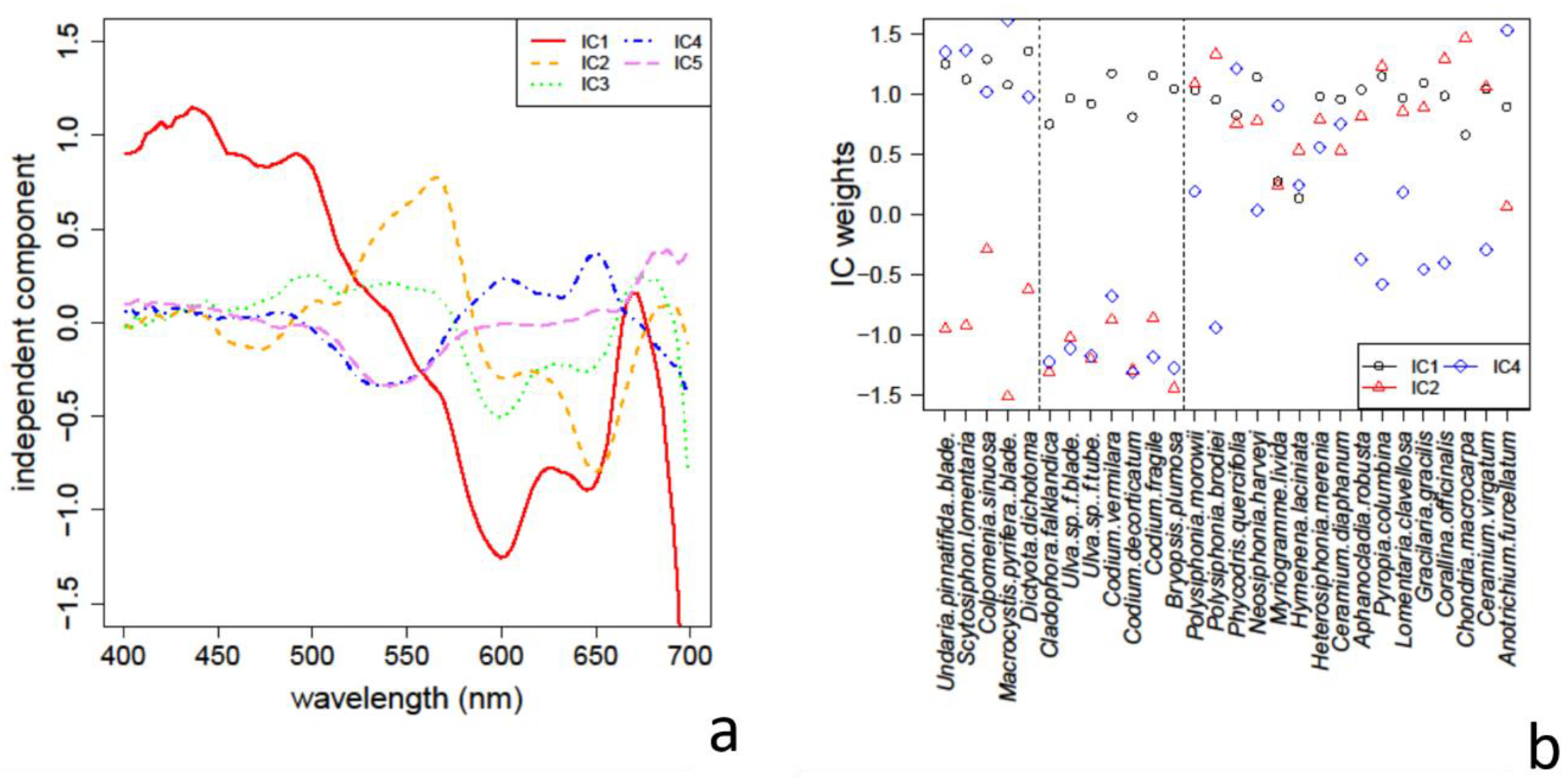
3.3. Absorption Feature Identification
3.4. Potential Applications and Future Work
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A




Appendix B



References
- Oppelt, N. Hyperspectral classification approaches for intertidal macroalgae habitat mapping: A case study in Heligoland. Opt. Eng. 2012, 51, 111703. [Google Scholar] [CrossRef]
- Wilson, K.L.; Kay, L.M.; Schmidt, A.L.; Lotze, H.K. Effects of increasing water temperatures on survival and growth of ecologically and economically important seaweeds in Atlantic Canada: Implications for climate change. Mar. Biol. 2015, 162, 2431–2444. [Google Scholar] [CrossRef]
- Harley, C.D.G.; Anderson, K.M.; Demes, K.W.; Jorve, J.P.; Kordas, R.L.; Coyle, T.A.; Graham, M.H. Effects of climate change on global seaweed communities. J. Phycol. 2012, 48, 1064–1078. [Google Scholar] [CrossRef] [PubMed]
- Rebours, C.; Marinho-Soriano, E.; Zertuche-González, J.A.; Hayashi, L.; Vásquez, J.A.; Kradolfer, P.; Soriano, G.; Ugarte, R.; Abreu, M.H.; Bay-Larsen, I.; et al. Seaweeds: An opportunity for wealth and sustainable livelihood for coastal communities. J. Appl. Phycol. 2014, 26, 1939–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, I.K.; Beardall, J.; Mehta, S.; Sahoo, D.; Stojkovic, S. Using marine macroalgae for carbon sequestration: A critical appraisal. J. Appl. Phycol. 2011, 23, 877–886. [Google Scholar] [CrossRef]
- Pallas, A.; Garcia-Calvo, B.; Corgos, A.; Bernardez, C.; Freire, J. Distribution and habitat use patterns of benthic decapod crustaceans in shallow waters: A comparative approach. Mar. Ecol. Prog. Ser. 2006, 324, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Shaffer, S. Preferential Use of Nearshore Kelp Habitats by Juvenile Salmon and Forage Fish. In Proceedings of the 2003 Georgia Basin/Puget Sound Research Conference, Vancouver, BC, Canada, 31 March–3 April 2003. [Google Scholar]
- Lorentsen, S.H.; Grémillet, D.; Nymoen, G.H. Annual variation in diet of breeding Great Cormorants: Does it reflect varying recruitment of Gadoids? Waterbirds 2004, 27, 161–169. [Google Scholar] [CrossRef]
- Buschmann, A.H.; Camus, C.; Infante, J.; Neori, A.; Israel, Á.; Hernández-González, M.C.; Pereda, S.V.; Gomez-Pinchetti, J.L.; Golberg, A.; Tadmor-Shalev, N.; et al. Seaweed production: Overview of the global state of exploitation, farming and emerging research activity. Eur. J. Phycol. 2017, 52, 391–406. [Google Scholar] [CrossRef]
- Vahtmäe, E.; Kutser, T. Classifying the Baltic Sea Shallow Water Habitats Using Image-Based and Spectral Library Methods. Remote Sens. 2013, 5, 2451–2474. [Google Scholar] [CrossRef] [Green Version]
- Mogstad, A.A.; Johnsen, G.; Ludvigsen, M. Shallow-Water Habitat Mapping using Underwater Hyperspectral Imaging from an Unmanned Surface Vehicle: A Pilot Study. Remote Sens. 2019, 11, 685. [Google Scholar] [CrossRef]
- Cavanaugh, K.; Siegel, D.; Reed, D.; Dennison, P. Environmental controls of giant-kelp biomass in the Santa Barbara Channel, California. Mar. Ecol. Prog. Ser. 2011, 429, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lõugas, L.; Kutser, T.; Kotta, J.; Vahtmäe, E. Detecting long time changes in benthic macroalgal cover using landsat image archive. Remote Sens. 2020, 12, 1901. [Google Scholar] [CrossRef]
- Glembocki, N.G.; Sánchez-Carnero, N.; Parma, A.M.; Orensanz, J.M. Remote Monitoring of a Remote Ecosystem-Pulsing Dynamics of Giant Kelp (Macrocystis pyrifera) Forests from Eastern Patagonia. in preparation.
- Dierssen, H.M.M.; Chlus, A.; Russell, B. Hyperspectral discrimination of floating mats of seagrass wrack and the macroalgae Sargassum in coastal waters of Greater Florida Bay using airborne remote sensing. Remote Sens. Environ. 2015, 167, 247–258. [Google Scholar] [CrossRef]
- Casal, G.; Kutser, T.; Domínguez-Gómez, J.A.; Sánchez-Carnero, N.; Freire, J. Mapping benthic macroalgal communities in the coastal zone using CHRIS-PROBA mode 2 images. Estuar. Coast. Shelf Sci. 2011, 94, 281–290. [Google Scholar] [CrossRef]
- Uhl, F.; Bartsch, I.; Oppelt, N. Submerged kelp detection with hyperspectral data. Remote Sens. 2016, 8, 487. [Google Scholar] [CrossRef] [Green Version]
- Vahtmäe, E.; Kutser, T.; Martin, G.; Kotta, J. Feasibility of hyperspectral remote sensing for mapping benthic macroalgal cover in turbid coastal waters—A Baltic Sea case study. Remote Sens. Environ. 2006, 101, 342–351. [Google Scholar] [CrossRef]
- Kislik, C.; Dronova, I.; Kelly, M. UAVs in Support of Algal Bloom Research: A Review of Current Applications and Future Opportunities. Drones 2018, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Rossiter, T.; Furey, T.; McCarthy, T.; Stengel, D.B. UAV-mounted hyperspectral mapping of intertidal macroalgae. Estuar. Coast. Shelf Sci. 2020, 242, 106789. [Google Scholar] [CrossRef]
- Tait, L.; Bind, J.; Charan-Dixon, H.; Hawes, I.; Pirker, J.; Schiel, D. Unmanned Aerial Vehicles (UAVs) for Monitoring Macroalgal Biodiversity: Comparison of RGB and Multispectral Imaging Sensors for Biodiversity Assessments. Remote Sens. 2019, 11, 2332. [Google Scholar] [CrossRef] [Green Version]
- Headwall Photonics Hyperspectral Inc. Hyperspectral Imaging Sensors: Nano-Hyperspec. Headwall Nano-Hyperspec. Available online: www.headwallphotonics.com/hyperspectral-sensors (accessed on 17 September 2020).
- Specim Spectral Imaging Ltd. Specim FX Series. Specim AFX Series. Available online: https://www.specim.fi/afx (accessed on 17 September 2020).
- Norsk Elektro Optikk AS HySpex Turnkey Solutions. Available online: https://www.hyspex.com (accessed on 17 September 2020).
- Brodie, J.; Ash, L.V.; Tittley, I.; Yesson, C. A comparison of multispectral aerial and satellite imagery for mapping intertidal seaweed communities. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 872–881. [Google Scholar] [CrossRef]
- Chao Rodríguez, Y.; Gómez Domínguez, J.A.; Sánchez-Carnero, N.; Rodríguez-Pérez, D. A comparison of spectral macroalgae taxa separability methods using an extensive spectral library. Algal Res. 2017, 26, 463–473. [Google Scholar] [CrossRef]
- Brodie, J.; Lewis, J. Introduction. In Unravelling the Algae: The Past, Present, and Future of Algal Systematics; System Association Special; Brodie, J., Lewis, J., Eds.; CRC Press: London, UK, 2007; Volume 75, pp. 1–4. ISBN 13:978-0-8493-7989-5. [Google Scholar]
- Mouritsen, O.G. The Biology of algae. In Seaweeds: Edible, Available, and Sustainable; University of Chicago Press: Chicago, IL, USA, 2013; pp. 21–37. ISBN 978-0-226-04453-8. [Google Scholar]
- Murtha, P.A. Detection and analysis of vegetation stresses. In Uses of Remote Sensing in Forest Pest Damage Appraisal, Proceedings of the a Seminar Held Inf. Rep. NOR-X-238, Edmonton, Alberta, 8 May 1981; Hall, R.J., Ed.; Canadian Forest Service Publications: Quebec, QC, Canada, 1982; pp. 2–24. [Google Scholar]
- Kutser, T.; Dekker, A.G.; Skirving, W. Modeling spectral discrimination of Great Barrier Reef benthic communities by remote sensing instruments. Limnol. Oceanogr. 2003, 48, 497–510. [Google Scholar] [CrossRef] [Green Version]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-wide electronic publication. Available online: http://www.algaebase.org (accessed on 25 August 2020).
- Boraso de Zaizso, A.L. Elementos Para el Estudio de las Macroalgas de Argentina, 1st ed.; Editorial Universitaria de la Patagonia (EDUPA): Comodoro Rivadavia, Argentina, 2013; ISBN 9789871937141. [Google Scholar]
- Eyras, M.C.; de Zaixso, A.L. Observaciones sobre la fertilidad de los esporofitos de Macrocystis pyrifera en la costa Argentina. Nat. Patagónica 1994, 2, 33–47. [Google Scholar]
- Liuzzi, M.G.; Gappa, J.L.; Piriz, M.L. Latitudinal gradients in macroalgal biodiversity in the Southwest Atlantic between 36 and 55° S. Hydrobiologia 2011, 673, 205–214. [Google Scholar] [CrossRef]
- Maritorena, S.; Morel, A.; Gentili, B. Diffuse reflectance of oceanic shallow waters: Influence of water depth and bottom albedo. Limnol. Oceanogr. 1994, 39, 1689–1703. [Google Scholar] [CrossRef]
- Anstee, J.M.; Dekker, A.G.; Byrne, G.T.; Daniel, P.; Held, A.; Miller, J. Hyperspectral Imaging for benthic species in shallow coastal waters. In Proceedings of the 10th Australian Remote Sensing and Photogrammetry Conference, Adelaide, Australia, 21–25 August 2000; pp. 1051–1061. [Google Scholar]
- Zimmerman, R.C.; Wittlinger, S.K. Hyperspectral Remote Sensing of Submerged Aquatic Vegetation in Optically Shallow Waters. In Proceedings of the Ocean Optics XV., Musée Océanographique, Monaco, 16–20 October 2000. CD-ROM Proc. Paper No. 1138, 6. [Google Scholar]
- Lubin, D.; Dustan, P.; Mazel, C.H.; Stannes, K. Spectral signatures of coral reefs: Feaures from space. Remote Sens. Environ. 2001, 75, 127–137. [Google Scholar] [CrossRef]
- Stekoll, M.S.; Deysher, L.E.; Hess, M. A remote sensing approach to estimating harvestable kelp biomass. J. Appl. Phycol. 2006, 18, 323–334. [Google Scholar] [CrossRef]
- Murfitt, S.L.; Allan, B.M.; Bellgrove, A.; Rattray, A.; Young, M.A.; Ierodiaconou, D. Applications of unmanned aerial vehicles in intertidal reef monitoring. Sci. Rep. 2017, 7, 10259. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Chehdi, K.; De Oliveira, E.; Cariou, C.; Charbonnier, B. Unsupervised Component Reduction of Hyperspectral Images and Clustering without Performance Loss: Application to Marine Algae Identification. In Image and Signal Processing for Remote Sensing XXII; International Society for Optics and Photonics: Edinburgh, UK, 18 October 2016; 100040Q. [Google Scholar]
- Schroeder, S.B.; Boyer, L.; Juanes, F.; Costa, M. Spatial and temporal persistence of nearshore kelp beds on the west coast of British Columbia, Canada using satellite remote sensing. Remote Sens. Ecol. Conserv. 2019, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Volent, Z.; Johnsen, G.; Sigernes, F. Kelp forest mapping by use of airborne hyperspectral imager. J. Appl. Remote Sens. 2007, 1, 011503. [Google Scholar] [CrossRef]
- Flynn, K.; Chapra, S. Remote Sensing of Submerged Aquatic Vegetation in a Shallow Non-Turbid River Using an Unmanned Aerial Vehicle. Remote Sens. 2014, 6, 12815–12836. [Google Scholar] [CrossRef] [Green Version]
- Kutser, T.; Metsamaa, L.; Vahtmäe, E.; Metsamaa, L. Spectral library of macroalgae and benthic substrates in Estonian coastal waters. Proc. Est. Acad. Sci. Biol. Ecol. 2006, 55, 329–340. [Google Scholar]
- Díaz, P.; López Gappa, J.J.; Piriz, M.L. Symptoms of eutrophication in intertidal macroalgal assemblages of Nuevo Gulf (Patagonia, Argentina). Bot. Mar. 2002, 45, 267–273. [Google Scholar] [CrossRef]
- Mendoza, M. Las Macroalgas Marinas Bentónicas De La Argentina. Cienc. Hoy 1999, 9, 40–49. [Google Scholar]
- Rivas, A.; Beier, E. Temperature and salinity fields in the northparagonic gulfs. Oceanol. Acta 1990, 13, 15–20. [Google Scholar]
- Esteves, J.L.; De Vido de Mattio, N. Influencia de Puerto Madryn en Bahía Nueva mediante salinidad y temperatura. Evidencia de fenómenos de Surgencia. Centro Nacional Patagónico-CONICET. Cent. Nac. Patagónico-CONICET 1980, 26, 1–40. [Google Scholar]
- NASA OBPG. MODIS Terra Level 3 SST MID-IR Annual 4km Nighttime V2019.0, Version 2019.0; PO.DAAC: California, CA, USA, 2020. [CrossRef]
- Casas, G.N.; Piriz, M.L. Surveys of Undaria pinnatifida (Laminariales, Phaeophyta) in Golfo Nuevo, Argentina. Hydrobiologia 1996, 326–327, 213–215. [Google Scholar] [CrossRef]
- Casas, G.N.; Piriz, M.L.; Parodi, E.R. Population features of the invasive kelp Undaria pinnatifida (Phaeophyceae: Laminariales) in Nuevo Gulf (Patagonia, Argentina). J. Mar. Biol. Assoc. U. K. 2008, 88, 21–28. [Google Scholar] [CrossRef]
- Irigoyen, A.J.; Trobbiani, G.; Sgarlatta, M.P.; Raffo, M.P. Effects of the alien algae Undaria pinnatifida (Phaeophyceae, Laminariales) on the diversity and abundance of benthic macrofauna in Golfo Nuevo (Patagonia, Argentina): Potential implications for local food webs. Biol. Invasions 2011, 13, 1521–1532. [Google Scholar] [CrossRef]
- Bianchi, A.A. Vertical stratification and air-sea CO2 fluxes in the Patagonian shelf. J. Geophys. Res. 2005, 110, C07003. [Google Scholar] [CrossRef] [Green Version]
- Boraso de Zaixsoa, A.; Ciancia, M.; Cerezo, A. The seaweed resources of Argentina. In Seaweed Resources of the World; Critchley, A.T., Ohno, M., Eds.; Japan International Cooperation Agency: Yokosuka, Japan, 1998; pp. 1–13. ISBN 90-75000-80-4. [Google Scholar]
- Hall, M.A.; de Zaixso, A.L. Ciclos de los Bosques de Macrocystis Pyrifera en Bahia Camarones, Provincia del Chubut, República Argentina. ECOSUR 1979, 6, 165–184. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 17 September 2020).
- Barnes, R.J.; Dhanoa, M.S.; Lister, S.J. Standard Normal Variate Transformation and De-Trending of Near-Infrared Diffuse Reflectance Spectra. Appl. Spectrosc. 1989, 43, 772–777. [Google Scholar] [CrossRef]
- Suzuki, R.; Shimodaira, H. Pvclust: An R package for assessing the uncertainty in hierarchical clustering. Bioinformatics 2006, 22, 1540–1542. [Google Scholar] [CrossRef]
- Lv, W.; Wang, X. Overview of Hyperspectral Image Classification. J. Sens. 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Clark, R.N.; Roush, T.L. Reflectance spectroscopy: Quantitative analysis techniques for remote sensing applications. J. Geophys. Res. Solid Earth 1984, 89, 6329–6340. [Google Scholar] [CrossRef]
- Tharwat, A. Independent component analysis: An introduction. Appl. Comput. Inform. 2020. [Google Scholar] [CrossRef]
- Helwig, N.E. ica: Independent Component Analysis. R Package Version 1.0-2. 2018. Available online: https://CRAN.R-project.org/package=ica (accessed on 17 September 2020).
- Du, H.; Qi, H.; Wang, X.; Ramanath, R.; Snyder, W.E. Band Selection Using Independent Component Analysis for Hyperspectral Image Processing. In Proceedings of the 32nd Applied Imagery Pattern Recognition Workshop, Washington, DC, USA, 15–17 October 2003; pp. 93–98. [Google Scholar]
- Carvalho, L.G.; Pereira, L. Review of marine algae as source of bioactive metabolites: A marine biotechnology approach. In Marine Algae: Biodiversity, Taxonomy, Environmental Assessment, and Biotechnology; Pereira, L., Neto, J.M., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 195–227. ISBN 978-1-4665-8181-4. [Google Scholar]
- Rowan, K.S. Photosynthetic Pigments of Algae, 1st ed.; Cambridge University Press: New York, NY, USA, 1989; ISBN 0-521-30176-9. [Google Scholar]
- Friedman, A.L.; Alberte, R.S. A Diatom Light-Harvesting Pigment-Protein Complex. Plant. Physiol. 1984, 76, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Friedman, A.L.; Alberte, R.S. Biogenesis and Light Regulation of the Major Light Harvesting Chlorophyll-Protein of Diatoms. Plant. Physiol. 1986, 80, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Tin, H.C.; O’Leary, M.; Fotedar, R.; Garcia, R. Spectral Response of Marine Submerged Aquatic Vegetation: A Case Study in Western Australia Coast. In Proceedings of the OCEANS 2015—MTS/IEEE Washington, Washington, DC, USA, 19–22 October 2015; pp. 1–5. [Google Scholar]
- Stengel, D.B.; Dring, M.J. Seasonal variation in the pigment content and photosynthesis of different thallus regions of Ascophyllum nodosum (Fucales, Phaeophyta) in relation to position in the canopy. Phycologia 1998, 37, 259–268. [Google Scholar] [CrossRef]
- Fyfe, S.K. Spatial and temporal variation in spectral reflectance: Are seagrass species spectrally distinct? Limnol. Oceanogr. 2003, 48, 464–479. [Google Scholar] [CrossRef]
- Kirk, J.T.O. Light and Photosynthesis in Aquatic Ecosystems, 2nd ed.; Cambridge University Press: New York, NY, USA, 2011; ISBN 978-0-521-15175-7. [Google Scholar]
- Schmid, M.; Stengel, D.B. Intra-thallus differentiation of fatty acid and pigment profiles in some temperate Fucales and Laminariales. J. Phycol. 2015, 51, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, C.; Ramlov, F.; de Lucena, L.A.F.; Uarrota, V.; Batista, M.B.; Sissini, M.N.; Oliveira, I.; Briani, B.; Martins, C.D.L.; de Nunes, J.M.C.; et al. UVR and PAR absorbing compounds of marine brown macroalgae along a latitudinal gradient of the Brazilian coast. J. Photochem. Photobiol. B Biol. 2018, 178, 165–174. [Google Scholar] [CrossRef]
- Jensen, J.R.; Estes, J.E.; Tinney, L. Remote sensing techniques for kelp surveys. Photogramm. Eng. Remote Sens. 1980, 46, 743–755. [Google Scholar]
- Cavanaugh, K.C.; Siegel, D.A.; Kinlan, B.P.; Reed, D.C. Scaling giant kelp field measurements to regional scales using satellite observations. Mar. Ecol. Prog. Ser. 2010, 403, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Arzee, T.; Polne, M.; Neushul, M.; Gibor, A. Morphogenetic aspects in Macrocystis development. Bot. Gaz. 1985, 146, 365–374. [Google Scholar] [CrossRef]
- Salavarria, E.; Benavente, M.; Kodaka, P.G. Histología de Macrocystis pyrifera (Linnaeus) C. Agardh 1820 (Phaeophyceae: Laminariales) en la costa centro del Perú. Arnaldoa 2014, 21, 69–80. [Google Scholar]
- Garbary, D.J.; Kim, K.Y. Anatomical Differentiation and Photosynthetic Adaptation in Brown Algae Anatomical Differentiation and Photosynthetic Adaptation in Brown Algae. Algae 2005, 20, 233–238. [Google Scholar] [CrossRef]
- Fernandes, F.; Barbosa, M.; Oliveira, A.P.; Azevedo, I.C.; Sousa-pinto, I.; Valentão, P.; Andrade, P.B. The pigments of kelps ( Ochrophyta ) as part of the flexible response to highly variable marine environments. J. Appl. Phycol. 2016, 3689–3696. [Google Scholar] [CrossRef]
- Castric-Fey, A.; Beaupoil, C.; Bouchain, J.; Pradier, E.; L’Hardy-Halos, M.T. The introduced alga Undaria pinnatifida (Laminariales, Alariaceae) in the rocky shore ecosystem of the St Malo area: Morphology and growth of the sporophyte. Bot. Mar. 1999, 42, 71–82. [Google Scholar] [CrossRef]
- Hedley, J.D.; Mumby, P.J. Biological and Remote Sensing Perspectives of Pigmentation in Coral Reef Organisms. In Advances in Marine Biology; Elsevier: Amsterdam, The Netherlands, 2002; pp. 277–317. [Google Scholar] [CrossRef]
- Kleinig, H. Carotenoids of siphonous green algae: A chemotaxonomical study. J. Phycol. 1969, 5, 281–284. [Google Scholar] [CrossRef]
- Yokohama, Y.; Kageyama, A. A carotenoid characteristic of chlorophycean seaweeds living in deep coastal waters. Bot. Mar. 1977, 20, 433–436. [Google Scholar] [CrossRef]
- Yokohama, Y. Distribution of the green light-absorbing pigments siphonaxanthin and siphonein in marine green alga. Bot. Mar. 1981, 24, 637–640. [Google Scholar]
- Boraso, A.L.; Piriz de Nuñez de la Rosa, M.L. Las especies del género Codium (Chlorophycophyta) en la costa Argentina. Physis 1975, 34, 245–256. [Google Scholar]
- Giovagnetti, V.; Han, G.; Ware, M.A.; Ungerer, P.; Qin, X.; Wang, W.D.; Kuang, T.; Shen, J.R.; Ruban, A.V. A siphonous morphology affects light-harvesting modulation in the intertidal green macroalga Bryopsis corticulans (Ulvophyceae). Planta 2018, 247, 1293–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganzon-Fortes, E.T. Influence of tidal location on morphology, photosynthesis and pigments of the agarophyte, Gelidiella acerosa, from Northern Philippines. Hydrobiologia 1999, 321–328. [Google Scholar] [CrossRef]
- Sfriso, A.A.; Gallo, M.; Baldi, F. Phycoerythrin productivity and diversity from five red macroalgae. J. Appl. Phycol. 2018, 30, 2523–2531. [Google Scholar] [CrossRef]
- Mogstad, A.A.; Johnsen, G. Spectral characteristics of coralline algae: A multi-instrumental approach, with emphasis on underwater hyperspectral imaging. Appl. Opt. 2017, 56, 9957. [Google Scholar] [CrossRef]
- Vásquez-Elizondo, R.M.; Enríquez, S.; Vasquez-Elizondo, R.M.; Enriquez, S.; Vásquez-Elizondo, R.M.; Enríquez, S. Light absorption in coralline algae (Rhodophyta): A morphological and functional approach to understanding species distribution in a coral reef lagoon. Front. Mar. Sci. 2017, 4, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Beach, K.S.; Borgeas, H.B.; Nishimura, N.J.; Smith, C.M. In vivo absorbance spectra and the ecophysiology of reef macroalgae. Coral Reefs 1997, 16, 21–28. [Google Scholar] [CrossRef]
- Kotta, J.; Remm, K.; Vahtmäe, E.; Kutser, T.; Orav-Kotta, H. In-air spectral signatures of the Baltic Sea macrophytes and their statistical separability. J. Appl. Remote Sens. 2014, 8, 083634. [Google Scholar] [CrossRef] [Green Version]
- Vroom, P.S.; Smith, C.M.; Keeley, S.C. Cladistics of the Bryopsidales: A preliminary analysis. J. Phycol. 1998, 34, 351–360. [Google Scholar] [CrossRef]
- Sugawara, T.; Ganesan, P.; Li, Z.; Manabe, Y.; Hirata, T. Siphonaxanthin, a Green Algal Carotenoid, as a Novel Functional Compound. Mar. Drugs 2014, 12, 3660–3668. [Google Scholar] [CrossRef] [Green Version]
- Yokohama, Y. Vertical Distribution and Photosynthetic Pigments of Marine Green Algae. Korean J. Phycol. 1989, 4, 149–163. [Google Scholar]
- Anderson, J.M. Chlorophyll-protein complexes of a Codium species, including a light-harvesting siphonaxanthin-Chlorophylla ab-protein complex, an evolutionary relic of some Chlorophyta. Biochim. Biophys. Acta Bioenerg. 1983, 724, 370–380. [Google Scholar] [CrossRef]
- Yokohama, Y. A Xanthophyll Characteristic of Deep-Water Green Algae Lacking Siphonaxanthin. Bot. Mar. 1983, 26, 45–48. [Google Scholar] [CrossRef]
- Enríquez, S.; Agustí, S.; Duarte, C.M. Light absorption by marine macrophytes. Oecologia 1994, 98, 121–129. [Google Scholar] [CrossRef]
- Lüning, K.; Dring, M.J. Action spectra and spectral quantum yield of photosynthesis in marine macroalgae with thin and thick thalli. Mar. Biol. 1985, 87, 119–129. [Google Scholar] [CrossRef]
- Barrett, J.; Anderson, J.M. Thylakoid membrane fragments with different chlorophyll A, chlorophyll C and fucoxanthin compositions isolated from the brown seaweed Ecklonia radiata. Plant. Sci. Lett. 1977, 9, 275–283. [Google Scholar] [CrossRef]
- Colombo-Pallotta, M.F.; García-Mendoza, E.; Ladah, L.B. Photosynthetic performance, light absorption, and pigment composition of Macrocystis pyrifera (Laminariales, Phaeophyceae) blades from different depths. J. Phycol. 2006, 42, 1225–1234. [Google Scholar] [CrossRef]
- De Martino, A.; Douady, D.; Quinet-szely, M.; Rousseau, B.; Cre, F.; Apt, K.; Caron, L. The light-harvesting antenna of brown algae Highly homologous proteins encoded by a multigene family. Eur. J. Biochem. 2000, 267, 5540–5549. [Google Scholar] [CrossRef] [PubMed]
- Poza, A.M.; Gauna, M.C.; Escobar, J.F.; Parodi, E.R. Temporal dynamics of algal epiphytes on Leathesia marina and Colpomenia sinuosa macrothalli (Phaeophyceae). Mar. Biol. Res. 2018, 14, 65–75. [Google Scholar] [CrossRef]
- Ramus, J. Seaweed anatomy and photosynthetic performance: The ecological significance of light guides, heterogeneous absorption and multiple scatter. J. Phycol. 1978, 14, 352–362. [Google Scholar] [CrossRef]
- Markager, S. Light Absorption and Quantum Yield for Growth in Five Species of Marine Macroalga. J. Phycol. 1993, 29, 54–63. [Google Scholar] [CrossRef]
- Uhl, F.; Oppelt, N.; Bartsch, I. Spectral mixture of intertidal marine macroalgae around the island of Helgoland (Germany, North Sea). Aquat. Bot. 2013, 111, 112–124. [Google Scholar] [CrossRef]
- Hoang, T.C.; Leary, M.J.O.; Fotedar, R.K. Remote-Sensed Mapping of Sargassum spp. Distribution around Rottnest Island, Western Australia, Using High-Spatial Resolution WorldView-2 Satellite Data. J. Coast. Res. 2015, 32, 1310. [Google Scholar] [CrossRef]
- Kutser, T.; Vahtmäe, E.; Martin, G. Assessing suitability of multispectral satellites for mapping benthic macroalgal cover in turbid coastal waters by means of model simulations. Estuar. Coast. Shelf Sci. 2006, 67, 521–529. [Google Scholar] [CrossRef]
- Hochberg, E. Capabilities of remote sensors to classify coral, algae, and sand as pure and mixed spectra. Remote Sens. Environ. 2003, 85, 174–189. [Google Scholar] [CrossRef]
- Casal, G.; Sánchez-Carnero, N.; Sánchez-Rodríguez, E.; Freire, J. Remote sensing with SPOT-4 for mapping kelp forests in turbid waters on the south European Atlantic shelf. Estuar. Coast. Shelf Sci. 2011, 91, 371–378. [Google Scholar] [CrossRef]
- Nijland, W.; Reshitnyk, L.; Rubidge, E. Satellite remote sensing of canopy-forming kelp on a complex coastline: A novel procedure using the Landsat image archive. Remote Sens. Environ. 2019, 220, 41–50. [Google Scholar] [CrossRef]
- Anderson, R.J.; Rand, A.; Rothman, M.D.; Share, A.; Bolton, J.J. Mapping and quantifying the South African kelp resource. Afr. J. Mar. Sci. 2007, 29, 369–378. [Google Scholar] [CrossRef]
- Ruffin, C.; King, R.L. Analysis of hyperspectral data using Savitzky-Golay filtering—theoretical basis (Part 1). Int. Geosci. Remote Sens. Symp. 1999, 2, 756–758. [Google Scholar]
- Aasen, H.; Honkavaara, E.; Lucieer, A.; Zarco-Tejada, P. Quantitative Remote Sensing at Ultra-High Resolution with UAV Spectroscopy: A Review of Sensor Technology, Measurement Procedures, and Data Correction Workflows. Remote Sens. 2018, 10, 1091. [Google Scholar] [CrossRef] [Green Version]
- Kruse, F.A.; Lefkoff, A.B.; Boardman, J.W.; Heidebrecht, K.B.; Shapiro, A.T.; Barloon, P.J.; Goetz, A.F.H. The spectral image processing system (SIPS)—Interactive visualization and analysis of imaging spectrometer data. Remote Sens. Environ. 1993, 44, 145–163. [Google Scholar] [CrossRef]
- Jiménez, M.; Díaz-Delgado, R. Towards a Standard Plant Species Spectral Library Protocol for Vegetation Mapping: A Case Study in the Shrubland of Doñana National Park. ISPRS Int. J. Geo-Inf. 2015, 4, 2472–2495. [Google Scholar] [CrossRef] [Green Version]
- Medina Machín, A.; Marcello, J.; Hernández-Cordero, A.I.; Martín Abasolo, J.; Eugenio, F. Vegetation species mapping in a coastal-dune ecosystem using high resolution satellite imagery. GIScience Remote Sens. 2019, 56, 210–232. [Google Scholar] [CrossRef]
- Knipling, E. Physical and physiological basis for the reflectance of visible and near-infrared radiation from vegetation. Remote Sens. Environ. USDA 1970, 1, 155–159. [Google Scholar] [CrossRef]
- Hochberg, E.J.; Atkinson, M.J. Spectral discrimination of coral reef benthic communities. Coral Reefs 2000, 19, 164–171. [Google Scholar] [CrossRef]
- Rico, A.; Lanas, P.; Lopez-Gappa, J. Colonization potential of the genus Ulva (Chlorophyta, Ulvales) in Comodoro Rivadavia Harbor (Chubut, Argentina). Ciencias Mar. 2005, 31, 719–735. [Google Scholar] [CrossRef] [Green Version]
- Casas, G.; Scrosati, R.; Luz Piriz, M. The invasive kelp Undaria pinnatifida (Phaeophyceae, Laminariales) reduces native seaweed diversity in Nuevo Gulf (Patagonia, Argentina). Biol. Invasions 2004, 6, 411–416. [Google Scholar] [CrossRef]
- Dellatorre, F.G.; Avaro, M.G.; Commendatore, M.G.; Arce, L.; Díaz de Vivar, M.E. The macroalgal ensemble of Golfo Nuevo (Patagonia, Argentina) as a potential source of valuable fatty acids for nutritional and nutraceutical purposes. Algal Res. 2020, 45, 101726. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Date | Site | TIM | AD | Reference | |
|---|---|---|---|---|---|---|
| PHAEOPHYCEAE (Brown Algae) | Colpomenia sinuosa | 12/09/2018 | PE | 1 | D1 | Tin et al., 2015 |
| Dictyota dichotoma | 18/03, 14/04/2015 | BC-PE | 10 | D1 | Schmitz et al., 2018 | |
| Macrocystis pyrifera (blade) | 18/03/2015 | BC | 12 | D1 | Jensen et al., 1980, Cavanaugh et al., 2010 | |
| Scytosiphon lomentaria | 12/09/2018 | PE | 2 | D1 | ||
| Undaria pinnatifida (blade) | 13/03/2018 | PE | 2 | D1 | ||
| CHLOROPHYTA (Green Algae) | Bryopsis plumosa | 14/04/2015 | PE | 6 | D1 | * B. vestita Tin et al., 2015 * B. corticulans Giovagnetti et al., 2018 |
| Cladophora falklandica | 12, 14/09/2018 | PE-CP | 2 | D2 | * C. glomerata Kutser et al., 2006, Kotta et al., 2014 | |
| Codium decorticatum | 18/03/2015 | BC | 1 | D1 | * C. duthieae Tin et al., 2015 * C. tomentosum Chao Rodríguez et al., 2017 | |
| Codium fragile | 14/04/2015 | PE | 2 | D1 | ||
| Codium vermilara | 14/04/2015 | PE | 7 | D1 | ||
| Ulva sp. (blade-form) | 18/03, 14/04/2015 | BC-PE | 25 | D1 | * U. fasciata Beach et al., 1997 * U. australis Tin et al., 2015 * U. spp. Chao Rodríguez et al., 2017 * U. instestinalis Kotta et al., 2014 | |
| Ulva sp. (tube-form) | 14/04/2015 | PE | 6 | D1 | Kutser et al., 2006, Tin et al., 2015, Chao Rodríguez et al., 2017 | |
| RHODOPHYTA (Red Algae) | Anotrichium furcellatum | 18/03, 14/04/2015 | BC-PE | 10 | D1 | |
| Aphanocladia robusta | 13/09/2018 | PE | 2 | D2 | ||
| Ceramium diaphanum | 13/09/2018 | PE | 2 | D2 | * C. tenuicorne Kotta et al., 2014 C. virgatum as C. rubrum Chao Rodríguez et al., 2017 | |
| Ceramium virgatum | 18/03, 14/04/2015 | BC-PE | 8 | D1 | ||
| Chondria macrocarpa | 18/03/2015 | BC | 7 | D1 | * C. dasyphylla Chao Rodríguez, et al., 2017 | |
| Corallina officinalis | 18/03,14/04/2015 | BC-PE | 11 | D1 | Chao Rodríguez et al., 2017, Mogstad and Johnsen 2017 | |
| Gracilaria gracilis | 14/04/2015 | PE | 4 | D1 | * G.salicornia Beach et al., 1997 | |
| Heterosiphonia merenia | 13, 14/09/2018 | PE-CP | 3 | D2 | ||
| Hymenena laciniata | 13/09/2018 | PE | 5 | D1 | ||
| Lomentaria clavellosa | 14/04/2015 | PE | 12 | D1 | ||
| Myriogramme livida | 14/09/2018 | CP | 1 | D2 | ||
| Neosiphonia harveyi | 13/09/2018 | PE | 1 | D2 | ||
| Phycodris quercifolia | 14/09/2018 | CP | 2 | D2 | ||
| Polysiphonia brodiei | 14/09/2018 | CP | 2 | D2 | * P. fucoides Kotta et al., 2014 | |
| Polysiphonia morowii | 12,14/09/2018 | CP | 3 | D2 | ||
| Pyropia columbina | 18/03/2015 | BC | 18 | D1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olmedo-Masat, O.M.; Raffo, M.P.; Rodríguez-Pérez, D.; Arijón, M.; Sánchez-Carnero, N. How Far Can We Classify Macroalgae Remotely? An Example Using a New Spectral Library of Species from the South West Atlantic (Argentine Patagonia). Remote Sens. 2020, 12, 3870. https://doi.org/10.3390/rs12233870
Olmedo-Masat OM, Raffo MP, Rodríguez-Pérez D, Arijón M, Sánchez-Carnero N. How Far Can We Classify Macroalgae Remotely? An Example Using a New Spectral Library of Species from the South West Atlantic (Argentine Patagonia). Remote Sensing. 2020; 12(23):3870. https://doi.org/10.3390/rs12233870
Chicago/Turabian StyleOlmedo-Masat, O. Magalí, M. Paula Raffo, Daniel Rodríguez-Pérez, Marianela Arijón, and Noela Sánchez-Carnero. 2020. "How Far Can We Classify Macroalgae Remotely? An Example Using a New Spectral Library of Species from the South West Atlantic (Argentine Patagonia)" Remote Sensing 12, no. 23: 3870. https://doi.org/10.3390/rs12233870