Bioactive Phytochemicals Isolated from Akebia quinata Enhances Glucose-Stimulated Insulin Secretion by Inducing PDX-1

 , , and
, , and 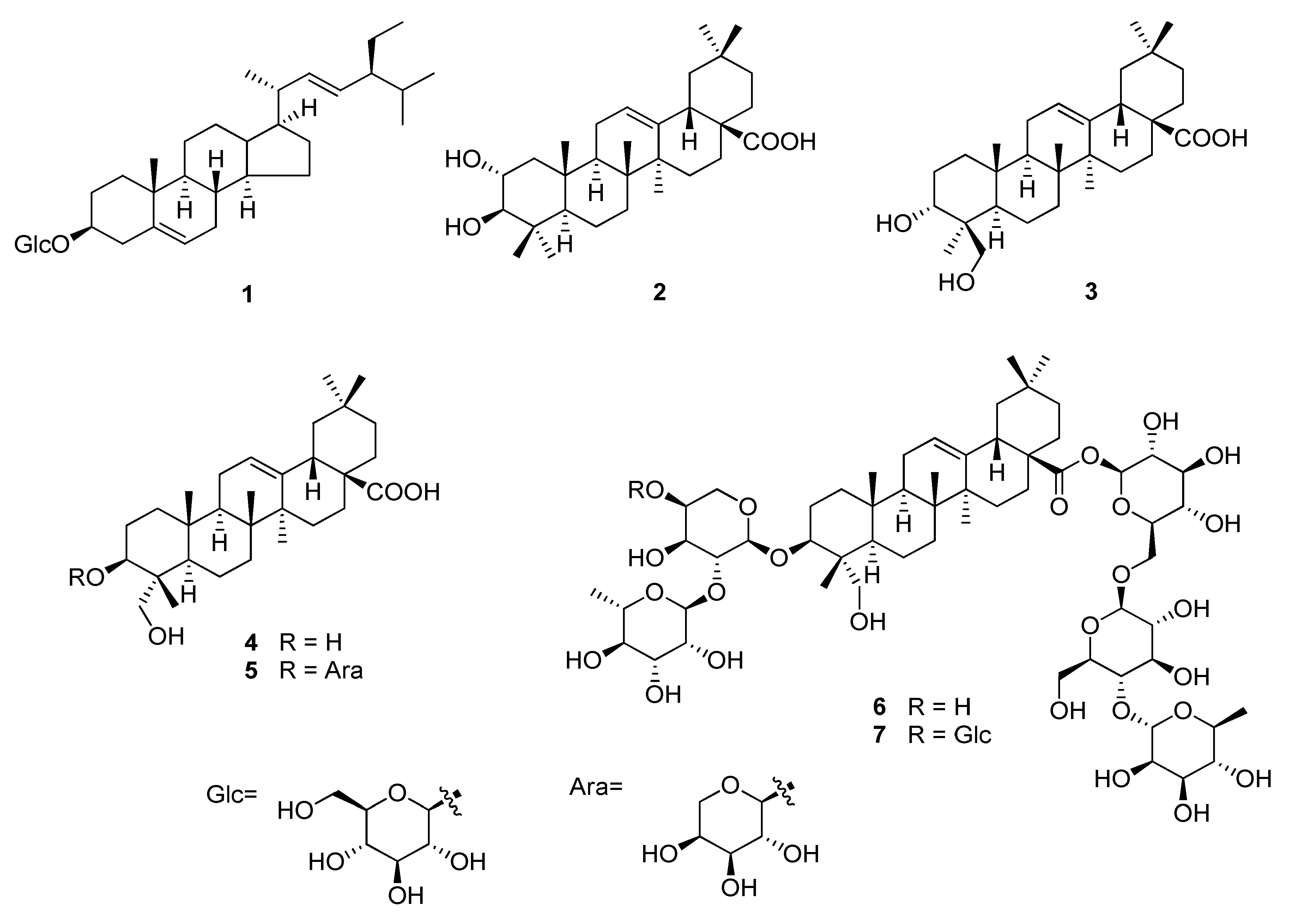{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of Compounds 1–7
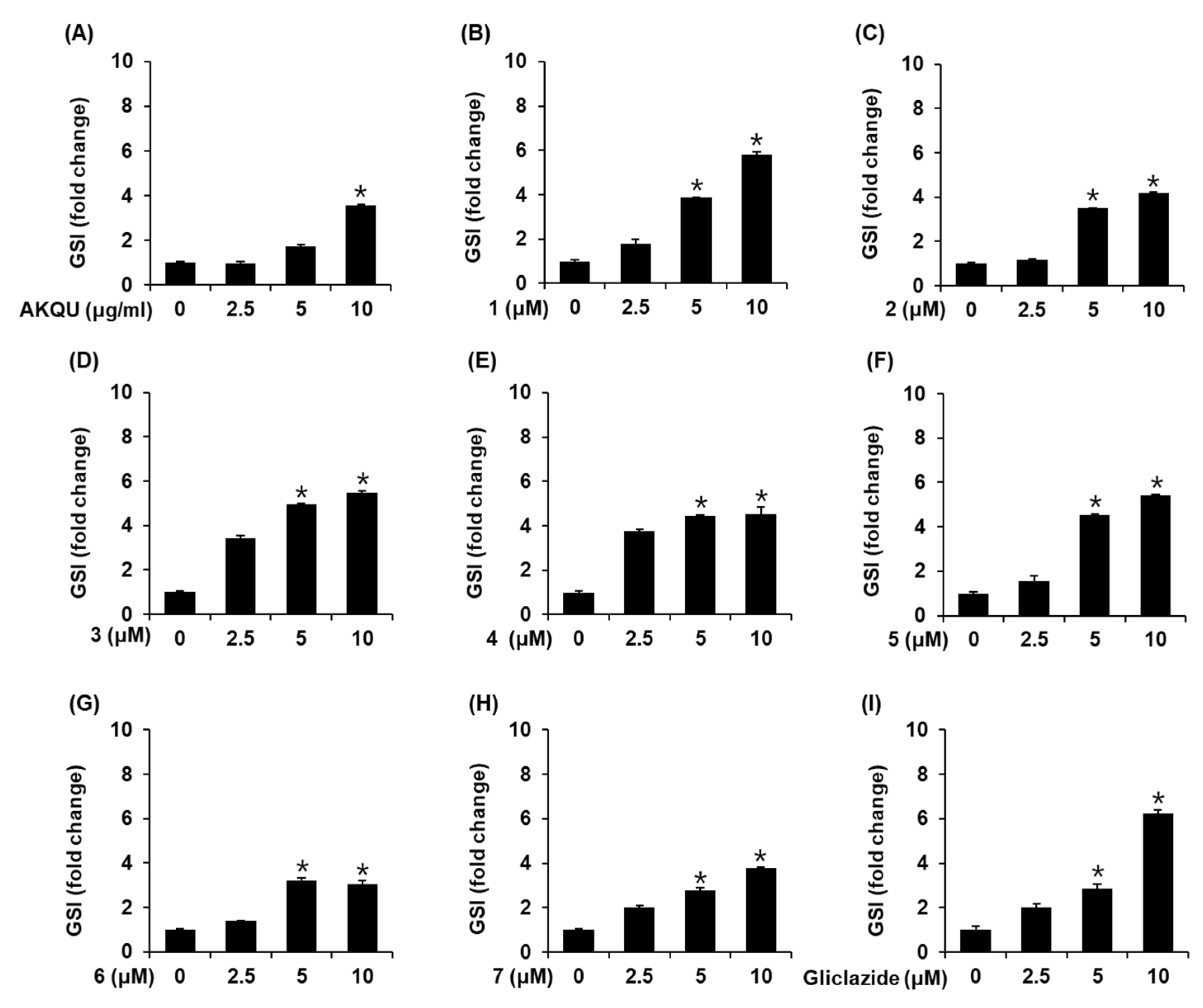
2.2. Effects of AKQU and Compounds 1–7 on GSIS
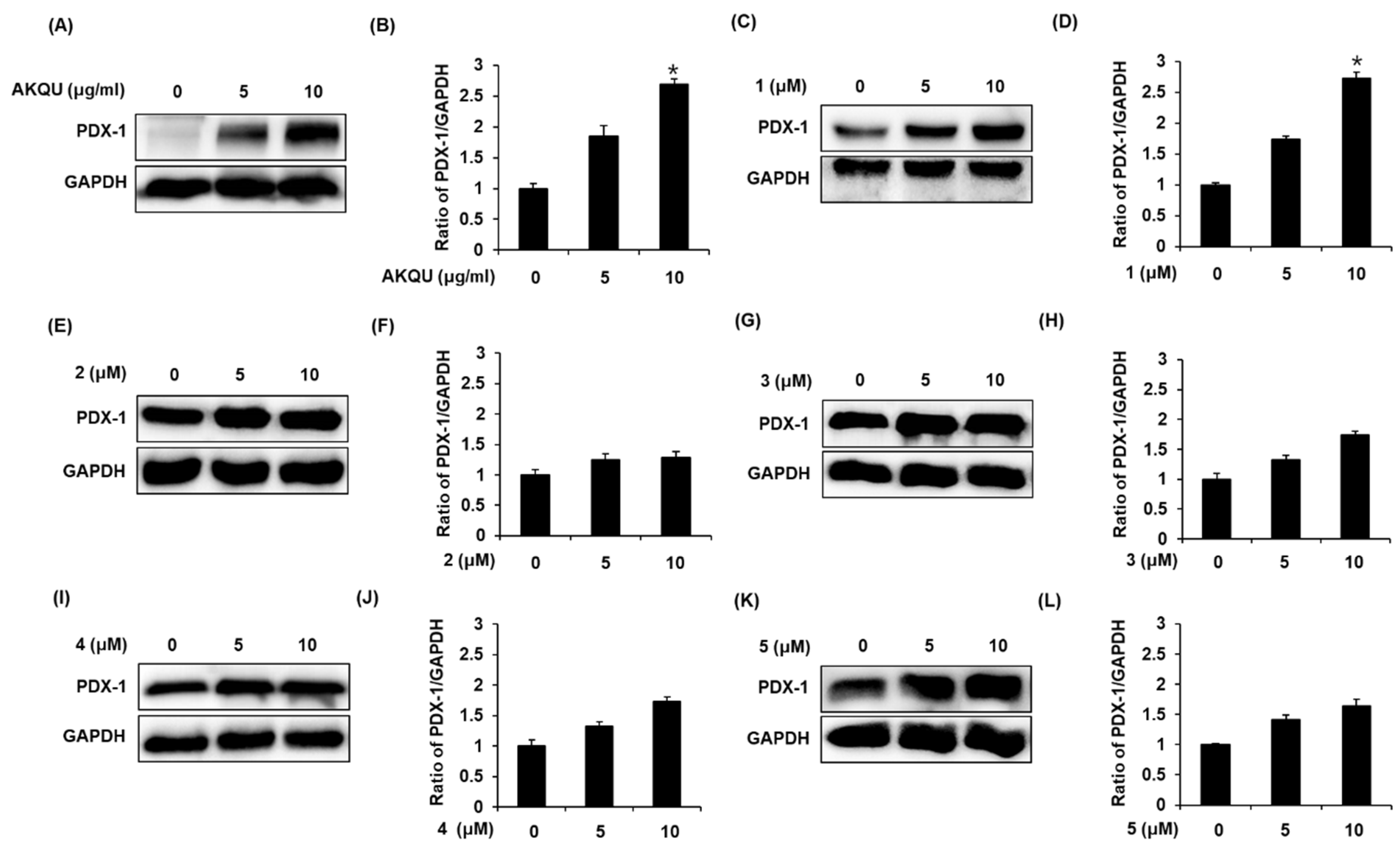
2.3. Effect of AKQU and Compounds 1–5 on the Expression of PDX-1 Protein
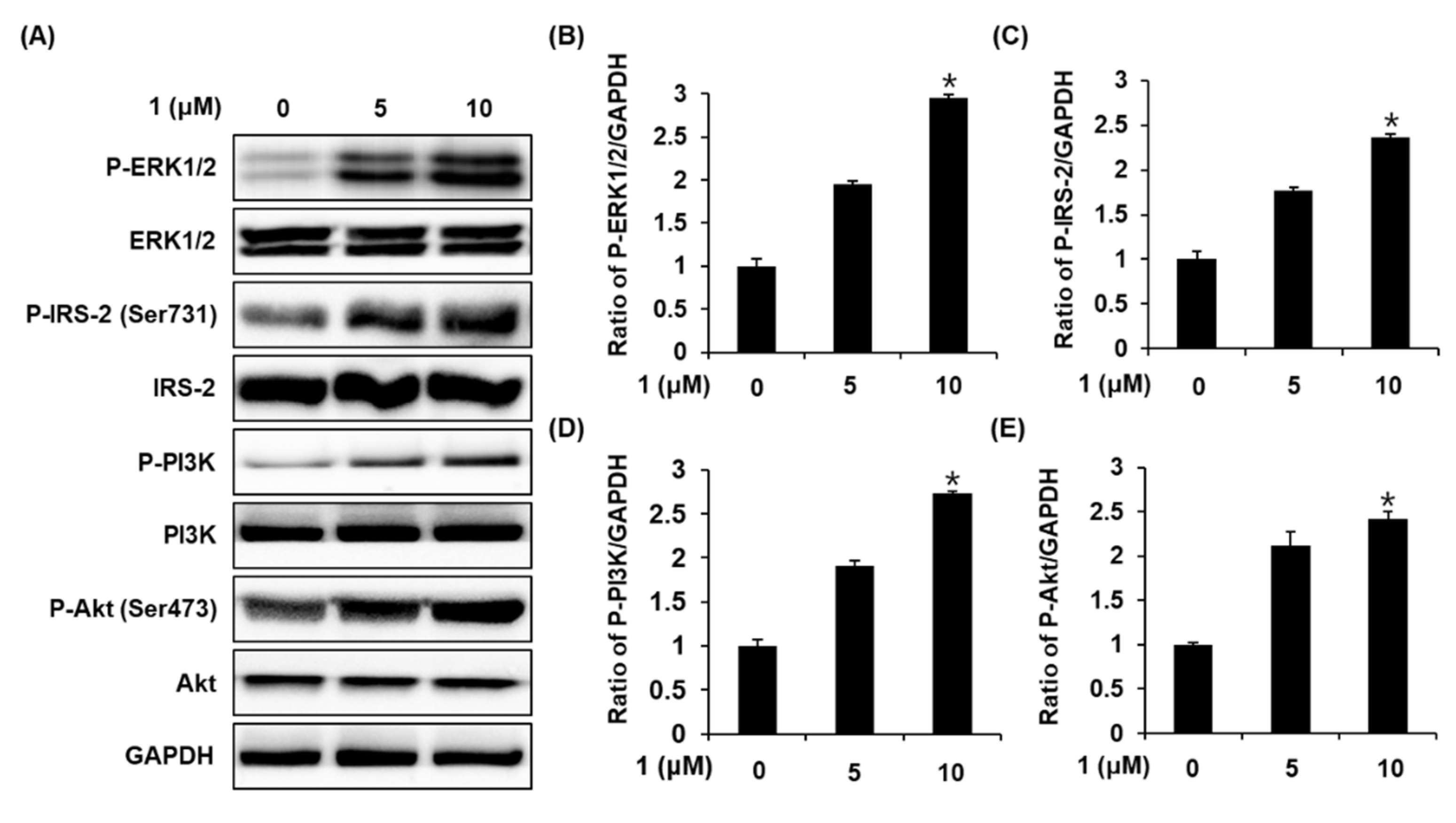
2.4. Effect of Stigmasterol-3-O-β-d-Glucoside (1) on the Protein Expression of P-ERK1/2, ERK1/2, P-IRS-2, IRS-2, P-PI3K, PI3K, P-Akt (Ser473), and Akt
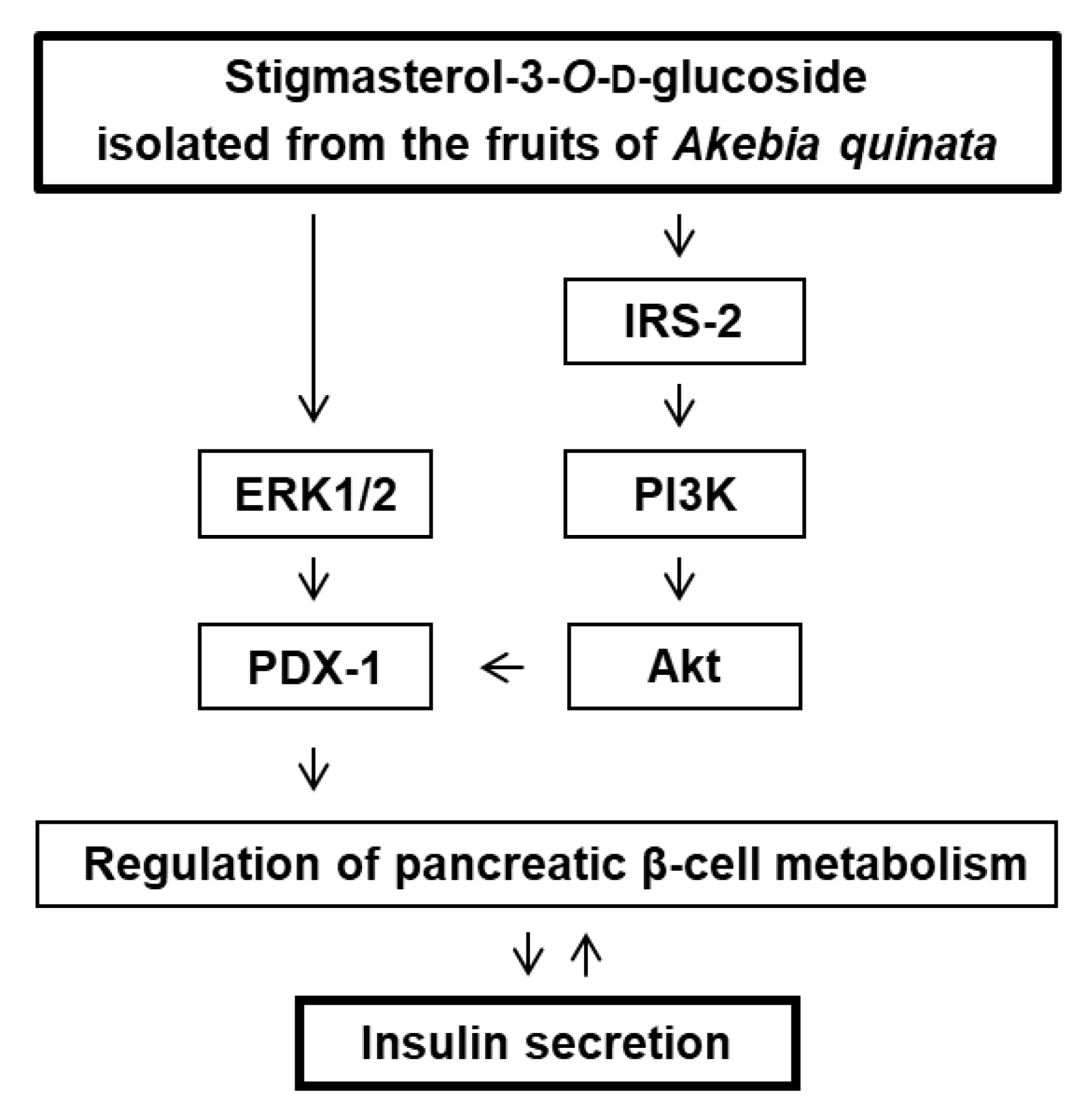
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Extraction and Isolation
4.4. Cell Culture
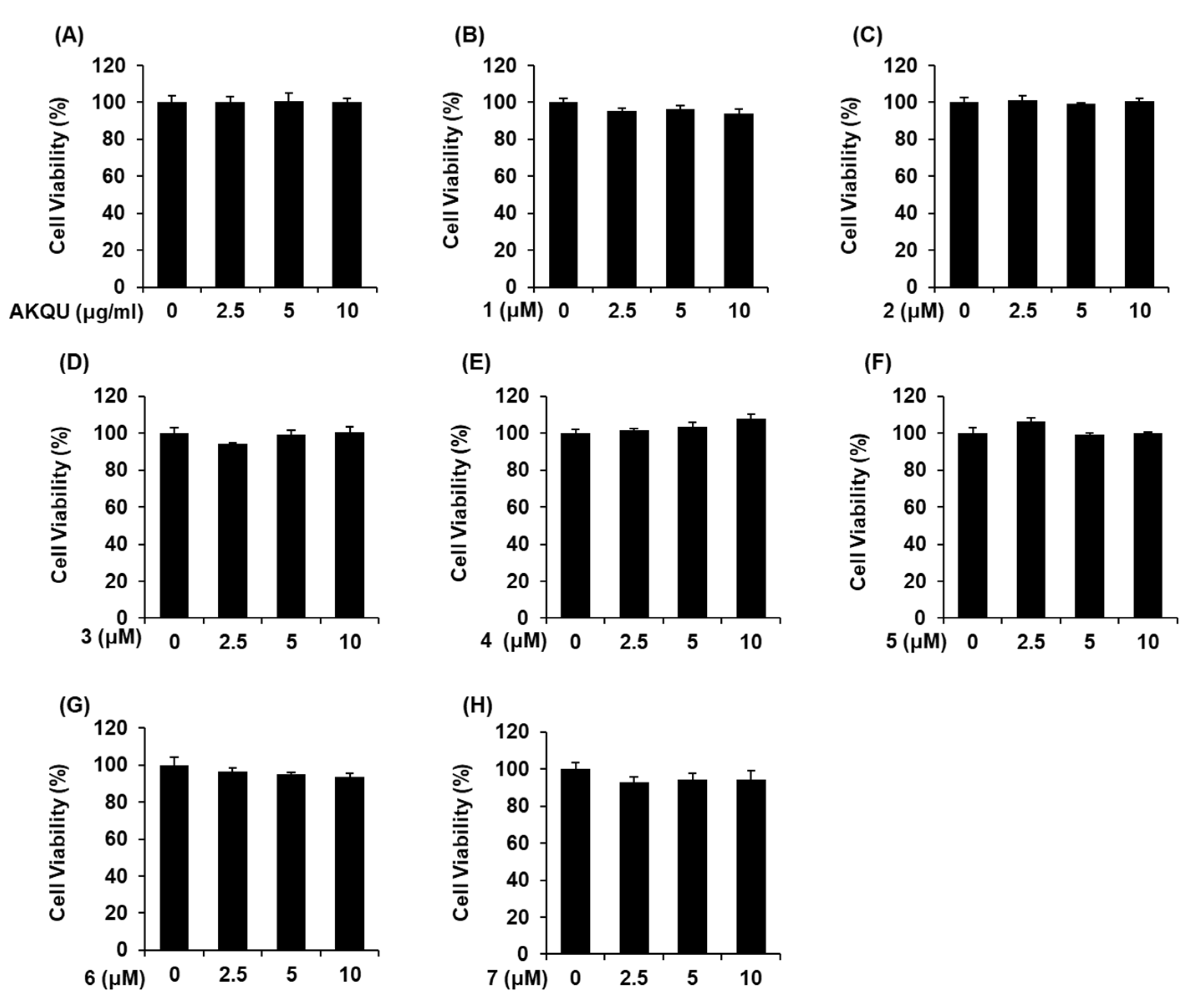
4.5. Ez-Cytox Cell Viability Assay
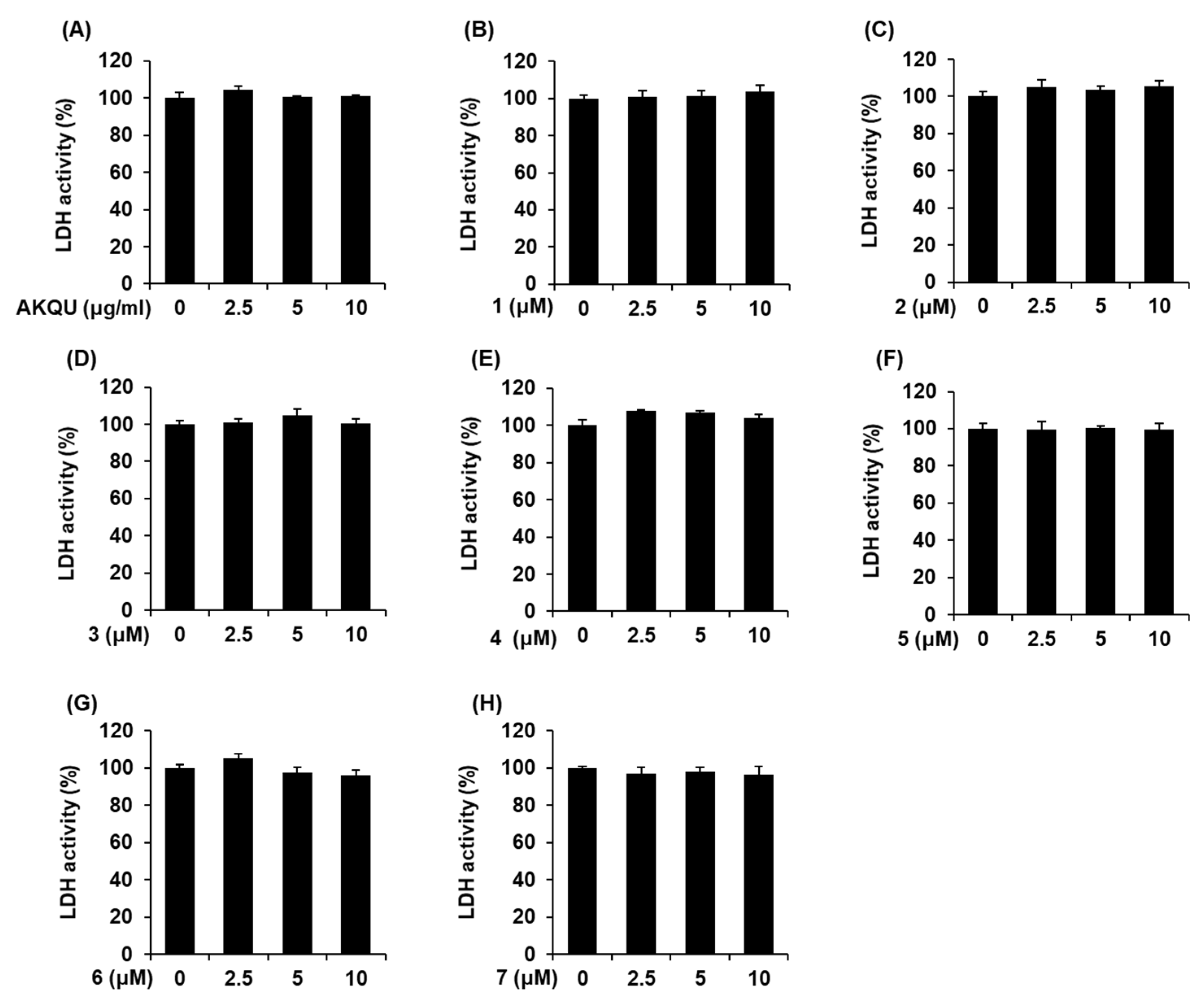
4.6. LDH Release Assay
4.7. GSIS Assay
4.8. Western Blotting
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shaw, J.E.; Sicree, R.A.; Zimmet, P.Z. Global estimates of the prevalence of diabetes for 2010 and 2030. Diabetes Res. Clin. Pract. 2010, 87, 4–14. [Google Scholar] [CrossRef]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [Green Version]
- Goyal, R.; Jialal, I. Diabetes Mellitus Type 2; StatPearls Publishing: St. Petersburg, FL, USA, 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK513253/ (accessed on 3 June 2020).
- Nathan, D.M.; Buse, J.B.; Davidson, M.B.; Ferrannini, E.; Holman, R.R.; Sherwin, R.; Zinman, B. Medical management of hyperglycemia in type 2 diabetes: A consensus algorithm for the initiation and adjustment of therapy: A consensus statement of the American Diabetes Association and the European Association for the Study of Diabetes. Diabetes Care 2009, 32, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.H. Can Diabetes Be Controlled by Lifestyle Activities? Curr. Res. Diabetes Obes. J. 2017, 1, 555568. [Google Scholar]
- Sheehan, M.T. Current therapeutic options in type 2 diabetes mellitus: A practical approach. Clin. Med. Res. 2003, 1, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Sola, D.; Rossi, L.; Schianca, G.P.C.; Maffioli, P.; Bigliocca, M.; Mella, R.; Corlianò, F.; Fra, G.P.; Bartoli, E.; Derosa, G. Sulfonylureas and their use in clinical practice. Arch. Med. Sci. 2015, 11, 840. [Google Scholar] [CrossRef]
- Miller, B.R.; Nguyen, H.; Hu, C.J.-H.; Lin, C.; Nguyen, Q.T. New and emerging drugs and targets for type 2 diabetes: Reviewing the evidence. Am. Health Drug Benefits 2014, 7, 452. [Google Scholar]
- Standl, E.; Erbach, M.; Schnell, O. Dipeptidyl-peptidase-4 inhibitors and heart failure: Class effect, substance-specific effect, or chance effect? Curr. Treat. Options Cardiovasc. Med. 2014, 16, 353. [Google Scholar] [CrossRef]
- Hung, H.-Y.; Qian, K.; Morris-Natschke, S.L.; Hsu, C.-S.; Lee, K.-H. Recent discovery of plant-derived anti-diabetic natural products. Nat. Prod. Rep. 2012, 29, 580–606. [Google Scholar] [CrossRef]
- Kalaitzidis, R.G.; Sarafidis, P.A.; Bakris, G.L. Effects of thiazolidinediones beyond glycaemic control. Curr. Pharm. Des. 2009, 15, 529–536. [Google Scholar] [CrossRef]
- Oh, Y.S. Plant-derived compounds targeting pancreatic beta cells for the treatment of diabetes. Evid. Based Complement. Alternat. Med. 2015. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.-H.; Ngoh, G.-C.; Yusoff, R. A brief review on anti diabetic plants: Global distribution, active ingredients, extraction techniques and acting mechanisms. Pharmacogn. Rev. 2012, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Rauter, A.P.; Martins, A.; Lopes, R.; Ferreira, J.; Serralheiro, L.M.; Araújo, M.-E.; Borges, C.; Justino, J.; Silva, F.V.; Goulart, M. Bioactivity studies and chemical profile of the antidiabetic plant Genista tenera. J. Ethnopharmacol. 2009, 122, 384–393. [Google Scholar] [CrossRef] [Green Version]
- Jesus, A.R.; Dias, C.; Matos, A.M.; de Almeida, R.F.; Viana, A.S.; Marcelo, F.; Ribeiro, R.T.; Macedo, M.P.; Airoldi, C.; Nicotra, F. Exploiting the therapeutic potential of 8-β-d-glucopyranosylgenistein: Synthesis, antidiabetic activity, and molecular interaction with islet amyloid polypeptide and amyloid β-peptide (1–42). J. Med. Chem. 2014, 57, 9463–9472. [Google Scholar] [CrossRef]
- Cernea, S.; Dobreanu, M. Diabetes and beta cell function: From mechanisms to evaluation and clinical implications. Biochem. Med. 2013, 23, 266–280. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, Q.; Zhou, Z.; Ikeda, Y. PDX1, Neurogenin-3, and MAFA: Critical transcription regulators for beta cell development and regeneration. Curr. Stem Cell Res. Ther. 2017, 8, 240. [Google Scholar] [CrossRef] [Green Version]
- Mosley, A.L.; Corbett, J.A.; Özcan, S. Glucose regulation of insulin gene expression requires the recruitment of p300 by the β-cell-specific transcription factor Pdx-1. Mol. Endocrinol. 2004, 18, 2279–2290. [Google Scholar] [CrossRef] [Green Version]
- Gadot, M.; Leibowitz, G.; Shafrir, E.; Cerasi, E.; Gross, D.; Kaiser, N. Hyperproinsulinemia and insulin deficiency in the diabetic Psammomys obesus. Endocrinology 1994, 135, 610–616. [Google Scholar] [CrossRef]
- Tang, W.; Eisenbrand, G. Chinese Drugs of Plant Origin: Chemistry, Pharmacology, and Use in Traditional and Modern Medicine; Springer: Berlin, Germany, 1992. [Google Scholar]
- Jin, H.-G.; Kim, A.R.; Ko, H.J.; Lee, S.K.; Woo, E.-R. Three new lignan glycosides with IL-6 inhibitory activity from Akebia quinata. Chem. Pharm. Bull. 2014, 62, 288–293. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Shi, S.P.; Cao, J.J.; Gao, Q.P.; Tu, P.F. Triterpene saponins from the fruits of Akebia quinata. Biochem. Syst. Ecol. 2008, 2, 138–141. [Google Scholar] [CrossRef]
- Li, L.; Yao, X.; Zhong, C.; Chen, X.; Huang, H. Akebia. A potential new fruit crop in China. HortScience 2010, 45, 4–10. [Google Scholar] [CrossRef]
- Lee, S.H.; Song, Y.S.; Jeong, Y.; Ko, K.S. Antioxidative and anti-inflammatory activities of Akebia quinata extracts in an in vitro model of acute alcohol-induced hepatotoxicity. J. Med. Food. 2017, 20, 912–922. [Google Scholar] [CrossRef] [PubMed]
- Merglen, A.; Theander, S.; Rubi, B.; Chaffard, G.; Wollheim, C.B.; Maechler, P. Glucose sensitivity and metabolism-secretion coupling studied during two-year continuous culture in INS-1E insulinoma cells. Endocrinology 2004, 145, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Sato, N.; Hatano, A.; Ogura, H. Sterol glucosides from Prunella vulgaris. Phytochemistry 1990, 29, 2351–2355. [Google Scholar] [CrossRef]
- Zucaro, Z.; Yasmin, L.; Compagnone, R.S.; Hess, S.C.; Monache, F.D. 6 beta-hydroxymaslinic acid, a triterpene from Vochysia ferruginea. J. Brazil. Chem. Soc. 2000, 11, 241–244. [Google Scholar] [CrossRef]
- Kizu, H.; Tomimori, T. Studies on the constituents of Clematis species. V. On the saponins of the root of Clematis chinensis osbeck. Chem. Pharm. Bull. 1982, 30, 3340–3346. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Van Puyvelde, L.; Maes, L.; Bosselaers, J.; De Kimpe, N. Antitrichomonas in vitro activity of Cussonia holstii Engl. Nat. Prod. Res. 2003, 17, 127–133. [Google Scholar] [CrossRef]
- Shao, C.-J.; Kasai, R.; Xu, J.-D.; Tanaka, O. Saponins from roots of Kalopanax septemlobus (Thunb.) Koidz, Ciqiu: Structures of Kalopanax-saponins C, D, E and F. Chem. Pharm. Bull. 1989, 37, 311–314. [Google Scholar] [CrossRef] [Green Version]
- Glebko, L.I.; Krasovskaj, N.P.; Strigina, L.I.; Ulanova, K.P.; Denisenko, V.A.; Dmitrenok, P.S. Triterpene glycosides from Pulsatilla chinensis. Russ. Chem. Bull. 2002, 51, 1945–4950. [Google Scholar] [CrossRef]
- Cantley, J.; Ashcroft, F.M. Q&A: Insulin secretion and type 2 diabetes: Why do β-cells fail? BMC Biol. 2015, 13, 33. [Google Scholar]
- Fukumitsu, S.; Villareal, M.O.; Aida, K.; Hino, A.; Hori, N.; Isoda, H.; Naito, Y. Maslinic acid in olive fruit alleviates mild knee joint pain and improves quality of life by promoting weight loss in the elderly. J. Clin. Biochem. Nutr. 2016, 59, 220–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokhtari, K.; Rufino-Palomares, E.E.; Pérez-Jiménez, A.; Reyes-Zurita, F.J.; Figuera, C.; García-Salguero, L.; Medina, P.P.; Peragón, J.; Lupiáñez, J.A. Maslinic acid, a triterpene from olive, affects the antioxidant and mitochondrial status of B16F10 melanoma cells grown under stressful conditions. Evid. Based. Complement. Alternat. Med. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, N.; Yagyu, S.; Hata, A.; Nirengi, S.; Kotani, K.; Moritani, T.; Sakane, N. Maslinic acid derived from olive fruit in combination with resistance training improves muscle mass and mobility functions in the elderly. J. Clin. Biochem. Nutr. 2019, 64, 224–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, X.; Zhang, P.; Liu, J.; Zhang, L.; Wu, X.; Ni, P.; Sun, H. Pentacyclic triterpenes. Part 2: Synthesis and biological evaluation of maslinic acid derivatives as glycogen phosphorylase inhibitors. Bioorg. Med. Chem. Lett. 2006, 16, 722–726. [Google Scholar] [CrossRef]
- Wen, X.; Sun, H.; Liu, J.; Wu, G.; Zhang, L.; Wu, X.; Ni, P. Pentacyclic triterpenes. Part 1: The first examples of naturally occurring pentacyclic triterpenes as a new class of inhibitors of glycogen phosphorylases. Bioorg. Med. Chem. Lett. 2005, 15, 4944–4948. [Google Scholar] [CrossRef]
- Liu, J.; Sun, H.; Duan, W.; Mu, D.; Zhang, L. Maslinic acid reduces blood glucose in KK-Ay mice. Biol. Pharm. Bull. 2007, 30, 2075–2078. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Huang, T.; Xue, M.; Chen, J.; Feng, L.; Du, R.; Feng, Y. Current knowledge and development of hederagenin as a promising medicinal agent: A comprehensive review. RSC Adv. 2018, 8, 24188–24202. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.G.; Liu, J.; Kong, L.Y. New pentacyclic triterpenes from Gypsophila oldhamiana and their biological evaluation as glycogen phosphorylase inhibitors. Chem. Biodivers. 2008, 5, 751–757. [Google Scholar] [CrossRef]
- Park, H.-J.; Kim, D.-H.; Choi, J.-W.; Park, J.-H.; Han, Y.-N. A potent anti-diabetic agent from Kalopanax pictus. Arch. Pharm. Res. 1998, 21, 24–29. [Google Scholar] [CrossRef]
- Ling, Y.; Zhang, Q.; Zhu, D.D.; Chen, F.; Kong, X.H.; Liao, L. Identification and characterization of the major chemical constituents in fructus Akebiae by high-performance liquid chromatography coupled with electrospray ionization-quadrupole-time-of-flight mass spectrometry. J. Chromatogr. Sci. 2016, 54, 148–157. [Google Scholar] [CrossRef]
- Sarkar, A.; Tiwari, A.; Bhasin, P.S.; Mitra, M. Pharmacological and pharmaceutical profile of gliclazide: A review. J. Appl. Pharm. Sci. 2011, 1, 11–19. [Google Scholar]
- Ju, L.; Wen, X.; Wang, C.; Wei, Y.; Peng, Y.; Ding, Y.; Feng, L.; Shu, L. Salidroside, a natural antioxidant, improves β-cell survival and function via activating AMPK pathway. Front. Pharmacol. 2017, 8, 749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, J.A.; Szabat, M.; Hoesli, C.A.; Gage, B.K.; Yang, Y.H.C.; Williams, D.E.; Riedel, M.J.; Luciani, D.S.; Kalynyak, T.B.; Tsai, K. A multi-parameter, high-content, high-throughput screening platform to identify natural compounds that modulate insulin and Pdx1 expression. PLoS ONE 2010, 5, e12958. [Google Scholar] [CrossRef] [PubMed]
- Hay, C.W.; Docherty, K. Comparative analysis of insulin gene promoters: Implications for diabetes research. Diabetes 2006, 55, 3201–3213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remedi, M.S.; Emfinger, C. Pancreatic β-cell identity in diabetes. Diabetes Obes. Metab. 2016, 18, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Boulton, T.; Cobb, M. Identification of multiple extracellular signal-regulated kinases (ERKs) with antipeptide antibodies. Cell. Regul. 1991, 2, 357–371. [Google Scholar] [CrossRef] [Green Version]
- Milewski, W.; Duguay, S.; Chan, S.; Steiner, D. Conservation of PDX-1 structure, function, and expression in zebrafish. Endocrinology 1998, 139, 1440–1449. [Google Scholar] [CrossRef]
- Lingohr, M.K.; Briaud, I.; Dickson, L.M.; McCuaig, J.F.; Alárcon, C.; Wicksteed, B.L.; Rhodes, C.J. Specific regulation of IRS-2 expression by glucose in rat primary pancreatic islet β-cells. J. Biol. Chem. 2006, 281, 15884–15892. [Google Scholar] [CrossRef] [Green Version]
- Park, S.M.; Hong, S.M.; Sung, S.R.; Lee, J.E.; Kwon, D.Y. Extracts of Rehmanniae radix, Ginseng radix and Scutellariae radix improve glucose-stimulated insulin secretion and β-cell proliferation through IRS2 induction. Genes. Nutr. 2008, 2, 347–351. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, K.; Polonsky, K.S. Pdx1 and other factors that regulate pancreatic β-cell survival. Diabetes Obes. Metab. 2009, 11, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, H.; Saito, H.; Nishimura, H.; Ueda, J.; Evers, B.M. Activation of phosphatidylinositol-3 kinase regulates pancreatic duodenal homeobox-1 in duct cells during pancreatic regeneration. Pancreas 2008, 36, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafiq, I.; da Silva Xavier, G.; Hooper, S.; Rutter, G.A. Glucose-stimulated preproinsulin gene expression and nucleartrans-location of pancreatic duodenum homeobox-1 require activation of phosphatidylinositol 3-kinase but not p38 MAPK/SAPK2. J. Biol. Chem. 2000, 275, 15977–15984. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Lee, J.S.; Sezirahiga, J.; Kwon, H.C.; Jang, D.S.; Kang, K.S. Bioactive Phytochemicals Isolated from Akebia quinata Enhances Glucose-Stimulated Insulin Secretion by Inducing PDX-1. Plants 2020, 9, 1087. https://doi.org/10.3390/plants9091087
Lee D, Lee JS, Sezirahiga J, Kwon HC, Jang DS, Kang KS. Bioactive Phytochemicals Isolated from Akebia quinata Enhances Glucose-Stimulated Insulin Secretion by Inducing PDX-1. Plants. 2020; 9(9):1087. https://doi.org/10.3390/plants9091087
Chicago/Turabian StyleLee, Dahae, Jin Su Lee, Jurdas Sezirahiga, Hak Cheol Kwon, Dae Sik Jang, and Ki Sung Kang. 2020. "Bioactive Phytochemicals Isolated from Akebia quinata Enhances Glucose-Stimulated Insulin Secretion by Inducing PDX-1" Plants 9, no. 9: 1087. https://doi.org/10.3390/plants9091087