Xyloglucan Is Not Essential for the Formation and Integrity of the Cellulose Network in the Primary Cell Wall Regenerated from Arabidopsis Protoplasts



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
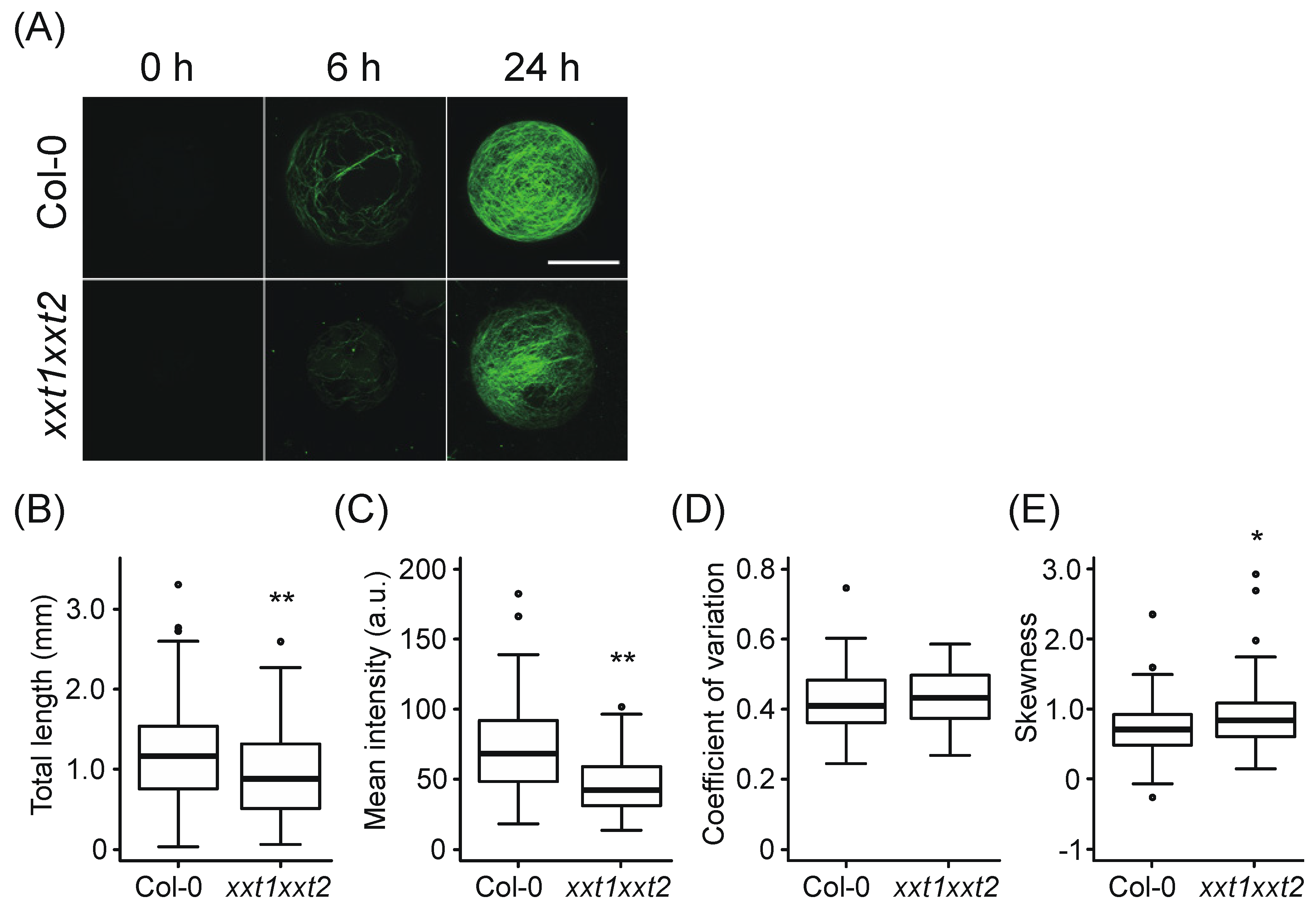
2.1. Comparative Analysis of Network Structure in Cell Walls Regenerated from xxt1 xxt2 and Wild-Type (WT) Protoplasts
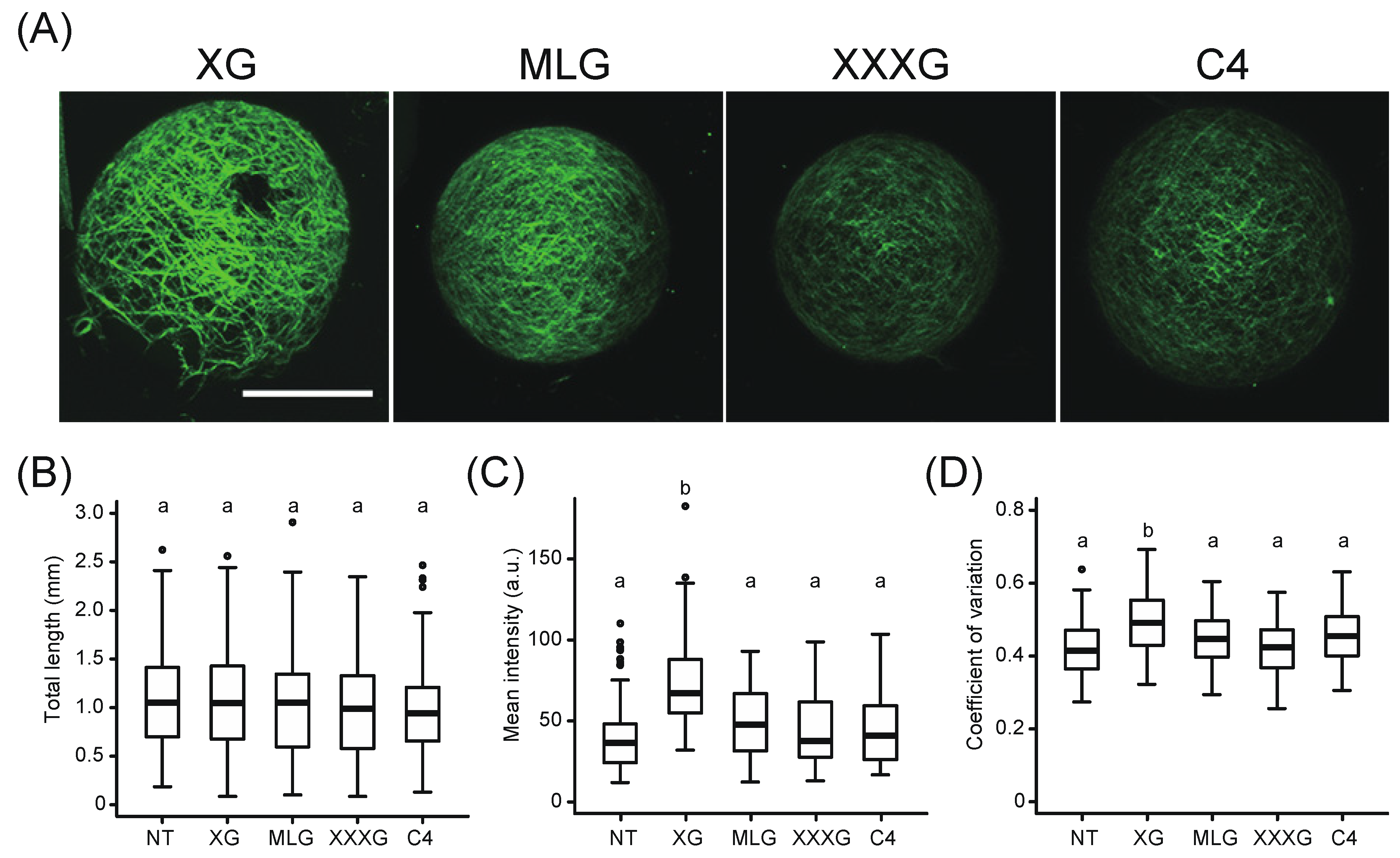
2.2. Evidence of Exogenous Xyloglucan Incorporation into the Cell Wall Network during Regeneration from xxt1 xxt2 Protoplasts
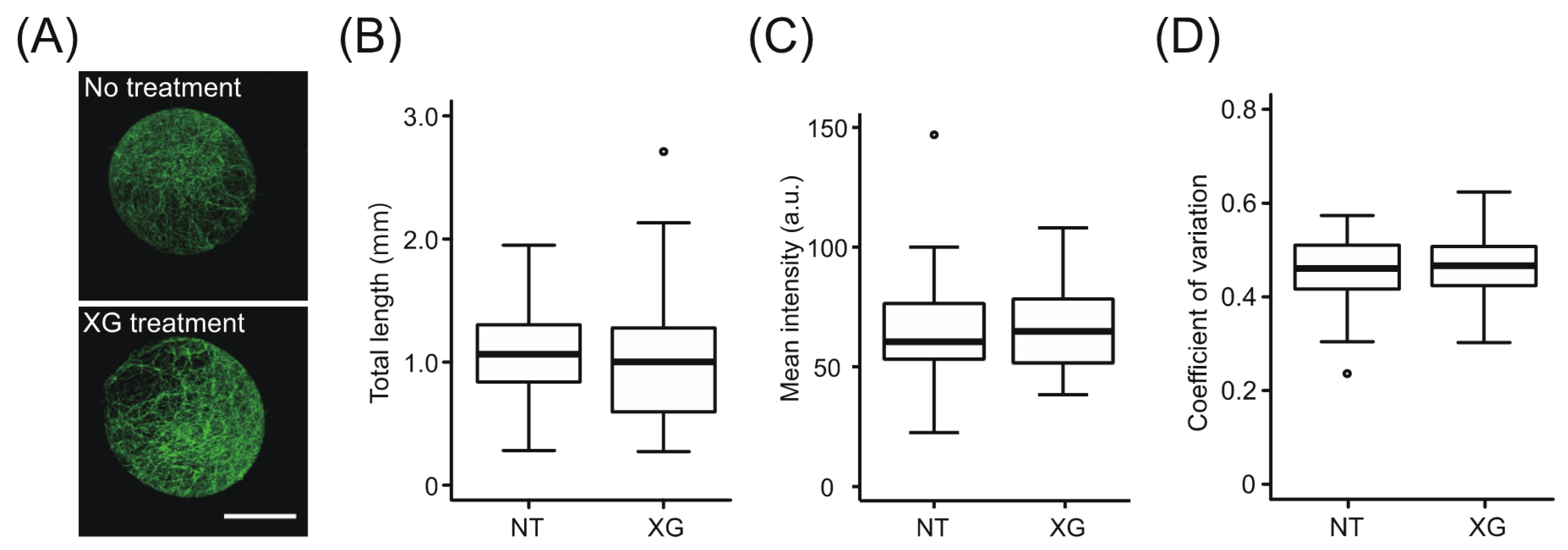
2.3. Effect of Exogenous Xyloglucan on the State of Cellulose Microfibrils during Regeneration of xxt1 xxt2 Protoplasts
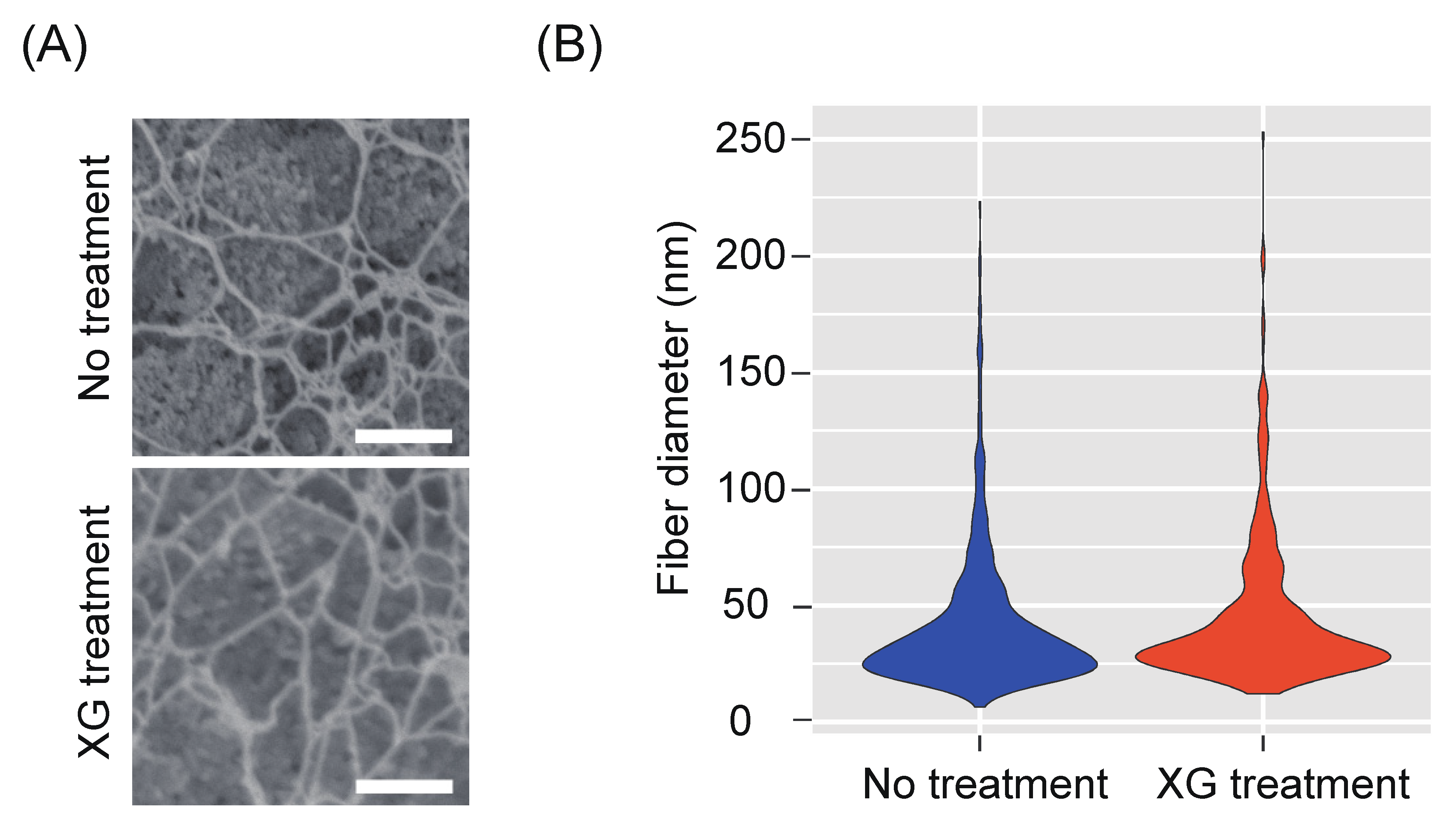
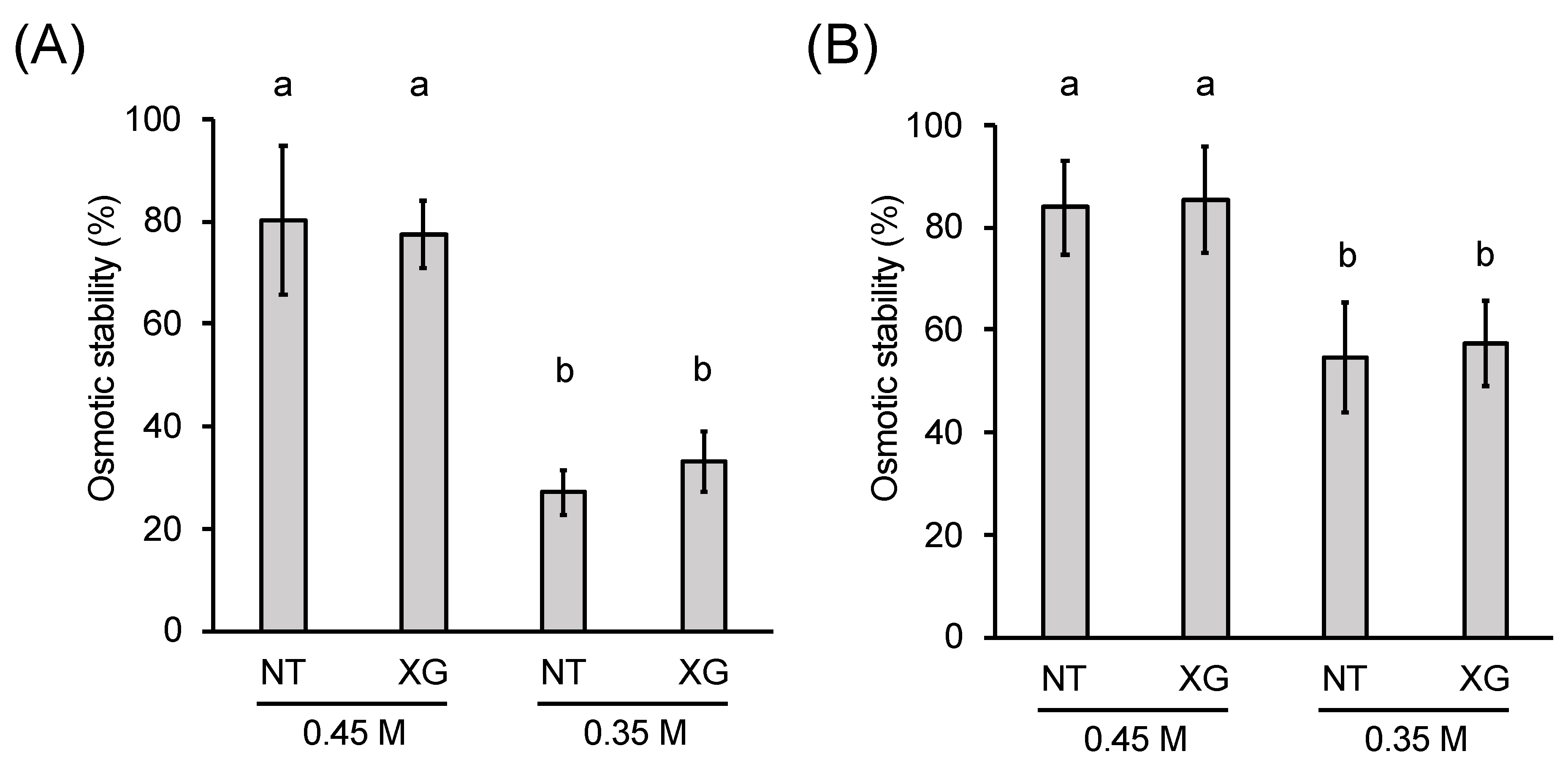
2.4. Role of Xyloglucan (XG) in the Osmotic Stability of Protoplasts
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Condition
4.2. Protoplast Isolation and Incubation
4.3. Calcofluor Staining and Image Acquisition
4.4. Image Analysis
4.5. Immunocytochemistry
4.6. Field-Emission Scanning Electron Microscopy (FE-SEM)
4.7. Osmotic Stability Test
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kimura, S.; Laosinchai, W.; Itoh, T.; Cui, X.; Linder, C.R.; Brown, R.M. Immunogold labeling of rosette terminal cellulose-synthesizing complexes in the vascular plant Vigna angularis. Plant Cell 1999, 11, 2075–2085. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, H.E.; Doring, A.; Persson, S. The cell biology of cellulose synthesis. Annu. Rev. Plant Biol. 2014, 65, 69–94. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Plant cell growth and elongation. eLS 2014. [Google Scholar] [CrossRef]
- Nishitani, K. The role of endoxyloglucan transferase in the organization of plant cell walls. Int. Rev. Cytol. 1997, 173, 157–206. [Google Scholar] [PubMed]
- Keegstra, K.; Talmadge, K.W.; Bauer, W.D.; Albersheim, P. The structure of plant cell walls: III. A model of the walls of suspension-cultured sycamore cells based on the interconnections of the macromolecular components. Plant Physiol. 1973, 51, 188–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Maclachlan, G. Pea xyloglucan and cellulose: I. Macromolecular organization. Plant Physiol. 1984, 75, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Labavitch, J.M.; Ray, P.M. Relationship between promotion of xyloglucan metabolism and induction of elongation by indoleacetic acid. Plant Physiol. 1974, 54, 499–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishitani, K.; Masuda, Y. Auxin-induced changes in the cell wall structure: Changes in the sugar compositions, intrinsic viscosity and molecular weight distributions of matrix polysaccharides of the epicotyl cell wall of Vigna angularis. Physiol. Plant 1981, 52, 482–494. [Google Scholar] [CrossRef]
- Nishitani, K.; Masuda, Y. Auxin-induced changes in cell wall xyloglucans: Effects of auxin on the two different subfractions of xyloglucans in the epicotyl cell wall of Vigna angularis. Plant Cell Physiol. 1983, 24, 345–355. [Google Scholar]
- Carpita, N.C.; Gibeaut, D.M. Structural models of primary cell walls in flowering plants: Consistency of molecular structure with the physical properties of the walls during growth. Plant J. 1993, 3, 1–30. [Google Scholar] [CrossRef]
- Nishitani, K. Construction and Restructuring of the cellulose-xyloglucan framework in the apoplast as mediated by the xyloglucan-related protein family—A hypothetical scheme. J. Plant Res. 1998, 111, 159–166. [Google Scholar] [CrossRef]
- Somerville, C.; Bauer, S.; Brininstool, G.; Facette, M.; Hamann, T.; Milne, J.; Osborne, E.; Paredez, A.; Persson, S.; Raab, T.; et al. Toward a systems approach to understanding plant cell walls. Science 2004, 306, 2206–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fry, S.C.; Smith, R.C.; Renwick, K.F.; Martin, D.J.; Hodge, S.K.; Matthews, K.J. Xyloglucan endotransglycosylase a new wall-loosening enzyme activity from plants. Biochem. J. 1992, 282, 821–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishitani, K.; Tominaga, T. Endo-xyloglucan transferase, a novel class of glycosyltransferase that catalyzes transfer of a segment of xyloglucan molecule to another xyloglucan molecule. J. Biol. Chem. 1992, 267, 21058–21064. [Google Scholar]
- Rose, J.K.; Braam, J.; Fry, S.C.; Nishitani, K. The XTH family of enzymes involved in xyloglucan endotransglucosylation and endohydrolysis: Current perspectives and a new unifying nomencalture. Plant Cell Physiol. 2002, 43, 1421–1435. [Google Scholar] [CrossRef] [Green Version]
- Cavalier, D.M.; Lerouxel, O.; Neumetzler, L.; Yamauchi, K.; Reinecke, A.; Freshour, G.; Zabotina, O.A.; Hahn, M.G.; Burgert, I.; Pauly, M.; et al. Disrupting two Arabidopsis thaliana xylosyltransferase genes results in plants deficient in xyloglucan, a major primary cell wall component. Plant Cell 2008, 20, 1519–1537. [Google Scholar] [CrossRef] [Green Version]
- Dick-Perez, M.; Wang, T.; Salazar, A.; Zabotina, O.A.; Hong, M. Multidimensional solid-state NMR studies of the structure and dynamics of pectic polysaccharides in uniformly 13C-labeled Arabidopsis primary cell walls. Magn. Reson. Chem. 2012, 50, 539–550. [Google Scholar] [CrossRef]
- Dick-Perez, M.; Zhang, Y.; Hayes, J.; Salazar, A.; Zabotina, O.A.; Hong, M. Structure and interactions of plant cell-wall polysaccharides by two- and three-dimensional magic-angle-spinning solid-state NMR. Biochemistry 2011, 50, 989–1000. [Google Scholar] [CrossRef]
- Wang, T.; Park, Y.B.; Cosgrove, D.J.; Hong, M. Cellulose-pectin spatial contacts are inherent to never-dried Arabidopsis primary cell walls: Evidence from solid-state nuclear magnetic resonance. Plant Physiol. 2015, 168, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Zabotina, O.; Hong, M. Pectin-cellulose interactions in the Arabidopsis primary cell wall from two-dimensional magic-angle-spinning solid-state nuclear magnetic resonance. Biochemistry 2012, 51, 9846–9856. [Google Scholar] [CrossRef]
- Bootten, T.J.; Harris, P.J.; Melton, L.D.; Newman, R.H. Solid-state 13C-NMR spectroscopy shows that the xyloglucans in the primary cell walls of mung bean (Vigna radiata L.) occur in different domains: A new model for xyloglucan-cellulose interactions in the cell wall. J. Exp. Bot. 2004, 55, 571–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrove, D.J. Re-constructing our models of cellulose and primary cell wall assembly. Curr. Opin. Plant Biol. 2014, 22, 122–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.B.; Cosgrove, D.J. A revised architecture of primary cell walls based on biomechanical changes induced by substrate-specific endoglucanases. Plant Physiol. 2012, 158, 1933–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.B.; Cosgrove, D.J. Xyloglucan and its interactions with other components of the growing cell wall. Plant Cell Physiol. 2015, 56, 180–194. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.T.; Carroll, A.; Akhmetova, L.; Somerville, C. Real-time imaging of cellulose reorientation during cell wall expansion in Arabidopsis roots. Plant Physiol. 2010, 152, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.W.; Zhang, T.; Zheng, Y.Z.; Cosgrove, D.J.; Anderson, C.J. Xyloglucan deficiency disrupts microtubule stability and cellulose biosynthesis in Arabidopsis, altering cell growth and morphogenesis. Plant Physiol. 2016, 170, 234–249. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.B.; Cosgrove, D.J. Changes in cell wall biomechanical properties in the xyloglucan-deficient xxt1/xxt2 mutant of Arabidopsis. Plant Physiol. 2012, 158, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Fisher, D.D.; Cyr, R.J. Extending the microtubule/microfibril paradigm−Cellulose synthesis is required for normal cortical microtubule alignment in elongating cells. Plant Physiol. 1998, 116, 1043–1051. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Polonenko, D.R.; Camirand, A.; Maclachlan, G. Pea xyloglucan and cellulose: IV. Assembly of beta-glucans by Pea protoplasts. Plant Physiol. 1986, 82, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Nagata, T.; Takebe, I. Cell wall regeneration and cell division in isolated Tobacco mesophyll protoplasts. Planta 1970, 92, 301–308. [Google Scholar] [CrossRef]
- Shea, E.M.; Gibeaut, D.M.; Carpita, N.C. Structural analysis of the cell walls regenerated by carrot protoplasts. Planta 1989, 179, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, R.; Kuki, H.; Kuroha, T.; Nishitani, K. Arabidopsis regenerating protoplast: A powerful model system for combining the proteomics of cell wall proteins and the visualization of cell wall dynamics. Proteomes 2016, 4, 34. [Google Scholar] [CrossRef] [PubMed]
- Kuki, H.; Higaki, T.; Yokoyama, R.; Kuroha, T.; Shinohara, N.; Hasezawa, S.; Nishitani, K. Quantitative confocal imaging method for analyzing cellulose dynamics during cell wall regeneration in Arabidopsis mesophyll protoplasts. Plant Direct 2017. [Google Scholar] [CrossRef]
- Wood, P.J. Specificity in the interaction of direct dyes with polysaccharides. Carbohydr. Res. 1980, 85, 271–287. [Google Scholar] [CrossRef]
- Hernandez-Gomez, M.C.; Rydahl, M.G.; Rogowski, A.; Morland, C.; Cartmell, A.; Crouch, L.; Labourel, A.; Fontes, C.M.G.A.; Willats, W.G.T.; Gilbert, H.J.; et al. Recognition of xyloglucan by the crystalline cellulose-binding stite of a family 3a carbohydrate-binding module. FEBS Lett. 2015, 589, 2297–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.; Coen, E. Interaction between autonomous and microtubule guidance systems controls cellulose synthase trajectories. Curr. Biol. 2020, 30, 941–947. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Bak, G.; Burgin, T.; Barnes, W.J.; Mayes, H.B.; Pena, M.J.; Urbanowicz, B.; Nielsen, E. Biochemical and genetic analysis identify CSLD3 as a beta-1,4-glucan synthase that functions during plant cell wall synthesis. Plant Cell 2020. [Google Scholar] [CrossRef] [Green Version]
- Gu, F.; Bringmann, M.; Combs, J.R.; Yang, J.; Bergmann, D.C.; Nielsen, E. Arabidopsis CSLD5 functions in cell plate formation in a cell cycle-dependent manner. Plant Cell 2016, 28, 1722–1737. [Google Scholar] [CrossRef] [Green Version]
- Bernal, A.J.; Yoo, C.M.; Mutwil, M.; Jensen, J.K.; Hou, G.; Blaukopf, C.; Sorensen, I.; Blancaflor, E.B.; Scheller, H.V.; Willats, W.G.T. Functional analysis of the cellulose synthase-Like genes CSLD1, CSLD2, and CSLD4 in tip-growing Arabidopsis cells. Plant Physiol. 2008, 148, 1238–1253. [Google Scholar] [CrossRef] [Green Version]
- Zablackis, E.; Hugang, J.; Muller, B.; Darvill, A.G.; Albersheim, P. Characterization of the cell-wall polysaccharides of Arabidopsis thaliana leaves. Plant Physiol. 1995, 107, 1129–1138. [Google Scholar] [CrossRef] [Green Version]
- Levy, S.; Maclachlan, G.; Staehelin, L.A. Xyloglucan sidechains modulate binding to cellulose during in vitro binding assays as predicted by conformational dynamics simulations. Plant J. 1997, 11, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Ogawa, K.; Mitsuishi, Y. Characterization of the adsorption of xyloglucan to cellulose. Plant Cell Physiol. 1994, 35, 1199–1205. [Google Scholar] [CrossRef]
- Zhao, F.; Chen, W.; Sechet, J.; Martin, M.; Bovio, S.; Lionnet, C.; Long, Y.; Battu, V.; Mouille, G.; Moneger, F.; et al. Xyloglucans and microtubules synergistically maintain meristem geometry and phyllotaxis. Plant Physiol. 2019, 181, 1191–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armezzani, T.; Abad, U.; Ali, O.; Robin, A.A.; Vachez, L.; Larrieu, A.; Mellerowicz, E.J.; Taconnat, L.; Battu, V.; Stanislas, T.; et al. Transcriptional indution of cell wall remodeling genes is coupled to microtubule-driven growth isotropy at the shoot apex in Arabidopsis. Development 2018. [Google Scholar] [CrossRef] [Green Version]
- Tsukaya, H.; Ohshima, T.; Naito, S.; Chino, M.; Komeda, Y. Sugar-dependent expression of the CHS-A gene for chalcone synthase from petunia in transgenic Arabidopsis. Plant Physiol. 1991, 97, 1414–1421. [Google Scholar] [CrossRef] [Green Version]
- LPIXEL ImageJ Plugins. Available online: https://lpixel.net/en/products/lpixel-imagej-plugins/ (accessed on 14 May 2020).
- Higaki, T.; Kutsuna, N.; Sano, T.; Kondo, N.; Hasezawa, S. Quantification and cluster analysis of actin cytoskeletal structures in plant cells: Role of actin bundling in stomatal movement during diurnal cycles in Arabidopsis guard cells. Plant J. 2010, 61, 156–165. [Google Scholar] [CrossRef]
- PlantProbes. Available online: http://www.plantprobes.net/index.php (accessed on 14 May 2020).
- CarboSource Services. Available online: http://www.carbosource.net (accessed on 14 May 2020).






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuki, H.; Yokoyama, R.; Kuroha, T.; Nishitani, K. Xyloglucan Is Not Essential for the Formation and Integrity of the Cellulose Network in the Primary Cell Wall Regenerated from Arabidopsis Protoplasts. Plants 2020, 9, 629. https://doi.org/10.3390/plants9050629
Kuki H, Yokoyama R, Kuroha T, Nishitani K. Xyloglucan Is Not Essential for the Formation and Integrity of the Cellulose Network in the Primary Cell Wall Regenerated from Arabidopsis Protoplasts. Plants. 2020; 9(5):629. https://doi.org/10.3390/plants9050629
Chicago/Turabian StyleKuki, Hiroaki, Ryusuke Yokoyama, Takeshi Kuroha, and Kazuhiko Nishitani. 2020. "Xyloglucan Is Not Essential for the Formation and Integrity of the Cellulose Network in the Primary Cell Wall Regenerated from Arabidopsis Protoplasts" Plants 9, no. 5: 629. https://doi.org/10.3390/plants9050629