Gibberellins and Heterosis in Crops and Trees: An Integrative Review and Preliminary Study with Brassica

Department of Biological Sciences, University of Lethbridge, Lethbridge, Alberta, T1K 3M4, Canada
*
Author to whom correspondence should be addressed.
Plants 2020, 9(2), 139; https://doi.org/10.3390/plants9020139
Submission received: 10 December 2019
/
Revised: 17 January 2020
/
Accepted: 20 January 2020
/
Published: 22 January 2020
(This article belongs to the Special Issue Plants Heterosis)
Abstract
:Heterosis, or hybrid vigor, has contributed substantially to genetic improvements in crops and trees and its physiological basis involves multiple processes. Four associations with the phytohormone gibberellin (GA) indicate its involvement in the regulation of heterosis for shoot growth in maize, sorghum, wheat, rice, tomato and poplar. (1) Inbreds somewhat resemble GA-deficient dwarfs and are often highly responsive to exogenous GA3. (2) Levels of endogenous GAs, including the bioeffector GA1, its precursors GA19 and GA20, and/or its metabolite GA8, are higher in some fast-growing hybrids than parental genotypes. (3) Oxidative metabolism of applied [3H]GAs is more rapid in vigorous hybrids than inbreds, and (4) heterotic hybrids have displayed increased expression of GA biosynthetic genes including GA 20-oxidase and GA 3-oxidase. We further investigated Brassica rapa, an oilseed rape, by comparing two inbreds (AO533 and AO539) and their F1 hybrid. Seedling emergence was faster in the hybrid and potence ratios indicated dominance for increased leaf number, area and mass, and stem mass. Overdominance (heterosis) was displayed for root mass, leading to slight heterosis for total plant mass. Stem contents of GA19,20,1 were similar across the Brassica genotypes and increased prior to bolting; elongation was correlated with endogenous GA but heterosis for shoot growth was modest. The collective studies support a physiological role for GAs in the regulation of heterosis for shoot growth in crops and trees, and the Brassica study encourages further investigation of heterosis for root growth.
1. Introduction
1.1. Gibberellins and Heterosis in Crops and Trees: Four Lines of Evidence
Heterosis, or hybrid vigor, represents hybrid performance that exceeds that of both parents [1]. Heterosis is common in crop plants and in some trees and has been responsible for a substantial proportion of the genetic improvement through plant breeding over the past century [2,3]. However, while heterosis is widely utilized for agriculture, horticulture and silviculture, and despite a century of investigation, its physiological basis is poorly understood [4].
Studies indicate that there are multiple contributing genetic and physiological factors [5,6,7] and phytohormones should be involved since these provide master regulators of plant form and function [8,9]. Of the phytohormones, there is substantial evidence that the promotive regulator gibberellin (GA) is especially involved in heterosis for shoot growth. There have been preliminary reviews of this association [10,11] and we extend the analyses to incorporate recent findings, expand the range of study approaches and increase the number of plant species.
1.2. Inbreds are Highly Responsive to Exogenous GA3
With plant breeding and genetics programs, several single gene dwarf mutants were identified in maize (or corn, Zea mays L.), peas, Arabidopsis, Brassica and other plants [9]. Some dwarfs have mutations in GA biosynthetic genes and can be restored through the exogenous application of gibberellic acid (GA3), indicating that their small size is due to a GA deficiency. With inbreeding, shoot growth is often depressed and with short stature, inbreds somewhat resemble dwarf mutants. A half century ago, Nickerson [12] recognized this similarity and applied GA3 to different maize genotypes. He found that maize inbreds were highly responsive to exogenous GA3 suggesting that, like the dwarf mutants, the inbreds could be limited by low levels of endogenous GAs. At that same time, Chawdhry [13] found variation in responsivity to GA3 across tomato genotypes and proposed a link between GAs and heterosis. The association was emphasized by Sarkissian et al. [14] and supported in an early review by Paleg [8]. Further studies with maize, the model plant for heterosis (Table 1), and other plants (Table 2) have confirmed the common correspondence, with smaller genotypes including inbreds often being highly responsive to exogenous GA3.
1.3. Hybrids Have Higher Levels of Endogenous GAs
The hypothesis that inbreds are GA deficient was subsequently tested with advancing techniques including selective GA bioassays, GA immunoassays, and definitive physicochemical analyses with combined gas chromatography-mass spectrometry (GC-MS). For GC-MS analyses, internal standard [2H]GAs are added to account for variable losses during the extensive purification [15,16]. Studies with different species and in different research labs supported the hypothesis, as heterotic hybrids contain higher levels of endogenous GAs (Table 1 and Table 2). This provides the second line of evidence supporting a regulatory role of GAs in heterosis for shoot growth.
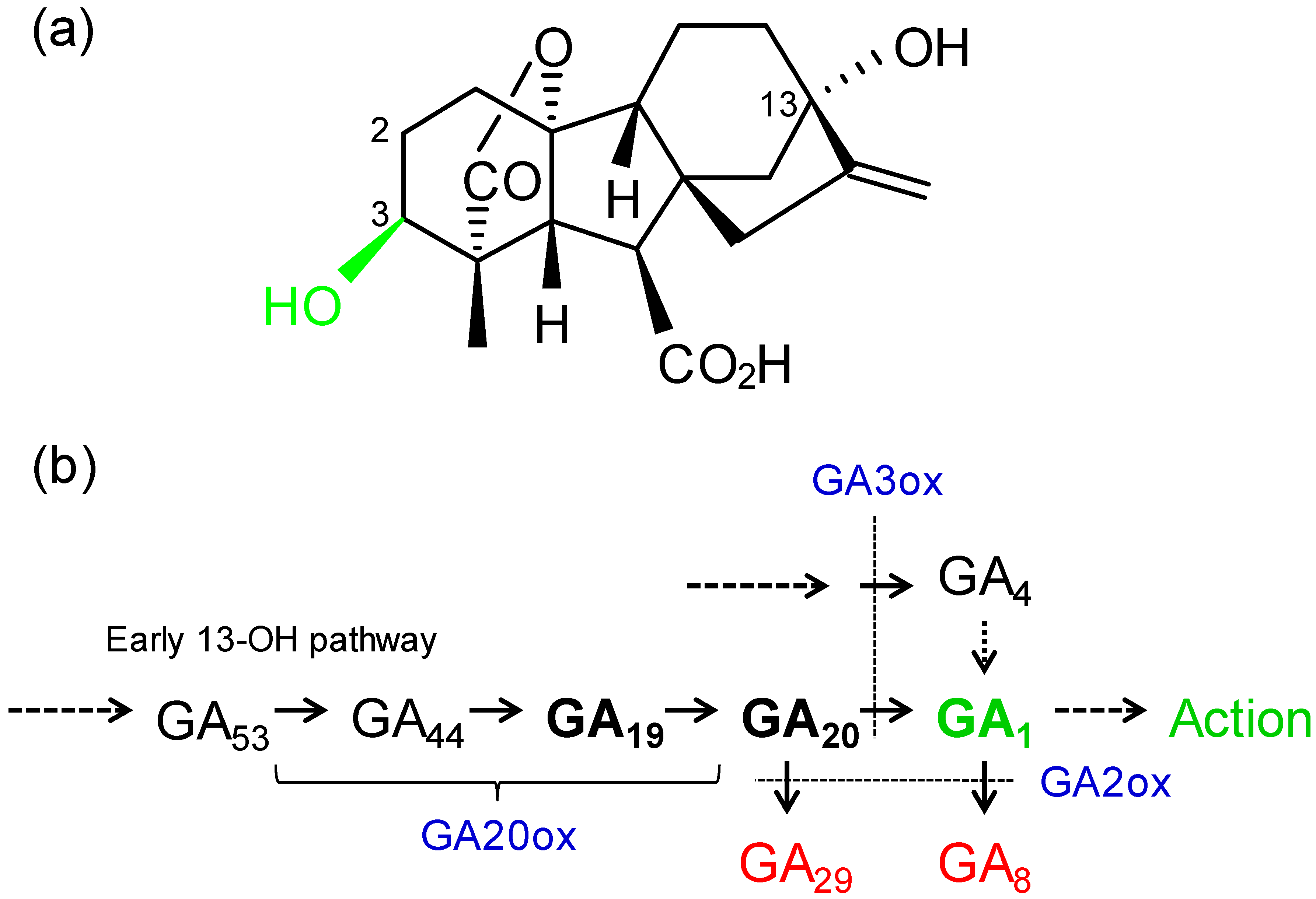
Multiple GAs are native in plants and GA1 is probably the bioeffector GA in most angiosperms for shoot elongation and some other processes (Figure 1a). The biosynthetic pathways have been determined and the “early 13-hydroxylation pathway” is probably the primary pathway in most crops and broad-leaved trees (Figure 1b). Another biosynthetic pathway leads to GA4, a bioactive GA [42] or an alternate GA1 precursor (Figure 1b) [43]. With 2-hydroxylation, GA1 is inactivated to form GA8 and different GAs are often analyzed to provide a record of present (GA1), past (GA8) and future (GA19, GA20, GA4) GA biosynthesis (Figure 1b). Researchers have primarily analyzed these endogenous GAs which may display different patterns across the genotypes (Table 1 and Table 2).
1.4. GA Metabolism is Rapid in Heterotic Hybrids
Analyses of endogenous GAs provide instantaneous measurements, while studies of GA metabolism capture physiological effects over hours or days. [3H]GA20 has been applied to parents and hybrids and the [3H] products analyzed. [2H]GA20 was also applied, permitting GC-MS confirmation of the 3-hydroxylation to [2H]GA1 (Table 1 and Table 2; Figure 1b). In maize and sorghum, metabolism of [3H]GA20 was faster in heterotic hybrids than parental genotypes, as was the 2-hydroxylation of [3H]GA1 to [2H]GA8 or its glucosyl conjugates. These findings indicate faster oxidative metabolism in fast-growing hybrids and provide the third line of evidence for a role of GAs in the regulation of heterosis for shoot growth.
1.5. Expression of GA Biosynthetic Genes is Increased in Some Hybrids
After the GA biosynthetic sequence was determined, the responsible genes were identified and cloned [42,44]. The multi-gene GA 20-oxidase family is responsible for sequential conversions from GA53 through GA44 and GA19 to GA20 (Figure 1b). GA19 is often an abundant endogenous GA and its conversion to GA20 is regarded as rate-limiting and hence, regulatory [44]. This gene has been especially studied and its expression has been associated with heterosis in some plants (Table 1 and Table 2). The final conversion from GA20 to GA1 is enabled by GA 3-oxidase, another multi-gene family (formerly 3β-hydroxylase), and the expression of this gene has also been found to be correlated with heterosis (Table 2). This association between the expression of the GA biosynthetic genes and hybrid vigor provides the fourth line of evidence supporting the role of GAs in the regulation of heterosis for shoot growth in crops and trees.
With multiple studies over a half century, by different researchers and with different plants, there has been substantial support for the association between GAs and heterosis for shoot growth. However, there has also been opposing evidence. In some studies, there have been no correspondences but there are confounding influences. A challenge is the tissue sampling since GAs are differentially distributed within plants, commonly with higher concentration in the shoot apices and subtending elongation zones [45]. Below those zones, the concentrations are lower and if whole stems are sampled there could be greater dilution by the larger stems of hybrids, thus obscuring and even inverting the endogenous GA pattern. This challenge can be increased for trees, which are much larger, and consequently bark scrapings are often sampled (Table 2). However, the locations relative to the growth zones remain a complexity and the timing of sampling further challenges these studies [36,45].
Another complexity relates to the type of growth. The association between GA and stem height is well established across plants, and there are also correspondences with other elongation measures such as leaf lengths, but there may be weaker associations with some other growth measures. A study by Auger et al. [22] applied an interesting approach with GA dwarfing genes in maize, and heterosis persisted. Auger et al. [22] considered this as opposing a regulatory role for GAs but the dwarfing genes would lead to GA reduction [46] rather than elimination, which would be lethal. Consequently, without analyses of the GA contents, that study was less complete.
2. Results
2.1. Gibberellins and Heterosis in Brassica
To further explore the growth components that contribute to heterosis, and to investigate their association with endogenous GAs, Brassica was used as another study system. Heterosis is observed in some Brassica hybrids [47,48] and genetic diversity is incorporated with the allotetraploid genotypes of some rapeseed and canola cultivars [49]. Brassica is a close crop relative to Arabidopsis, allowing for the application of knowledge from that primary model plant system, which also displays heterosis [40,50,51]. Consequently, this study was undertaken with a triplet family consisting of two inbreds, AO533 and AO539, and their F1 hybrid, and we expected: (1) heterosis in the hybrid and (2) correspondence between GA content and stem growth within and across the genotypes.
2.2. Growth and Dominance Patterns
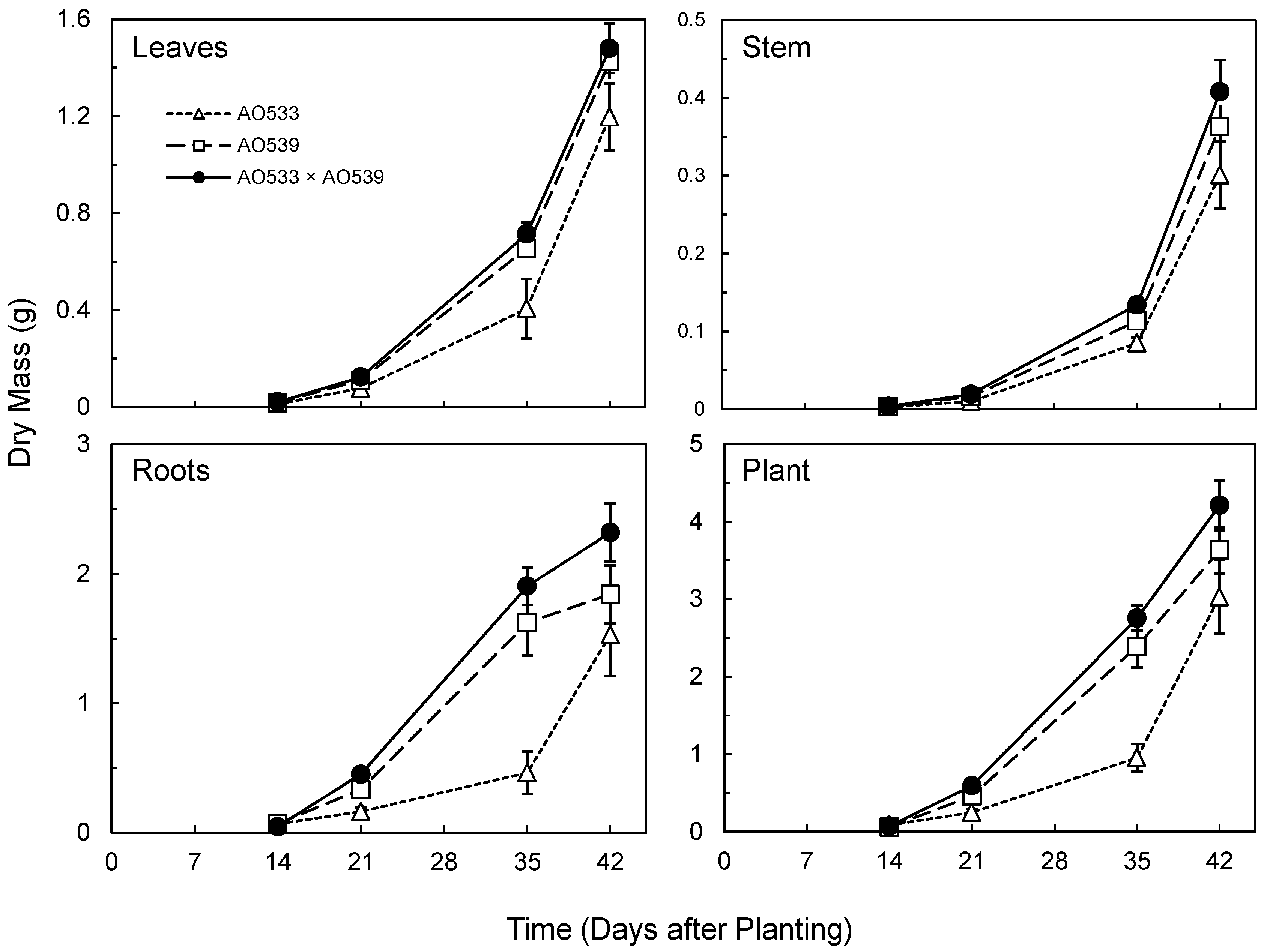
Initial seedling emergence was more rapid in the Brassica hybrid (Table 3). Subsequently, the parent AO539 and the F1 displayed similar seedling emergence percentages, while AO533 displayed slower and less complete emergence. This pattern of dominance or slight overdominance for the increased trait was also observed for most of the subsequent shoot characteristics; the F1 was slightly superior to AO539, while AO533 was consistently less vigorous (Table 3 and Table 4). This was the case for leaf number while leaf sizes were similar. As the product of these two traits, total leaf areas and leaf masses were greater in the F1 and AO539 than AO533 (Table 3 and Table 4, Figure 2).
The sequential harvests displayed the increasing growth of the Brassica plants, with the F1 displaying similar or slightly greater growth than AO539, and both showing greater growth than AO533 (Figure 2). There was apparently slight heterosis for stem mass and the largest difference was for root mass (Figure 2). The F1 root mass was consistently larger and there was a lag in root growth in AO533. This provided the greatest contribution to the total plant mass, which displayed slight heterosis (Figure 2).
The analyses of covariance included the combined results from harvests on days 21, 35 and 42, with day as the covariate. The primary factor of genotype provided significant effects (p < 0.05) for seven of the eight growth measures (Table 4). Subsequent post-hoc pairings with the marginal means confirmed the pattern displayed in the growth plots, with the F1 being similar to or slightly larger than AO539, while AO533 was smaller (Table 4, Figure 2).
Potence ratios (PR) quantified the degree of dominance with values substantially greater than 1 indicating overdominance, or heterosis (Table 4). PR averages from the three harvests are provided (Table 4) and were highest for juvenile height, which extended from the early advantage of the hybrid, and root mass, which contributed to the moderate overdominance for overall plant mass. Root sampling is imprecise due to some adhering substrate particles and the loss of some fine roots. The root rinsing with a water spray appeared complete and we expect similar root recoveries across the harvests and genotypes, providing a valid comparison. The challenge with root sampling should be recognized but the results indicate substantial heterosis for root growth with this triplet of genotypes.
2.3. Endogenous Gibberellins
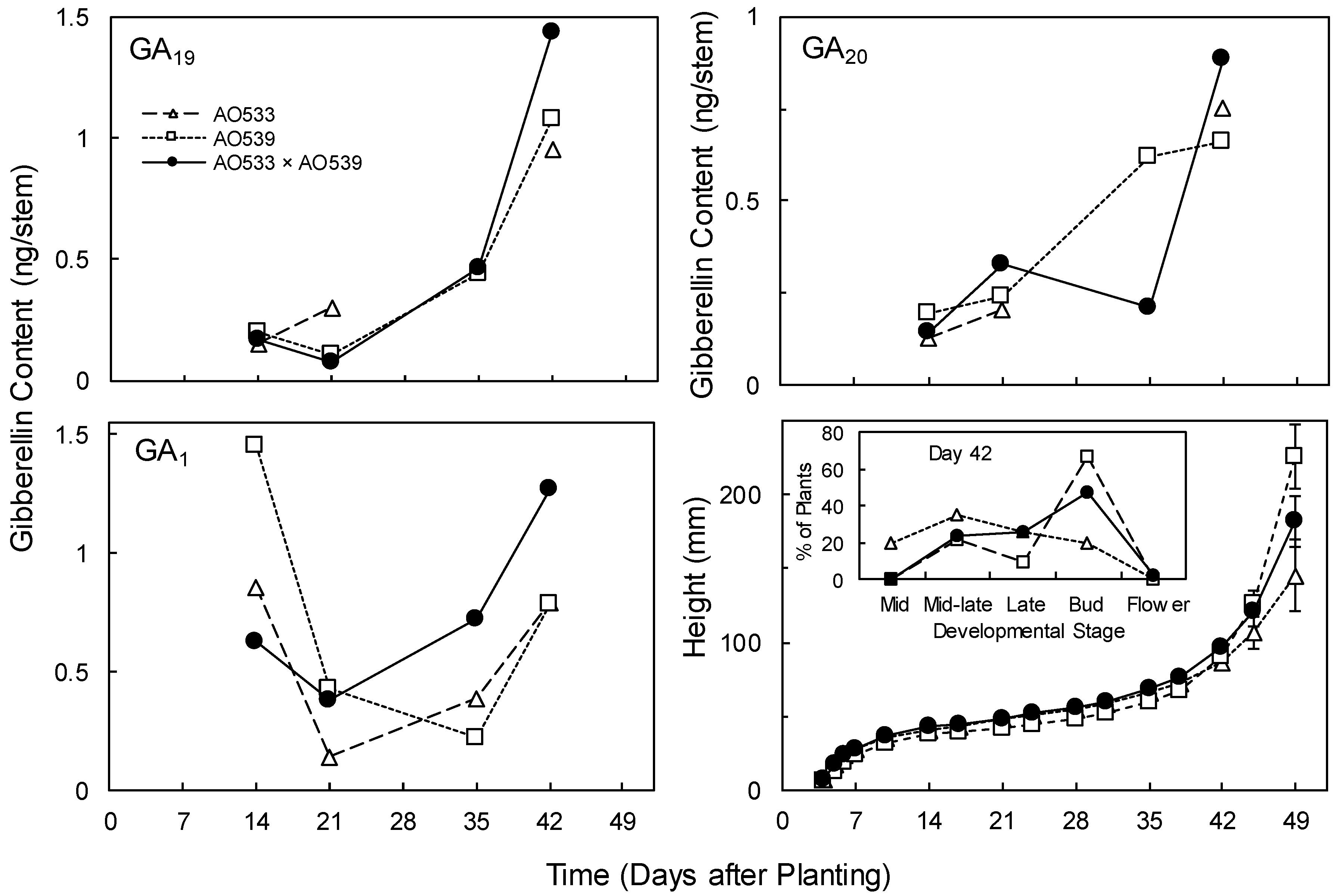
Generally, similar patterns across the genotypes were observed for GA1 and its precursors, GA20 and GA19 (Figure 3). There were often lower GA contents at the early harvests and increases by day 42, when stem bolting was commencing. This was also during the transition to the reproductive phase (Figure 3) and it is likely that endogenous GA would be involved in that developmental process as well as stem elongation. There were no significant differences across the genotypes for GA content or concentration at day 21 or 35, and at day 42 GA contents per stem were significantly higher in the F1 hybrid than in either parent (Figure 3; ANOVA with combined GAs: F(2,4) = 11.3, p = 0.023). Concentrations were higher in the F1 than in AO539 but not significantly different from AO533 (3.16, 2.60 and 3.83 ng/g for GA19; 2.50, 1.59 and 2.36 for GA20; and 2.65, 1.90, and 3.39 for GA1 in AO533, AO539 and the F1, respectively; ANOVA: F(2,4) = 15.49, p = 0.013). These results support the involvement of endogenous GAs in the regulation of stem elongation but are uncertain relative to the relationship with heterosis for shoot growth.
3. Discussion
3.1. Heterosis and Gibberellins in Brassica
In this study, hybrid vigor, superior performance of the hybrid, was slight. As occurs for other plants [14,53,54], seedling emergence was faster in the hybrid. Subsequently, for the different growth traits, the hybrid performance matched or slightly exceeded the superior inbred parent. The expression of heterosis through the multiple traits supports a polygenic foundation and the hybrid would benefit from superior performance for many traits, both independent and coordinated [55]. We expected heterosis for shoot growth, but an unexpected finding was that heterosis was greatest for root growth. There have been fewer studies of heterosis and root growth but early research with tomato hybrids recognized heterosis for root growth [56] and this pattern has been subsequently demonstrated for sorghum and maize [53,57]. At the cellular level, heterosis for gene expression in roots has also been demonstrated, but the function of some genes is less clear [58].
Root mass is a product of root numbers and their sizes, including lengths. The observed overdominance for total root mass might reflect the additive benefit from dominance for higher root numbers and lengths, and it would be useful to resolve the root components and even cell sizes and numbers. If GAs were involved, this might relate to root lengths, since GAs contribute strongly to the regulation of elongation growth and GA manipulation through recombinant GA 2-oxidase resulted in denser but shorter root systems in hybrid poplar [59]. Heterosis for root growth might contribute to yield stability and could influence adaptation to drought or salinity [60].
Relative to endogenous GAs, GA1 and GA19 contents per stem were apparently higher in the hybrid on day 42, when the bolting stage commenced, and the hybrid displayed slight heterosis for increased stem mass. However, this Brassica study was preliminary and future research should focus on hybrid versus parent combinations that display greater hybrid vigor in the growth patterns [61,62], as has been useful for maize inbreds versus hybrids [15,17,23,63]. A comparison for Brassica could include family combinations with increased hybrid vigor [64] and it would be useful to analyze GA contents and especially GA1 along with its precursors and metabolite, as well as with the expression of the key GA biosynthesis genes [39,42] (Table 1). A genetic approach could also be undertaken such as investigating the influence on heterosis from dwarfing genes in Brassica that block GA biosynthesis or action [22,46,65]. Due to the genetic, molecular and physiological similarity, directions from studies of heterosis in Arabidopsis should also be productive and these have supported the involvement of phytohormonal regulation [50]. And while GA could provide a focus, there are likely to be coordinated interactions with other phytohormones and especially auxins [58,66], along with other signaling substances such as brassinosteroids [25].
3.2. A Final Consideration for GAs and Heterosis: Cause or Correlate
While this study with Brassica provided provisional outcomes regarding heterosis, it is clear that GAs are involved in the regulation of shoot growth in Brassica, as is observed for the other plants studied: maize, sorghum, wheat, rice and poplar (Table 1 and Table 2). There have been numerous studies that provide direct or indirect support for the role of GAs in the regulation of heterosis for shoot growth, including stem and leaf growth, in crops and trees. However, while the findings indicate that slow-growing inbreds are limited due to relative GA deficiency, or conversely that hybrids display increased GA biosynthesis and contents, the subsequent question arises, “how are the GA biosynthetic genes regulated?” Thus, GAs may serve as an intermediate between heterotic factors and especially genetic heterozygosity and the phenotypic outcome of hybrid vigor. Also, rather than individual genes being central, as is the case for single gene dwarf mutants of maize, Brassica and other plants, a number of heterozygous genes would probably contribute, providing a polygenic foundation. The consequent up-regulation of the GA biosynthesis genes would provide a mechanism to amplify the outcome, since this would promote a range of morphological and physiological traits that contribute to the phenotypic outcome of hybrid vigor.
4. Material and Methods
4.1. Plant Material and Growth Conditions
Seeds of two inbred parental lines of Brassica rapa L. (syn. B. campestris L.), AO533 and AO539, and their F1 hybrid, AO533 × AO539 (from the Agriculture Canada canola breeding program, Saskatoon, Canada), were sown in pots containing Metro-mix (Ajax, ON, Canada), a Sphagnum peat moss and vermiculite-containing potting medium. Plant pots were randomly arranged in the University of Lethbridge greenhouse (49°70′ N, 112°86′ W), with a 16 h photoperiod that was provided by a combination of natural sunlight and light from Na vapour lamps (260 µmol m−2s−1; Reflector PL90M [medium] N400; P.L. Light Systems Canada, Inc., Beamsville, ON, Canada), and with temperatures maintained between 21–24 °C. Pots were watered daily to saturation and fertilized monthly with a water-soluble 20-14-14 (N-P-K) fertilizer with supplemental trace elements (Professional Gardener Co., Ltd., Calgary, AB, Canada).
4.2. Growth Measurements
Seedling emergence was noted and height and developmental stages [52] were recorded biweekly. Ten individual plants of each genotype were harvested at each of five harvest dates, at 14, 21, 35, 42 and 70 days after planting and for each plant, leaf number, total leaf area using a portable LI-COR photometric area meter (Lincoln, NB, USA), and organ dry weights (leaf, stem and root) were measured. Prior to weighing, leaf and root tissues were oven dried, while stem tissues were flash frozen in liquid nitrogen and then freeze dried for dry weight determinations and to permit analysis of endogenous GAs.
4.3. Potence Ratios
Inheritance patterns were quantified by comparing the performance of the hybrids with that of the two parents [17,67]. The additive component of variation (H) was calculated as the absolute value of either parent (P2 represents the superior parent for that trait) minus the mean of the two parents, the midparent value (mp). The dominance component of variation (D) was calculated as the hybrid value minus mp and the potence ratios represent the overall degree of dominance as:
Potence ratio (PR) = D/H = (Hybrid − mp)/(P2 − mp)
Recognizing some quantitative variation, PR values around 0 (from about −0.5 to 0.5) indicate additive inheritance with the hybrids intermediate between the parents. PR values from ~ −1.5 to −0.5 or 0.5 to 1.5 indicate dominance of the smaller or larger characteristic, respectively. PR values exceeding ~ 1.5 indicate overdominance or hybrid vigor, while values below ~ −1.5 indicate hybrid break-down.
4.4. Analyses of Endogenous Gibberellins
Gibberellins (GAs) were quantified in one of the two replicates, with each consisting of 5-6 stems from a genotype at a harvest. Analyses were undertaken as described by Zanewich and Rood [68] and others [16,69]. Briefly, tissue was extracted in aqueous methanol with internal standards of [2H2]-GA1, -GA19 and -GA20 (from L. Mander, Australian National University), filtered, and buffered (pH 9). Following in vacuo reduction, the aqueous phase was partitioned twice with water-saturated ether. The pH was then adjusted to 3 and the sample was further partitioned 3 times with ethyl acetate. Extracts were frozen, filtered to remove ice, and dried in vacuo. Residues were resuspended and transferred to glass microfiber discs and trace quantities of [3H]GA1 and [3H]GA4 were added for tracking. Samples were further purified using silica columns [68] and respective GA-containing fractions were separated using reversed-phase C18 high performance liquid chromatography. Grouped fractions containing the three GAs of interest were methylated and trimethylsilylated, and GAs were detected using gas chromatography-mass spectrometry (GC-MS) with selected ion monitoring. Quantities were calculated based in the paired ion abundances from the internal standard [2H2]GAs and endogenous forms [16,43].
4.5. Statistical Analyses
Analyses of variance (ANOVA) with SPSS v.19 (IBM, Armonk, NY, USA) provided the primary assessment for each growth measure with genotype and harvest day as fixed factors. These analyses used the data collected from harvest days 21, 35 and 42, which consistently displayed strong linear or log-linear patterns. Gibberellin content and concentrations were also analyzed by ANOVA, for all harvest days (three-factor: GA, genotype and day) or individual days (two-factor: GA and genotype).
Author Contributions
Literature review, S.B.R. and K.P.Z.; Brassica study, conceptualization K.P.Z. and S.B.R.; plant cultivation and harvesting, gibberellin sample preparation and quantification, K.P.Z.; data analyses, K.P.Z. and S.B.R.; manuscript writing and editing, S.B.R. and K.P.Z.; funding acquisition, S.B.R. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by funding from Alberta Innovates and the Natural Sciences and Engineering Research Council of Canada.
Acknowledgments
We thank Keith Topinka (University of Alberta) for providing the Brassica genotypes, and David Major (Agriculture and Agri-Foods Canada) and David Pearce (University of Lethbridge) for helpful manuscript recommendations.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Shull, G.H. Beginnings of the heterosis concept. In Heterosis, Gowen, J.W., Ed.; Iowa State College Press: Ames, IA, USA, 1952; pp. 14–48. [Google Scholar]
- Lippman, Z.B.; Zamir, D. Heterosis: Revisiting the magic. Trends Genet. 2007, 23, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Tollenaar, M.; Ahmadzadeh, A.; Lee, E.A. Physiological basis of heterosis for grain yield in maize. Crop Sci. 2004, 44, 2086–2094. [Google Scholar] [CrossRef]
- Schnable, P.S.; Springer, N.M. Progress toward understanding heterosis in crop plants. Annu. Rev. Plant Biol. 2013, 64, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Birchler, J.A.; Auger, D.L.; Riddle, N.C. In search of the molecular basis of heterosis. Plant Cell 2003, 15, 2236–2239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.J. Genomic and epigenetic insights into the molecular bases of heterosis. Nat. Rev. Genet. 2013, 14, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, R.; Uezono, K.; Ishikura, S.; Peacock, W.J.; Dennis, E.S. Recent research on the mechanism of heterosis is important for crop and vegetable breeding systems. Breed. Sci. 2008, 68, 148–158. [Google Scholar] [CrossRef] [Green Version]
- Paleg, L.G. Physiological effect of gibberellins. Annu. Rev. Plant Physiol. 1965, 16, 291–322. [Google Scholar] [CrossRef]
- Reid, J.B. Plant hormone mutants. J. Plant Growth Regul. 1993, 12, 207–226. [Google Scholar] [CrossRef]
- Pharis, R.P.; Zhang, R.; Jiang, I.B.J.; Dancik, B.P.; Yeh, F.C. Differential efficacy of gibberellins in flowering and vegetative shoot growth, including heterosis and inherently rapid growth. In Progress in Plant Growth Regulation, Karssen, C.M., van Loon, L.C., Vreugdenhil, D., Eds.; Springer: Dordrecht, The Netherlands, 1992; pp. 13–27. [Google Scholar] [CrossRef]
- Rood, S.B.; Pharis, R.P. Hormones and heterosis in plants. In Plant Hormones and their Role in Plant Growth and Development, Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 1987; pp. 463–473. [Google Scholar] [CrossRef]
- Nickerson, N.H. Sustained treatment with gibberellic acid of five different kinds of maize. Ann. Mo. Bot. Gard. 1959, 46, 19–37. [Google Scholar] [CrossRef]
- Chawdhry, M.Y. Relation of Gibberellin Treatment to Heterosis and Parthenocarpy in Tomato. Master’s Thesis, University of Arizona, Tuscon, AZ, USA, 1959. [Google Scholar]
- Sarkissian, I.V.; Kessinger, M.A.; Harris, W. Differential rates of development of heterotic and nonheterotic young maize seedlings, I. Correlation of differential morphological development with physiological differences in germinating seeds. Proc. Natl. Acad. Sci. USA 1964, 51, 212. [Google Scholar] [CrossRef] [Green Version]
- Rood, S.B.; Buzzell, R.I.; Mander, L.N.; Pearce, D.; Pharis, R.P. Gibberellins: A phytohormonal basis for heterosis in maize. Science 1988, 241, 1216–1218. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Viswanath, V.; Ma, C.; Etherington, E.; Dharmawardhana, P.; Shevchenko, O.; Strauss, S.H.; Pearce, D.W.; Rood, S.B.; Busov, V. Recombinant DNA modification of gibberellin metabolism alters growth rate and biomass allocation in Populus. Tree Genet. Genomes 2015, 11, 127. [Google Scholar] [CrossRef] [Green Version]
- Rood, S.B.; Pharis, R.P.; Koshioka, M.; Major, D.J. Gibberellins and heterosis in maize I. Endogenous gibberellin-like substances. Plant Physiol. 1983, 71, 639–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rood, S.B.; Blake, T.J.; Pharis, R.P. Gibberellins and heterosis in maize II. Response to gibberellic acid and metabolism of [3H]GA20. Plant Physiol. 1983, 71, 645–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rood, S.B. Heterosis and the metabolism of [3H]gibberellin A1 in maize. Can. J. Bot. 1986, 64, 2160–2164. [Google Scholar] [CrossRef]
- Rood, S.B.; Larsen, K.M. Gibberellins, amylase, and the onset of heterosis in maize seedlings. J. Exp. Bot. 1988, 39, 223–233. [Google Scholar] [CrossRef]
- Rood, S.B.; Buzzell, R.I.; Major, D.J.; Pharis, R.P. Gibberellins and heterosis in maize: Quantitative relationships between GA content, GA3 response, and growth. Crop Sci. 1990, 30, 281–286. [Google Scholar] [CrossRef]
- Auger, D.L.; Peters, E.M.; Birchler, J.A. A genetic test of bioactive gibberellins as regulators of heterosis in maize. J. Hered. 2005, 96, 614–617. [Google Scholar] [CrossRef] [Green Version]
- Użarowska, A.; Keller, B.; Piepho, H.-P.; Schwarz, G.; Ingvardsen, C.; Wenzel, G.; Lübberstedt, T. Comparative expression profiling in meristems of inbred-hybrid triplets of maize based on morphological investigations of heterosis for plant height. Plant Mol. Biol. 2007, 63, 21–34. [Google Scholar] [CrossRef]
- Du, J.; Yao, Y.; Ni, Z.; Sun, Q. Cloning and characterization of an up-regulated GA 20-oxidase gene in hybrid maize. Prog. Nat. Sci. 2009, 19, 161–166. [Google Scholar] [CrossRef]
- Hu, S.; Wang, C.; Sanchez, D.L.; Lipka, A.E.; Liu, P.; Yin, Y.; Blanco, M.; Lübberstedt, T. Gibberellins promote brassinosteroids action and both increase heterosis for plant height in maize (Zea mays L.). Front. Plant Sci. 2017, 8, 1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feys, K.; Demuynck, K.; De Block, J.; Bisht, A.; De Vliegher, A.; Inzé, D.; Nelissen, H. Growth rate rather than growth duration drives growth heterosis in maize B104 hybrids. Plant Cell Environ. 2018, 41, 374–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, P.W.; Miller, F.R.; Quinby, J.R. Manipulation of sorghum growth and development with gibberellic acid. Agronomy 1977, 69, 789–793. [Google Scholar] [CrossRef]
- Williams, D.J.; Dancik, B.P.; Pharis, R.P. Early progeny testing and evaluation of controlled crosses of black spruce. Can. J. For. Res. 1987, 17, 1442–1450. [Google Scholar] [CrossRef]
- Bate, N.J.; Rood, S.B.; Blake, T.J. Gibberellins and heterosis in poplar. Can. J. Bot. 1988, 66, 1148–1152. [Google Scholar] [CrossRef]
- Dijkstra, P.; Kuiper, P.J.C. Effects of exogenously applied growth regulators on shoot growth of inbred lines of Plantago major differing in relative growth rate: Differential response to gibberellic acid and (2-chloroethyl)-trimethylammonium chloride. Physiol. Plant. 1989, 77, 512–518. [Google Scholar] [CrossRef]
- Dijkstra, P.; Ter Reegen, H.; Kuiper, P.J.C. Relation between relative growth rate, endogenous gibberellins, and the response to applied gibberellic acid for Plantago major. Physiol. Plant. 1990, 79, 629–634. [Google Scholar] [CrossRef]
- Pharis, R.P.; Yeh, F.C.; Dancik, B.P. Superior growth potential in trees: What is its basis, and can it be tested for at an early age? Can. J. For. 1991, 21, 368–374. [Google Scholar] [CrossRef]
- Rood, S.B.; Witbeck, T.J.; Major, D.J.; Miller, F.R. Gibberellins and heterosis in sorghum. Crop Sci. 1992, 32, 713–718. [Google Scholar] [CrossRef]
- Rood, S.B. Heterosis and the metabolism of gibberellin A20 in sorghum. Plant Growth Regul. 1995, 16, 271–278. [Google Scholar] [CrossRef]
- Rashid, M.A. Physiological basis of heterosis: Gibberellins and heterosis in eggplant. In CIMMYT, Book of Abstracts: The Genetics and Exploitation of Heterosis in Crops; International Symposium; International Maize and Wheat Improvement Center: Mexico City, Mexico, 1997; pp. 126–127. [Google Scholar]
- Pearce, D.W.; Rood, S.B.; Wu, R. Phytohormones and shoot growth in a three-generation hybrid poplar family. Tree Physiol. 2004, 24, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Ni, Z.; Yao, Y.; Nie, X.; Sun, Q. Gibberellins and heterosis of plant height in wheat (Triticum aestivum L.). BMC Genet. 2007, 8, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Yao, Y.; Peng, H.; Zhang, Y.; Lu, L.; Ni, Z.; Sun, Q. The relationship of differential expression of genes in GA biosynthesis and response pathways with heterosis of plant height in a wheat diallel cross. Chin. Sci. Bull. 2009, 54, 3029–3034. [Google Scholar] [CrossRef]
- Ma, Q.; Hedden, P.; Zhang, Q. Heterosis in rice seedlings: Its relationship to gibberellin content and expression of gibberellin metabolism and signaling genes. Plant Physiol. 2011, 156, 1905–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groszmann, M.; Gonzalez-Bayon, R.; Greaves, I.K.; Wang, L.; Huen, A.K.; Peacock, W.J.; Dennis, E.S. Intraspecific Arabidposis hybrids show different patterns of heterosis despite the close relatedness of the parental genomes. Plant Physiol. 2014, 166, 265–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Thomas, B.R. Hormones and heterosis in hybrid balsam poplar (Populus balsmifera L.). Forests 2019, 10, 143. [Google Scholar] [CrossRef] [Green Version]
- Hedden, P.; Sponsel, V. A century of gibberellin research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef] [Green Version]
- Rood, S.; Hedden, P. Convergent pathways of gibberellin A1 biosynthesis in Brassica. Plant Growth Regul. 1994, 15, 241–246. [Google Scholar] [CrossRef]
- Hedden, P. Gibberellin biosynthesis in higher plants. Annu. Plant Rev. 2016, 49, 37–72. [Google Scholar] [CrossRef]
- Zanewich, K.P.; Rood, S.B. Distribution of endogenous gibberellins in vegetative and reproductive organs of Brassica. J. Plant Growth Regul. 1993, 12, 41–46. [Google Scholar] [CrossRef]
- Zanewich, K.P.; Rood, S.B.; Southworth, C.E.; Williams, P.H. Dwarf mutants of Brassica: Responses to applied gibberellins and gibberellin content. J. Plant Growth Regul. 1991, 10, 121–127. [Google Scholar] [CrossRef]
- Diers, B.W.; McVetty, P.B.E.; Osborn, T.C. Relationship between heterosis and genetic distance based on restriction fragment length polymorphism markers in oilseed rape (Brassica napus L.). Crop Sci. 1996, 36, 79–83. [Google Scholar] [CrossRef]
- Hale, A.L.; Farnham, M.W.; Nzaramba, M.N.; Kimbeng, C.A. Heterosis for horticultural traits in broccoli. Theor. Appl. Genet. 2007, 115, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Abel, S.; Möllers, C.; Becker, H.C. Development of synthetic Brassica napus lines for the analysis of “fixed heterosis” in allopolyploid plants. Euphytica 2005, 146, 157–163. [Google Scholar] [CrossRef]
- Groszmann, M.; Gonzalez-Bayon, R.; Lyons, R.L.; Greaves, I.K.; Kazan, K.; Peacock, W.J.; Dennis, E.S. Hormone-regulated defense and stress response networks contribute to heterosis in Arabidposis F1 hybrids. Proc. Natl. Acad. Sci. USA 2015, 112, E6397–E6406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Wang, X.; Ren, D.; Huang, H.; Xu, M.; He, G.; Deng, X. Genomic architecture of biomass heterosis in Arabidposis. Proc. Natl. Acad. Sci. USA 2017, 114, 8101–8106. [Google Scholar] [CrossRef] [Green Version]
- Harper, F.R.; Berkencamp, B. Revised growth-stage key for Brassica campestris and B. napus. Can. J. Plant Sci. 1975, 55, 657–658. [Google Scholar] [CrossRef] [Green Version]
- Hoecker, N.; Keller, B.; Piepho, H.P.; Hochholdinger, F. Manifestation of heterosis during early maize (Zea mays L.) root development. Theor. Appl. Genet. 2006, 112, 421–429. [Google Scholar] [CrossRef]
- Zhu, A.; Greaves, I.K.; Liu, P.-C.; Wu, L.; Dennis, E.S.; Peacock, W.J. Early changes of gene activity in developing seedlings of Arabidopsis hybrids relative to parents may contribute to hybrid vigour. Plant J. 2016, 88, 597–607. [Google Scholar] [CrossRef]
- Meyer, S.; Pospisil, H.; Scholten, S. Heterosis associated gene expression in maize embryos 6 days after fertilization exhibits additive, dominant and overdominant pattern. Plant Mol. Biol. 2007, 63, 381–391. [Google Scholar] [CrossRef]
- Robbins, W.J. Growth of excised roots and heterosis in tomato. Am. J. Bot. 1941, 28, 216–225. [Google Scholar] [CrossRef]
- Blum, A.; Jordan, W.R.; Arkin, G.F. Sorghum root morphogenesis and growth. II. Manif. Heterosis Crop Sci. 1977, 17, 153–157. [Google Scholar] [CrossRef]
- Wang, L.; Wu, L.M.; Greaves, I.K.; Zhu, A.; Dennis, E.S.; Peacock, W.J. PIF4-controlled auxin pathway contributes to hybrid vigor in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2017, 114, E3555–E3562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elias, A.A.; Busov, V.B.; Kosola, K.R.; Ma, C.; Etherington, E.; Shevchenko, O.; Gandhi, H.; Pearce, D.W.; Rood, S.B.; Strauss, S.H. Green revolution trees: Semidwarfism transgenes modify gibberellins, promote root growth, enhance morphological diversity, and reduce competitiveness in hybrid poplar. Plant Physiol. 2012, 160, 1130–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaleel, C.A.; Manivannan, P.A.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.A.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Nassimi, A.W.; Raziuddin, A.; Sardar, N.; Naushad, A. Study on heterosis in agronomic characters of rapeseed (Brassica napus L.) using diallel. J. Agron. 2006, 5, 505–508. [Google Scholar] [CrossRef]
- Ali, M.; Copeland, L.O.; Elias, S.G.; Kelly, J.D. Relationship between genetic distance and heterosis for yield and morphological traits in winter canola (Brassica napus L.). Theor. Appl. Genet. 1995, 91, 118–121. [Google Scholar] [CrossRef]
- Stupar, R.M.; Gardiner, J.M.; Oldre, A.G.; Haun, W.J.; Chandler, V.L.; Springer, N.M. Gene expression analyses in maize inbreds and hybrids with varying levels of heterosis. BMC Plant Bio. 2008, 8, 33. Available online: http://www.biomedcentral.com/1471-2229/8/33 (accessed on 21 January 2020). [CrossRef] [Green Version]
- Rahman, H.; Bennett, R.A.; Yang, R.-C. Patterns of heterosis in three distinct inbred populations of spring Brassica napus canola. Crop Sci. 2016, 56, 2536–2545. [Google Scholar] [CrossRef] [Green Version]
- Muangprom, A.; Thomas, S.G.; Sun, T.P.; Osborn, T.C. A novel dwarfing mutation in a green revolution gene from Brassica rapa. Plant Physiol. 2005, 137, 931–938. [Google Scholar] [CrossRef] [Green Version]
- Ozga, J.A.; Reinecke, D.M. Hormonal interactions in fruit development. J. Plant Growth Regul. 2003, 22, 73–81. [Google Scholar] [CrossRef]
- Mather, K.; Jinks, J.L. Biometrical Genetics, 2nd ed.; Chapman and Hall: London, UK, 1971; pp. 65–68. [Google Scholar]
- Zanewich, K.P.; Rood, S.B. Vernalization and gibberellin physiology of winter canola (endogenous gibberellin (GA) content and metabolism of [3H]GA1 and [3H]GA20. Plant Physiol. 1995, 108, 615–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busov, V.; Meilan, R.; Pearce, D.W.; Rood, S.B.; Ma, C.; Tschaplinski, T.J.; Strauss, S.H. Transgenic modification of gai or rgl1 causes dwarfing and alters gibberellins, root growth, and metabolite profiles in Populus. Planta 2006, 224, 288–299. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a) Structure of gibberellin A1 (GA1), the bioeffector GA involved in the regulation of shoot growth and other processes in many higher plants. Numbers indicate positions of hydroxylations. (b) The early 13-hydroxylation GA biosynthetic pathway, including GAs and enzymes (blue; ox = oxidase) that were assessed in this study with Brassica or in the studies listed in Table 1 and Table 2. GA19 and GA20 are the primary GA1 precursors and GA4 provides an alternate precursor [16]. Following 2-hydroxylation, GA29 and GA8 (red) are biologically inactive.
Figure 1.
(a) Structure of gibberellin A1 (GA1), the bioeffector GA involved in the regulation of shoot growth and other processes in many higher plants. Numbers indicate positions of hydroxylations. (b) The early 13-hydroxylation GA biosynthetic pathway, including GAs and enzymes (blue; ox = oxidase) that were assessed in this study with Brassica or in the studies listed in Table 1 and Table 2. GA19 and GA20 are the primary GA1 precursors and GA4 provides an alternate precursor [16]. Following 2-hydroxylation, GA29 and GA8 (red) are biologically inactive.

Figure 2.
Dry mass of organs and whole plants of two Brassica inbreds and their F1 hybrid grown in greenhouse conditions (Means ± SE, n = 10).
Figure 2.
Dry mass of organs and whole plants of two Brassica inbreds and their F1 hybrid grown in greenhouse conditions (Means ± SE, n = 10).

Figure 3.
Contents of endogenous gibberellin A1 (GA1) and its precursors GA19 and GA20 in stems, and heights (Means ± SE provided for day 49, n = 10) of two Brassica inbreds and their F1 hybrid grown in greenhouse conditions. The inset plot displays the developmental stages at day 42, in accordance with Harper and Berkencamp [52].
Figure 3.
Contents of endogenous gibberellin A1 (GA1) and its precursors GA19 and GA20 in stems, and heights (Means ± SE provided for day 49, n = 10) of two Brassica inbreds and their F1 hybrid grown in greenhouse conditions. The inset plot displays the developmental stages at day 42, in accordance with Harper and Berkencamp [52].


{kind=link}
{kind=link}
{kind=link}
Table 1.
Studies of gibberellins (GAs) and heterosis for growth in maize, or corn, Zea mays L., sequenced chronologically (GA3 = gibberellic acid).
Table 1.
Studies of gibberellins (GAs) and heterosis for growth in maize, or corn, Zea mays L., sequenced chronologically (GA3 = gibberellic acid).
| Findings | Reference | Exogenous GA3 | Endogenous GAs | GA metabolism | Gene expression | Other |
|---|---|---|---|---|---|---|
| Higher sensitivity of inbreds to exogenous GA3 | [12] | X | ||||
| Higher levels of GA-like substances in shoot apices from a hybrid than parental inbreds | [17] | X | ||||
| Increased responsivity of inbreds, faster [3H]GA20 metabolism in a hybrid | [18] | X | X | |||
| Faster [3H]GA1 metabolism in a heterotic hybrid | [19] | X | ||||
| Higher levels of GA19 and GA1 in hybrids than parental inbreds | [15] | X | ||||
| Increased GA1 and correspondence with amylase and seedling growth in a hybrid family | [20] | X | X | |||
| Correspondences between GA3 responsivity and GA content across inbreds and hybrids | [21] | X | X | |||
| Heterosis persists with a GA dwarfing gene | [22] | X | ||||
| Increased levels of ‘gibberellin-stimulated transcript 1 like protein’ in hybrids | [23] | X | ||||
| Increased expression of GA 20-oxidase in hybrids over parental inbreds, in multiple tissues | [24] | X | ||||
| Genome-wide associations support GAs as promoting brassinosteroids and heterosis for height | [25] | X | ||||
| Induction of GA 20-oxidase promoted heterosis | [26] | X |
Table 2.
Studies of gibberellins (GAs) and heterosis in crop plants other than maize and trees, sequenced chronologically (GA3 = gibberellic acid).
Table 2.
Studies of gibberellins (GAs) and heterosis in crop plants other than maize and trees, sequenced chronologically (GA3 = gibberellic acid).
| Plant | Findings | Reference | Exogenous GA3 | Endogenous GAs | GA metabolism | Gene expression | Other |
|---|---|---|---|---|---|---|---|
| Tomato | Greater growth promotion by GA3 of slow-growing inbreds | [13] | X | ||||
| Sorghum | Greater growth promotion by GA3 of shorter varieties | [27] | X | ||||
| Spruce | Increased responsivity to GA4/7 in slower growing F1 crosses | [28] | X | ||||
| Poplar | Higher levels of GA-like substances in bark scrapings from hybrids, GA19 and GA1 identified | [29] | X | ||||
| Plantago | Slower growing inbreds were more responsive to GA3; a GA inhibitor reduced growth especially in faster growing lines | [30] | X | X | |||
| Plantago | Decreased apparent GA, by immunoassay, in a slower growing inbred | [31] | X | ||||
| Poplar | Higher GA-like substances in bark scrapings from hybrids than parents | [32] | X | ||||
| Sorghum | Higher GA1 in fast-growing hybrids than parents, 2 triplets | [33] | X | ||||
| Sorghum | Increased [3H]GA20 metabolism in fast- growing hybrids | [34] | X | ||||
| Eggplant | Shorter genotypes were more responsive to GA3 for inbreds and hybrids | [35] | X | ||||
| Poplar | No increase in levels in subapical internodes of GA44,20,29,1,8 but harvests were after the growth interval | [36] | X | ||||
| Wheat | Higher GA4 in hybrids; increased expression of GA 20-oxidase and GA 3-oxidase | [37] | X | X | |||
| Wheat | Heterosis for height correlated with expression of GA 20-oxidase and GA 3-oxidase variants | [38] | X | ||||
| Rice | Higher levels of GA53,44,4,1 but lower GA20, and increased expression of GA biosynthesis and action genes in hybrids | [39] | X | X | |||
| Arabidopsis | A GA biosynthetic inhibitor blocks heterosis | [40] | X | ||||
| Poplar | Across hybrids, growth positively correlated with GA8 but negatively correlated with GA19 and GA20 | [41] | X | ||||
| Brassica | Shoot elongation correlated with GA19,20,1 but slight heterosis, primarily for increased root growth | This study | X |
Table 3.
Characteristics of two Brassica inbreds (AO533 and AO539) and their F1 hybrid, grown in greenhouse conditions (± SE, n = 10). Statistical comparisons including these results along with measures from plants harvested on days 21 and 35 are provided in Table 4.
Table 3.
Characteristics of two Brassica inbreds (AO533 and AO539) and their F1 hybrid, grown in greenhouse conditions (± SE, n = 10). Statistical comparisons including these results along with measures from plants harvested on days 21 and 35 are provided in Table 4.
| Genotype | Seedling Emergence (%) | Foliar Characteristics at Day 42 | |||
|---|---|---|---|---|---|
| Day 4 | Day 7 | Leaf Number | Leaf Size (cm2) | Leaf Area (cm2) | |
| AO533 | 25.9 | 58.8 | 10.6 ± 0.4 | 32.5 ± 2.3 | 352.4 ± 37.5 |
| AO539 | 41.2 | 89.4 | 12.3 ± 0.3 | 30.3 ± 0.9 | 371.2 ± 16.4 |
| AO533 × AO539 | 71.8 | 95.3 | 12.8 ± 0.3 | 30.0 ± 1.7 | 384.6 ± 23.8 |
Table 4.
Analyses of covariance for growth characteristics of two Brassica inbreds (AO533 = “A3” and AO539 = “A9”) and their F1 hybrid grown in greenhouse conditions. Plants were harvested at 21, 35 and 42 days, and day provided the covariate. There were generally 99 plants producing F(2,96), except for GA content (F2,20) and *, ** = p < 0.05, p < 0.01, respectively. For comparisons, genotypes are sequenced by decreasing values with least significant difference (LSD) pairwise comparisons: ‘~’ not significantly different, or greater (‘≥’ = p < 0.1; ‘>’ = p < 0.05, and ‘>>’ = p < 0.01). The potence ratio (PR, averages from 3 harvests for each value) provides a measure of dominance with values substantially exceeding 1 indicating overdominance, or heterosis (PR > 1.5 in red). The PR value for GAs is inflated due to similarity of the two parents for some GA measures.
Table 4.
Analyses of covariance for growth characteristics of two Brassica inbreds (AO533 = “A3” and AO539 = “A9”) and their F1 hybrid grown in greenhouse conditions. Plants were harvested at 21, 35 and 42 days, and day provided the covariate. There were generally 99 plants producing F(2,96), except for GA content (F2,20) and *, ** = p < 0.05, p < 0.01, respectively. For comparisons, genotypes are sequenced by decreasing values with least significant difference (LSD) pairwise comparisons: ‘~’ not significantly different, or greater (‘≥’ = p < 0.1; ‘>’ = p < 0.05, and ‘>>’ = p < 0.01). The potence ratio (PR, averages from 3 harvests for each value) provides a measure of dominance with values substantially exceeding 1 indicating overdominance, or heterosis (PR > 1.5 in red). The PR value for GAs is inflated due to similarity of the two parents for some GA measures.
| Characteristic | Genotype F Value | Comparisons | Potence Ratio |
|---|---|---|---|
| Number of Leaves | 22.10 ** | F1 ~ A9 >> A3 | 1.57 |
| Leaf Size | 2.60 | F1 ~ A9 > A3 | 1.35 |
| Leaf Area | 5.76 ** | F1 ~ A9 > A3; F1 >> A3 | 1.81 |
| Leaf Mass | 8.53 ** | F1 ~ A9 >> A3 | 1.58 |
| Juvenile Height | 5.48 ** | F1 ~ A3 > A9; F1 >> A9 | 2.71 |
| Stem Mass | 4.45 * | F1 ~ A9 > A3; F1 >> A3 | 2.36 |
| Root Mass | 15.78 ** | F1 > A9 >> A3 | 2.64 |
| Total Plant Mass | 15.76 ** | F1 > A9 >> A3 | 2.23 |
| Gibberellins (GAs; content/stem) | 1.83 | F1 ~ A9 ~ A3 | 15.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zanewich, K.P.; Rood, S.B. Gibberellins and Heterosis in Crops and Trees: An Integrative Review and Preliminary Study with Brassica. Plants 2020, 9, 139. https://doi.org/10.3390/plants9020139
AMA Style
Zanewich KP, Rood SB. Gibberellins and Heterosis in Crops and Trees: An Integrative Review and Preliminary Study with Brassica. Plants. 2020; 9(2):139. https://doi.org/10.3390/plants9020139
Chicago/Turabian StyleZanewich, Karen P., and Stewart B. Rood. 2020. "Gibberellins and Heterosis in Crops and Trees: An Integrative Review and Preliminary Study with Brassica" Plants 9, no. 2: 139. https://doi.org/10.3390/plants9020139
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.