1. Introduction
Forest ecosystems are essential for capturing atmospheric carbon, which is then deposited in above- and below-ground biomass [
1]. The woody bole and branches where a substantial portion of atmospheric carbon is stored form this aboveground biomass [
2]. Landscape-level distribution of aboveground biomass has been well documented [
3] but the fundamental mechanisms of producing and retaining aboveground biomass, particularly in restored forest ecosystems is less explored. Logically, aboveground biomass is expected to increase during secondary succession [
4] but the time required to reach the maximum level of aboveground biomass might vary across forest ecosystems [
5]. The variation of aboveground biomass might be influenced by several factors including environmental variables such as soil properties and forest community characteristics such as plant taxonomic diversity (TD) and functional diversity. Functional diversity is defined as the distribution and relative abundance of functional traits in a community [
6]. Functional dispersion is one of the multidimensional indexes of FD which captures the spread of species traits from the centroid. It has been used in this article to represent FD. However, our understanding of the relative importance of functional and taxonomic diversity in affecting aboveground biomass in restored woody forest ecosystems as opposed to the herbaceous grassland ecosystem is relatively understudied.
Taxonomic diversity and plant functional diversity have linkage with aboveground biomass and soil properties [
7,
8]. Species richness has been positively linked with aboveground biomass from strata (understorey and overstorey) to the whole community [
7]. Moreover, species richness explained 28.5% of the variation in the total carbon stock in a subtropical forest ecosystem in southeastern China [
9]. Species richness creates more niche space and improves efficient nutrient utilization, therefore increases aboveground biomass [
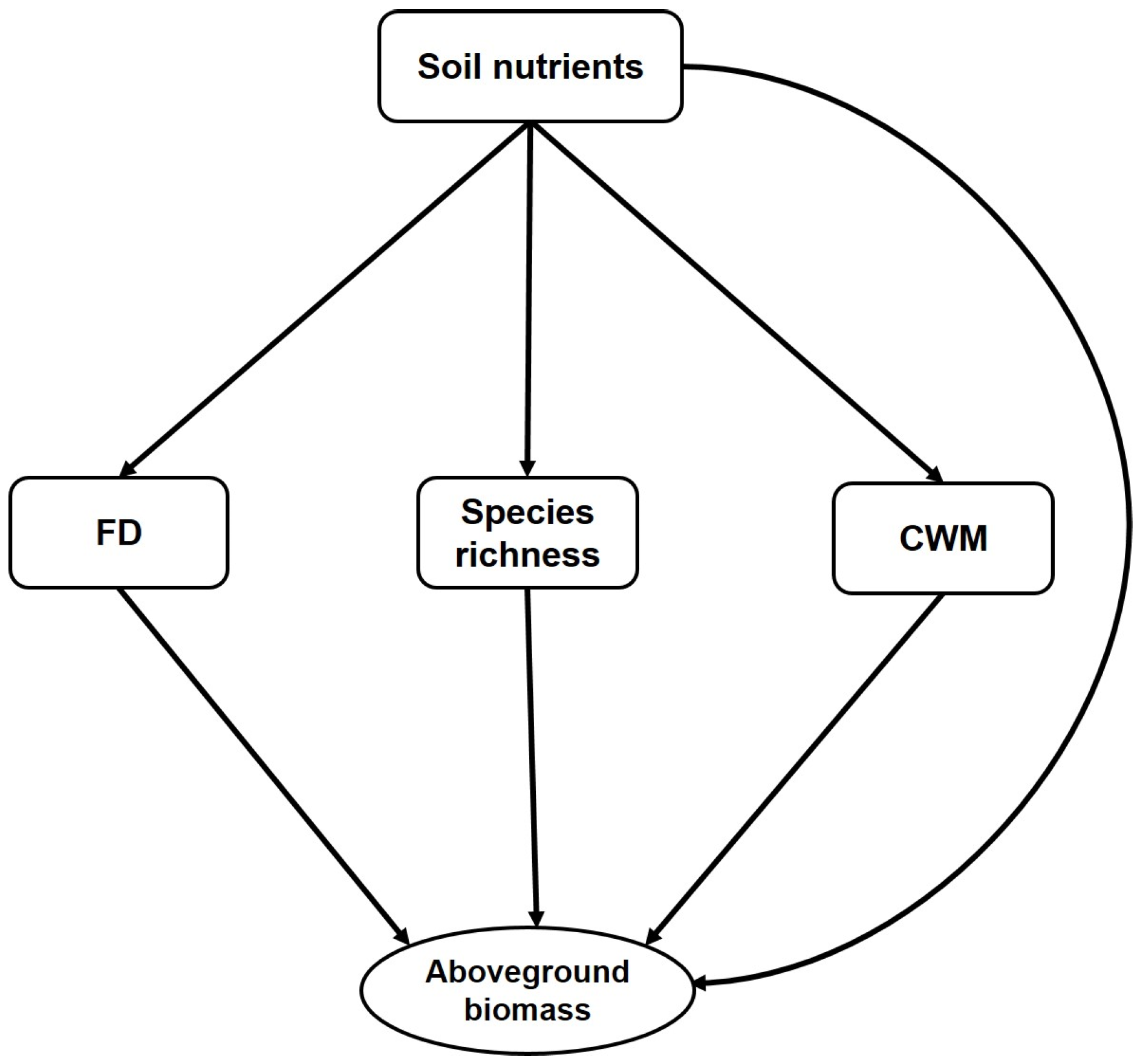
10]. According to our hypothesis (
Figure 1), the relationship between aboveground biomass and plant functional diversity fundamentally relies on species richness and soil nutrients. Functional diversity has important linkages with ecosystem processes. Functional diversity has been used for describing the variability in carbon stock deposited in the aboveground biomass [
11]. The relationship between carbon accumulation in the aboveground biomass and plant functional diversity depends upon the forest types and the diversity of functional traits in a plant community [
1,
12]. A higher diversity of traits associated with resource acquisition creates niche complementarity which allows the community to have greater access to the entire resource pool [
6]. Therefore, functional diversity should have a positive correlation with aboveground biomass when resource complementarity is the underlying driver. However, if the community has a higher abundance of productive species, then species richness would have a stronger relationship with aboveground biomass than functional diversity [
1].
There are two important hypotheses by which we can characterize the effect of functional diversity on aboveground biomass. The mass ratio hypothesis states that dominant species and their traits have a relatively stronger effect on ecosystem processes than do functionally diverse rare species [
13]. The mass ratio or community weighted mean (CWM) is the trait value of the species weighted by the species relative abundance in a community [
14,
15]. The niche complementary hypothesis posits that higher aboveground biomass in a forest community is regulated by diverse species that create more available niche space and the availability of diverse resources ensuring proper utilization of the resources [
12,
16,
17]. According to our hypothesis (
Figure 1), functional diversity, species richness, and CWM traits will correlate with and contribute to aboveground biomass. Previous empirical studies showed evidence of CWM and functional diversity alone [
18,
19] or jointly [
12] influencing aboveground biomass. Additionally, ecosystem processes can be directly influenced by soil properties by altering the ecosystem flux rates of matter and energy [
20]. Soil nutrients were also found to influence aboveground biomass indirectly through species richness and plant functional traits in tropical forests [
6,
20]. Therefore, the conceptual framework (
Figure 1) of this study postulates that soil nutrients will directly influence aboveground biomass and that species richness and plant functional diversity will do so indirectly. Previously we found that the taxonomic and phylogenetic diversity of the studied forest has been increased steadily during restoration, and deterministic and stochastic processes drove community composition of the forest [
21]. Former studies in other regions of the globe focused on the plant trait and ecosystem functioning linkages in artificially managed plant communities or environmental gradients. In these communities, functional diversity would be very low compared to under natural conditions. Furthermore, along environmental gradients, it is very difficult to differentiate the effects of plant communities on ecosystem functions and soil properties. Therefore, in realistic communities, the linkages between plant traits and ecosystem functions need to be explored [
22].
Plant functional traits have a direct and indirect effect on plant fitness and productivity [
23]. Aboveground biomass might be mechanistically correlated with the plant traits of a community related to the growth rate, and the ability to capture, store, and release carbon and resources [
19]. Important ecosystem functions such as nutrient cycling, respiration, and productivity were found to be strongly influenced by specific leaf area (SLA) and the leaf nitrogen content (LN) [
24]. SLA, LN, and the leaf dry matter content (LDMC) have been found to have a positive relationship with aboveground biomass [
6,
12,
18]. Moreover, it is important to identify key plant functional traits, both above- and below-ground, which can be used to predict individual or multiple functions of the ecosystem. Previous studies mainly focused on aboveground plant traits as the variables of plant functional diversity, but ecosystem functions can be directly influenced by root traits, so these need to be incorporated in the study of the biodiversity-ecosystem functioning relationship [
25]. Root traits have wider implications for soil processes likely to impact ecosystem functioning [
26,
27] and have been rarely studied.
Herein, we aim to decipher the relationship between biodiversity metrics (taxonomic and functional diversity) and ecosystem function (aboveground biomass) in a restored forest landscape in subtropical China. The forest landscape consists of four plantation types: an Acacia mangium (AM) plantation, a mixed Eucalyptus (EE) species plantation, a mixed coniferous species (MC) plantation, and a mixed native species (NS) plantation. We address the following questions: i) Which biodiversity metrics (taxonomic diversity or functional diversity) is the more important regulator of aboveground biomass? ii) What are the important plant functional traits driving aboveground biomass?
2. Results
The results of the Principal Component Analysis (PCA) of 15 functional traits (CWM) and 4 soil properties are shown in
Appendix Table A1. The first PC axis for CWM traits explained 91.44% of overall variability and the second PC axis accounted for 6.71% of overall variability (
Table A1). The first PC axis dominated by traits related to statute, morphological and elemental concentration in leaf and roots (
Table A1). In case of soil PCA, the first PC axis which explained 66.05% of overall variability, which mainly represented the variability in soil total carbon (SOC) and soil total nitrogen (TN). The second soil PCA axis, which explained 33.82% overall variance, mainly represented the variability in total phosphorus content (TP) and total potassium content (TK) in soil (
Table A1).
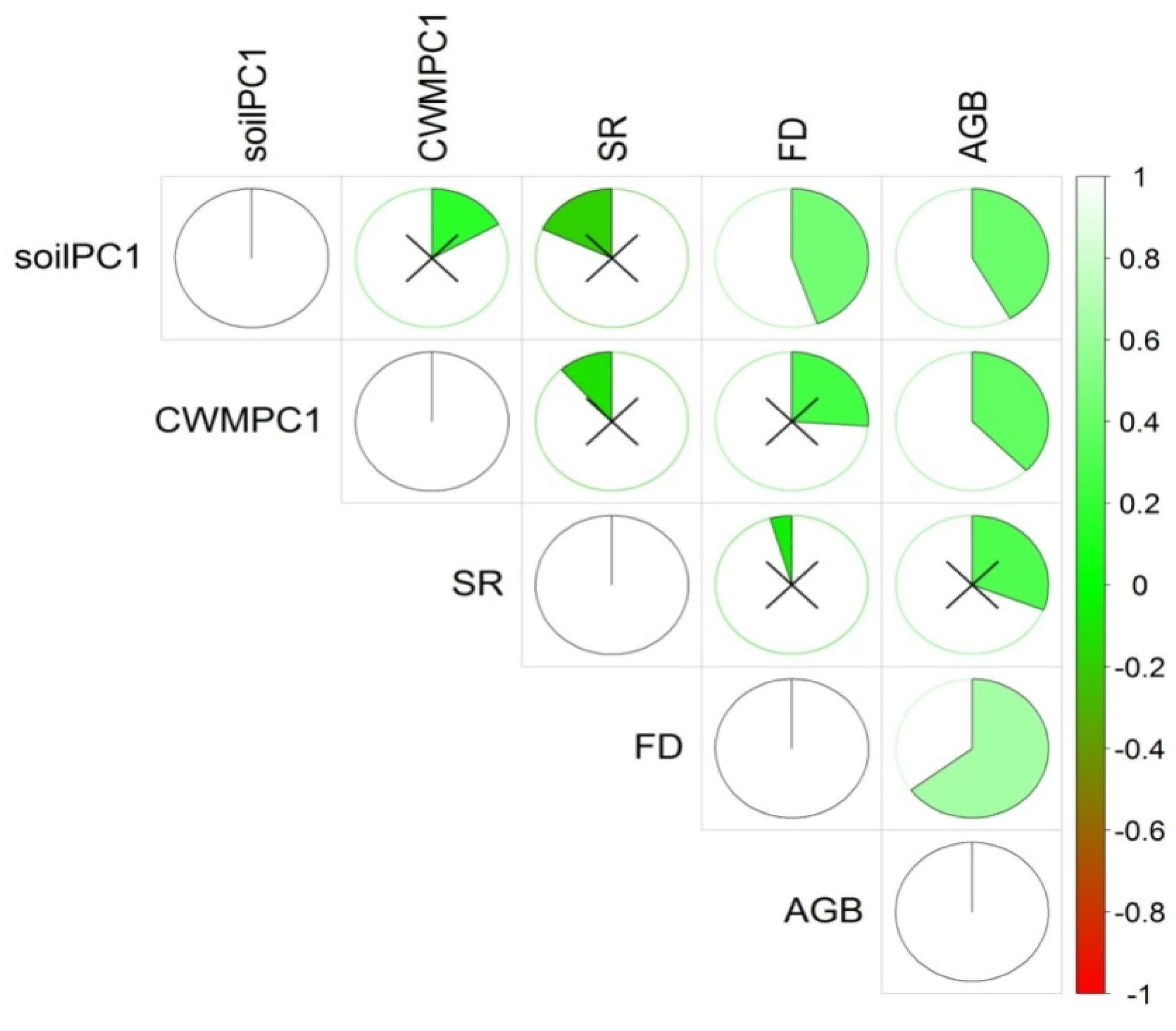
The ordinary least squares (OLS) linear regression analysis showed that aboveground biomass was significantly (
p < 0.01) and positively associated with TD and FD (
Figure 2a,b). The relationship of aboveground biomass with FD (R
2 = 0.45) was stronger than that with TD (R
2 = 0.13). FD (R
2 = 0.45 vs. 0.13) explained more variations in aboveground biomass than did TD. Aboveground biomass was also significantly related to CWM (R
2 = 0.08) traits (
Figure 2c) and soil nutrients (R
2 = 0.22) (
Figure 2d). Soil nutrients had a significant (
p < 0.01) relationship with FD and nonsignificant (
p > 0.05) relationship with TD and CWM traits (
Figure 2e,f). The analysis of covariance (ANCOVA) showed that plantations types had an influence on aboveground biomass (AGB) and TD bivariate relationships, while the other bivariate relationship between AGB and predictor variables was not influenced by plantation types (
Table A2).
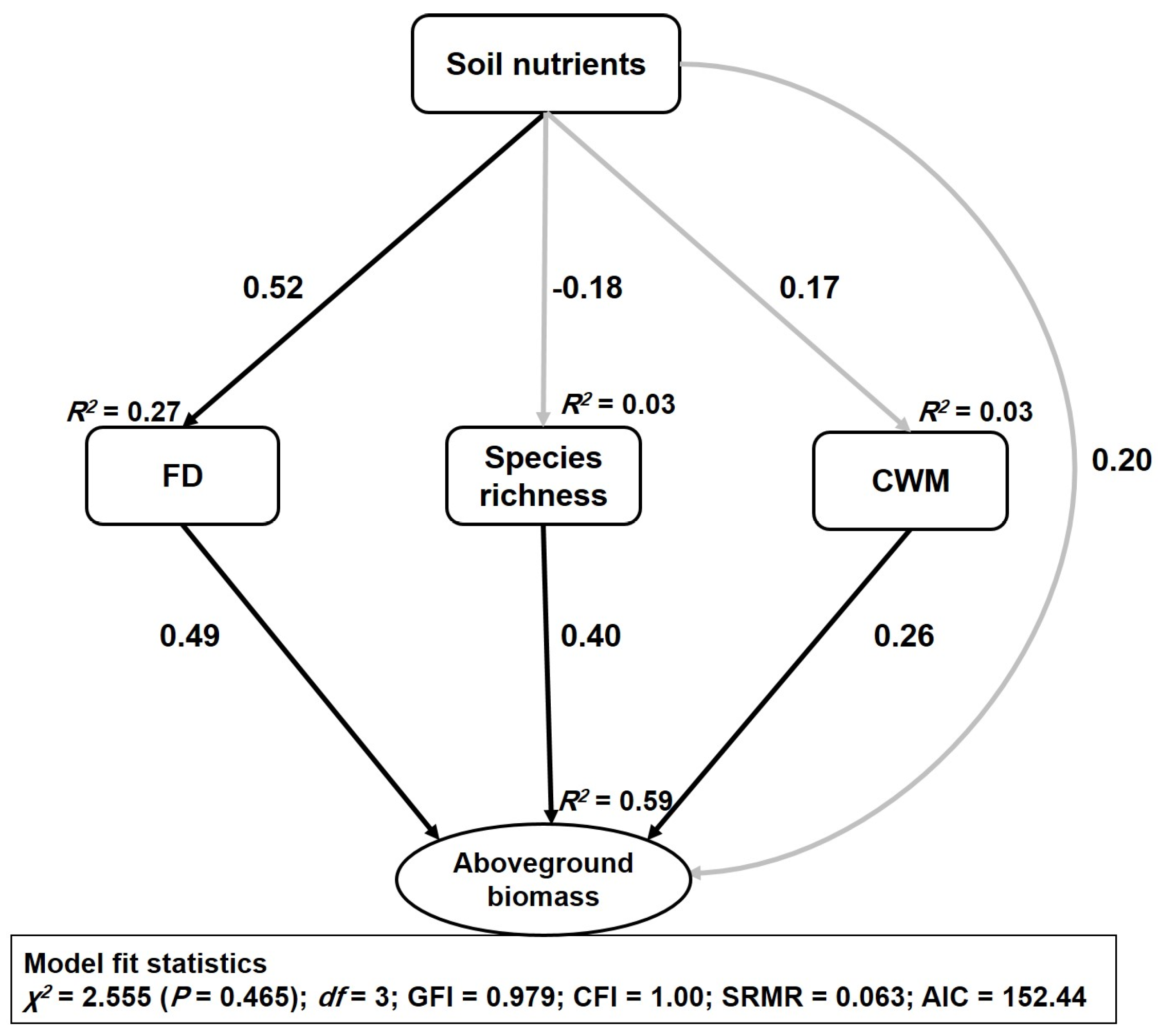
Structural equation modelling (SEM) accounted for 59% of the variation in aboveground biomass (
Figure 3). FD had the strongest positive direct relationship with aboveground biomass (β = 0.49,
p < 0.001), followed by species richness (β = 0.40,
p < 0.001) and CWM traits (β = 0.26,
p < 0.01). The direct effect of soil nutrients on aboveground biomass was found to be nonsignificant (β = 0.20,
p > 0.05), but the indirect effect of soil nutrients through FD was found to be significant (β = 0.25,
p < 0.01). The SEM also attributed 27% variation to FD and 3% variation to each of the species richness and CWM traits (
Figure 3).
Soil nutrients had a significant direct positive relationship with FD (β = 0.52,
p < 0.001) (
Table 1), but the relationship was nonsignificant between soil nutrients, species richness, and CWM traits (
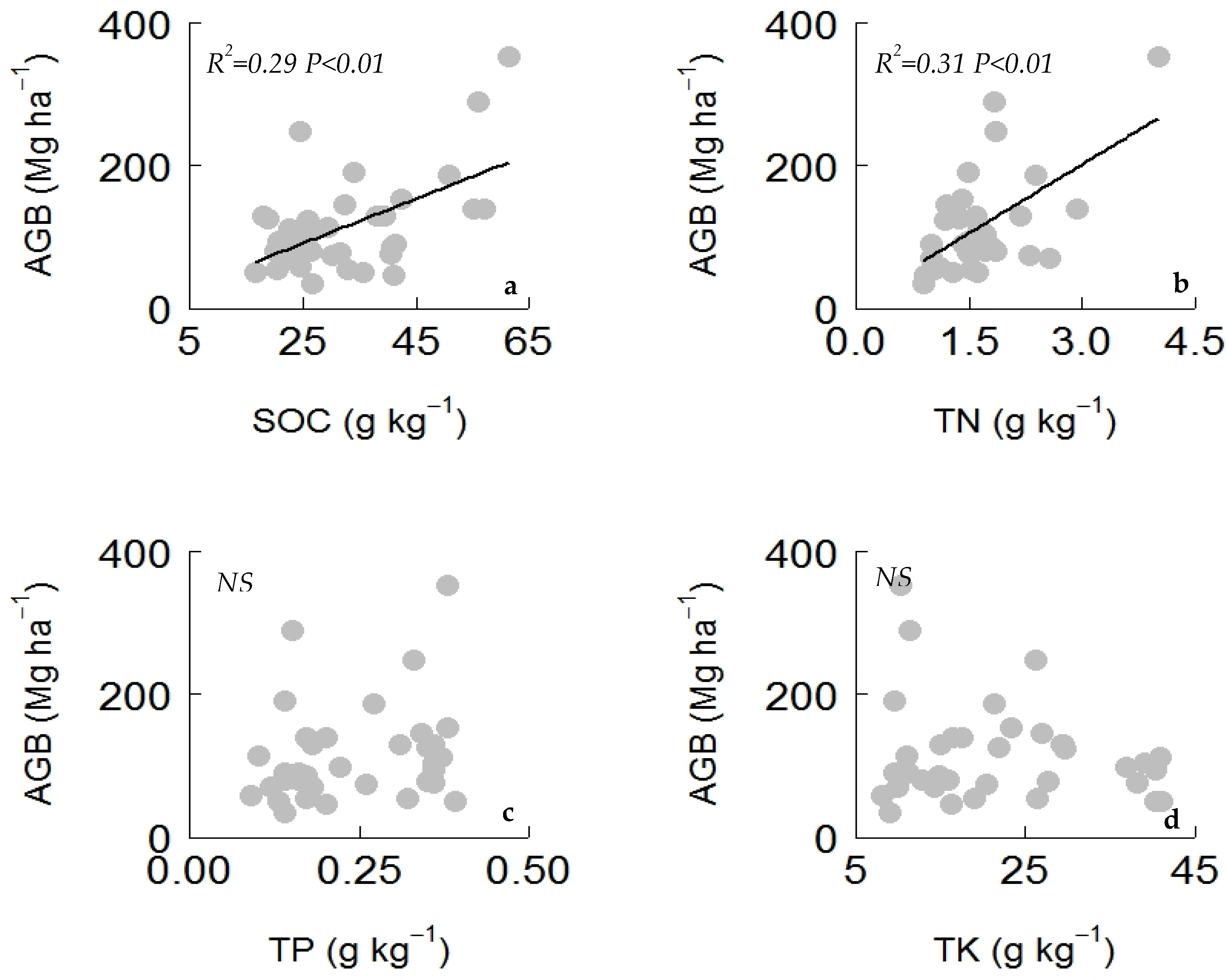
Figure A1). SOC and TN had a significant relationship with aboveground biomass, while the relationship of TP and TK with aboveground biomass was found to be nonsignificant (
Figure A2). The AGB and TN relationship was influenced by the plantation type, while other bivariate relationships was not influenced by plantation types (
Table A2).
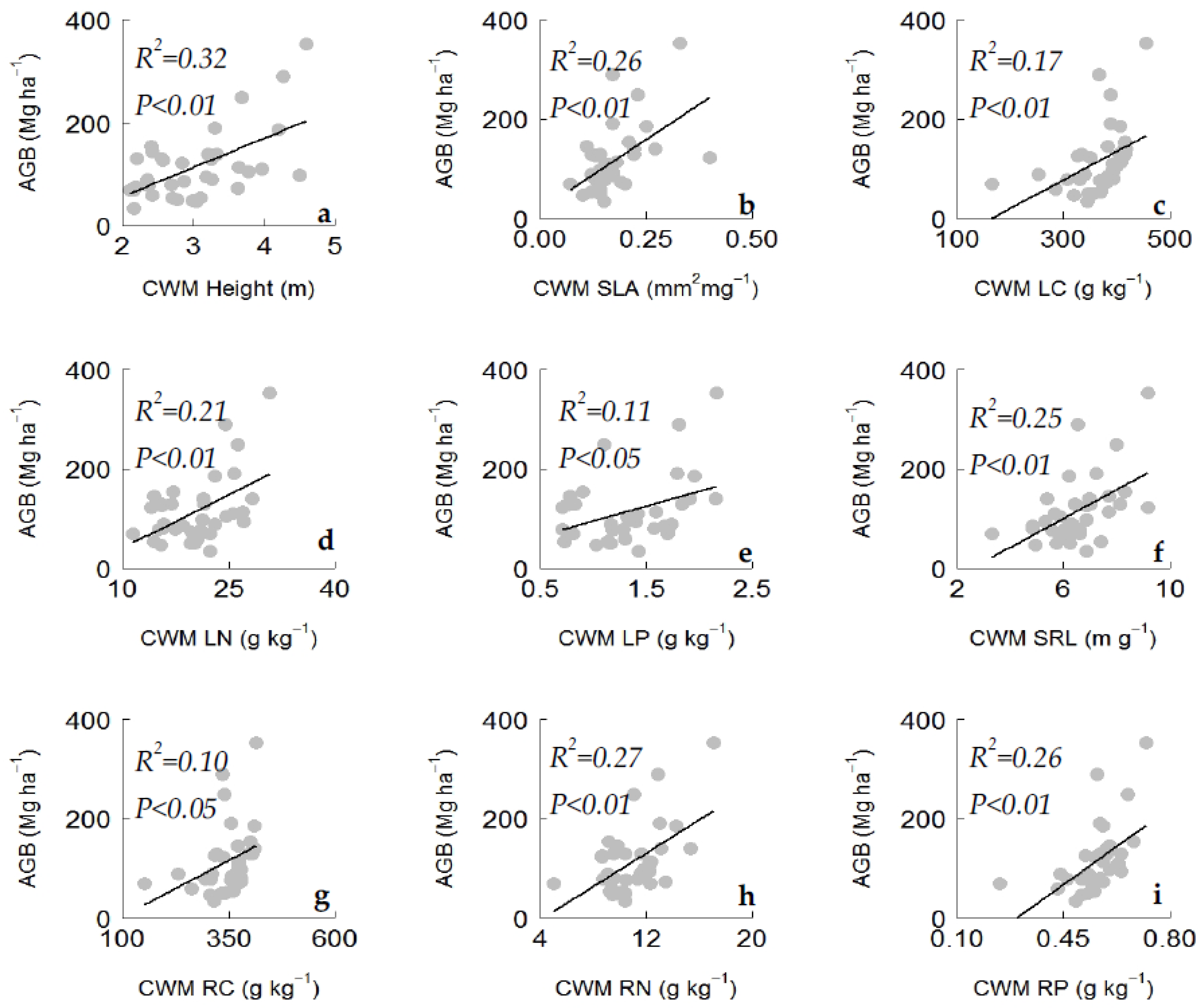
Random forest modelling was applied to identify the most important plant functional traits that contribute in predicting variations in aboveground biomass. The random forest model explained 33% of the variability in aboveground biomass (
Table A3). Plant functional traits related to carbon (leaf carbon content, LC; root carbon content, RC), nitrogen (LN, RN), and phosphorus (leaf phosphorus content, LP; root phosphorus content, RP) were found to be the dominant predictors of aboveground biomass (
Figure 4). Plant height, SLA, and root tissue density (RTD) were also found to be among the most important predictors of aboveground biomass (
Figure 4). We also tested the bivariate relationships between aboveground biomass and the significant plant traits identified by random forest analysis. We found that the plant traits were significantly and positively related to aboveground biomass (
Figure 5). The ANCOVA results showed that plantation type had an influence on the AGB–LN and AGB–RN relationships (
Table A2), while other bivariate relations was not influenced by plantation types. The results of the bivariate relationships and PCA indicated the existence of a leaf and root economic spectrum with an acquisitive resource use strategy (
Table A1).
The correlation analysis showed that SOC was positively (
p < 0.05) correlated with the CWM of the plant height, SLA, LC, root dry matter content (RDMC), and RC (
Table A4), while TN was positively and significantly (
p < 0.05) correlated with plant height, SLA, LDMC, LN, RDMC, and RN (
Table A6). TP was positively (
p < 0.05) correlated with specific root length (SRL), RP, RTD, root branching intensity (RBI) and negatively correlated with LP (
Table A4).
3. Discussion
Understanding how biodiversity attributes determine ecosystem functioning (aboveground biomass) is one of the central goals of ecology. This empirical study focused on the relative importance of taxonomic (species richness) diversity and functional diversity in regulating aboveground biomass. Consistent with our hypothesis, the aboveground biomass was found to have a positive relationship with taxonomic and functional diversity (
Figure 2). The results from OLS regression and SEM analysis revealed that FD had relatively more explanatory power in predicting variation in the aboveground biomass than did TD in the restored forest ecosystem (
Figure 3). Higher FD might provide lower niche overlap among the constituent species and ensure maximum utilization of the available resources which, in turn, can positively influence the aboveground biomass [
28]. The dominant species and their functional traits (mass-ratio hypothesis) also increased aboveground biomass. The positive relationship of aboveground biomass with CWM and FD indicates that increased aboveground biomass in the restored forest is driven by the dominant species with rapid acquisitive strategies; and diverse species with diversified nutrient use strategies [
1,
12]. Therefore, aboveground biomass in the restored forest is regulated by both the mass-ratio hypothesis and the niche complementarity hypothesis (
Figure 3). The findings of this study—that aboveground biomass in the restored forest is jointly regulated by the mass-ratio and niche complementarity mechanisms—are consistent with the findings of Ali et al. [
12] but contrast with empirical studies that did not find support for either of the two mechanisms [
14,
29]. Moreover, species richness might have increased aboveground biomass through greater resource capture and efficient utilization of the available resources [
9,
30]. Additionally, diversity of resources due to diverse species richness across plantation types might have influenced aboveground biomass. Consistent with our findings, species richness was also previously found to have a positive link with aboveground biomass in a subtropical forest [
6]. However, some empirical studies found that species richness had a weak or negative relationship with aboveground biomass in sites where the heterogeneity of environmental factors was dominant [
1,
3]. These results, that plant functional diversity better explained the aboveground biomass than did species richness, are in accordance with previous studies on secondary forests and grassland [
18,
31].
Plant functional traits that enhance acquisition, sustain plant growth, and allow proficient use of natural resources (solar radiation) and nutrients are essential for sustainable increases in aboveground biomass [
20,
32]. In our study, plant functional traits related to growth and nutrient acquisition and utilization were found to be the dominant predictors of aboveground biomass. The PCA results revealed that the first PCA axis was driven by CWM traits related to morphology and elemental concentration in plant tissues, representing a community-level leaf and root economic spectrum. These results also suggest that at the community level, plant traits associated with construction cost (leaf and root) and resource acquisition potential might covary, highlighting the presence of an economic spectrum (leaf and root) in the restored forest landscape. Moreover, the PCA loadings and bivariate relationships suggest an acquisitive strategy that tends to influence aboveground biomass. The increase in aboveground biomass with increasing plant height might be due to the presence of fast-growing tall species that are proactive in their efficient utilization of solar radiation. A positive link between aboveground biomass and plant height was previously reported in several empirical studies [
1,
2,
12,
29]. The positive relationship between CWM SLA and aboveground biomass increase might be related to the leaf economic spectrum [
15], the exploitative species with high SLA facilitate rapid nutrient acquisition and turnover, which might enhance rapid growth and maximize aboveground biomass [
24]. Indeed, the CWM of SLA following the carbon gain concept [
33] might facilitate higher carbon accumulation; the positive effect of SLA on stand level productivity was also found in tropical rainforests of Australia [
34]. The presence of a maximum number of unshaded leaves (e.g., in
Eucalyptus and coniferous vegetation) is another probable reason for the positive relationship between aboveground biomass and CWM SLA. Therefore, high aboveground biomass in forest communities might be associated with a high proportion of unshaded leaves [
12]. Furthermore, the significant increase in aboveground biomass with increasing CWM LC, CWM LN, and CWM LP also suggests that short lived leaves (high SLA), having a higher leaf elemental concentration through their canopy properties, might improve soil properties by adding nutrients into the soil, which, in turn, might influence aboveground biomass [
11]. These findings are in agreement with previous empirical studies in a tropical forest [
29] and successional biomes of temperate [
8] and tropical regions [
12], which implies that the presence of a leaf economic spectrum [
24] will have a significant impact on canopy properties and ecosystem processes.
Our results also found that high aboveground biomass is associated with root traits. The positive associations of CWM SRL and CWM RN with aboveground biomass indicate a root economic spectrum [
26]. SRL and RN have a strong effect on nitrification and have linkage with the plant nitrogen cycling [
27]. The rapid acquisition and turnover of fine roots can enhance nutrient cycling which might facilitate plant growth and increased aboveground biomass. The positive association among CWM RN and aboveground biomass might be due to the leguminous species (e.g., the
Acacia mangium in AM plantation) which fixes atmospheric nitrogen and added to the soil. Moreover, phosphorus is considered to be the most limiting nutrient in the tropical forest ecosystem [
30]; thus, an increase in CWM RP might increase phosphorus cycling and more available phosphorus might facilitate plant growth and increase the aboveground biomass of the restored forest community. Therefore, high aboveground biomass in the restored forest is scaled up by the economic spectra and the acquisitive strategies in the leaf and root traits. Moreover, the SEM did not find a significant direct effect of soil nutrients on aboveground biomass, but the bivariate relationship found a positive effect of SOC and TN on aboveground biomass (
Figure A2). These results indicate that an increase in SOC and TN during forest restoration through plant-mediated inputs might positively (
Table A2) influence plant growth and aboveground biomass [
20,
35]. FD had a positive relationship with soil nutrients, indicating that soil properties (mainly SOC and TN as suggested by the PC1 loadings) favor a higher dispersion of functional traits that might facilitate the germination of dispersed species or survival of planted/dispersed species. Moreover, SOC and TN were positively correlated with CWM traits (e.g., SLA, LC, LN, RC, RN), suggesting that the dominant species of the community have higher carbon and nitrogen use efficiency [
36]. Fast-growing species possessing acquisitive traits might facilitate litter (above and belowground) decomposition and rapid turnover to the soil which in turn might significantly influence the aboveground biomass [
32]. Therefore, the greater nutrient retention and resource use capacity might support plant growth and aboveground biomass [
37].



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







