A Review on Si Uptake and Transport System


Department of Ecology, Environment and Plant Sciences Stockholm University, 10691 Stockholm, Sweden
*
Author to whom correspondence should be addressed.
Plants 2019, 8(4), 81; https://doi.org/10.3390/plants8040081
Submission received: 4 February 2019
/
Revised: 4 March 2019
/
Accepted: 26 March 2019
/
Published: 29 March 2019
Abstract
:Silicon (Si) was long listed as a non-essential component for plant growth and development because of its universal availability. However, there has been a resurgence of interest in studying the underlying uptake and transport mechanism of silicon in plants because of the reported dynamic role of silicon in plants under stressed environmental conditions. This uptake and transport mechanism is greatly dependent upon the uptake ability of the plant’s roots. Plant roots absorb Si in the form of silicic acid from the soil solution, and it is moved through different parts of the plant using various influx and efflux transporters. Both these influx and efflux transporters are mostly found in the plasma membrane; however, their location and pattern of expression varies among different plants. The assessment of these features provides a new understanding of different species-dependent Si accumulations, which have been studied in monocots but are poorly understood in other plant groups. Therefore, the present review provides insight into the most recent research exploring the use of Si transporters in angiosperms and cryptogams. This paper presents an extensive representation of data from different families of angiosperms, including monocots and eudicots. Eudicots (previously referred to as dicots) have often been neglected in the literature, because they are categorized as low/intermediate Si accumulators. However, in this review, we attempt to highlight the accumulating species of different plant groups in which Si uptake is mediated through transporters.
1. Introduction
Silicon holds a unique position in plant biology because of its relative abundance (28.8%) in the Earth’s crust [1]. Though, not deemed to be a vital element for most plants, its uptake is associated with a number of benefits impacting normal growth and development. Crop plants, such as rice and sugarcane, are sometimes supplemented with Si fertilizers to enhance plant yield and crop quality [2,3]. The benefits of Si include resistance to biotic and abiotic stresses, such as drought, low temperature, salinity, metal toxicity, diseases and pests, enhancement of secondary metabolites in various medicinal plants, nutrient imbalances, and stimulation of photosynthesis [4,5,6]. The essentiality of Si has always been a topic of debate, as Si does not alter the normal growth and development of plants. Nevertheless, debates regarding essentiality must consider that food products derived from Si-exposed plants offer greater bone strength and improved nervous and immune system functions in human beings [7]. However, the beneficial effects of Si are greatly hampered by the absorption ability of plants, which, in turn, results in the arbitrary classification of plants as low accumulators (<0.1% Si), intermediary accumulators (1%), and high accumulators (more than 5% Si on dry weight basis) with respect to Si absorption [8]. For instance, rice, which is a monocot, accumulates up to 10% Si in shoots, whereas cucumber, a eudicot, is an intermediate accumulator that accumulates less Si than rice, and tomato, which is also a eudicot, is an extremely low-accumulating plant [9]. Therefore, in general, it is believed that monocots are high Si accumulators compared with eudicots. Different plants of the monocot families, such as Poaceae and Cyperaceae, accumulate more Si (>4%) than eudicot clades, such as Urticaceae and Cucurbitaceae, which accumulate 2–4% Si [10,11]. Table 1 shows different plant species with high, low, and intermediate degrees of Si accumulation in different groups of plants.
Therefore, efforts were made in this paper to corroborate the uptake of Si in different plant groups. Collective data representation was developed for Si transporters in angiosperms (monocots and eudicots) and cryptogams. Here, we focused on the channels/transporters responsible for Si uptake and transport in different groups of plants.
2. Mode of Cellular Si Uptake and Accumulation
Si content varies greatly in different plant tissues, as it undergoes several chemical transformations during the storage, deposition and transport phases. More often, it is also due to the differences in the uptake and transport mechanism, which takes place in plants [13,14]. Three different types of water-based Si uptake mechanisms have been anticipated based upon the Si content in plants and their uptake rates relative to water [15,16]. Plants having higher Si uptake levels than water are classified as active (e.g., rice, wheat, and barley), whereas those with similar rates are classified as passive (e.g., oat) and those with lower uptake rates are classified as rejective. The uptake mechanism of Si requires specific transmembrane proteins identified as Si transporters to mediate the whole process [17]. These protein channels are passive transporters, which do not require energy to transport silicic acid across the plasma membrane. Some examples of plants with energy-dependent Si transport include rice, barley, maize, wheat, banana, and cucumber [18].
Si is transported from roots to shoots as silicic acid [Si (OH)4], which at biological pH has the ability to move across the plasma membrane [19]. In a comparative analysis of high, medium, and low Si-accumulating plants such as rice, cucumber, and tomato, it was observed that Si transport is facilitated by transporters (Km value 0.15 mM) across the cortical cells, whereas Vmax value followed a decreasing trend from rice to cucumber to tomato, thereby showing a significant variation in the transport density. Following primary uptake from the roots, Si is further transported to shoots via xylem. In shoots, silicic acid is polymerized into silica gel due to loss of water (transpiration) and is usually deposited in different plant parts [20]. For example, in rice, Si is deposited as a dense layer (2.5 μm) beneath the cuticle layer (0.1 μm), thus acting as a first line of defense against various kinds of stresses [21]. It is also deposited in bulliform cells of the leaf epidermis of durum wheat and forms a 1–4 μm thick layer, which provides mechanical support and protection [22].
3. Si Facilitation via Transporter Genes
3.1. Angiosperms
3.1.1. In Monocots
The main reservoir of Si (50–70%) in the form of Si dioxide is in the solid phase of soil. However, its availability to plants is relatively low. The mechanism of uptake of Si varies amongst different plant species and apparently depends on the presence of specific Si transporters. The very first report of the identification of Si transporters in plants was in rice [23]. More recently, transporters were also exposed in other plants including barley, wheat, and pumpkin [24,25]. The discovery of such evidence in rice, which suggests that the uptake mechanism of Si is quite active and even faster than that of water, made it quite clear that a specific system exists in rice that facilitates the uptake of Si. This led to the identification and cloning of Lsi1 genes using a defective rice mutant, which was believed to be responsible for Si uptake in rice [26]. Transporter genes were localized on chromosome 2, with cDNA of 1409 bp consisting of 5 exons and 4 introns. Blast and ClustalW analysis showed that this transporter belongs to a NIP3 major protein family and an aquaporins subfamily [27]. When present in the plasma membrane, this transporter facilitates Si uptake into the root stele, which is otherwise prevented by the presence of casparian strips. Lsi1 is expressed in the main and lateral roots but not in the root hairs, thereby justifying a previously reported study that found that root hairs lack participation in the Si uptake process [28]. Also, when cRNA-encoded Lsi1 was inserted into a Xenopus laevis oocyte, it was seen that silicic acid transport activity increased significantly. All these findings confirm that Lsi1 is a major influx transporter of Si into rice roots.
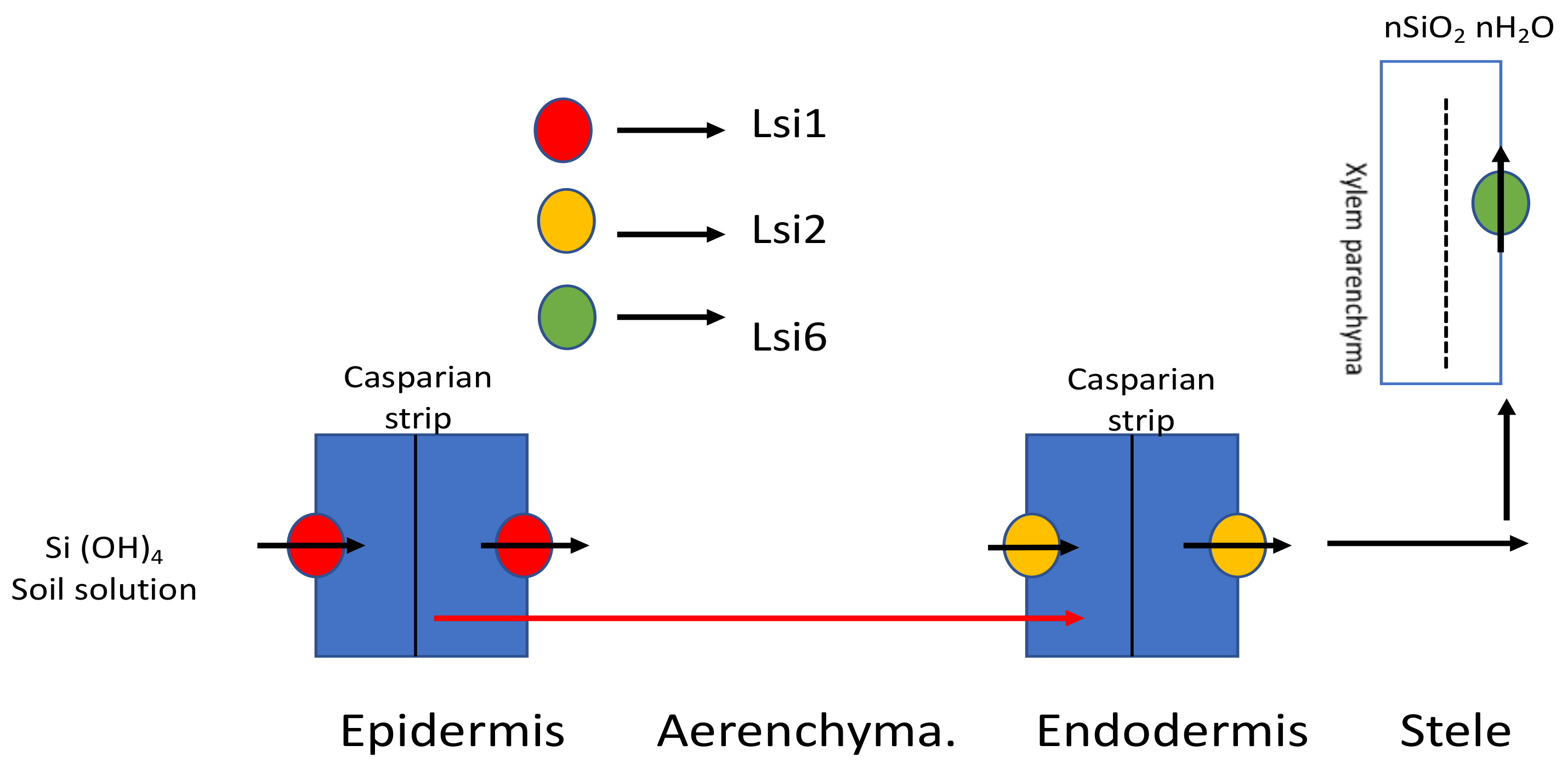
Lsi2, belonging to an anion transport family, facilitates Si uptake from the roots to the vascular tissues in rice [29,30]. Lsi2 shares location similarity with Lsi1 but lies towards the proximal side of the membrane, building up in the mature root zone instead of the root tips. Moreover, with injection into Xenopus oocytes, Lsi2 showed completely efflux activity rather than any influx activity for silicic acid. Another Si transporter Lsi6, which moves silicic acid across the vascular bundle, has been identified in rice [31]. Lsi6 is expressed in the parenchyma cells of the leaves. The absence of Lsi6 has no negative effect on Si uptake except that it can lead to excess silicified epidermal depositions and also outburst the flow into the guttation fluid [32]. Si uptake and transport using Si transporters is depicted in Figure 1.
Influx transporter (Lsi1) takes silicic acid from soil solution up to the exodermis, followed by the efflux transporter (Lsi2), which takes it further across the aerenchyma. Then, it is further moved up the aerial parts of the plants by another influx transporter, Lsi6 [32].
Erysiphe graminis f.sp. tritici causes powdery mildew in wheat, which is inhibited by the addition of Si to the wheat nutrient solution. This finding led to the discovery of TaLsi1, an influx transporter of Si [25,33]. This transporter was found to be quite similar to the OsLsi1 influx transporter in rice in terms of localization and activity in oocytes. In another study, discovery of the HvLsi1 influx transporter was made in barley; it was located in the distal side of the plasma membrane and showed 81% similarity with rice influx transporter, OsLsi1 [34]. Despite such a high degree of similarity, it differed in terms of expression patterns. Maize (Zea mays L.) is a high Si accumulator crop plant, in which Si uptake is mediated by two genes ZmLsi1 and ZmLsi6. ZmLsi1 is located in the roots, while ZmLsi6 is located in the parenchyma cells of the leaves. When expressed in the Xenopus laevis oocyte, both showed higher levels of permeability towards silicic acid.
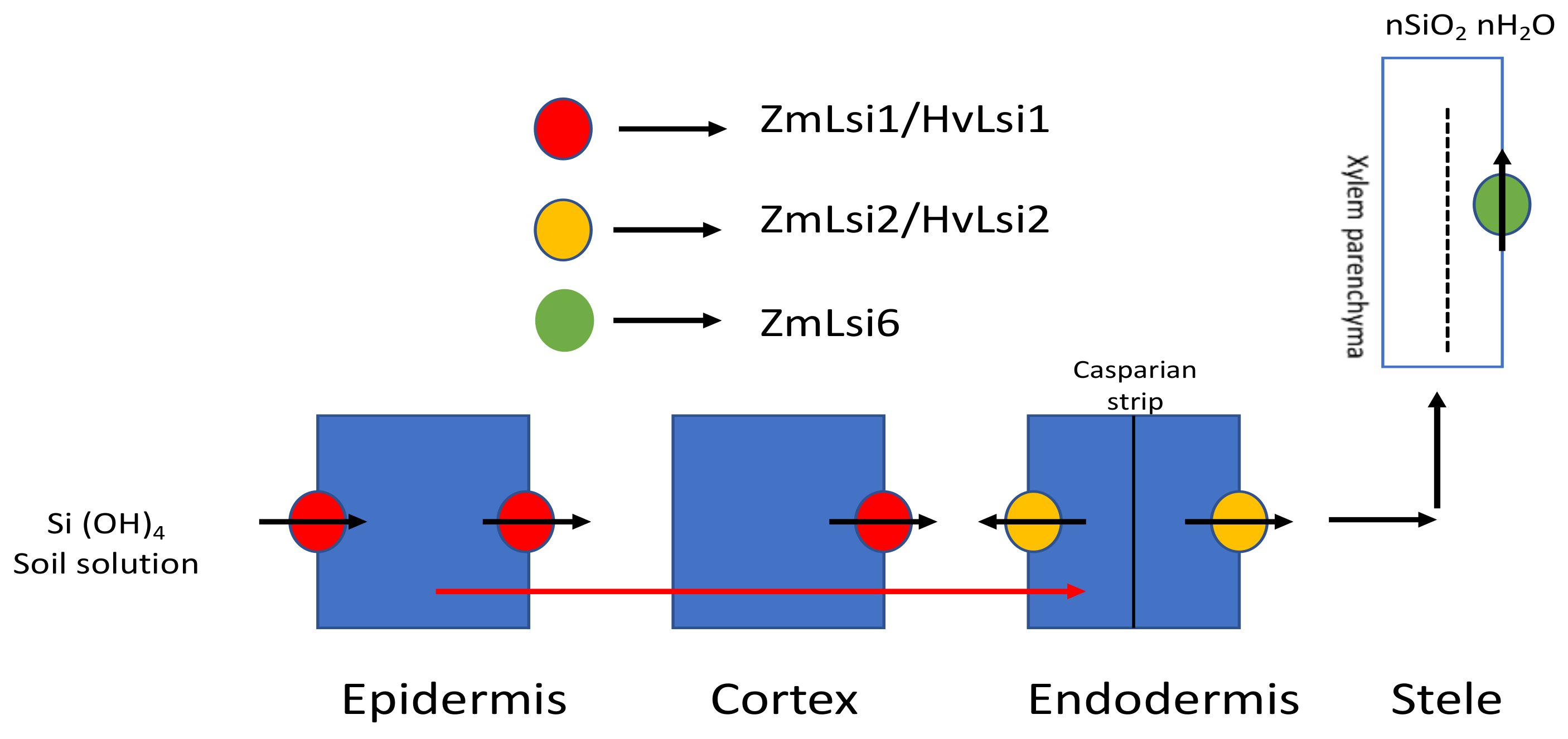
OsLsi2 efflux homologues have also been found in barley (HvLsi2) and maize (ZmLsi2), although they have more than 80% similarity with Lsi2 but differ in terms of localization, as both are present only in the endodermis and do not show polar localization [32]. HvLsi2 and ZmLsi2 also share functional similarity with OsLsi2 in terms of activity and expression patterns. In corn, ZmLsi6, an influx Si transporter similar to OsLsi6, has also been isolated and characterized [35]. ZmLsi6 is mainly responsible for xylem unloading and is localized in the parenchyma cells of xylem. There is a disparity in the uptake system between different plants, which is attributed to different root structures. In rice roots, there are two casparian strips at the exodermis and endodermis, whereas in maize and barley roots there is one casparian strip at the endodermis. Figure 2 shows the Si uptake mechanism mediated by different transporters in maize and barley.
3.1.2. In Eudicots
Analysis of the literature reveals that little information is available about Si transporters in eudicots, except with regard to a few members of the Cucurbitaceae, Urticaceae, and Asteraceae families, which show additional benefits with Si applications [11,24]. In pumpkin (Cucurbita moschata), CmLsi1 was the first Si influx gene found in eudicots [24]. Two different pumpkin cultivars (bloom and bloomless Cucumis sativus) were studied for Si uptake activity and expression patterns. Heterogeneous expression in the Xenopus oocyte showed enhanced Si influx activity in the rootstock from the bloom pumpkin (CmLsi1B+), whereas no activity was reported in the bloomless rootstock (CmLsi1B−). Unlike the rice transporter, bloom pumpkin transporters were localized in the plasma membrane, while bloomless pumpkin transporters were found in the endoplasmic reticulum. Two efflux Si transporters have also been isolated and characterized in pumpkin [36]. CmLsi2-1 and CmLsi2-2 seem to be expressed in both roots and shoots with no sequence disparity. This confirms that if there is a mutation in CmLsi1, it may lead to low Si uptake in bloomless pumpkins rather than any mutation in CmLsi2-1 and CmLsi2-2.
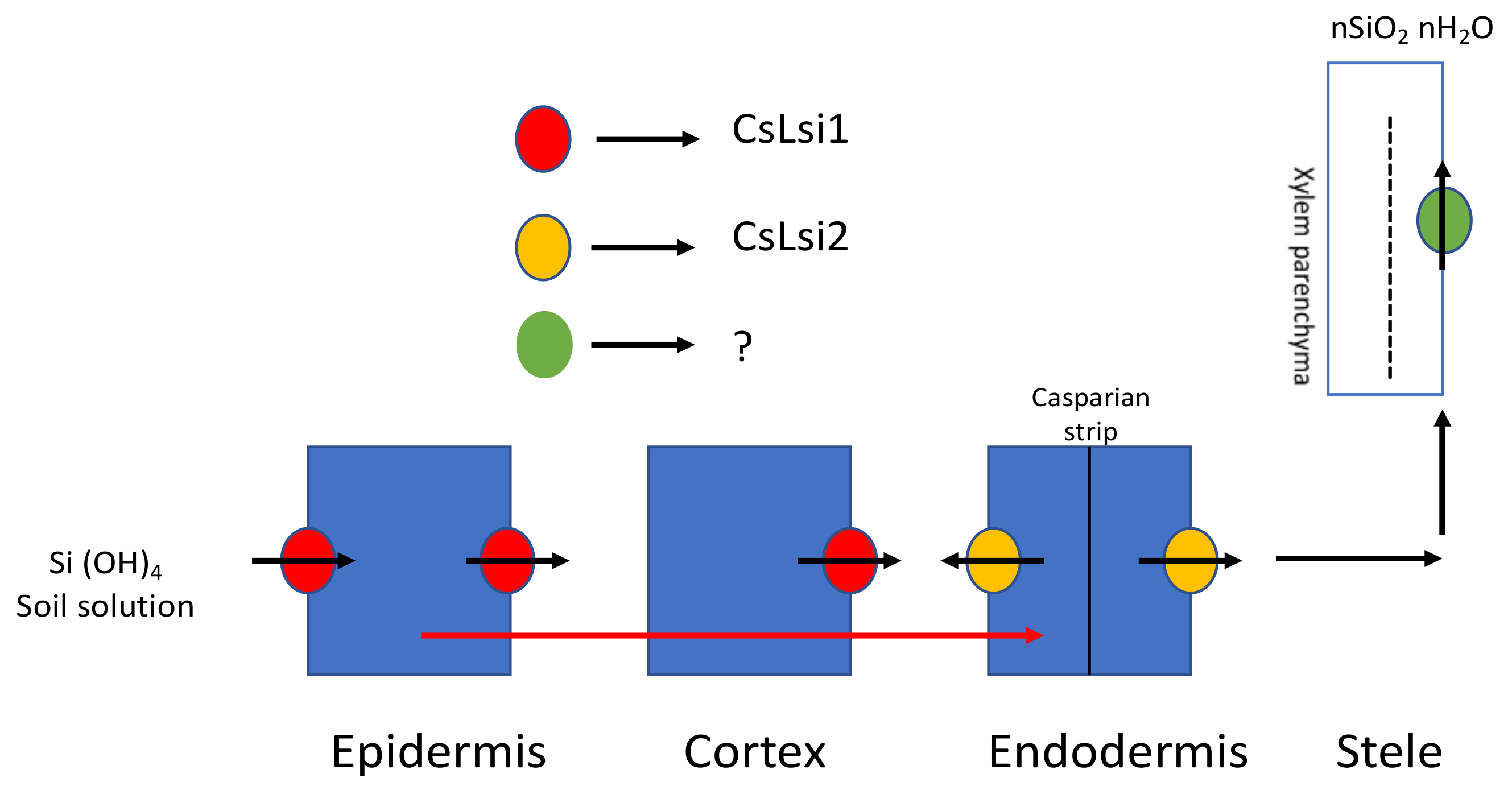
Cucumber (Cucumis sativus), a commonly grown cucurbitaceous vegetable, accumulates more Si compared with other eudicots. In a study, it was concluded that more than half of the Si taken up by cucumber roots reaches the shoots [18], while, in another study, it was shown that Si uptake is a concentration-independent, active process that follows the Michaelis curve [16]. CsLsi1, the Si cucumber influx transporter gene with localization in the distal side of the endodermis and the cortical cells in the root tips, was isolated and characterized in root hairs [37]. CsLsi1 showed 55.7% and 90.6% homology with rice and pumpkin Lsi1 transporter genes, respectively. CsLsi2, an efflux Si transporter, has also been identified and characterized recently; it is located in the plasma membrane, and its uptake is an energy-dependent process [38]. Transient expression in a Xenopus laevis oocyte showed efflux activity for transport of Si. Synchronization of both CsLsi1 and CsLsi2 mediates Si uptake and transport across the cell (Figure 3).
GmNIP2-1 and GmNIP2-2, two Si influx transporter genes have been identified and characterized in soybean in a comparative study of a Si-accumulating (rice) and a non-accumulating (Arabidopsis) species [39]. The permeability of these transporters towards Si uptake was tested using Xenopus oocyte bioassay. Potato, the third most important crop plant after wheat and rice, shows a potential response to the application of Si fertilizers. A Si transporter homologous with that of pumpkin led to the discovery of a putative Si transporter, StLsi1, in potato [40]. StLsi1 is localized in the roots and leaves; however, its expression rates were much higher in the roots compared with the leaves.
3.2. Cryptogams
Equisetum arvense (horsetail) is the highest Si accumulator; however, it has always been neglected, as early efforts by scientists to identify Si transporters proved unsuccessful. Based upon the phylogenetic analysis of its sequence, it was revealed that horsetail belongs to a new multigene family of NIPs, which is a different group compared with that of the already identified plant Si transporters [41]. This newly identified protein group contains a STAR pore in contrast with the previously identified GSGR pore in other known Si transporters. These plants are more inclined, in terms of homology, towards the NIP II group than the NIP III group. The NIP II group comprises of transporters for other small, uncharged molecules. In order to study the transmembrane flux, heterologous protein expressions were carried out in a Xenopus oocyte, and it was found that aquaporins from three different horsetail subgroups, EaNIP3;1, EaNIP3;3, and EaNIP3;4, with STAR pores were very efficient Si transporters. Therefore, it was concluded that EaNIP3;1 and EaNIP3;4 were the most efficient, known plant Si transporters, exhibiting even higher activity than rice transporters.
EaLsi2-1 and EaLsi2-2, two putative Si efflux transporter genes, have also been identified in horsetail, showing less than 50% homology to rice gene transporters [42]. In these plants, structural differences with rice Lsi2 genes can also been attributed to the presence of two introns instead of one. Both these transporter genes are expressed in the roots and shoots, though level of expression were much higher in the shoots than the roots, suggesting that both contribute to Si transport.
4. Conclusions and Future Prospects
Si provides a number of benefits for plants; although it is not regarded as an essential element, it plays an important role against several biotic and abiotic factors. Unfortunately, the majority of plants do not benefit from these effects, as they are either deprived of an efficient Si transport system or the molecular mechanism underlying the uptake and transport of Si in these plants is poorly understood. It is believed that various transporters are involved in the Si uptake mechanism, but only a few could be identified so far. Table 2 shows the presence and absence of Si transporters in different plant families.
To study the Si uptake and transport mechanism via transporters into the plants at the cellular level, the extraction and characterization of the amino-acid sequence in relation to functional proteins needs to be explored. Moreover, understanding the polar localization of these transporters is another crucial step to identify the directional flow of Si into and outside the cells. The resurgence of interest in studying the Si uptake mechanism through the identification of putative genes and controlled gene expressions will also fast-track this research; as with molecular approaches, specific Si transporter genes can be addressed, whereas others involved in general regulatory functions can be eliminated.
In the current review, we tried to highlight the transport mechanism involved in Si uptake in monocots, eudicots, and few cryptogams. However, in investigating the benefits of Si, it is of utmost importance to study more closely the underlying molecular mechanism of Si uptake in other plant species. Owing to recent developments in this field of research, it is quite clear that many other mechanisms can be explored and understood in greater depth.
Author Contributions
This is a review article and both the authors contributed their part.
Funding
This research received no external funding.
Acknowledgments
No special acknowledgment.
Conflicts of Interest
No conflict of interest.
References
- Wedepohl, K.H. The composition of the continental crust. Geochim. Cosmochim. Acta 1995, 59, 1217–1232. [Google Scholar] [CrossRef]
- Pereira, C.E.D.B.; Felcman, J. Correlation between five minerals and the healing effect of Brazilian medicinal plants. Biol. Trace Elem. Res. 1998, 65, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Savant, N.K.; Korndörfer, G.H.; Datnoff, L.E.; Snyder, G.H. Si nutrition and sugarcane production: A review. J. Plant Nutr. 1999, 22, 1853–1903. [Google Scholar] [CrossRef]
- Ma, J.F. Role of Si in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Zhu, Y.; Gong, H. Beneficial effects of Si on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F. Mechanisms of Si-mediated alleviation of heavy metal toxicity in plants: A review. Ecotox. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.A.; Dietz, K.J. Silicon as versatile player in plant and human biology-Overlooked and poorly understood. Front. Plant Sci. 2015, 6, 994. [Google Scholar] [CrossRef]
- Deshmukh, R.; Belanger, R.R. Molecular evolution of aquaporins and Si influx in plants. The functional role of Si in plant Biology. Funct. Ecol. 2016, 30, 1277–1285. [Google Scholar] [CrossRef]
- Mitani, N.; Ma, J.F. Uptake system of Si in different plant species. J. Exp. Bot. 2005, 56, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Takahashi, E. Si-Accumulating Plants in the Plant Kingdom. In Soil, Fertilizer, and Plant Si Research in Japan; Elsevier Science: Amsterdam, The Netherlands, 2002; pp. 63–71. [Google Scholar]
- Hodson, M.; White, P.; Mead, A.; Broadley, M. Phylogenetic variation in the Si composition of plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef]
- Cornelis, J.T.; Delvaux, B.; Georg, R.B.; Lucas, Y.; Ranger, J.; Opfergelt, S. Tracing the origin of dissolved Si transferred from various soil plant systems towards rivers: A review. Biogeosciences 2011, 8, 89–112. [Google Scholar] [CrossRef]
- Epstein, E. The anomaly of Si in plant biology. Proc. Natl. Acad. Sci. USA 1994, 91, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Miyakeb, Y.; Takahashi, E. Si as a Beneficial Element for Crop Plants. In Si in Agriculture; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; Volume 8, pp. 17–39. [Google Scholar]
- Takahashi, E.; Ma, J.F.; Miyake, Y. The possibility of Si as an essential element for higher plants. Comments Agric. Food Chem. 1990, 2, 99–122. [Google Scholar]
- Liang, Y.; Si, J.; Römheld, V. Si uptake and transport is an active process in Cucumis sativus. New Phytol. 2005, 167, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Marron, A.O.; Sarah, R.; Wheeler, G.L.; Goldstein, R.E.; King, N.; Not, F.; Vargas, C.D.; Richter, D.J. The evolution of Si transport in eukaryotes. Mol. Biol. Evol. 2016, 33, 3226–3248. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, M.; Nikolic, N.; Liang, Y.C.; Kirkby, E.A.; Römheld, V. Germanium-68 as an adequate tracer for Si transport in plants. Characterization of Si uptake in different crop species. Plant Physiol. 2007, 143, 495–503. [Google Scholar] [CrossRef]
- Raven, J.A. Si Transport at the Cell and Tissue Level. In Si in Agriculture; Studies in Plant Science; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; Volume 8, pp. 41–55. [Google Scholar]
- Reynolds, O.L.; Keeping, M.G.; Meyer, J.H. Si-augmented resistance of plants to herbivorous insects: A review. Ann. Appl. Biol. 2009, 155, 171–186. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Si uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef]
- Meunier, J.D.; Barboni, D.; Anwarul Haq, M.; Levard, C.; Chaurand, P.; Vidal, V.; Grauby, O.; Huc, R.; Laffont-Schwob, I.; Rabier, J.; et al. Effect of phytoliths for mitigating water stress in durum wheat. New Phytol. 2017, 215, 229–239. [Google Scholar] [CrossRef]
- Ma, J.F. Si Transporters in Higher Plants. In MIPs and Their Role in the Exchange of Metalloids. Advances in Experimental Medicine and Biology; Jahn, T.P., Bienert, G.P., Eds.; Springer: New York, NY, USA, 2010; Volume 679, pp. 99–109. [Google Scholar]
- Mitani, N.; Yamaji, N.; Ago, Y.; Iwasaki, K.; Ma, J.F. Isolation and functional characterization of an influx Si transporter in two pumpkin cultivars contrasting in Si accumulation. Plant J. 2011, 66, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Montpetit, J.; Vivancos, J.; Mitani-Ueno, N.; Yamaji, N.; Rémus-Borel, W.; Belzile, F.; Ma, J.F.; Bélanger, R.R. Cloning, functional characterization and heterologous expression of TaLsi1, a wheat Si transporter gene. Plant Mol. Biol. 2012, 79, 35–46. [Google Scholar] [CrossRef]
- Ma, J.F.; Mitani, N.; Nagao, S.; Konishi, S.; Tamai, K.; Iwashita, T.; Yano, M. Characterization of Si uptake system and molecular mapping of Si transporter gene in rice. Plant Physiol. 2004, 136, 3284–3289. [Google Scholar] [CrossRef]
- Gomes, D.; Agasse, A.; Thiebaud, P.; Delrot, S.; Geros, H.; Chaumont, F. Aquaporins are multifunctional water and solute transporters highly divergent in living organisms. Biochim. Biophys. Acta Biomemb. 2009, 1788, 1213–1228. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Tamai, K.; Ichii, M.; Wu, G.F. A rice mutant defective in Si uptake. Plant Physiol. 2002, 130, 2111–2117. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of Si in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef]
- Yamaji, N.; Ma, J.F. Further characterization of a rice Si efflux transporter, Lsi2. Soil Sci. Plant Nutr. 2011, 57, 259–264. [Google Scholar] [CrossRef]
- Yamaji, N.; Mitani, N.; Ma, J.F. A transporter regulating Si distribution in rice shoots. Plant Cell 2008, 20, 1381–1389. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N.; Mitani-Ueno, N. Transport of Si from roots to panicles in plants. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2011, 87, 377. [Google Scholar] [CrossRef]
- Remus-Borel, W.; Menzies, J.; Belanger, R. Si induces antifungal compounds in powdery mildew-infected wheat. Physiol. Mol Plant Pathol. 2005, 66, 108–115. [Google Scholar] [CrossRef]
- Chiba, Y.; Mitani, N.; Yamaji, N.; Ma, J.F. HvLsi1 is a Si influx transporter in barley. Plant J. 2009, 57, 810–818. [Google Scholar] [CrossRef]
- Mitani, N.; Yamaji, N.; Ma, J.F. Identification of maize Si influx transporters. Plant Cell Physiol. 2009, 50, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Mitani-Ueno, N.; Yamaji, N.; Ma, J.F. Si efflux transporters isolated from two pumpkin cultivars contrasting in Si uptake. Plant Signal. Behav. 2011, 6, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Guo, J.; Duan, Y.; Zhang, T.; Huo, H.; Gong, H. Isolation and functional characterization of CsLsi1, a Si transporter gene in Cucumis sativus. Physiol. Plant 2017, 159, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Duan, Y.; Qi, X.; Zhang, L.; Huo, H.; Gong, H. Isolation and characterization of CsLsi2, a cucumber Si efflux transport gene. Ann. Bot. 2018, 122. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.K.; Vivabcos, J.; Guerin, V.; Sonah, H.; Labbe, C.; Belzile, F.; Belanger, R.R. Identification and functional characterization of Si transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef]
- Vulavala, V.K.R.; Elbaum, R.; Yermiyahu, U.; Fogelman, E.; Kumar, A.; Ginzberg, I. Si fertilization of potato: Expression of putative transporters and tuber skin quality. Planta 2016, 243, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, C.; Remus-Borel, W.; Vivancos, J.; Labbe, C.; Belzile, F.; Belanger, R.R. Discovery of a multigene family of aquaporin Si transporters in the primitive plants Equisetum arvense. Plant J. 2012, 72, 320–330. [Google Scholar] [CrossRef]
- Vivancos, J.; Deshmukh, R.; Gregoire, C.; Remus-Borel, W.; Belzile, F.; Belanger, R.R. Identification and characterization of Si efflux transporters in horsetail (Equisteum arvense). J. Plant Physiol. 2016, 200, 82–89. [Google Scholar] [CrossRef]
- Trembath-Reichert, E.; Wilson, J.P.; McGlynn, S.E.; Fischer, W.W. Four hundred million years of silica bio mineralization in land plants. Proc. Natl. Acad. Sci. USA 2015, 112, 5449–5454. [Google Scholar] [CrossRef]
Figure 1.
Flow of Si across the plant using Si transporters in rice. Lsi1, the influx transporter gene localized in the plasma membrane, is accountable for Si uptake into the exodermis of the cell; Lsi2, an efflux transporter, moves Si into the apoplast across the aerenchyma; Lsi6 further takes it up the aerial parts of the plant. Modified with reference [32].
Figure 1.
Flow of Si across the plant using Si transporters in rice. Lsi1, the influx transporter gene localized in the plasma membrane, is accountable for Si uptake into the exodermis of the cell; Lsi2, an efflux transporter, moves Si into the apoplast across the aerenchyma; Lsi6 further takes it up the aerial parts of the plant. Modified with reference [32].

Figure 2.
Si transporter-mediated Si transport in maize and barley. In maize and barley, Si is absorbed from the external solution using ZmLsi1/HvLsi1 and transported further into the root cells from which, through the symplastic pathway, it is passed into the endodermis and further to the stele. ZmLsi6 mainly functions as a Si transporter for xylem unloading. Modified with reference [32].
Figure 2.
Si transporter-mediated Si transport in maize and barley. In maize and barley, Si is absorbed from the external solution using ZmLsi1/HvLsi1 and transported further into the root cells from which, through the symplastic pathway, it is passed into the endodermis and further to the stele. ZmLsi6 mainly functions as a Si transporter for xylem unloading. Modified with reference [32].

Figure 3.
Si uptake and transport in Cucumis sativus. CsLsi1, the influx transporter, takes up Si from an external solution, and CsLsi2, the efflux transporter, further transports it to the endodermis by symplastic pathway. However, the transporter for xylem unloading has not yet been identified. (?—No information is available in literature so far). Modified with reference [32].
Figure 3.
Si uptake and transport in Cucumis sativus. CsLsi1, the influx transporter, takes up Si from an external solution, and CsLsi2, the efflux transporter, further transports it to the endodermis by symplastic pathway. However, the transporter for xylem unloading has not yet been identified. (?—No information is available in literature so far). Modified with reference [32].


{kind=link}
{kind=link}
{kind=link}
Table 1.
Different classes of Si accumulators, including high, low, and intermediate accumulator plant species.
Table 1.
Different classes of Si accumulators, including high, low, and intermediate accumulator plant species.
| Group | Species | Si Accumulation |
|---|---|---|
| Angiosperms | Acoraceae Acorus calamus | - |
| Asteraceae Anaphalis margarita | - | |
| Helianthus annuus | + | |
| Helianthus maximilianii, | + | |
| Helianthus atrorubens | + | |
| Inula helenium, Inula viscosa | ± | |
| Lactuca serriola | ± | |
| Cyperaceae (Carex cinica) | + | |
| Poaceae Agrostis spp. | + | |
| Andropogon scoparius | + | |
| Arundinaria gigantean | + | |
| Bouteloua hirsuta | + | |
| Brachypodium sylvaticum | + | |
| Chasmanthium latifolium | + | |
| Ctenium aromaticum | + | |
| Echinochloa colona | ± | |
| Elymus molli | ± | |
| Oryza sativa | + | |
| Zea mays | ± | |
| Cucurbitaceae Benincasa hispida | ± | |
| Citrullus lanatus | ± | |
| Ecballium elaterium | ± | |
| Equisetophyta | Equisetaceae Equisetum arvense | + |
| Equisetum hyemale | + |
Table 2.
The presence of Si transporters in various plant families.
| Plant Species | Si Transporters |
|---|---|
| Angiosperms (monocots and a few eudicots) | + |
| Equisetopsida | + |
| Gymnosperms | - |
| Lycophytes | - |
| Bryophytes (Liverworts, Mosses, Hornworts) | - |
| Gnetophytes | - |
Modified with reference from Trembath [43].
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kaur, H.; Greger, M. A Review on Si Uptake and Transport System. Plants 2019, 8, 81. https://doi.org/10.3390/plants8040081
AMA Style
Kaur H, Greger M. A Review on Si Uptake and Transport System. Plants. 2019; 8(4):81. https://doi.org/10.3390/plants8040081
Chicago/Turabian StyleKaur, Harmanjit, and Maria Greger. 2019. "A Review on Si Uptake and Transport System" Plants 8, no. 4: 81. https://doi.org/10.3390/plants8040081
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.