

Plant Growth-Promoting Rhizobacteria as Tools to Improve the Growth of Kohlrabi (Brassica oleracea var. gongylodes) Plants in an Aquaponics System
 , ,
, ,  and
and
Abstract
:1. Introduction
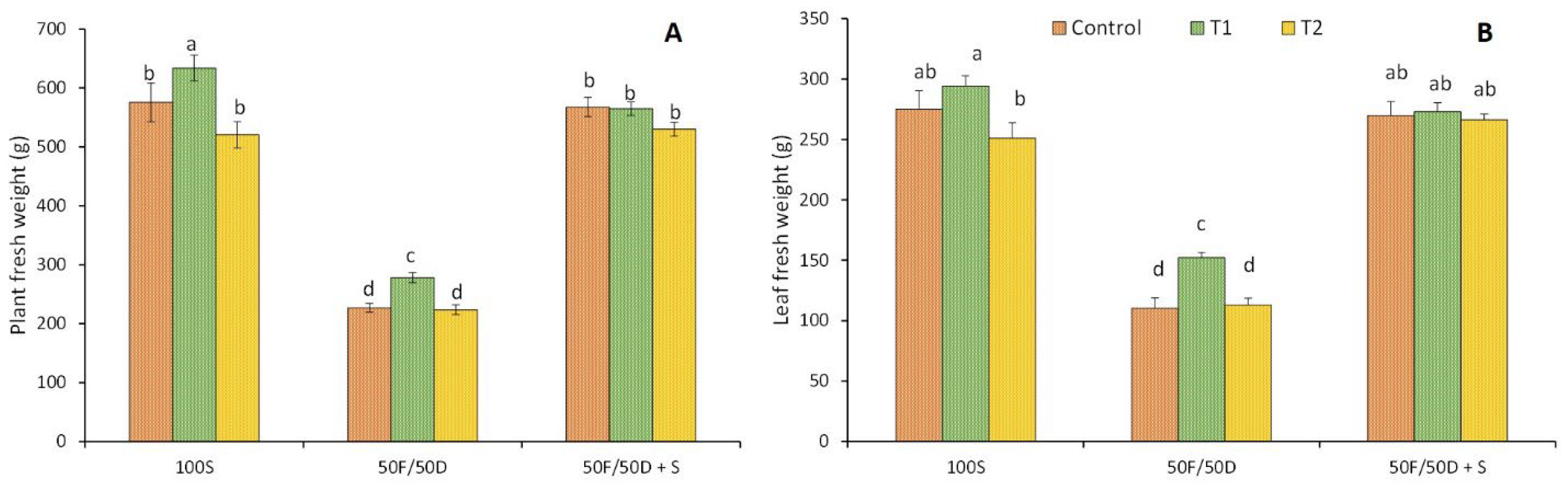
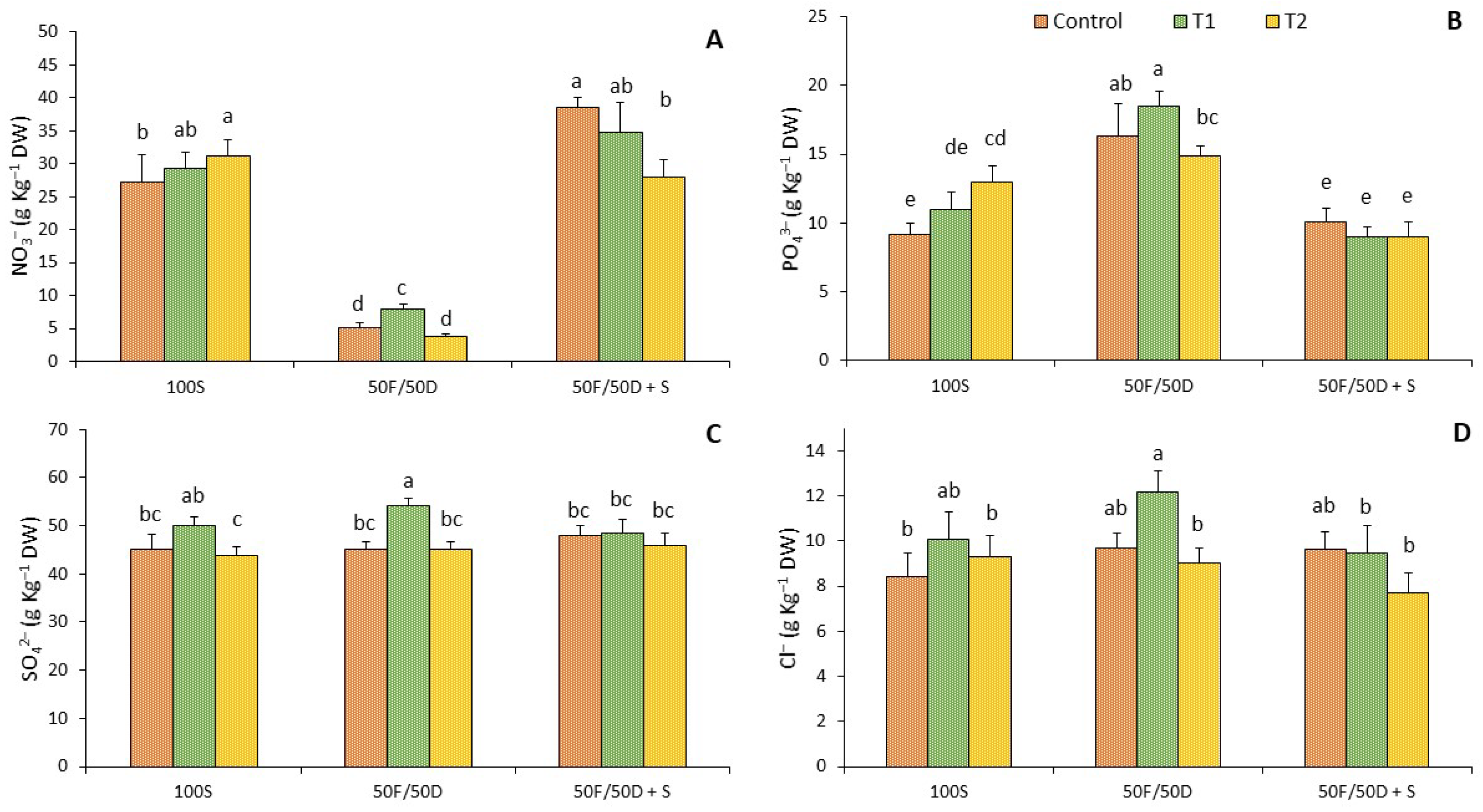
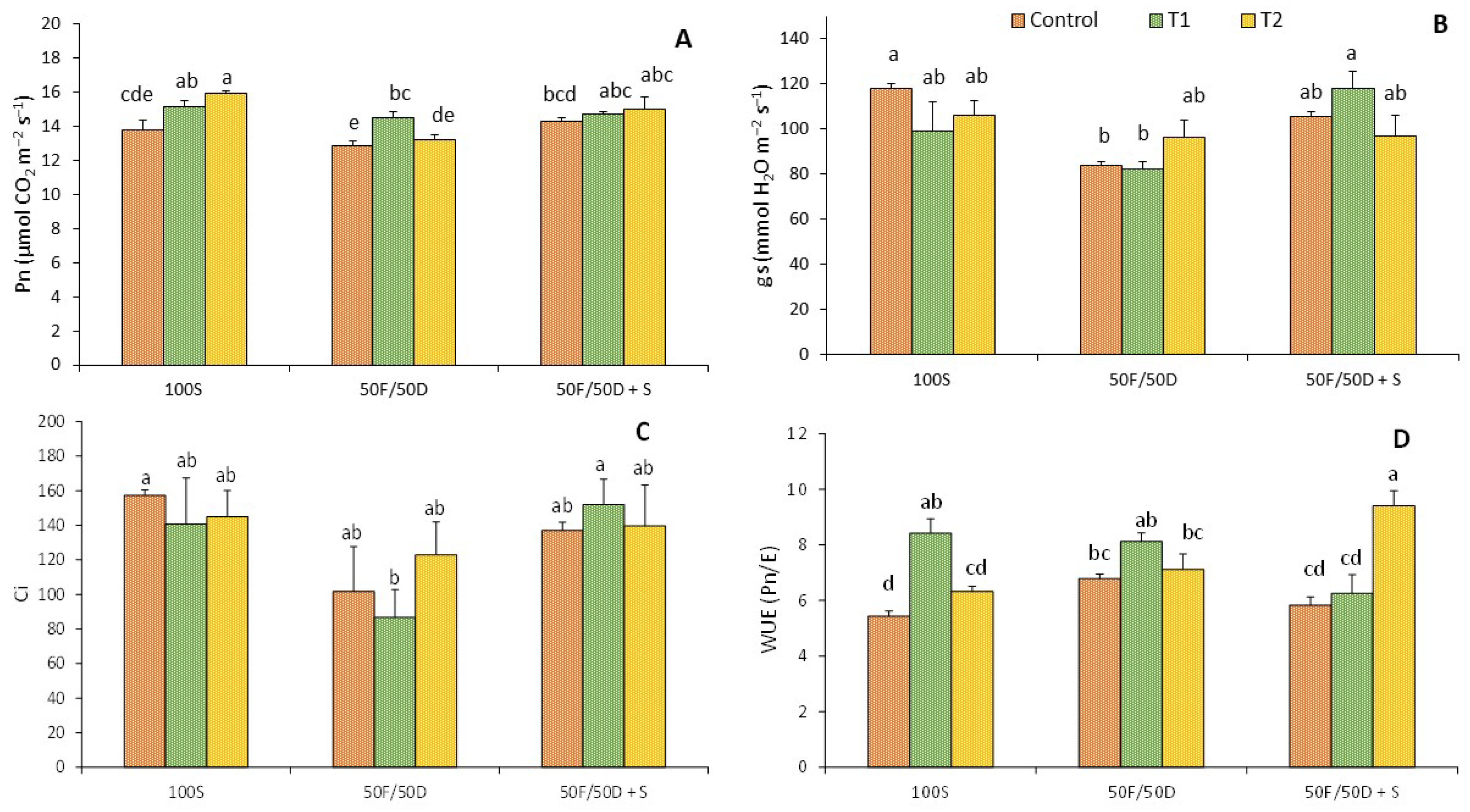
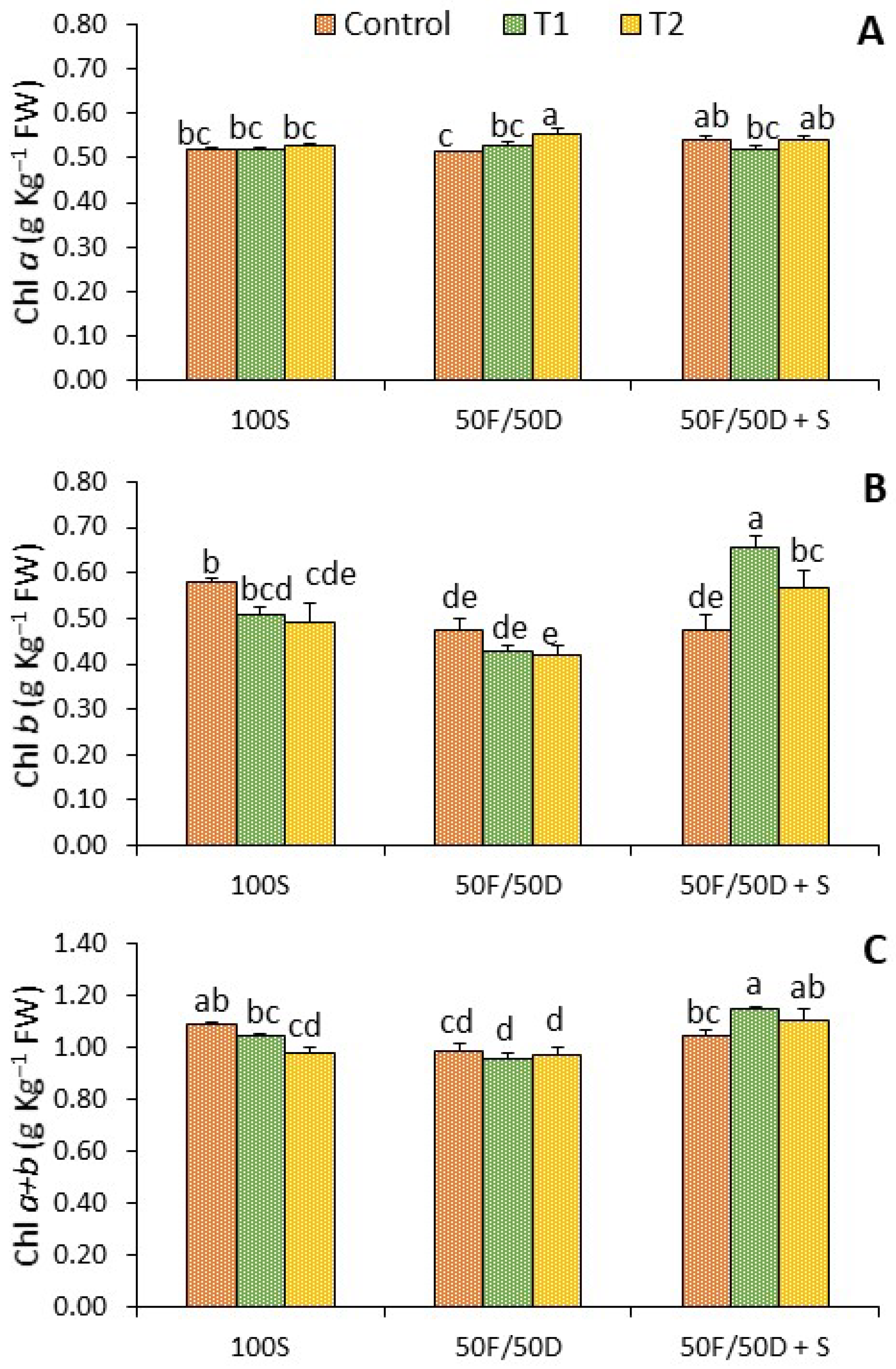
2. Results
2.1. Plant Material and Growth Conditions
2.2. Ion Determination
2.3. Gas Exchange
2.4. Chlorophylls
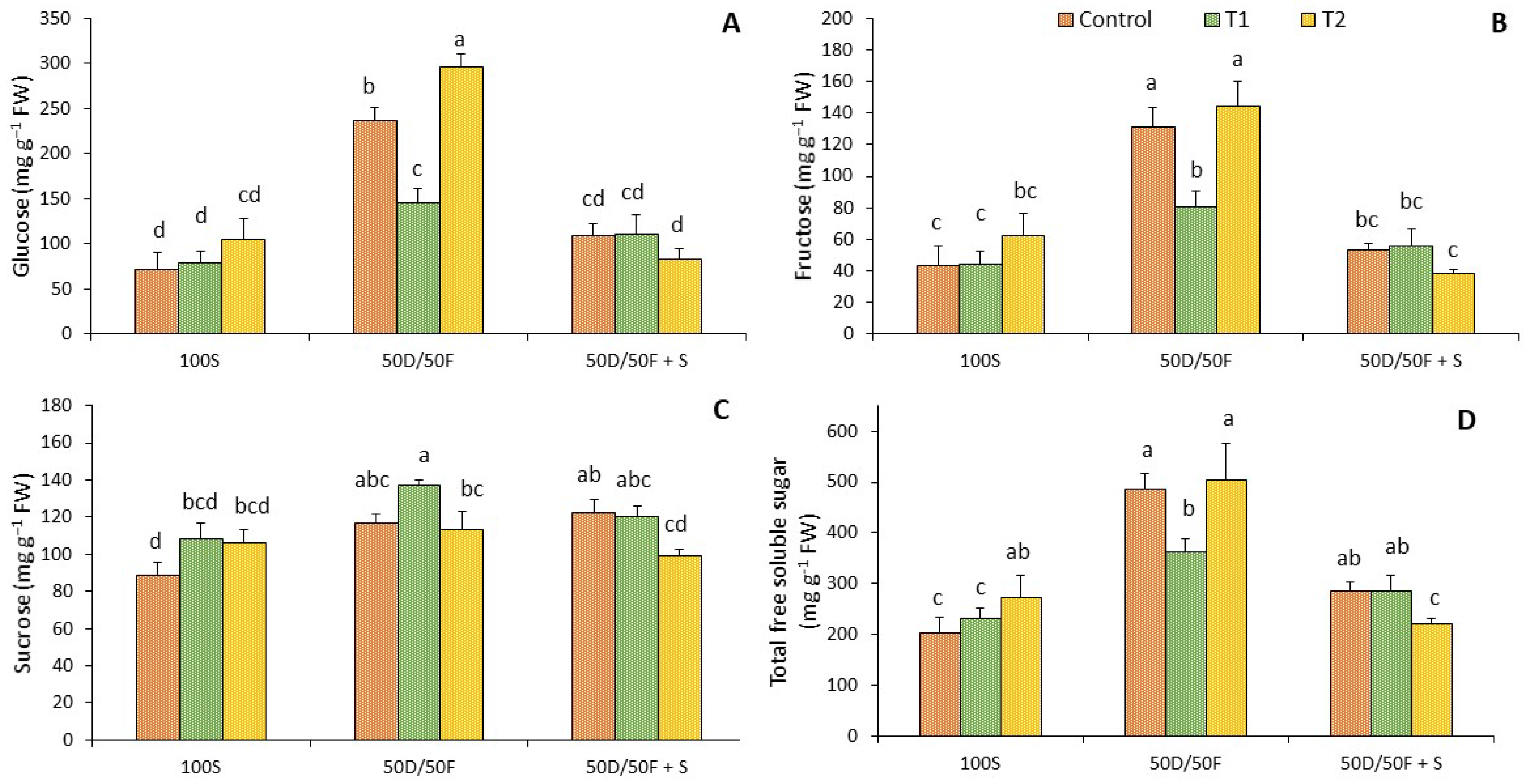
2.5. Total Soluble Sugars
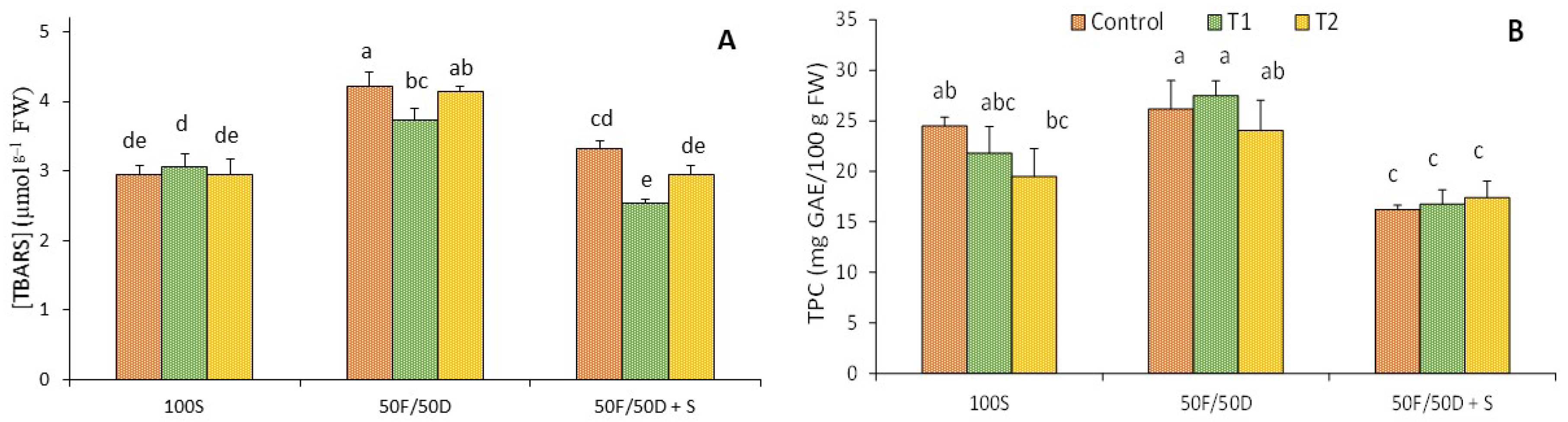
2.6. Lipid Peroxidation and Total Phenolic Compounds
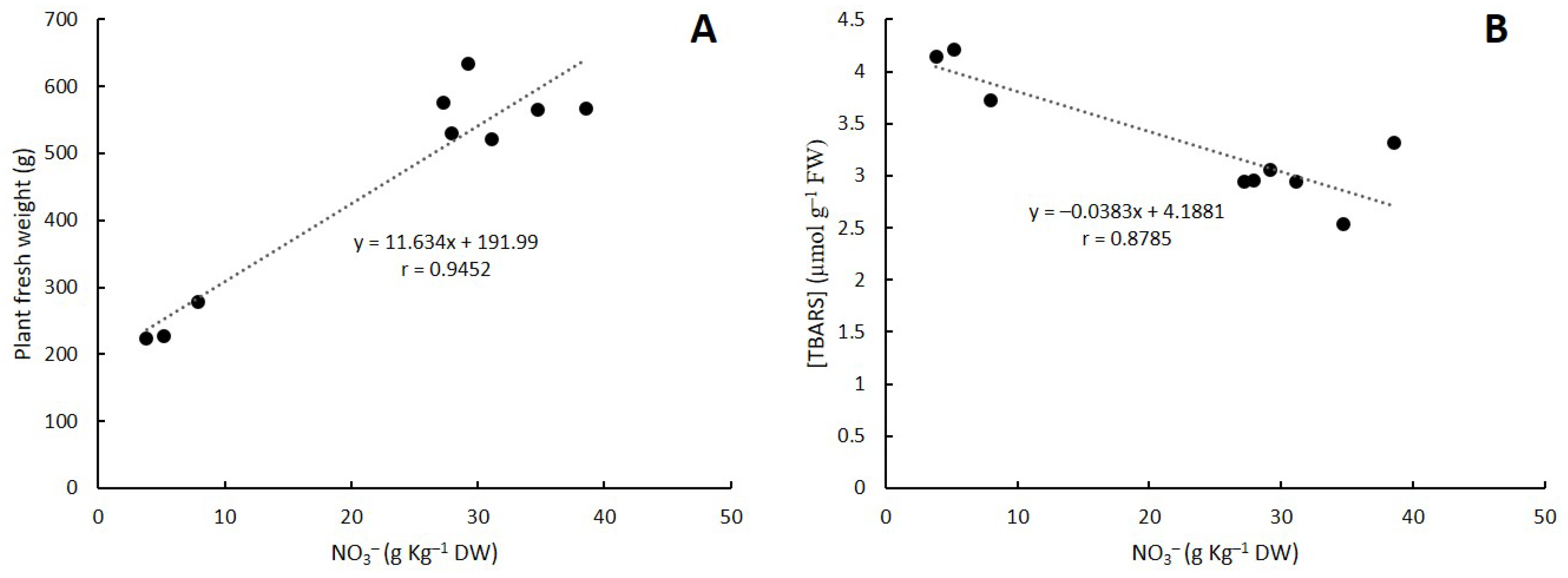
3. Discussion
4. Materials and Methods
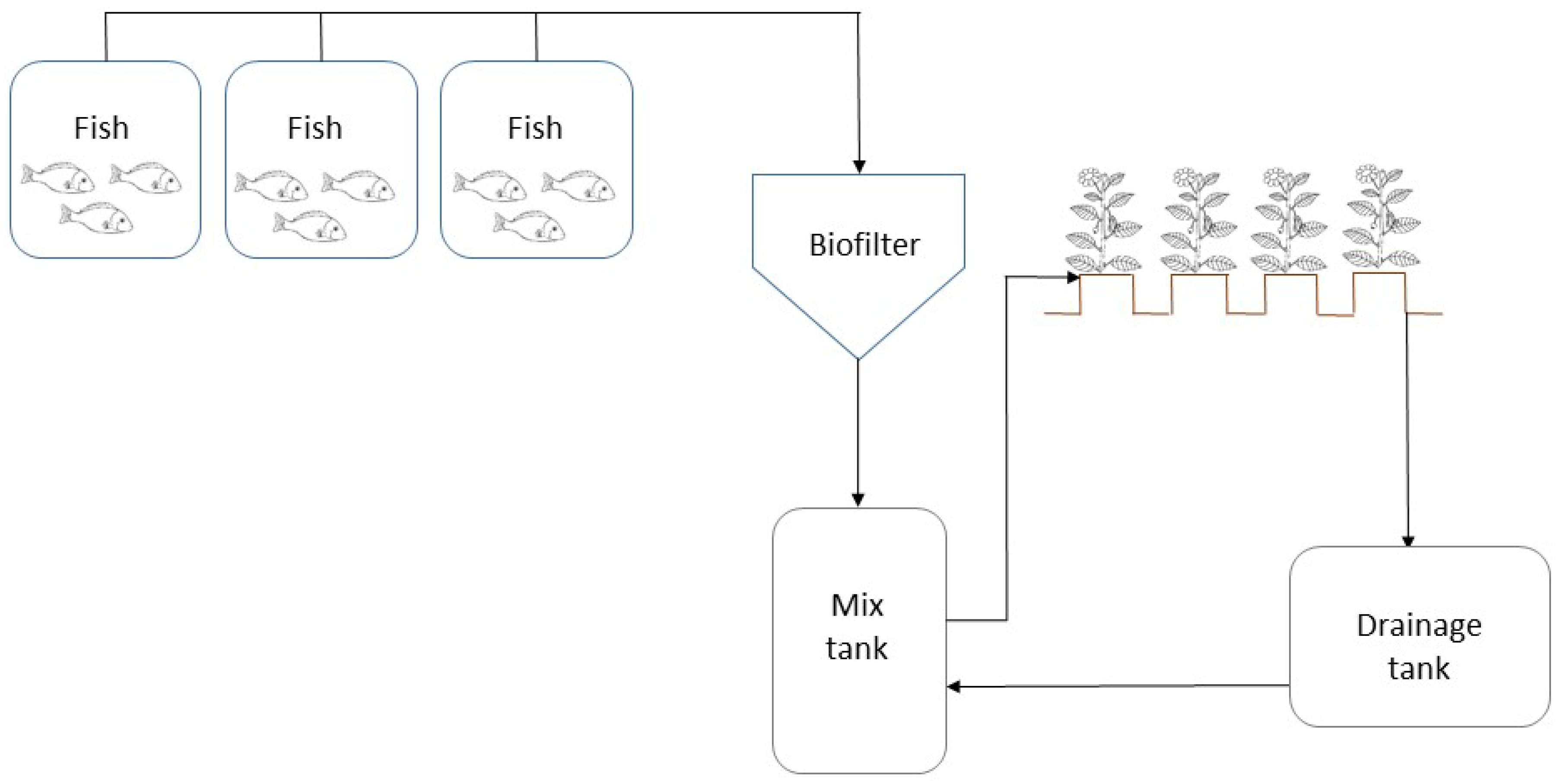
4.1. Plant Material and Growth Conditions
4.2. Ion Determination
4.3. Gas Exchange
4.4. Chlorophylls
4.5. Total Soluble Sugars
4.6. Lipid Peroxidation and Total Phenolic Compounds
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Nocerino, S.; Rouphael, Y.; Colla, G.; El-Nakhel, C.; Mori, M. Nitrogen use and uptake efficiency and crop performance of baby spinach (Spinacia oleracea L.) and Lamb’s Lettuce (Valerianella locusta L.) grown under variable sub-optimal N regimes combined with plant-based biostimulant application. Agronomy 2020, 10, 278. [Google Scholar] [CrossRef]
- Navarro-León, E.; Javier López-Moreno, F.; Borda, E.; Marín, C.; Sierras, N.; Blasco, B.; Ruiz, J.M. Effect of l-amino acid-based biostimulants on nitrogen use efficiency (NUE) in lettuce plants. Wiley Online Libr. 2022, 102, 7098–7106. [Google Scholar] [CrossRef]
- Sutton, M.; Howard, C.; Erisman, J.; Billen, G. The European Nitrogen Assessment; Cambridge University Press: Cambridge, UK, 2011; ISBN 9780511976988. [Google Scholar]
- Waqas, M.; Hawkesford, M.; Geilfus, C. Feeding the world sustainably: Efficient nitrogen use. Trends Plant Sci. 2023, 28, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Goddek, S.; Alyssa, J.; Kotzen, B.; Burnell, G.M. Aquaponics Food Production Systems Combined Aquaculture and Hydroponic Production Technologies for the Future. In Aquaponics Food Production Systems; Springer: Cham, Switzerland, 2019; pp. 353–378. [Google Scholar] [CrossRef]
- Piñero, M.C.; Otálora, G.; Collado-González, J.; López-Marín, J.; del Amor, F.M. Effects triggered by foliar selenium application on growth, enzyme activities, mineral nutrients and carbohydrates in lettuce under an aquaculture system. Plant Physiol. Biochem. 2022, 180, 1–8. [Google Scholar] [CrossRef]
- Zhao, C.F.; Zhou, L.H.; Zhang, Y.D.; Zhu, Z.; Chen, T.; Zhao, Q.Y.; Yao, S.; Yu, X.; Wang, C.L. QTL mapping for seedling traits associated with low-nitrogen tolerance using a set of advanced backcross introgression lines of rice. Plant Breed. 2014, 133, 189–195. [Google Scholar] [CrossRef]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.L.; Zeng, X.-P.; Liu, Y.; Li, Y.R. Fate of nitrogen in agriculture and environment: Agronomic, eco-physiological and molecular approaches to improve nitrogen use efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ.-Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef]
- Cassán, F.; Coniglio, A.; López, G.; Molina, R.; Nievas, S.; Le Noir De Carlan, C.; Donadio, F.; Torres, D.; Rosas, S.; Olivera Pedrosa, F.; et al. Everything you must know about Azospirillum and its impact on agriculture and beyond. Biol. Fertil. Soils 2020, 56, 461–479. [Google Scholar] [CrossRef]
- Sharma, S.D.; Kumar, P.; Raj, H.; Bhardwaj, S.K. Isolation of arbuscular mycorrhizal fungi and Azotobacter chroococcum from local litchi orchards and evaluation of their activity in the air-layers system. Sci. Hortic. 2009, 123, 117–123. [Google Scholar] [CrossRef]
- Muthukumar, T.; Udaiyan, K. Growth of nursery-grown bamboo inoculated with arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria in two tropical soil types with and without fertilizer application. New For. 2006, 31, 469–485. [Google Scholar] [CrossRef]
- Soltan, H.A.H.; Osman, S.A.M.; Tantawy, I.A.A. The Role of New Transformants Phosphate Bio-Stimulates (PBS) Bacteria Inoculates on Growth, Yield and Quality of Kohlrabi Plants. J. Plant Prod. 2021, 12, 541–551. [Google Scholar] [CrossRef]
- Cechin, I.; Valquilha, É.M. Nitrogen effect on gas exchange characteristics, dry matter production and nitrate accumulation of Amaranthus cruentus L. Rev. Bras. Bot. 2019, 42, 373–381. [Google Scholar] [CrossRef]
- Hu, W.; Lu, Z.; Meng, F.; Li, X.; Cong, R.; Ren, T.; Sharkey, T.D.; Lu, J. The reduction in leaf area precedes that in photosynthesis under potassium deficiency: The importance of leaf anatomy. New Phytol. 2020, 227, 1749–1763. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Long, S.P.; Zhu, X.-G.; Naidu, S.L.; Ort, D.R. Can improvement in photosynthesis increase crop yields? Wiley Online Libr. 2006, 29, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.W. Photosynthesis, productivity and environment. J. Exp. Bot. 1995, 46, 1449–1461. [Google Scholar] [CrossRef]
- Efthimiadou, A.; Bilalis, D.; Karkanis, A.; Froud-Williams, B. Combined organic/inorganic fertilization enhance soil quality and increased yield, photosynthesis and sustainability of sweet maize crop. Aust. J. Crop Sci. 2010, 4, 722–729. Available online: https://search.informit.org/doi/10.3316/informit.859484046375351 (accessed on 21 February 2023).
- Zhang, J.; Xie, J.; Gan, Y.; Coulter, J.A.; Dawuda, M.M.; Yu, J.; Lv, J.; Li, J.; Zhang, X.; Tang, C.; et al. Promoting pepper (Capsicum annuum) photosynthesis via chloroplast ultrastructure and enzyme activities by optimising the ammonium to nitrate ratio. Funct. Plant Biol. 2020, 47, 303. [Google Scholar] [CrossRef]
- Liu, X.; Herbert, S.J.; Jin, J.; Zhang, Q.; Wang, G. Responses of photosynthetic rates and yield/quality of main crops to irrigation and manure application in the black soil area of Northeast China. Plant Soil 2004, 261, 55–60. [Google Scholar] [CrossRef]
- Jiang, H.M.; Yang, J.C.; Zhang, J.F. Effects of external phosphorus on the cell ultrastructure and the chlorophyll content of maize under cadmium and zinc stress. Environ. Pollut. 2007, 147, 750–756. [Google Scholar] [CrossRef]
- Zhao, D.; Oosterhuis, D.M.; Bednarz, C.W. Influence of potassium deficiency on photosynthesis, chlorophyll content, and chloroplast ultrastructure of cotton plants. Photosynthetica 2001, 39, 103–109. [Google Scholar] [CrossRef]
- Collado-González, J.; Piñero, M.C.; Otálora, G.; López-Marín, J.; Del Amor, F.M. Effects of different nitrogen forms and exogenous application of putrescine on heat stress of cauliflower: Photosynthetic gas exchange, mineral concentration and lipid peroxidation. Plants 2021, 10, 152. [Google Scholar] [CrossRef] [PubMed]
- Ayuso-Calles, M.; Flores-Félix, J.D.; Rivas, R. Overview of the Role of Rhizobacteria in Plant Salt Stress Tolerance. Agronomy 2021, 11, 1759. [Google Scholar] [CrossRef]
- Boleta, E.H.M.; Shintate Galindo, F.; Jalal, A.; Santini, J.M.K.; Rodrigues, W.L.; de Lima, B.H.; Arf, O.; da Silva, M.R.; Buzetti, S.; Teixeira Filho, M.C.M. Inoculation With Growth-Promoting Bacteria Azospirillum brasilense and Its Effects on Productivity and Nutritional Accumulation of Wheat Cultivars. Front. Sustain. Food Syst. 2020, 4, 607262. [Google Scholar] [CrossRef]
- Muhammad, I.; Yang, L.; Ahmad, S.; Farooq, S.; Al-Ghamdi, A.A.; Khan, A.; Zeeshan, M.; Elshikh, M.S.; Mehmood Abbasi, A.; Zhou, X.-B. Nitrogen fertilizer modulates plant growth, chlorophyll pigments and enzymatic activities under different irrigation regimes. Agronomy 2022, 12, 845. [Google Scholar] [CrossRef]
- Lara, T.S.; de Lima Lessa, J.H.; de Souza, K.R.D.; Corguinha, A.P.B.; Martins, F.A.D.; Lopes, G.; Guilherme, L.R.G. Selenium biofortification of wheat grain via foliar application and its effect on plant metabolism. J. Food Compos. Anal. 2019, 81, 10–18. [Google Scholar] [CrossRef]
- Bashan, Y.; De-Bashan, L.E. How the plant growth-promoting bacterium azospirillum promotes plant growth-a critical assessment. Adv. Agron. 2010, 108, 77–136. [Google Scholar] [CrossRef]
- Sun, N.; Huang, L.; Zhao, H.; Zhang, N.; Lin, X.; Sun, C. Beneficial Bacterium Azospirillum brasilense Induces Morphological, Physiological and Molecular Adaptation to Phosphorus Deficiency in Arabidopsis. Plant Cell Physiol. 2022, 63, 1273–1284. [Google Scholar] [CrossRef]
- Pinero, M.C.; Perez-Jimenez, M.; Lopez-Marin, J.; del Amor, F.M. Changes in the salinity tolerance of sweet pepper plants as affected by nitrogen form and high CO2 concentration. J. Plant Physiol. 2016, 200, 18–27. [Google Scholar] [CrossRef]
- Nagata, M.; Yamashita, I. Simple Method for Simultaneous Determination of Chlorophyll and Carotenoids in Tomato Fruit. J. Jpn. Soc. Food Sci. Technol. Shokuhin Kagaku Kogaku Kaishi 1992, 39, 925–928. [Google Scholar]
- Balibrea, M.E.; Cuartero, J.; Bolarín, M.C.; Pérez-Alfocea, F. Sucrolytic activities during fruit development of Lycopersicon genotypes differing in tolerance to salinity. Physiol. Plant. 2003, 118, 38–46. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts: I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Balestrasse, K.B.; Gallego, S.M.; Tomaro, M.L. Aluminium stress affects nitrogen fixation and assimilation in soybean (Glycine max L.). Plant Growth Regul. 2006, 48, 271–281. [Google Scholar] [CrossRef]
- Kahkonen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant activity of plant extracts containing phenolic compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient (mg L−1) | Control | 50F/50D | 50F/50D + S |
|---|---|---|---|
| NO3− | 606.70 ± 54.19 | 249.74 ± 38.27 | 550.22 ± 36.31 |
| PO43− | 367.73 ± 10.50 | 290.54 ± 8.09 | 445.75 ± 7.97 |
| SO42− | 380.46 ± 4.24 | 338.76 ± 2.19 | 392.04 ± 4.48 |
| K+ | 458.44 ± 12.32 | 193.32 ± 7.59 | 429.89 ± 7.15 |
| Ca2+ | 76.70 ± 3.07 | 80.61 ± 2.86 | 81.06 ± 2.84 |
| Mg2+ | 65.68 ± 0.97 | 66.20 ± 0.96 | 68.60 ± 1.29 |
| Irrigation | PGPR | Treatments |
|---|---|---|
| 100S | 0 | 100S |
| T1 | 100S + T1 | |
| T2 | 100S + T2 | |
| 50F/50D | 0 | 50F/50D |
| T1 | 50F/50D + T1 | |
| T2 | 50F/50D + T2 | |
| 50F/50D + S | 0 | 50F/50D + S |
| T1 | 50F/50D + S + T1 | |
| T2 | 50F/50D + S + T2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piñero, M.C.; Collado-González, J.; Otálora, G.; López-Marín, J.; del Amor, F.M. Plant Growth-Promoting Rhizobacteria as Tools to Improve the Growth of Kohlrabi (Brassica oleracea var. gongylodes) Plants in an Aquaponics System. Plants 2024, 13, 595. https://doi.org/10.3390/plants13050595
Piñero MC, Collado-González J, Otálora G, López-Marín J, del Amor FM. Plant Growth-Promoting Rhizobacteria as Tools to Improve the Growth of Kohlrabi (Brassica oleracea var. gongylodes) Plants in an Aquaponics System. Plants. 2024; 13(5):595. https://doi.org/10.3390/plants13050595
Chicago/Turabian StylePiñero, María Carmen, Jacinta Collado-González, Ginés Otálora, Josefa López-Marín, and Francisco M. del Amor. 2024. "Plant Growth-Promoting Rhizobacteria as Tools to Improve the Growth of Kohlrabi (Brassica oleracea var. gongylodes) Plants in an Aquaponics System" Plants 13, no. 5: 595. https://doi.org/10.3390/plants13050595