Structurally Preserved Liquidambar Infructescences, Associated Pollen, and Leaves from the Late Oligocene of the Nanning Basin, South China

 , , and
, , and
Abstract
:
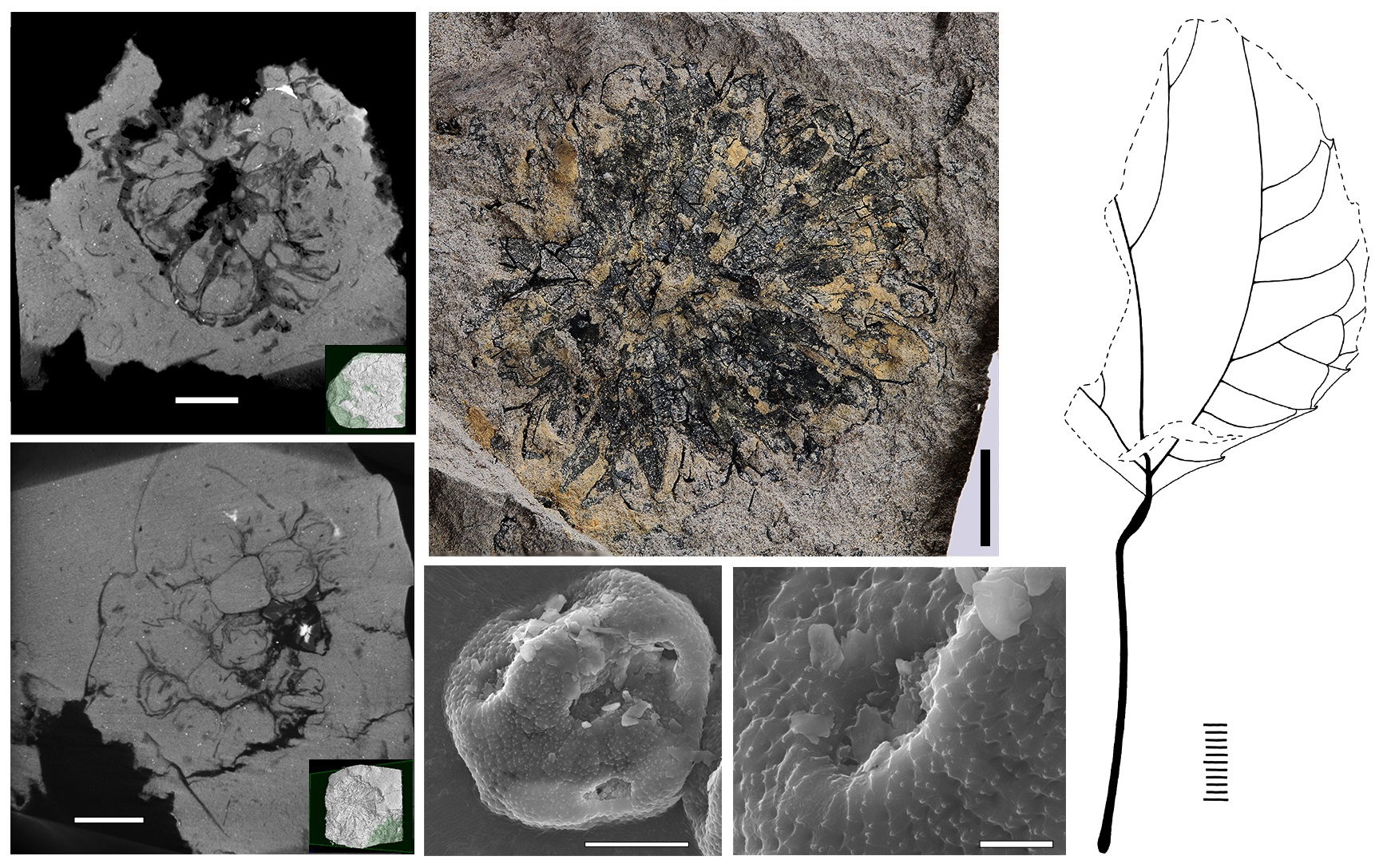{kind=link}
{kind=link}
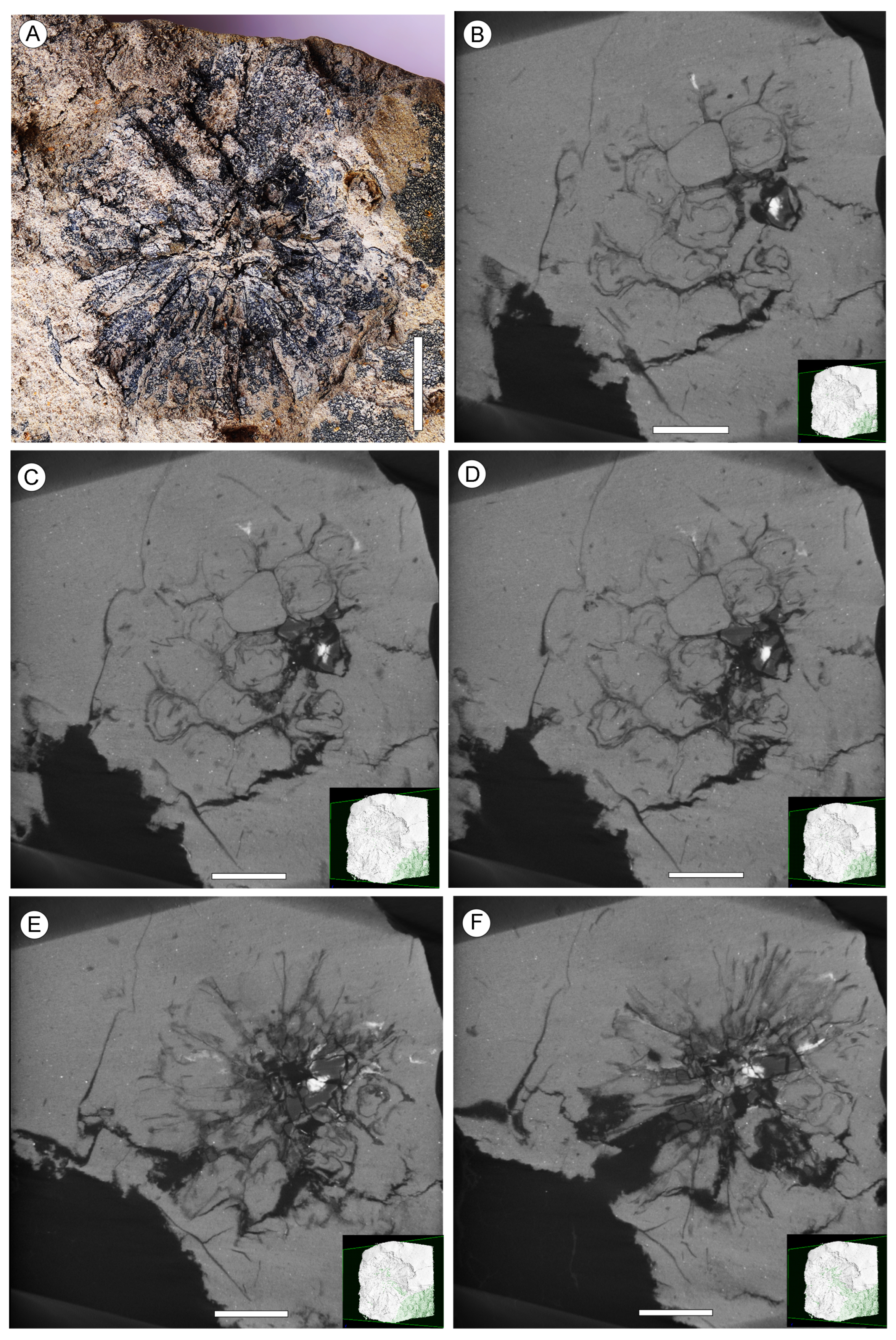{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Systematic Botany
2.2. Liquidambar sp. 1
2.3. Liquidambar sp. 2
3. Discussion
3.1. Morphological Comparison of Studied Fossils with Extant and Fossil Asian Liquidambar Species
3.1.1. Reproductive Structures
3.1.2. Leaves
3.2. Fungi Associated with L. nanningensis Heads
3.3. Taxonomic Significance of New Liquidambar Fossils from the Nanning Basin
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ickert-Bond, S.; Wen, J. A taxonomic synopsis of Altingiaceae with nine new combinations. Phytokeys 2013, 31, 21–61. [Google Scholar] [CrossRef] [PubMed]
- Ickert-Bond, S.M.; Pigg, K.B.; Wen, J. Comparative infructescence morphology in Liquidambar (Altingiaceae) and its evolutionary significance. Am. J. Bot. 2005, 92, 1234–1255. [Google Scholar] [CrossRef] [PubMed]
- Ickert-Bond, S.M.; Pigg, K.B.; Wen, J. Comparative infructescence morphology in Altingia (Altingiaceae) and discordance between morphological and molecular phylogenies. Am. J. Bot. 2007, 94, 1094–1115. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, F.H. Fossil Flora of the John Day Basin, Oregon; U.S. Geological Survey: Washington, DC, USA, 1902; Volume 204, pp. 1–153.
- Endo, S.; Morita, H. Notes on the genera Comptoniphyllum and Liquidambar. Sci. Rep. Tohoku Imp. Univ. 1932, 15, 41–53. [Google Scholar]
- Brown, R.W. A Cretaceous Sweet Gum. Bot. Gaz. 1933, 94, 611–615. [Google Scholar] [CrossRef]
- Hu, H.H.; Chaney, R.W. A Miocene Flora from Shantung Province, China; Carnegie Institution of Washington Publication: Washington, DC, USA, 1940. [Google Scholar]
- MacGinitie, H.D. A Middle Eocene Flora from the Central Sierra Nevada; Carnegie Institution of Washington Publication: Washington, DC, USA, 1941. [Google Scholar]
- Boytsova, E.P.; Pokrovskaya, I.M. Contributions to the stratigraphy of continental Oligocene and Miocene deposits of the Turgay Depression. In Materialy po Palinologii i Stratigrafii (Sbornik Statey); Pokrovskaya, I.M., Ed.; Gosudarstvennoe Nauchno-tekhnicheskoe Izdatelstvo Literatury po Geologii i Okhrane Nedr: Moscow, Russia, 1954; pp. 86–113. (In Russian) [Google Scholar]
- Makarova, Z.I. On the history of the genus Liquidambar L. Bot. Zh. 1957, 42, 1182–1195. [Google Scholar]
- Chaney, R.W.; Axelrod, D.I. Miocene Floras of the Columbia Plateau; Carnegie Institution of Washington Publication: Washington, DC, USA, 1959; Volume 134. [Google Scholar]
- Suzuki, K. The important and characteristic Pliocene and Miocene species of plants from the southern parts of the Tohoku District, Japan. Sci. Rep. Fac. Arts Sci. Fukushima Univ. 1961, 10, 1–95. [Google Scholar]
- Ferguson, D.K. The Miocene flora of Kreuzau, Western Germany. I. The Leaf-Remains. Verh. K. Ned. Akad. Wet. Afd. Natuurkd. Ser. II 1971, 60, 1–297. [Google Scholar]
- Ferguson, D.K. A survey of the Liquidambaroideae (Hamamelidaceae) with a view to elucidating its fossil record. In Evolution, Systematics, and Fossil History of the Hamamelidae; Crane, P.R., Blackmore, S., Eds.; Clarendon Press: Oxford, UK, 1989; Volume 1, pp. 249–272. [Google Scholar]
- Huzioka, K. The Tertiary floras of Korea. J. Min. Coll. Akita Univ. Ser. A 1972, 5, 1–83. [Google Scholar]
- Koch, B.E.; Friedrich, W.L.; Christensen, E.F.; Friis, E.M. Den miocæne brunkulsflora og dens geologiske miljø i Søby-Fasterholt området sydøst for Herning. Dansk Geol. Foren. Årsskr. for 1972 1973, 1–57. [Google Scholar]
- Onoe, T.A. Middle Miocene flora from Ogunimachi, Yamagata Prefecture, Japan. Rep. Geol. Surv. Jpn. 1974, 253, 1–644. [Google Scholar]
- Huzioka, K.; Uemura, K. The Comptonia-Liquidambar forest during middle Miocene Daijima age in Japan. Rep. Res. Inst. Undergr. Resour. Min. Coll. Akita Univ. 1979, 45, 37–50. [Google Scholar]
- Wolfe, J.A.; Tanai, T. The Miocene Seldovia Point Flora from the Kenai Group, Alaska; Geological Survey Professional Paper; U.S. Department of the Interior: Washington, DC, USA, 1980; Volume 1105, pp. 1–52.
- Uemura, K. Late Neogene Liquidambar (Hamamelidaceae) from the southern part of northeast Honshu, Japan. Mem. Natl. Sci. Mus. Tokyo 1983, 16, 25–36. [Google Scholar]
- Ozaki, K. Late Miocene and Pliocene floras in Central Honshu, Japan. Bull. Kanagawa Prefect. Mus. (Nat. Sci.) Spec. Issue 1991, 1–244. [Google Scholar]
- Maslova, N.P. Liquidambar L. from the Cenozoic of eastern Asia. Paleontol. J. 1995, 29, 145–158. [Google Scholar]
- Zhang, Z.; Lu, A. Hamamelidaceae: Geographic distribution, fossil history and origin. Acta Phytotax. Sin. 1995, 33, 313–339. [Google Scholar]
- Meyer, H.W.; Manchester, S.R. The Oligocene Bridge Creek flora of the John Day Formation, Oregon; University of California Press: Berkeley, CA, USA, 1997. [Google Scholar]
- Martinetto, E. East Asian elements in the Plio-Pleistocene floras of Italy. In Proceedings of the International Symposium of Floristic Characteristics and Diversity of East Asian Plants, Kunming, China, 25–27 July 1996; Zhang, A., Wu, S.G., Eds.; Springer: Berlin, Germany, 1998; pp. 71–87. [Google Scholar]
- Stults, D.Z.; Axsmith, B.J. Filling the gaps in the Neogene plant fossil record of Eastern North America: New data from the Pliocene of Alabama. Rev. Palaeobot. Palynol. 2011, 167, 1–9. [Google Scholar] [CrossRef]
- Vieira, M.; Poças, E.; Pais, J.; Pereira, D. Pliocene flora from S. Pedro da Torre deposits (Minho, NW Portugal). Geodiversitas 2011, 33, 71–85. [Google Scholar] [CrossRef]
- Xiao, L.; Sun, B.N.; Li, X.C.; Ren, W.X.; Jia, H. Anatomical variations of living and fossil Liquidambar leaves: A proxy for paleoenvironmental reconstruction. Sci. China Earth Sci. 2011, 54, 493–508. [Google Scholar] [CrossRef]
- Worobiec, G.; Worobiec, E.; Szynkiewicz, A. Plant assemblage from the Upper Miocene deposits of the Bełchatow Lignite Mine (Central Poland). Acta Palaeobot. 2012, 52, 369–413. [Google Scholar]
- Maslova, N.P.; Kodrul, T.M.; Song, Y.S.; Volkova, L.D.; Jin, J.H. Liquidambar maomingensis sp. nov. (Altingiaceae) from the late Eocene of South China. Am. J. Bot. 2015, 102, 1356–1370. [Google Scholar] [CrossRef]
- Maslova, N.P.; Kodrul, T.M.; Herman, A.B.; Tu, M.; Liu, X.Y.; Jin, J.H. A new species of Liquidambar (Altingiaceae) from the late Eocene of South China. J. Plant Res. 2019, 132, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Maslova, N.P.; Kodrul, T.M.; Kachkina, V.V.; Hofmann, C.C.; Xu, S.L.; Liu, X.Y.; Jin, J.H. Evidence for the evolutionary history and diversity of fossil sweetgums: Leaves and associated capitate reproductive structures of Liquidambar from the Eocene of Hainan Island, South China. Pap. Palaeontol. 2023, 9, e1540. [Google Scholar] [CrossRef]
- Dong, J.L.; Sun, B.N.; Mao, T.; Yan, D.F.; Liu, C.H.; Wang, Z.X.; Jin, P.H. Liquidambar (Altingiaceae) and associated insect herbivory from the Miocene of southeastern China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 497, 11–24. [Google Scholar] [CrossRef]
- Denk, T.; Bouchal, J.M.; Smirnov, P.; Trubin, Y. Late Oligocene leaf and pollen flora of Southwestern Siberia: Taxonomy, biogeography and palaeoenvironments. Hist. Biol. 2021, 33, 2951–2976. [Google Scholar] [CrossRef]
- Graham, A. The Sucker Creek and Trout Creek Miocene Floras of Southeastern Oregon; Kent State University Press: Kent, OH, USA, 1965. [Google Scholar]
- Kuprianova, L.A. Palynological data contributing to the history of Liquidambar. Pollen Spores 1960, 2, 71–88. [Google Scholar]
- Muller, J. Fossil pollen records of modern angiosperms. Bot. Rev. 1981, 47, 1–142. [Google Scholar] [CrossRef]
- Song, Z.C.; Zheng, Y.H.; Li, M.Y.; Zhang, Y.Y.; Wang, W.M.; Wang, D.N.; Zhao, C.B.; Zhou, S.F.; Zhu, Z.H.; Zhao, Y.N. The Late Cretaceous and Tertiary Spores and Pollen; Science Press: Beijing, China, 1999; Volume 1. [Google Scholar]
- Melchior, R.C. Paleobotany of the Williamsburg Formation (Paleocene) at the Santee Rediversion site, Berkeley County, South Carolina. In Paleobiology of the Williamsburg Formation (Black Mingo Group; Paleocene) of South Carolina, USA; Sanders, A.E., Ed.; American Philosophical Society: Philadelphia, PA, USA, 1998; Volume 4, pp. 49–121. [Google Scholar]
- Wheeler, E.A.; Lee, S.J.; Baas, P. Wood anatomy of the Altingiaceae and Hamamelidaceae. IAWA J. 2010, 31, 399–423. [Google Scholar] [CrossRef]
- Hofmann, C.C.; Spicer, R.A.; Ahlberg, A.; Herman, A.B. Scanning electron microscopy investigation of monads and tetrads of basal core eudicots from the Upper Cretaceous Vilui Basin, Siberia: Evidence for reticulate evolution. Rev. Palaeobot. Palynol. 2011, 167, 196–211. [Google Scholar] [CrossRef]
- Pigg, K.B.; Ickert-Bond, S.M.; Wen, J. Anatomically preserved Liquidambar (Altingiaceae) from the Middle Miocene of Yakima Canyon, Washingon State, USA, and its biogeographic implications. Am. J. Bot. 2004, 91, 499–509. [Google Scholar] [CrossRef]
- Zhou, Z.K.; Crepet, W.L.; Nixon, K.C. The earliest fossil evidence of the Hamamelidaceae: Late Cretaceous (Turonian) inflorescences and fruits of Altingioideae. Am. J. Bot. 2001, 88, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Gandolfo, M.A.; Crepet, W.L.; Nixon, K.C. Paleoaltingia Gen. Nov., a new genus of Altingiaceae from the Late Cretaceous of New Jersey. Appl. Plant Sci. 2021, 108, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Scharfstein, A.R.; Stockey, R.A.; Rothwell, G.W. Evolution and Phylogeny of Altingiaceae: Anatomically Preserved Infructescences from Late Cretaceous Deposits of Vancouver Island, British Columbia, Canada. Int. J. Plant Sci. 2020, 181, 452–463. [Google Scholar] [CrossRef]
- Kirchheimer, F. Über Steinhauera subglobosa Presl. und die Reste von Liquidambar-Fruchtständen aus dem Tertiär Mitteleuropas. Neues Jahrb. Miner. Geol. Paläontol. Monatsh. Abt. B 1943, 8/9, 216–236. [Google Scholar]
- Mai, H.D. Zwei ausgestorbene Gattungen im Tertiär Europas und ihre florengeschichtliche Bedeutung. Palaeontogr. Abt. B 1968, 123, 184–199. [Google Scholar]
- Strullu-Derrien, C.; Spencer, A.R.T.; Kenrick, P.; Judd, W.S.; De Franceschi, D.; Manchester, S.R. Revisions to the Eocene carpoflora of Anjou, western France, with new data from X-ray tomography. Bot. Lett. 2022, 169, 454–465. [Google Scholar] [CrossRef]
- Martinetto, E. The role of central Italy as a centre of refuge for thermophilous plants in the late Cenozoic. Acta Palaeobot. 2001, 41, 299–319. [Google Scholar]
- Li, X.C. The Late Cenozoic Floras from Eastern Zhejiang Province and Their Paleoclimatic Reconstruction. Ph.D. Thesis, Lanzhou University, College of Enviromental Sciences, Lanzhou, China, 2010. [Google Scholar]
- He, C.X.; Tao, J.R. A study on the Eocene flora in Yilan County, Heilongjiang. Acta Phytotax. Sin. 1997, 35, 249–256. [Google Scholar]
- Manchester, S.R.; Chen, Z.D.; Geng, B.Y.; Tao, J.R. Middle Eocene flora of Huadian, Jilin Province, Northeastern China. Acta Palaeobot. 2005, 45, 3–26. [Google Scholar]
- Oskolski, A.A.; Kodrul, T.M.; Jin, J.H. Altingioxylon hainanensis sp. nov.: Earliest fossil wood record of the family Altingiaceae in Eastern Asia and its implications for historical biogeography. Plant Syst. Evol. 2012, 298, 661–669. [Google Scholar] [CrossRef]
- Huang, L.L.; Jin, J.H.; Quan, C.; Oskolski, A.A. New occurrences of Altingiaceae fossil woods from the Miocene and upper Pleistocene of South China with phytogeographic implications. J. Palaeogeogr. 2021, 10, 482–493. [Google Scholar] [CrossRef]
- Maslova, N.P. Systematics of fossil platanoids and hamamelids. Paleontol. J. 2010, 44, 1379–1466. [Google Scholar] [CrossRef]
- Maslova, N.P. A new plant of the family Platanaceae from the early Paleogene reconstructed on the basis of leaves and inflorescences. Paleontol. J. 2002, 36, 207–218. [Google Scholar]
- Ickert-Bond, S.M.; Wen, J. Phylogeny and biogeography of Altingiaceae: Evidence from combined analysis of five non-coding chloroplast regions. Mol. Phylogenet. Evol. 2006, 39, 512–528. [Google Scholar] [CrossRef] [PubMed]
- Li, H.M.; Guo, S.X. Angiosperm. In Paleontology Atlas of East China (Mesozoic and Cenozoic volumes); Geology and Mineral Resources Ministry, Nanjing Geology and Mineral Resources Institute, Eds.; Geological Publishing House: Beijing, China, 1982; pp. 294–316. [Google Scholar]
- Ina, H.; Saito, T.; Kawase, M.; Wang, W.M. Fossil leaves, fruits and pollen of Liquidambar (Hamamelidaceae) from the lower Miocene Nakamura Formation in Gifu Prefecture, Central Japan. J. Geol. Soc. Jpn. 2007, 113, 542–545. [Google Scholar] [CrossRef]
- Kryshtofovich, A.N. Palaeobotanika, 4th ed.; Gosudarstvennoe Nauchno-Tekhnicheskoe Izdatelstvo Neftyanoi i Gorno-Toplivnoi Literatury, Leningradskoe Otdelenie: Leningrad, USSR, 1957. (In Russian) [Google Scholar]
- Aleksandrova, G.N.; Kodrul, T.M.; Jin, J.H. Palynological and paleobotanical investigations of Paleogene sections in the Maoming Basin, South China. Strat. Geol. Correl. 2015, 23, 300–325. [Google Scholar] [CrossRef]
- Wang, W.M.; Chen, G.J.; Liao, W. Oligocene palynoflora from Nanning Basin in Guangxi and its palaeoenvironmental significance. Quat. Sci. 2015, 35, 650–659. [Google Scholar]
- Ying, T.; Shaw, D.; Schneider, S. Oligocene fossil assemblages from Lake Nanning (Yongning Formation; Nanning Basin, Guangxi Province, SE China): Biodiversity and evolutionary implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 505, 100–119. [Google Scholar] [CrossRef]
- Tang, H.; Li, S.F.; Su, T.; Spicer, R.A.; Zhang, S.T.; Li, S.H.; Liu, J.; Lauretano, V.; Witkowski, C.R.; Spicer, T.E.V.; et al. Early Oligocene vegetation and climate of southwestern China inferred from palynology. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 560, 109988. [Google Scholar] [CrossRef]
- Bogle, A.L.; Philbrick, C.T. A Generic Atlas of Hamamelidaceous Pollens. Contrib. Gray Herb. 1980, 210, 29–103. [Google Scholar] [CrossRef]
- Zhang, J.T. Pollen morphological characteristics of modern and certain fossils of the genus Liquidambar. Acta Bot. Sin. 1958, 7, 215–229. [Google Scholar]
- Grand, L.F.; Vernia, C.S. A Preliminary checklist of fungi in the Nags Head Woods maritime forest in North Carolina. Castanea 2002, 67, 324–328. [Google Scholar]
- Han, J.G.; Shin, H.D. New Record of Xylaria persicaria on Liquidambar Fruits in Korea. Mycobiology 2007, 35, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Voglmayr, H.; Jaklitsch, W.M. Molecular data reveal high host specificity in the phylogenetically isolated genus Massaria (Ascomycota, Massariaceae). Fungal Divers. 2011, 46, 133–170. [Google Scholar] [CrossRef]
- Zhao, Z.R. The Vertebrate fossils and Lower Tertiary from Nanning Basin, Guangxi. Vertebr. Palasiat. 1981, 19, 218–227. [Google Scholar]
- Deng, D.G.; Wu, Y. Features of the Palaeogene sedimentary facies of the Nanning Basin, Guangxi. Guangxi Geol. 1992, 65, 23–30. [Google Scholar]
- Zhao, Z.R. New anthracothere materials from the Paleogene of Guangxi. Vertebr. Palasiat. 1993, 31, 13–190. [Google Scholar]
- Zhao, Z.R. A new species of anthracothere from Nanning Basin, Guangxi. Vertebr. Palasiat. 1983, 21, 266–270. [Google Scholar]
- Quan, C.; Fu, Q.Y.; Shi, G.L.; Liu, Y.S.; Li, L.; Liu, X.; Jin, J.H. First Oligocene mummified plant lagerstätte at the low latitudes of East Asia. Sci. China Earth Sci. 2016, 59, 445–448. [Google Scholar] [CrossRef]
- Huang, L.L.; Jin, J.H.; Quan, C.; Oskolski, A.A. Camellia nanningensis sp. nov.: The earliest fossil wood record of the genus Camellia (Theaceae) from East Asia. J. Plant Res. 2016, 129, 823–831. [Google Scholar] [CrossRef]
- Shi, X.G.; Fu, Q.Y.; Jin, J.H.; Quan, C. Mummified Oligocene fruits of Schima (Theaceae) and their systematic and biogeographic implications. Sci. Rep. 2017, 7, 4009. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.Y.; Li, L.; Jin, J.H.; Liu, X.Y.; Quan, C. Mummified fruits of Choerospondias nanningensis sp. nov. (Anacardiaceae) from the upper Oligocene of a low latitude site in East Asia. J. Syst. Evol. 2017, 55, 477–483. [Google Scholar] [CrossRef]
- Huang, L.L.; Jin, J.H.; Quan, C.; Oskolski, A.A. Mummified fossil woods of Fagaceae from the upper Oligocene of Guangxi, South China. J. Asian Earth Sci. 2018, 152, 39–51. [Google Scholar] [CrossRef]
- Ellis, B.; Daly, D.C.; Hickey, L.J.; Johnson, K.R.; Mitchell, J.D.; Wilf, P.; Wing, S.L. Manual of Leaf Architecture; Cornell University Press: Ithaca, NY, USA, 2009. [Google Scholar]
- Kvaček, Z. Whole-plant Reconstructions in Fossil Angiosperm Research. Int. J. Plant Sci. 2008, 169, 918–927. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, S.-L.; Maslova, N.; Kodrul, T.; Zdravchev, N.; Kachkina, V.; Liu, X.-Y.; Wu, X.-K.; Jin, J.-H. Structurally Preserved Liquidambar Infructescences, Associated Pollen, and Leaves from the Late Oligocene of the Nanning Basin, South China. Plants 2024, 13, 275. https://doi.org/10.3390/plants13020275
Xu S-L, Maslova N, Kodrul T, Zdravchev N, Kachkina V, Liu X-Y, Wu X-K, Jin J-H. Structurally Preserved Liquidambar Infructescences, Associated Pollen, and Leaves from the Late Oligocene of the Nanning Basin, South China. Plants. 2024; 13(2):275. https://doi.org/10.3390/plants13020275
Chicago/Turabian StyleXu, Sheng-Lan, Natalia Maslova, Tatiana Kodrul, Nikita Zdravchev, Vasilisa Kachkina, Xiao-Yan Liu, Xin-Kai Wu, and Jian-Hua Jin. 2024. "Structurally Preserved Liquidambar Infructescences, Associated Pollen, and Leaves from the Late Oligocene of the Nanning Basin, South China" Plants 13, no. 2: 275. https://doi.org/10.3390/plants13020275