
Genome and Transcriptome Analyses of Genes Involved in Ascorbate Biosynthesis in Pepper Indicate Key Genes Related to Fruit Development, Stresses, and Phytohormone Exposures
 , and
, and 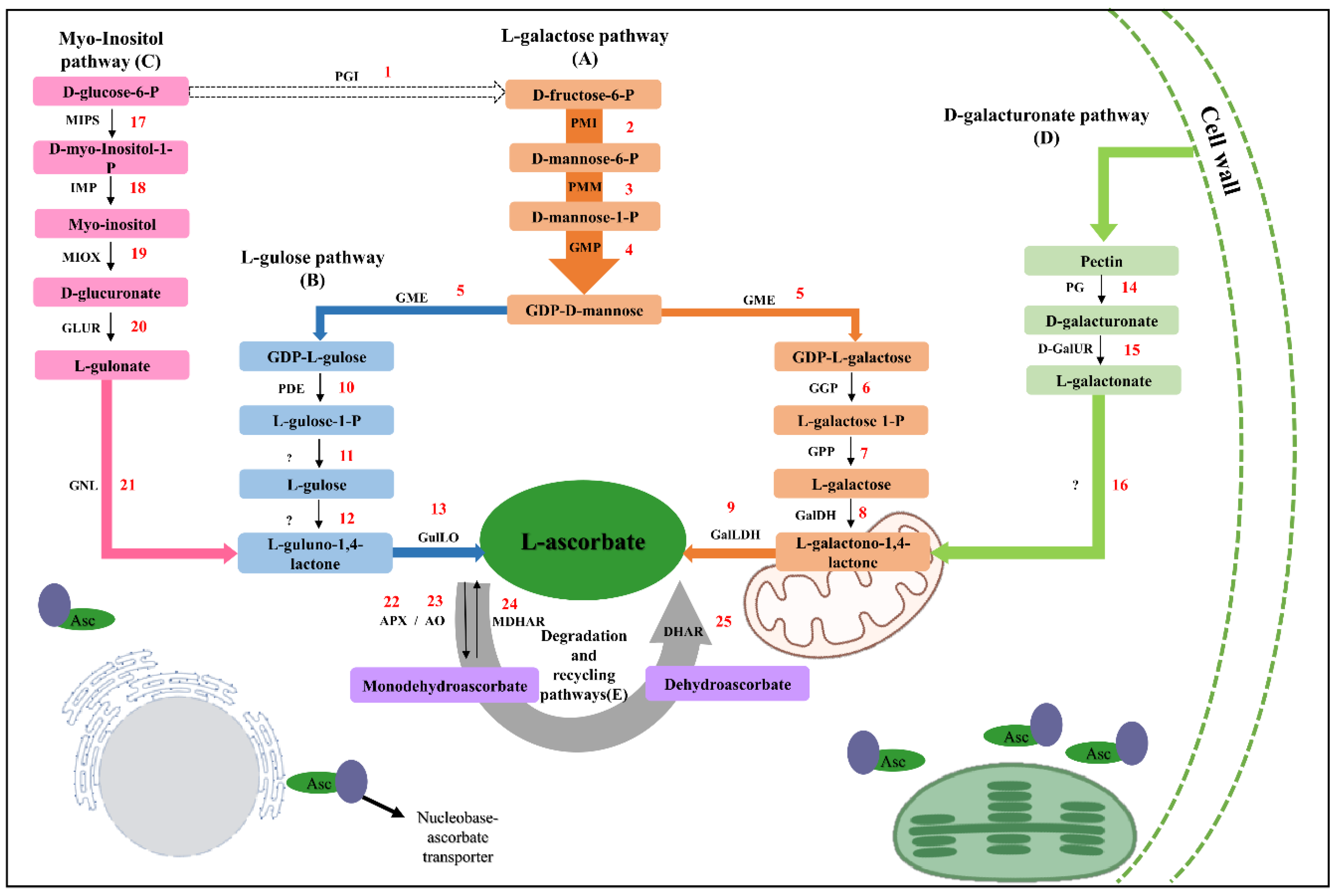{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of Genes Encoding Ascorbate Biosynthesis Enzymes
2.2. Expression Profile of Genes of Ascorbate Biosynthesis Enzymes during Fruit Development
2.3. Expression Profile of Genes of Ascorbate Biosynthesis Enzymes under Abiotic Stresses
2.4. Expression Profile of Genes of Ascorbate Biosynthesis under Biotic Stresses
2.5. Expression Profile of Genes of Ascorbate Biosynthesis Enzymes under Phytohormones Treatments
2.6. Cis-Acting Elements in the Promoter Region of Genes Involved in Ascorbate Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Identification and Annotation of Genes Involved in Ascorbate Biosynthesis
4.2. Gene Expression Analyses Using Capsicum Annuum RNA-Seq Experiments
4.3. Searches for Regulatory Cis-Elements in Genes Promoter Regions
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smirnoff, N.; Wheeler, G.L. Ascorbic Acid in Plants: Biosynthesis and Function. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 291–314. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, G.; Ishikawa, T.; Pornsaksit, V.; Smirnoff, N. Evolution of Alternative Biosynthetic Pathways for Vitamin C Following Plastid Acquisition in Photosynthetic Eukaryotes. Elife 2015, 4, e06369. [Google Scholar] [CrossRef] [PubMed]
- Nishikimi, M.; Koshizaka, T.; Ozawa, T.; Yagi, K. Occurrence in Humans and Guinea Pigs of the Gene Related to Their Missing Enzyme L-Gulono-γ-Lactone Oxidase. Arch. Biochem. Biophys. 1988, 267, 842–846. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N. Ascorbic Acid Metabolism and Functions: A Comparison of Plants and Mammals. Free Radic. Biol. Med. 2018, 122, 116–129. [Google Scholar] [CrossRef]
- Blaschke, K.; Ebata, K.T.; Karimi, M.M.; Zepeda-Martínez, J.A.; Goyal, P.; Mahapatra, S.; Tam, A.; Laird, D.J.; Hirst, M.; Rao, A.; et al. Vitamin C Induces Tet-Dependent DNA Demethylation and a Blastocyst-like State in ES Cells. Nature 2013, 500, 222–226. [Google Scholar] [CrossRef]
- Macknight, R.C.; Laing, W.A.; Bulley, S.M.; Broad, R.C.; Johnson, A.A.; Hellens, R.P. Increasing Ascorbate Levels in Crops to Enhance Human Nutrition and Plant Abiotic Stress Tolerance. Curr. Opin. Biotechnol. 2017, 44, 153–160. [Google Scholar] [CrossRef]
- Ali, B.; Pantha, S.; Acharya, R.; Ueda, Y.; Wu, L.B.; Ashrafuzzaman, M.; Ishizaki, T.; Wissuwa, M.; Bulley, S.; Frei, M. Enhanced Ascorbate Level Improves Multi-Stress Tolerance in a Widely Grown Indica Rice Variety without Compromising Its Agronomic Characteristics. J. Plant Physiol. 2019, 240, 152998. [Google Scholar] [CrossRef] [PubMed]
- Raja, V.; Wani, U.M.; Wani, Z.A.; Jan, N.; Kottakota, C.; Reddy, M.K.; Kaul, T.; John, R. Pyramiding Ascorbate–Glutathione Pathway in Lycopersicum esculentum Confers Tolerance to Drought and Salinity Stress. Plant Cell Rep. 2022, 41, 619–637. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant Adaptations to the Combination of Drought and High Temperatures. Physiol. Plant. 2018, 162, 2–12. [Google Scholar] [CrossRef]
- Wheeler, G.L.; Jones, M.A.; Smirnoff, N. The Biosynthetic Pathway of Vitamin C in Higher Plants. Nature 1998, 393, 365–369. [Google Scholar] [CrossRef]
- Lorence, A.; Chevone, B.I.; Mendes, P.; Nessler, C.L. Myo-Inositol Oxygenase Offers a Possible Entry Point into Plant Ascorbate Biosynthesis. Plant Physiol. 2004, 134, 1200–1205. [Google Scholar] [CrossRef] [PubMed]
- Badejo, A.A.; Wada, K.; Gao, Y.; Maruta, T.; Sawa, Y.; Shigeoka, S.; Ishikawa, T. Translocation and the Alternative D-Galacturonate Pathway Contribute to Increasing the Ascorbate Level in Ripening Tomato Fruits Together with the D-Mannose/L-Galactose Pathway. J. Exp. Bot. 2012, 63, 229–239. [Google Scholar] [CrossRef]
- Bulley, S.M.; Rassam, M.; Hoser, D.; Otto, W.; Schünemann, N.; Wright, M.; MacRae, E.; Gleave, A.; Laing, W. Gene Expression Studies in Kiwifruit and Gene Over-Expression in Arabidopsis Indicates That GDP-L-Galactose Guanyltransferase Is a Major Control Point of Vitamin C Biosynthesis. J. Exp. Bot. 2009, 60, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Agius, F.; González-Lamothe, R.; Caballero, J.L.; Muñoz-Blanco, J.; Botella, M.A.; Valpuesta, V. Engineering Increased Vitamin C Levels in Plants by Overexpression of a D-Galacturonic Acid Reductase. Nat. Biotechnol. 2003, 21, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Wolucka, B.A.; Van Montagu, M. GDP-Mannose 3′,5′-Epimerase Forms GDP-L-Gulose, a Putative Intermediate for the de Novo Biosynthesis of Vitamin C in Plants. J. Biol. Chem. 2003, 278, 47483–47490. [Google Scholar] [CrossRef] [PubMed]
- Ivanov Kavkova, E.; Blöchl, C.; Tenhaken, R. The Myo-Inositol Pathway Does Not Contribute to Ascorbic Acid Synthesis. Plant Biol. 2019, 21, 95–102. [Google Scholar] [CrossRef]
- Alós, E.; Rey, F.; Gil, J.V.; Rodrigo, M.J.; Zacarias, L. Ascorbic Acid Content and Transcriptional Profiling of Genes Involved in Its Metabolism during Development of Petals, Leaves, and Fruits of Orange (Citrus sinensis Cv. Valencia Late). Plants 2021, 10, 2590. [Google Scholar] [CrossRef]
- Loewus, F.A. Biosynthesis and Metabolism of Ascorbic Acid in Plants and of Analogs of Ascorbic Acid in Fungi. Phytochemistry 1999, 52, 193–210. [Google Scholar] [CrossRef]
- Truffault, V.; Fry, S.C.; Stevens, R.G.; Gautier, H. Ascorbate Degradation in Tomato Leads to Accumulation of Oxalate, Threonate and Oxalyl Threonate. Plant J. 2017, 89, 996–1008. [Google Scholar] [CrossRef]
- Castro, J.C.; Maddox, J.D.; Cobos, M.; Requena, D.; Zimic, M.; Bombarely, A.; Imán, S.A.; Cerdeira, L.A.; Medina, A.E. De Novo Assembly and Functional Annotation of Myrciaria dubia Fruit Transcriptome Reveals Multiple Metabolic Pathways for L-Ascorbic Acid Biosynthesis. BMC Genomics 2015, 16, 997. [Google Scholar] [CrossRef]
- Alós, E.; Rodrigo, M.J.; Zacarías, L. Differential Transcriptional Regulation of L-Ascorbic Acid Content in Peel and Pulp of Citrus Fruits during Development and Maturation. Planta 2014, 239, 1113–1128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Huang, J.; Li, X. Transcriptomic Analysis Reveals the Metabolic Mechanism of L-Ascorbic Acid in Ziziphus jujuba Mill. Front. Plant Sci. 2016, 7, 122. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Rus, E.; Amaya, I.; Sánchez-Sevilla, J.F.; Botella, M.A.; Valpuesta, V. Regulation of L-Ascorbic Acid Content in Strawberry Fruits. J. Exp. Bot. 2011, 62, 4191–4201. [Google Scholar] [CrossRef]
- de Ong, W.; Voo, L.Y.C.; Kumar, V.S. De Novo Assembly, Characterization and Functional Annotation of Pineapple Fruit Transcriptome through Massively Parallel Sequencing. PLoS ONE 2012, 7, e46937. [Google Scholar] [CrossRef]
- dos Santos, C.P.; Batista, M.C.; da Cruz Saraiva, K.D.; Roque, A.L.M.; de Souza Miranda, R.; Alexandre e Silva, L.M.; Moura, C.F.H.; Alves Filho, E.G.; Canuto, K.M.; Costa, J.H. Transcriptome Analysis of Acerola Fruit Ripening: Insights into Ascorbate, Ethylene, Respiration, and Softening Metabolisms. Plant Mol. Biol. 2019, 101, 269–296. [Google Scholar] [CrossRef]
- Yabuta, Y.; Mieda, T.; Rapolu, M.; Nakamura, A.; Motoki, T.; Maruta, T.; Yoshimura, K.; Ishikawa, T.; Shigeoka, S. Light Regulation of Ascorbate Biosynthesis Is Dependent on the Photosynthetic Electron Transport Chain but Independent of Sugars in Arabidopsis. J. Exp. Bot. 2007, 58, 2661–2671. [Google Scholar] [CrossRef]
- Badejo, A.A.; Fujikawa, Y.; Esaka, M. Gene Expression of Ascorbic Acid Biosynthesis Related Enzymes of the Smirnoff-Wheeler Pathway in Acerola (Malpighia glabra). J. Plant Physiol. 2009, 166, 652–660. [Google Scholar] [CrossRef]
- Zhang, G.Y.; Liu, R.R.; Zhang, C.Q.; Tang, K.X.; Sun, M.F.; Yan, G.H.; Liu, Q.Q. Manipulation of the Rice L-Galactose Pathway: Evaluation of the Effects of Transgene Overexpression on Ascorbate Accumulation and Abiotic Stress Tolerance. PLoS ONE 2015, 10, e0125870. [Google Scholar] [CrossRef]
- Ma, L.; Wang, Y.; Liu, W.; Liu, Z. Overexpression of an Alfalfa GDP-Mannose 3, 5-Epimerase Gene Enhances Acid, Drought and Salt Tolerance in Transgenic Arabidopsis by Increasing Ascorbate Accumulation. Biotechnol. Lett. 2014, 36, 2331–2341. [Google Scholar] [CrossRef]
- Liu, W.; An, H.M.; Yang, M. Overexpression of Rosa roxburghii L-Galactono-1,4-Lactone Dehydrogenase in Tobacco Plant Enhances Ascorbate Accumulation and Abiotic Stress Tolerance. Acta Physiol. Plant. 2013, 35, 1617–1624. [Google Scholar] [CrossRef]
- Lisko, K.A.; Torres, R.; Harris, R.S.; Belisle, M.; Vaughan, M.M.; Jullian, B.; Chevone, B.I.; Mendes, P.; Nessler, C.L.; Lorence, A. Elevating Vitamin C Content via Overexpression of Myo-Inositol Oxygenase and l-Gulono-1,4-Lactone Oxidase in Arabidopsis Leads to Enhanced Biomass and Tolerance to Abiotic Stresses. Vitr. Cell. Dev. Biol. Plant 2013, 49, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Zhang, C.; Ye, J.; Hu, T.; Ye, Z.; Li, H.; Zhang, Y. Ectopic Expression of FaGalUR Leads to Ascorbate Accumulation with Enhanced Oxidative Stress, Cold, and Salt Tolerance in Tomato. Plant Growth Regul. 2015, 76, 187–197. [Google Scholar] [CrossRef]
- Alós, E.; Rodrigo, M.J.; Zacarías, L. Transcriptomic Analysis of Genes Involved in the Biosynthesis, Recycling and Degradation of L-Ascorbic Acid in Pepper Fruits (Capsicum annuum L.). Plant Sci. 2013, 207, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Gómez-García, M.d.R.; Ochoa-Alejo, N. Predominant Role of the L-Galactose Pathway in l-Ascorbic Acid Biosynthesis in Fruits and Leaves of the Capsicum annuum L. Chili Pepper. Rev. Bras. Bot. 2016, 39, 157–168. [Google Scholar] [CrossRef]
- González-Gordo, S.; Bautista, R.; Claros, M.G.; Cañas, A.; Palma, J.M.; Corpas, F.J. Nitric Oxide-Dependent Regulation of Sweet Pepper Fruit Ripening. J. Exp. Bot. 2019, 70, 4557–4570. [Google Scholar] [CrossRef]
- Liu, Z.; Lv, J.; Zhang, Z.; Li, H.; Yang, B.; Chen, W.; Dai, X.; Li, X.; Yang, S.; Liu, L.; et al. Integrative Transcriptome and Proteome Analysis Identifies Major Metabolic Pathways Involved in Pepper Fruit Development. J. Proteome Res. 2019, 18, 982–994. [Google Scholar] [CrossRef]
- Liu, Y.; Lv, J.; Liu, Z.; Wang, J.; Yang, B.; Chen, W.; Ou, L.; Dai, X.; Zhang, Z.; Zou, X. Integrative Analysis of Metabolome and Transcriptome Reveals the Mechanism of Color Formation in Pepper Fruit (Capsicum annuum L.). Food Chem. 2020, 306, 125629. [Google Scholar] [CrossRef]
- Kang, W.H.; Sim, Y.M.; Koo, N.; Nam, J.Y.; Lee, J.; Kim, N.; Jang, H.; Kim, Y.M.; Yeom, S.I. Transcriptome Profiling of Abiotic Responses to Heat, Cold, Salt, and Osmotic Stress of Capsicum annuum L. Sci. Data 2020, 7, 17. [Google Scholar] [CrossRef]
- Miao, W.; Song, J.; Huang, Y.; Liu, R.; Zou, G.; Ou, L.; Liu, Z. Comparative Transcriptomics for Pepper (Capsicum annuum L.) under Cold Stress and after Rewarming. Appl. Sci. 2021, 11, 10204. [Google Scholar] [CrossRef]
- Guo, H.; Tian, H.; Wang, Y.; Xiong, X.; Tian, H.; Zhang, S.; Geng, G. Comparative Analysis of Growth, Physiological and Transcriptomic Response Revealed Mechanisms of Waterlogging Tolerance of Hot Pepper (Capsicum annuum var. conoides). Plant Breed. 2022, 141, 807–819. [Google Scholar] [CrossRef]
- Escalante, C.; Sela, N.; Valverde, R.A. Transcriptome Analysis of Two Near-Isogenic Lines of Bell Pepper (Capsicum annuum) Infected with Bell Pepper Endornavirus and Pepper Mild Mottle Virus. Front. Genet. 2023, 14, 1182578. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.F.; Tucker Hallmark, H.; Ramaraj, T.; Sundararajan, A.; Schilkey, F.; Rashotte, A.M. Three Strains of Tobacco Etch Virus Distinctly Alter the Transcriptome of Apical Stem Tissue in Capsicum annuum during Infection. Viruses 2021, 13, 741. [Google Scholar] [CrossRef]
- Lee, J.; Nam, J.Y.; Jang, H.; Kim, N.; Kim, Y.M.; Kang, W.H.; Yeom, S.I. Comprehensive Transcriptome Resource for Response to Phytohormone-Induced Signaling in Capsicum annuum L. BMC Res. Notes 2020, 13, 440. [Google Scholar] [CrossRef]
- Ioannidi, E.; Kalamaki, M.S.; Engineer, C.; Pateraki, I.; Alexandrou, D.; Mellidou, I.; Giovannonni, J.; Kanellis, A.K. Expression Profiling of Ascorbic Acid-Related Genes during Tomato Fruit Development and Ripening and in Response to Stress Conditions. J. Exp. Bot. 2009, 60, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.; Creissen, G.; Kular, B.; Firmin, J.; Robinson, S.; Verhoeyen, M.; Mullineaux, P. Changes in Oxidative Processes and Components of the Antioxidant System during Tomato Fruit Ripening. Planta 2002, 214, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Arce-Rodríguez, M.L.; Martínez, O.; Ochoa-Alejo, N. Genome-Wide Identification and Analysis of the Myb Transcription Factor Gene Family in Chili Pepper (Capsicum Spp.). Int. J. Mol. Sci. 2021, 22, 2229. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, T.; Tao, N.; Yan, T.; Wang, Q.; Li, Q. Nitric Oxide Is Essential to Keep the Postharvest Quality of Fruits and Vegetables. Horticulturae 2023, 9, 135. [Google Scholar] [CrossRef]
- Rodríguez-Ruiz, M.; Mateos, R.M.; Codesido, V.; Corpas, F.J.; Palma, J.M. Characterization of the Galactono-1,4-Lactone Dehydrogenase from Pepper Fruits and Its Modulation in the Ascorbate Biosynthesis. Role of Nitric Oxide. Redox Biol. 2017, 12, 171–181. [Google Scholar] [CrossRef]
- Liu, X.; Wu, R.; Bulley, S.M.; Zhong, C.; Li, D. Kiwifruit MYBS1-like and GBF3 Transcription Factors Influence l-Ascorbic Acid Biosynthesis by Activating Transcription of GDP-L-Galactose Phosphorylase 3. N. Phytol. 2022, 234, 1782–1800. [Google Scholar] [CrossRef]
- Zheng, X.; Gong, M.; Zhang, Q.; Tan, H.; Li, L.; Tang, Y.; Li, Z.; Peng, M.; Deng, W. Metabolism and Regulation of Ascorbic Acid in Fruits. Plants 2022, 11, 1602. [Google Scholar] [CrossRef]
- Xiao, M.; Li, Z.; Zhu, L.; Wang, J.; Zhang, B.; Zheng, F.; Zhao, B.; Zhang, H.; Wang, Y.; Zhang, Z. The Multiple Roles of Ascorbate in the Abiotic Stress Response of Plants: Antioxidant, Cofactor, and Regulator. Front. Plant Sci. 2021, 12, 598173. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, Z.W.; Wu, Z.J.; Wang, Y.X.; Teng, R.M.; Zhuang, J. Differentially Expressed Protein and Gene Analysis Revealed the Effects of Temperature on Changes in Ascorbic Acid Metabolism in Harvested Tea Leaves. Hortic. Res. 2018, 5, 65. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, M.; Liang, D.; Cui, M.; Ma, F. Expression Patterns and Promoter Characteristics of the Gene Encoding Actinidia deliciosa L-Galactose-1-Phosphate Phosphatase Involved in the Response to Light and Abiotic Stresses. Mol. Biol. Rep. 2013, 40, 1473–1485. [Google Scholar] [CrossRef] [PubMed]
- Tsaniklidis, G.; Delis, C.; Nikoloudakis, N.; Katinakis, P.; Aivalakis, G. Low Temperature Storage Affects the Ascorbic Acid Metabolism of Cherry Tomato Fruits. Plant Physiol. Biochem. 2014, 84, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Massot, C.; Bancel, D.; Lauri, F.L.; Truffault, V.; Baldet, P.; Stevens, R.; Gautier, H. High Temperature Inhibits Ascorbate Recycling and Light Stimulation of the Ascorbate Pool in Tomato despite Increased Expression of Biosynthesis Genes. PLoS ONE 2013, 8, e84474. [Google Scholar] [CrossRef]
- Galli, V.; Messias, R.S.; Guzman, F.; Perin, E.C.; Margis, R.; Rombaldi, C.V. Transcriptome Analysis of Strawberry (Fragaria × Ananassa) Fruits under Osmotic Stresses and Identification of Genes Related to Ascorbic Acid Pathway. Physiol. Plant. 2019, 166, 979–995. [Google Scholar] [CrossRef]
- Azuma, R.; Ito, N.; Nakayama, N.; Suwa, R.; Nguyen, N.T.; Larrinaga-Mayoral, J.Á.; Esaka, M.; Fujiyama, H.; Saneoka, H. Fruits Are More Sensitive to Salinity than Leaves and Stems in Pepper Plants (Capsicum annuum L.). Sci. Hortic. 2010, 125, 171–178. [Google Scholar] [CrossRef]
- Tajaragh, R.P.; Rasouli, F.; Giglou, M.T.; Zahedi, S.M.; Hassanpouraghdam, M.B.; Aazami, M.A.; Adámková, A.; Mlček, J. Morphological and Physiological Responses of In Vitro-Grown Cucurbita Sp. Landraces Seedlings under Osmotic Stress by Mannitol and PEG. Horticulturae 2022, 8, 1117. [Google Scholar] [CrossRef]
- Bai, T.; Ma, P.; Li, C.; Yin, R.; Ma, F. Role of Ascorbic Acid in Enhancing Hypoxia Tolerance in Roots of Sensitive and Tolerant Apple Rootstocks. Sci. Hortic. 2013, 164, 372–379. [Google Scholar] [CrossRef]
- Lin, H.H.; Lin, K.H.; Syu, J.Y.; Tang, S.Y.; Lo, H.F. Physiological and Proteomic Analysis in Two Wild Tomato Lines under Waterlogging and High Temperature Stress. J. Plant Biochem. Biotechnol. 2016, 25, 87–96. [Google Scholar] [CrossRef]
- Li, J.; Cui, M.; Li, M.; Wang, X.; Liang, D.; Ma, F. Expression Pattern and Promoter Analysis of the Gene Encoding GDP-D-Mannose 3’,5’-Epimerase under Abiotic Stresses and Applications of Hormones by Kiwifruit. Sci. Hortic. 2013, 150, 187–194. [Google Scholar] [CrossRef]
- Botanga, C.J.; Bethke, G.; Chen, Z.; Gallie, D.R.; Fiehn, O.; Glazebrook, J. Metabolite Profiling of Arabidopsis Inoculated with Alternaria brassicicola Reveals That Ascorbate Reduces Disease Severity. Mol. Plant-Microbe Interact. 2012, 25, 1628–1638. [Google Scholar] [CrossRef] [PubMed]
- Jawaid, M.Z.; Khalid, M.F.; Ahmed, T. Seed Priming Mitigates the Salt Stress in Eggplant (Solanum melongena) by Activating Antioxidative Defense Mechanisms. SSRN 2023. [Google Scholar] [CrossRef]
- Ergin, S.; Aydogan, C.; Ozturk, N.; Turhan, E. Effects of Ascorbic Acid Application in Strawberry Plants During Heat Stress. Türk Tarım ve Doğa Bilim. Derg. 2014, 1, 1486–1491. [Google Scholar]
- Shalata, A.; Neumann, P.M. Exogenous Ascorbic Acid (Vitamin C) Increases Resistance to Salt Stress and Reduces Lipid Peroxidation. J. Exp. Bot. 2001, 52, 2207–2211. [Google Scholar] [CrossRef] [PubMed]
- Khafagy, M.A.; Arafa, A.; El-Banna, M.F. Glycinebetaine and Ascorbic Acid Can Alleviate the Harmful Effects of NaCl Salinity in Sweet Pepper. Aust. J. Crop Sci. 2009, 3, 257–267. [Google Scholar]
- Khazaei, Z.; Estaji, A. Effect of Foliar Application of Ascorbic Acid on Sweet Pepper (Capsicum annuum) Plants under Drought Stress. Acta Physiol. Plant. 2020, 42, 118. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Ahmad, I.; Basit, A.; Shehata, W.F.; Hassan, U.; Shah, S.T.; Haleema, B.; Jalal, A.; Amin, R.; Khalid, M.A.; et al. Ascorbic Acid Enhances Growth and Yield of Sweet Peppers (Capsicum annum) by Mitigating Salinity Stress. Gesunde Pflanz. 2022, 74, 423–433. [Google Scholar] [CrossRef]
- Zarrillo, A.; Minutolo, M.; Alioto, D.; Errico, A. Ascorbic Acid Regulation in Leaves and Fruits of Tomato Ecotypes Infected by Eggplant Mottled Dwarf Virus. Sci. Hortic. 2017, 225, 512–524. [Google Scholar] [CrossRef]
- Vioque, B.; Castellano, J.M. In Vivo and in Vitro 1-Aminocyclopropane-1-Carboxylic Acid Oxidase Activity in Pear Fruit: Role of Ascorbate and Inactivation during Catalysis. J. Agric. Food Chem. 1998, 46, 1706–1711. [Google Scholar] [CrossRef]
- Ye, N.; Zhu, G.; Liu, Y.; Zhang, A.; Li, Y.; Liu, R.; Shi, L.; Jia, L.; Zhang, J. Ascorbic Acid and Reactive Oxygen Species Are Involved in the Inhibition of Seed Germination by Abscisic Acid in Rice Seeds. J. Exp. Bot. 2012, 63, 1809–1822. [Google Scholar] [CrossRef] [PubMed]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic Acid-a Potential Oxidant Scavenger and Its Role in Plant Development and Abiotic Stress Tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wang, J.; Li, S.; Kakan, X.; Zhou, Y.; Miao, Y.; Wang, F.; Qin, H.; Huang, R. Ascorbic Acid Integrates the Antagonistic Modulation of Ethylene and Abscisic Acid in the Accumulation of Reactive Oxygen Species. Plant Physiol. 2019, 179, 1861–1875. [Google Scholar] [CrossRef] [PubMed]
- López-Carbonell, M.; Munné-Bosch, S.; Alegre, L. The Ascorbate-Deficient Vtc-1 Arabidopsis Mutant Shows Altered ABA Accumulation in Leaves and Chloroplasts. J. Plant Growth Regul. 2006, 25, 137–144. [Google Scholar] [CrossRef]
- Guo, W.L.; Chen, R.G.; Gong, Z.H.; Yin, Y.X.; Ahmed, S.S.; He, Y.M. Exogenous Abscisic Acid Increases Antioxidant Enzymes and Related Gene Expression in Pepper (Capsicum annuum) Leaves Subjected to Chilling Stress. Genet. Mol. Res. 2012, 11, 4063–4080. [Google Scholar] [CrossRef]
- Magwaza, L.S.; Mditshwa, A.; Tesfay, S.Z.; Opara, U.L. An Overview of Preharvest Factors Affecting Vitamin C Content of Citrus Fruit. Sci. Hortic. 2017, 216, 12–21. [Google Scholar] [CrossRef]
- Sayyari, M.; Babalar, M.; Kalantari, S.; Serrano, M.; Valero, D. Effect of Salicylic Acid Treatment on Reducing Chilling Injury in Stored Pomegranates. Postharvest Biol. Technol. 2009, 53, 152–154. [Google Scholar] [CrossRef]
- Mustafa, M.A.; Ali, A.; Seymour, G.; Tucker, G. Enhancing the Antioxidant Content of Carambola (Averrhoa carambola) during Cold Storage and Methyl Jasmonate Treatments. Postharvest Biol. Technol. 2016, 118, 79–86. [Google Scholar] [CrossRef]
- Bao, G.; Zhuo, C.; Qian, C.; Xiao, T.; Guo, Z.; Lu, S. Co-Expression of NCED and ALO Improves Vitamin C Level and Tolerance to Drought and Chilling in Transgenic Tobacco and Stylo Plants. Plant Biotechnol. J. 2016, 14, 206–214. [Google Scholar] [CrossRef]
- Jiang, M.; Liu, Y.; Ren, L.; She, X.; Chen, H. Light Regulates Ascorbic Acid Accumulation and Ascorbic Acid-Related Genes Expression in the Peel of Eggplant. S. Afr. J. Bot. 2018, 114, 20–28. [Google Scholar] [CrossRef]
- Wang, H.; Lu, S.; Guan, X.; Jiang, Y.; Wang, B.; Hua, J.; Zou, B. Dehydration-Responsive Element Binding Protein 1C, 1E, and 1G Promote Stress Tolerance to Chilling, Heat, Drought, and Salt in Rice. Front. Plant Sci. 2022, 13, 851731. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.H.; McDonald, A.E.; Arnholdt-Schmitt, B.; Fernandes de Melo, D. A Classification Scheme for Alternative Oxidases Reveals the Taxonomic Distribution and Evolutionary History of the Enzyme in Angiosperms. Mitochondrion 2014, 19, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Li, Z.; Sun, Y.; Aluko, O.O.; Wu, X.; Wang, Q.; Liu, G. MG2C: A User-Friendly Online Tool for Drawing Genetic Maps. Mol. Hortic. 2021, 1, 16. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van De Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguiar, É.S.d.; Dias, A.N.; Sousa, R.M.; Germano, T.A.; de Sousa, R.O.; Miranda, R.d.S.; Costa, J.H.; dos Santos, C.P. Genome and Transcriptome Analyses of Genes Involved in Ascorbate Biosynthesis in Pepper Indicate Key Genes Related to Fruit Development, Stresses, and Phytohormone Exposures. Plants 2023, 12, 3367. https://doi.org/10.3390/plants12193367
Aguiar ÉSd, Dias AN, Sousa RM, Germano TA, de Sousa RO, Miranda RdS, Costa JH, dos Santos CP. Genome and Transcriptome Analyses of Genes Involved in Ascorbate Biosynthesis in Pepper Indicate Key Genes Related to Fruit Development, Stresses, and Phytohormone Exposures. Plants. 2023; 12(19):3367. https://doi.org/10.3390/plants12193367
Chicago/Turabian StyleAguiar, Évelyn Silva de, Abigailde Nascimento Dias, Raquel Mendes Sousa, Thais Andrade Germano, Renato Oliveira de Sousa, Rafael de Souza Miranda, José Hélio Costa, and Clesivan Pereira dos Santos. 2023. "Genome and Transcriptome Analyses of Genes Involved in Ascorbate Biosynthesis in Pepper Indicate Key Genes Related to Fruit Development, Stresses, and Phytohormone Exposures" Plants 12, no. 19: 3367. https://doi.org/10.3390/plants12193367