
Selection of Reference Genes in Evodia rutaecarpa var. officinalis and Expression Patterns of Genes Involved in Its Limonin Biosynthesis

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Analysis of Relative Content of Limonin in E. rutaecarpa var. officinalis
2.2. Selection of Reference Genes and Primer Design
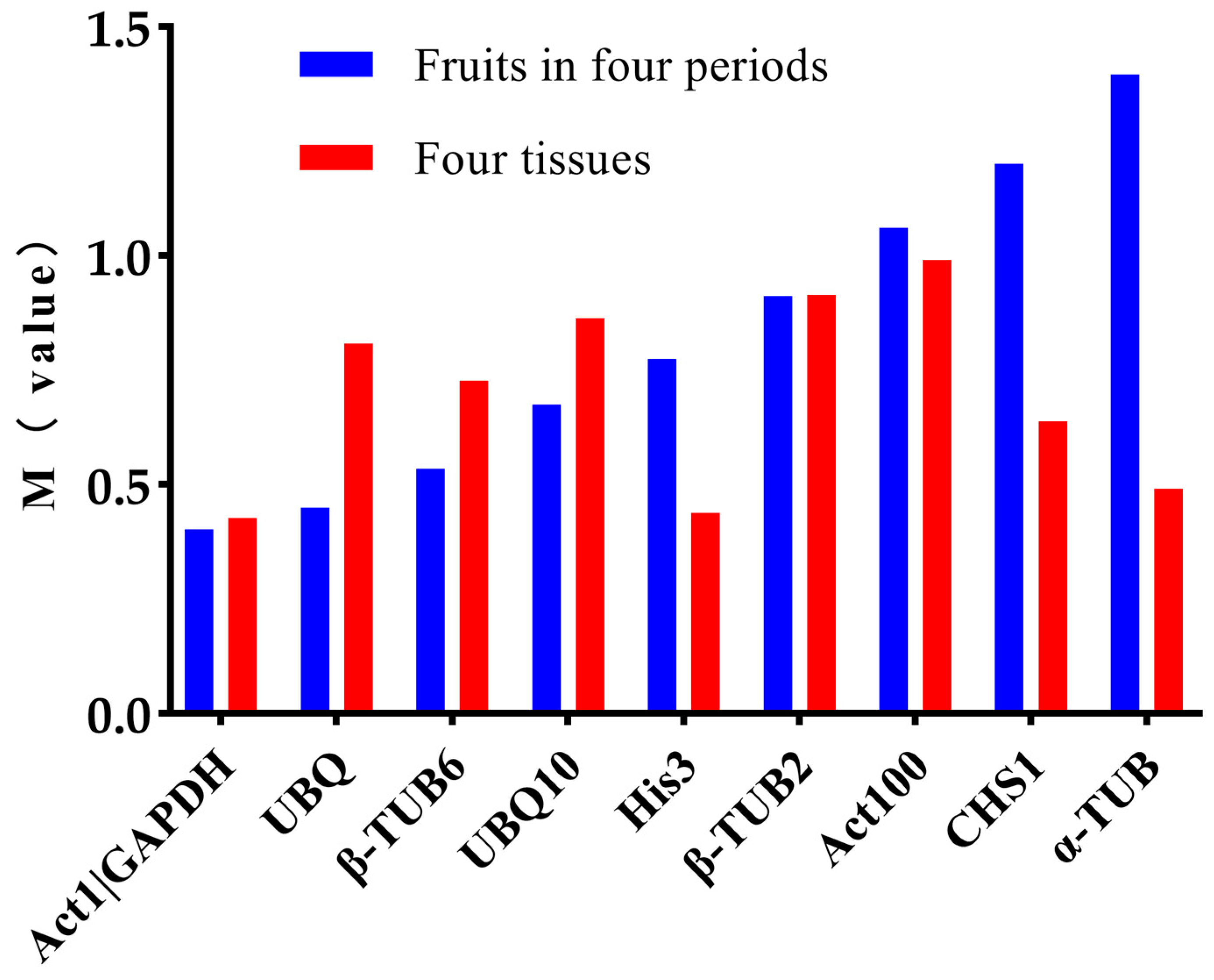
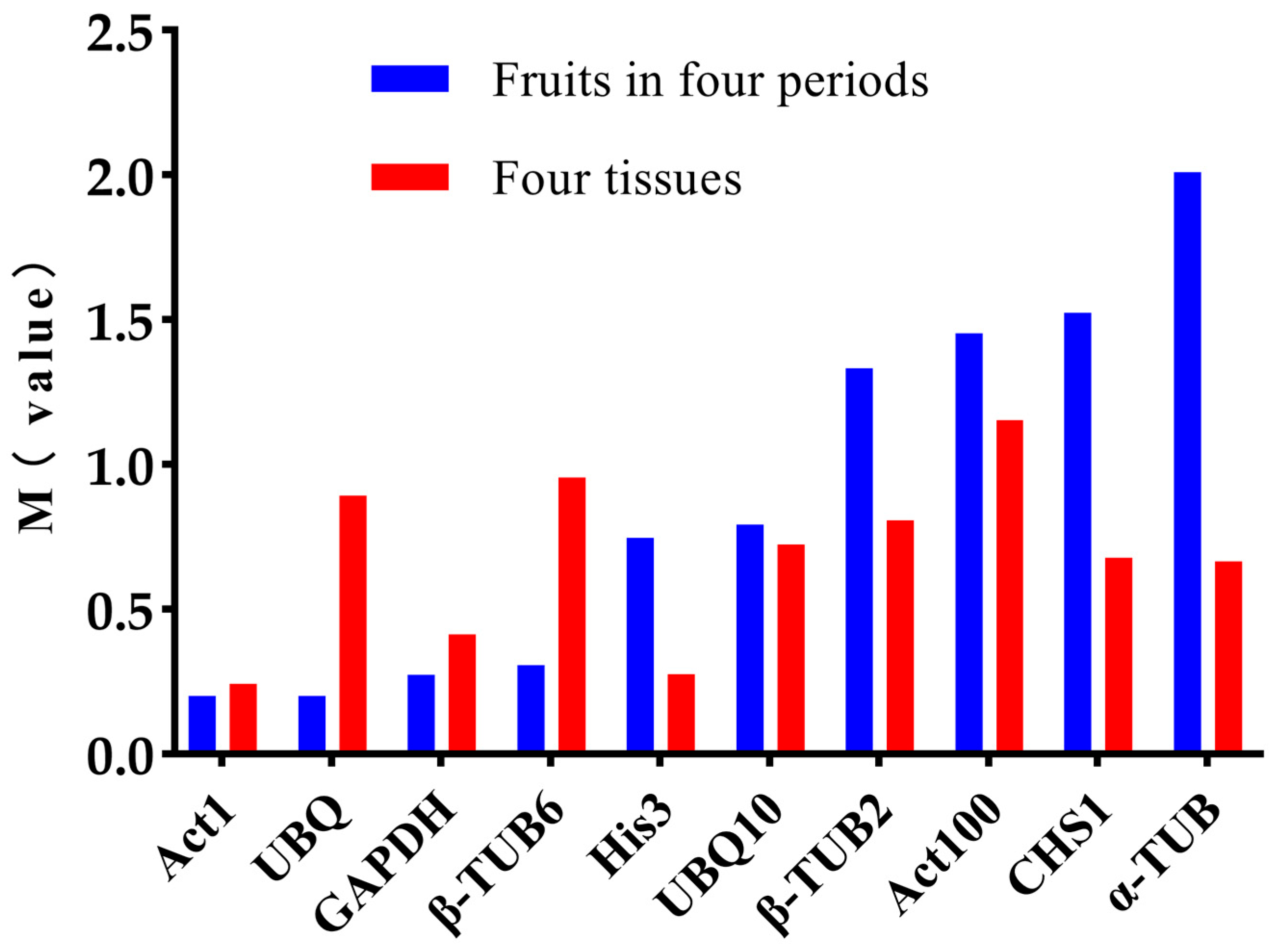
2.3. Expression Abundance Analysis of the Candidate Reference Genes
2.4. Stability Analysis of the Candidate Reference Genes
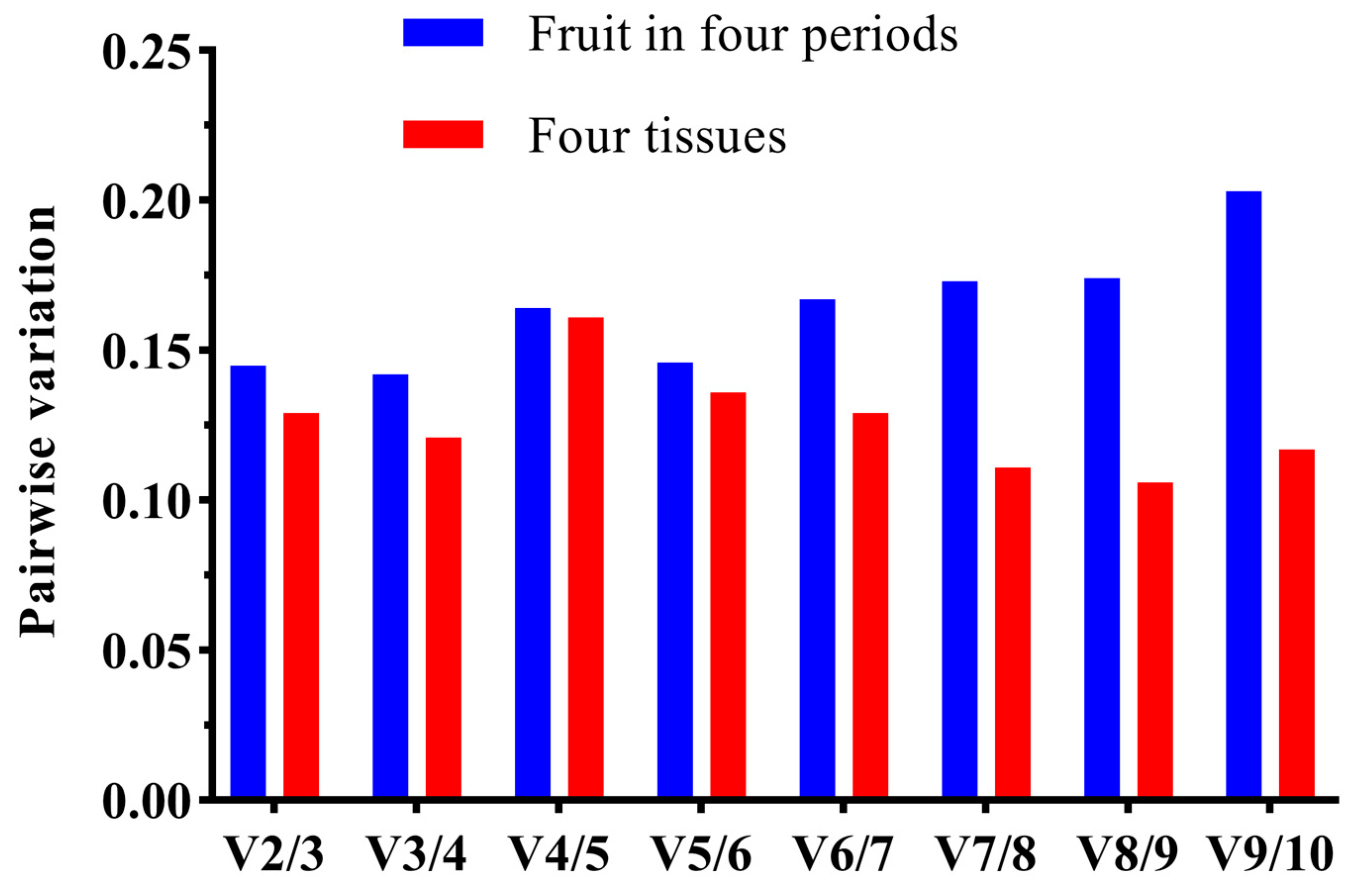
2.4.1. GeNorm Analysis
2.4.2. NormFinder Analysis
2.4.3. BestKeeper Analysis
2.4.4. Delta Ct Analysis
2.4.5. RefFinder Comprehensive Stability Analysis
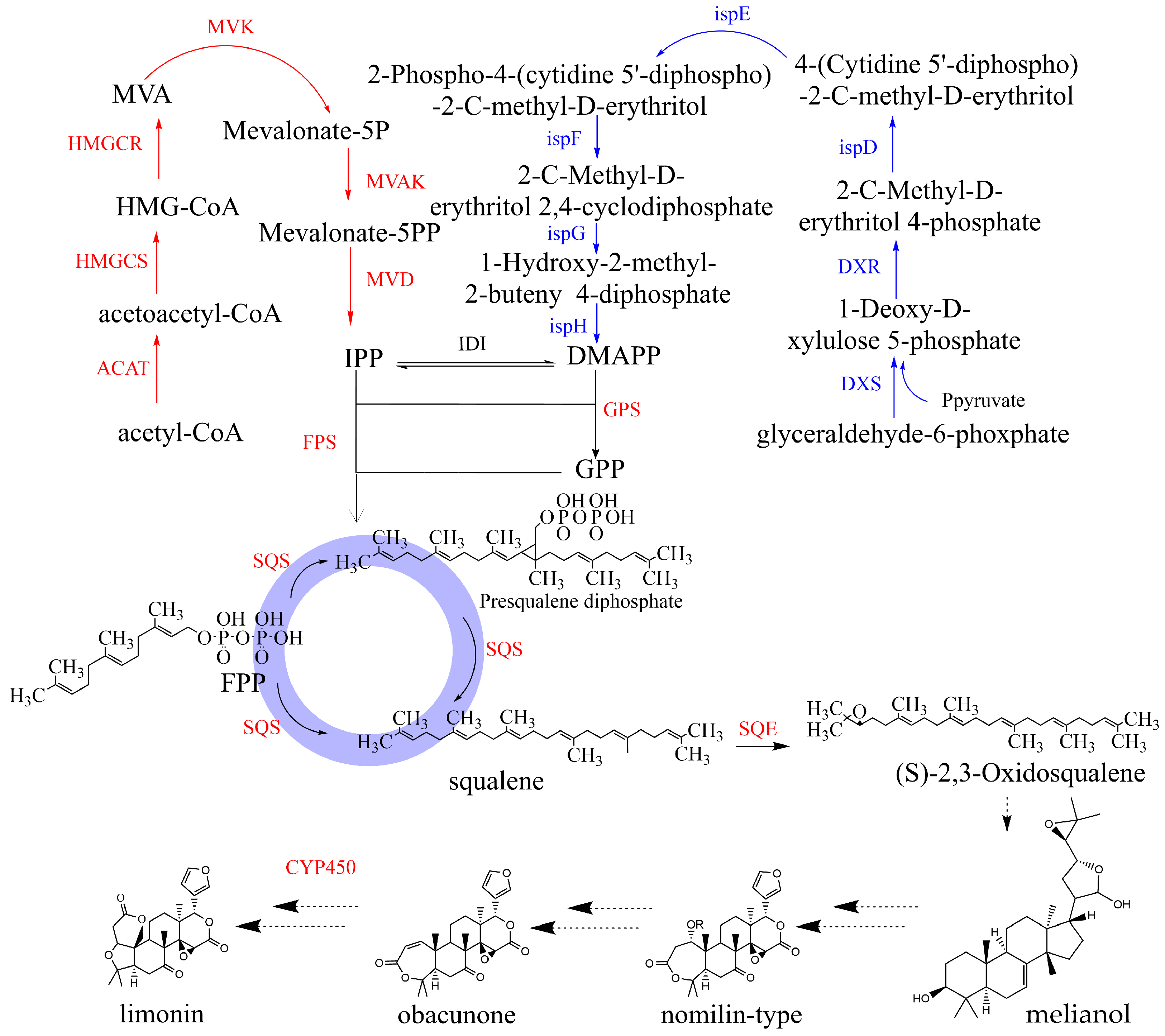
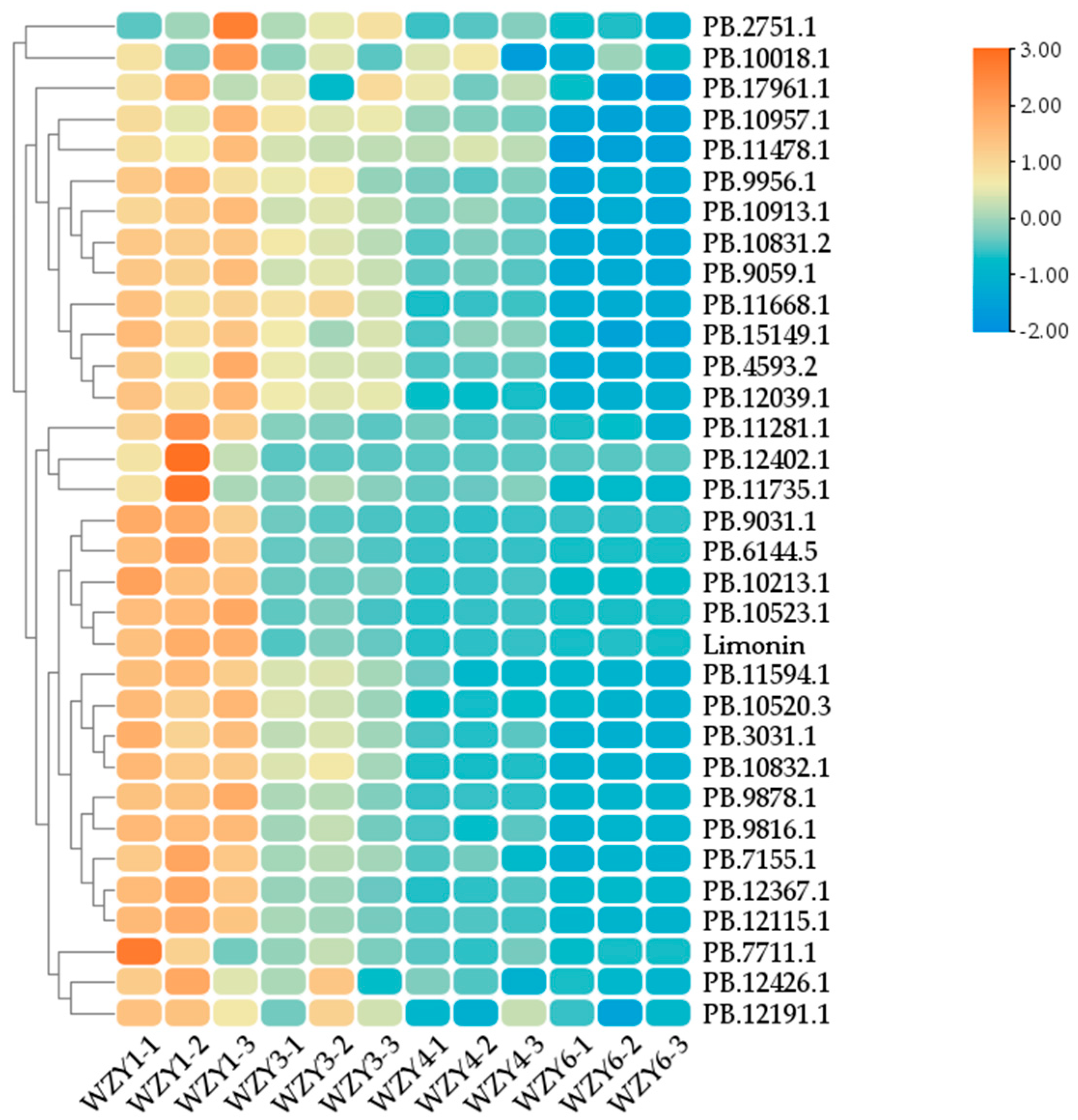
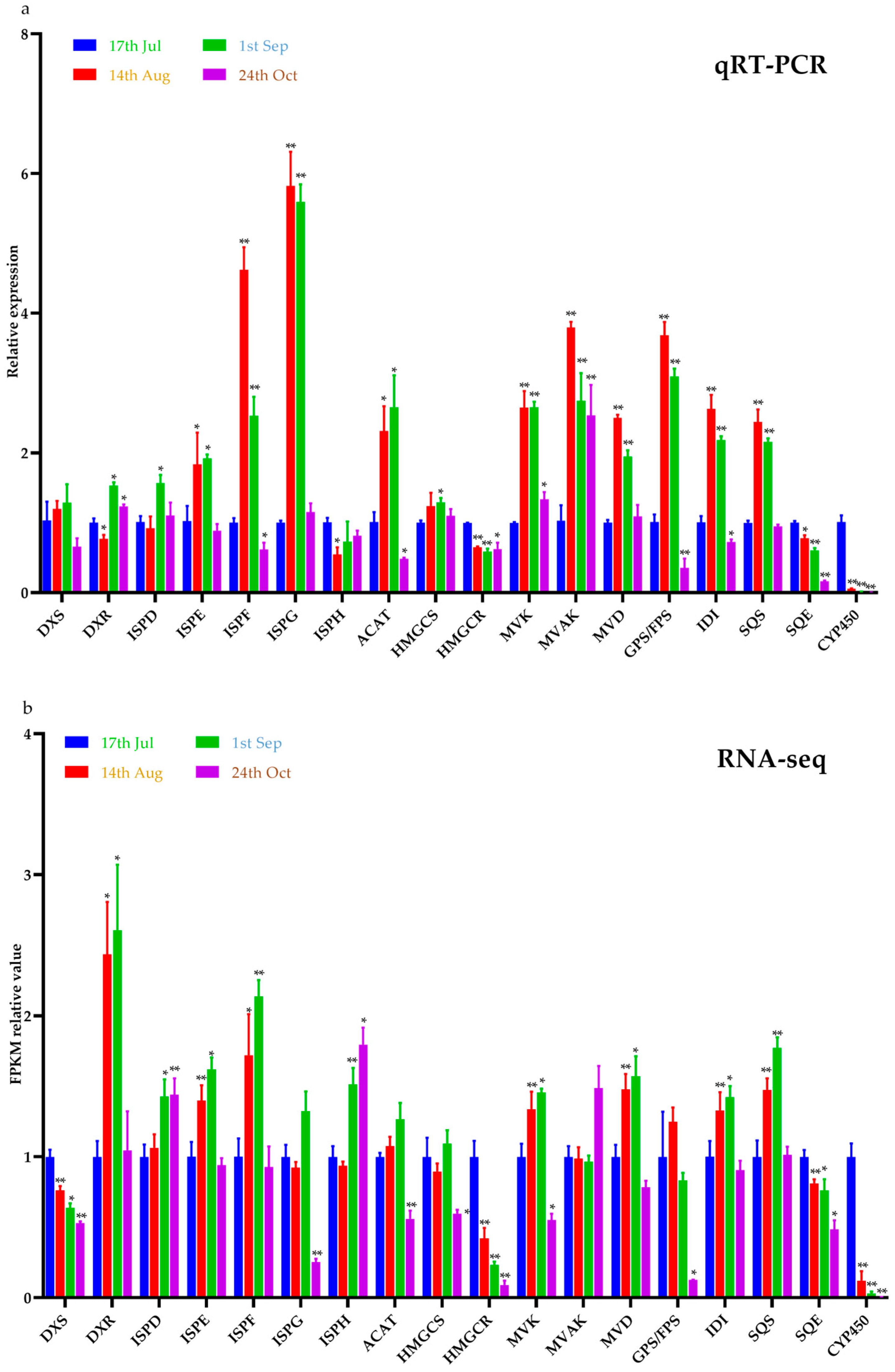
2.5. Gene Expression Analysis of the Related Enzymes in the Biosynthesis Pathway of Limonin in E. rutaecarpa var. officinalis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Extraction and Relative Content Calculation of Limonin
4.3. Total RNA Extraction and cDNA Synthesis
4.4. Selection of Reference Genes and Genes Involved in Limonin Biosynthesis
4.5. qRT-PCR Analysis
4.6. Analysis of the Candidate Reference Gene Expression Stability
4.7. Stastical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lee, S.H.; Son, J.-K.; Jeong, B.S.; Jeong, T.-C.; Chang, H.W.; Lee, E.-S.; Jahng, Y. Progress in the Studies on Rutaecarpine. Molecules 2008, 13, 272–300. [Google Scholar] [CrossRef]
- Yu, X.; Wu, D.-Z.; Yuan, J.-Y.; Zhang, R.-R.; Hu, Z.-B. Gastroprotective Effect of Fructus Evodiae Water Extract on Ethanol-Induced Gastric Lesions in Rats. Am. J. Chin. Med. 2006, 34, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, C.-P.; Deng, P.-Y.; Shen, S.-S.; Zhu, H.-Q.; Ding, J.-S.; Tan, G.-S.; Li, Y.-J. The protective effects of rutaecarpine on gastric mucosa injury in rats. Planta Medica 2005, 71, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, H.; Chen, L.; Zhao, H.; Liu, J.; Gong, S.; Ma, D.; Chen, C.; Zeng, S.; Long, H.; et al. Mechanism and Pharmacodynamic Substance Basis of Raw and Wine-Processed Evodia rutaecarpa on Smooth Muscle Cells of Dysmenorrhea Mice. Pain Res. Manag. 2023, 2023, 7711988. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, J.; Guo, J.; Wang, Y.; Wang, Q. Elucidation of Pharmacological Mechanism Underlying the Anti-Alzheimer’s Disease Effects of Evodia rutaecarpa and Discovery of Novel Lead Molecules: An In Silico Study. Molecules 2023, 28, 5846. [Google Scholar] [CrossRef] [PubMed]
- State Pharmacopoeia Commission. Pharmacopoeia of the China; China Pharmaceutical Science and Technology Press: Beijing, China, 2022. [Google Scholar]
- Zhang, P.-t.; Pan, B.-y.; Liao, Q.-f.; Yao, M.-c.; Xu, X.-j.; Wan, J.-z.; Liu, D.; Xie, Z.-y. Separation of Five Quinolone Alkaloids from Fruits of Evodia rutaecarpa by High-speed Counter-current Chromatography. Chin. Herb. Med. 2014, 6, 47–52. [Google Scholar] [CrossRef]
- Liang, J.; Chen, Y.; Ren, G.; Dong, W.; Shi, M.; Xiong, L.; Li, J.; Dong, J.; Li, F.; Yuan, J. Screening Hepatotoxic Components in Euodia rutaecarpa by UHPLC-QTOF/MS Based on the Spectrum-Toxicity Relationship. Molecules 2017, 22, 1264. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Q.-H.; Wu, L.-J.; Tashiro, S.-I.; Onodera, S.; Ikejima, T. Atypical apoptosis in L929 cells induced by evodiamine isolated from Evodia rutaecarpa. J. Asian Nat. Prod. Res. 2004, 6, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Wei, J.; Zhao, W.; Shi, S.; Zhang, Y.; Wei, R.; Zhang, Y.; Li, W.; Wang, Q. Toxicity of Evodiae fructus on Rat Liver Mitochondria: The Role of Oxidative Stress and Mitochondrial Permeability Transition. Molecules 2014, 19, 21168–21182. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.R.; Park, W.H.; Suh, D.H.; No, J.H.; Kim, Y.B.; Kim, K. Role of limonin in anticancer effects of Evodia rutaecarpa on ovarian cancer cells. BMC Complement. Med. Ther. 2020, 20, 94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, M.; Song, H.; Gao, C.; Wang, D.; Hua, H.; Pan, Y.; Liu, X. CYP3A4 inducer aggravates big flower Evodiae Fructus-induced hepatotoxicity whereas limonin attenuates its hepatotoxicity. J. Ethnopharmacol. 2021, 264, 113277. [Google Scholar] [CrossRef] [PubMed]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isopreno-id synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef] [PubMed]
- De La Peña, R.; Hodgson, H.; Liu, J.C.; Stephenson, M.J.; Martin, A.C.; Owen, C.; Harkess, A.; Leebens-Mack, J.; Jimenez, L.E.; Osbourn, A.; et al. Complex scaffold remodeling in plant triterpene biosynthesis. Science 2023, 379, 361–368. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Nolan, T.; Pfaffl, M.W. Quantitative real-time RT-PCR–A perspective. J. Mol. Endocrinol. 2005, 34, 597–601. [Google Scholar] [CrossRef]
- Ginzinger, D.G. Gene quantification using real-time quantitative PCR: An emerging technology hits the mainstream. Exp. Hematol. 2002, 30, 503–512. [Google Scholar] [CrossRef]
- Klein, D. Quantification using real-time PCR technology: Applications and limitations. Trends Mol. Med. 2002, 8, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Gaete-Eastman, C.; Mattus-Araya, E.; Herrera, R.; Moya-León, M.A. Evaluation of reference genes for transcript normalization in Fragaria chiloensis fruit and vegetative tissues. Physiol. Mol. Biol. Plants 2022, 28, 1535–1544. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, J.; Wang, X.; Xiong, Y.; Xiong, Y.; Dong, Z.; Lei, X.; Yan, L.; Ma, X. Selection and Validation of Reference Genes for qRT-PCR Gene Expression Analysis in Kengyilia melanthera. Genes 2022, 13, 1445. [Google Scholar] [CrossRef] [PubMed]
- Bharati, R.; Sen, M.K.; Kumar, R.; Gupta, A.; Sur, V.P.; Melnikovová, I.; Fernández-Cusimamani, E. Selection and Validation of the Most Suitable Reference Genes for Quantitative Real-Time PCR Normalization in Salvia rosmarinus under In Vitro Conditions. Plants 2022, 11, 2878. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Q.; Sun, M.; Zhu, L.; Yang, M.; Zhao, Y. Selection of Reference Genes for Quantitative Real-Time PCR Normalization in Panax ginseng at Different Stages of Growth and in Different Organs. PLoS ONE 2014, 9, e112177. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 31. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, J.; Liu, Y.; Chen, J.; Lin, H.; Huang, Y.; Bian, X.; Zhao, Y. Selection and validation of appropriate reference genes for real-time quantitative PCR analysis in Momordica charantia. Phytochemistry 2019, 164, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, J.; Eattock, N.; Van Deynze, A.; Blumwald, E. Selection and validation of reference genes for gene expression analysis in switchgrass (Panicum virgatum) using quantitative real-time RT-PCR. PLoS ONE 2014, 9, e91474. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, H.; Liu, L.; Li, W.; Wei, Y.; Shi, S. Validation of Reference Genes for RT-qPCR Studies of Gene Expression in Preharvest and Postharvest Longan Fruits under Different Experimental Conditions. Front. Plant Sci. 2016, 7, 780. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper--Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: a miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Wang, J.; Zhang, B. RefFinder: a web-based tool for comprehensively analyzing and identifying reference genes. Funct. Integr. Genom. 2023, 23, 125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Cai, Y.; Zhang, M.; Du, G.; Wang, J. Selection and Evaluation of Candidate Reference Genes for Quantitative Real-Time PCR in Aboveground Tissues and Drought Conditions in Rhododendron Delavayi. Front. Genet. 2022, 13, 876482. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, R.; Zhou, Z. Identification and Validation of Reference Genes for Gene Expression Analysis in Schima superba. Genes 2021, 12, 732. [Google Scholar] [CrossRef]
- Zhu, Z.; Yu, J.; Tang, X.; Xiong, A.; Sun, M. Selection and Validation of Reference Genes in Different Tissues of Okra (Abelmoschus esculentus L.) under Different Abiotic Stresses. Genes 2023, 14, 603. [Google Scholar] [CrossRef]
- Amil-Ruiz, F.; Garrido-Gala, J.; Blanco-Portales, R.; Folta, K.M.; Muñoz-Blanco, J.; Caballero, J.L. Identification and Validation of Reference Genes for Transcript Normalization in Strawberry (Fragaria × ananassa) Defense Responses. PLoS ONE 2013, 8, e70603. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Zhao, Z.; Qian, C.; Sui, Y.; Malik, A.A.; Chen, J. Selection of appropriate reference genes for gene expression studies by quantitative real-time polymerase chain reaction in cucumber. Anal. Biochem. 2010, 399, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xia, H.; Liu, X.; Lin, Z.; Guo, Y.; Deng, H.; Wang, J.; Lin, L.; Deng, Q.; Lv, X.; et al. Identification of Suitable Reference Genes for qRT-PCR Normalization in Kiwifruit. Horticulturae 2022, 8, 170. [Google Scholar] [CrossRef]
- Karuppaiya, P.; Yan, X.-X.; Liao, W.; Wu, J.; Chen, F.; Tang, L. Identification and validation of superior reference gene for gene expression normalization via RT-qPCR in staminate and pistillate flowers of Jatropha curcas–A biodiesel plant. PLoS ONE 2017, 12, e0172460. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, B.; Wang, X.; Wei, X. Screening of stable internal reference gene of Quinoa under hormone treatment and abiotic stress. Physiol. Mol. Biol. Plants 2021, 27, 2459–2470. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, H.; Yuan, Y. A Comparison of the Flavonoid Biosynthesis Mechanisms of Dendrobium Species by Analyzing the Transcriptome and Metabolome. Int. J. Mol. Sci. 2022, 23, 11980. [Google Scholar] [CrossRef]
- Lou, H.; Yang, Y.; Zheng, S.; Ma, Z.; Chen, W.; Yu, C.; Song, L.; Wu, J. Identification of key genes contributing to amino acid biosynthesis in Torreya grandis using transcriptome and metabolome analysis. Food Chem. 2022, 379, 132078. [Google Scholar] [CrossRef]
- Tang, Q.; Ma, X.; Mo, C.; Wilson, I.W.; Song, C.; Zhao, H.; Yang, Y.; Fu, W.; Qiu, D. An efficient approach to finding Siraitia grosvenorii triterpene biosynthetic genes by RNA-seq and digital gene expression analysis. BMC Genom. 2011, 12, 343. [Google Scholar] [CrossRef]
- Dai, L.; Liu, C.; Zhu, Y.; Zhang, J.; Men, Y.; Zeng, Y.; Sun, Y. Functional Characterization of Cucurbitadienol Synthase and Triterpene Glycosyltransferase Involved in Biosynthesis of Mogrosides from Siraitia grosvenorii. Plant Cell Physiol. 2015, 56, 1172–1182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Dai, L.; Yang, J.; Liu, C.; Men, Y.; Zeng, Y.; Cai, Y.; Zhu, Y.; Sun, Y. Oxidation of Cucurbitadienol Catalyzed by CYP87D18 in the Biosynthesis of Mogrosides from Siraitia grosvenorii. Plant Cell Physiol. 2016, 57, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Full Name | Primer Sequence (5′ to 3′) | Slope (k) | Amplification Efficiency (%) | Correlation Coefficients (R2) |
|---|---|---|---|---|---|
| α-TUB | α-tubulin | F: TCCTTTCAACCCATTCCC R: CTCGGTCACATCCACATTCA | −3.157 | 107.4 | 0.996 |
| β-TUB2 | β-tubulin2 | F: GCTACCTCACAGCCACAG R: CCATTTCATCCATACCCT | −3.113 | 109.5 | 0.990 |
| β-TUB6 | β-tubulin6 | F: AGTCGTGGAGCCATACAA R: AACGGAATCAGATTAACAGC | −3.140 | 108.2 | 0.996 |
| His 3 | Histone H3 | F: TAAATCAACTGGAGGAAAGG R: GAGACCGACGAGGTAAGC | −3.074 | 111.5 | 0.990 |
| CHS 1 | Chalcone synthase 1 | F: GCAAAGAAGCAGCCACCA R: TTAACGGAAGGGCGAAGG | −3.176 | 106.5 | 0.998 |
| UBQ10 | Ubiquitin10 | F: CCAGAAAGAATCCACGCTAC R: CACGGAGACGAGAACAAGA | −3.128 | 108.8 | 0.998 |
| UBQ | Ubiquitin | F: AAACCCTAACTGGGAAGAC R: TACCACGTAGACGAAGCAC | −3.335 | 99.5 | 0.997 |
| Act1 | Actin1 | F: TCCGTGACATGAAGGAGA R: AGAAATGGCTGGAAGAGG | −3.248 | 103.2 | 0.991 |
| Act100 | Actin100 | F: TTTCCCTTTATGCCAGTG R: TTTCCCGCTCAGCAGTAG | −3.465 | 94.4 | 0.985 |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | F: AACTGATTGGTATGGCTTTC R: CTCGGACTCCTCCTTGAT | −3.193 | 105.7 | 0.997 |
| Rank | Fruits in Different Periods | Different Tissues | ||
|---|---|---|---|---|
| Gene | CV ± SD | Gene | CV ± SD | |
| 1 | UBQ | 1.16 ± 0.24 | His3 | 1.16 ± 0.24 |
| 2 | Act1 | 1.7 ± 0.35 | Act1 | 1.55 ± 0.32 |
| 3 | β-TUB6 | 1.49 ± 0.36 | GAPDH | 1.55 ± 0.34 |
| 4 | GAPDH | 2.62 ± 0.53 | UBQ10 | 2.68 ± 0.51 |
| 5 | His3 | 3.43 ± 0.69 | β-TUB2 | 2.34 ± 0.54 |
| 6 | UBQ10 | 4.44 ± 0.9 | α-TUB | 2.57 ± 0.55 |
| 7 | CHS1 | 5.3 ± 1.19 | CHS1 | 3.08 ± 0.69 |
| 8 | β-TUB2 | 4.74 ± 1.24 | Act100 | 2.76 ± 0.79 |
| 9 | Act100 | 4.91 ± 1.26 | UBQ | 3.92 ± 0.8 |
| 10 | α-TUB | 6.12 ± 1.35 | β-TUB6 | 3.43 ± 0.91 |
| Gene | Full Name and the E.C. Number | Primer Sequence (5′ → 3′) |
|---|---|---|
| DXS | 1-deoxy-D-xylulose-5-phosphate synthase (EC: 2.2.1.7) | F:ACTGCCACCAGAAAACAAA |
| R:GCTACGAACGAGGGAATGA | ||
| DXR | 1-deoxy-D-xylulose-5-phosphate reductoisomerase (EC: 1.1.1.267) | F:TCCATCCGTGAATCTTTG |
| R:GCTGCGTAATCTCGTGCC | ||
| ISPD | 2-C-methyl-D-erythritol4-phosphate cytidylyltransferase (EC: 2.7.7.60) | F:AGGATTTCCATACCCAAGA |
| R:CAATCGGTTGACCCAGAA | ||
| ISPE | 4-diphosphocytidyl-2-C-methyl-D-erythritol kinase (EC: 2.7.1.148) | F:CCACCAACTCCATCTACCT |
| R:CCTGTCTACTTCATCTGCC | ||
| ISPF | 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase (EC: 4.6.1.12) | F:CGTCCAAATCAAAATCACT |
| R:AATCGCATCCACAACACAG | ||
| ISPG | (E)-4-hydroxy-3-methylbut-2-enyl-diphosphate synthase (EC: 1.17.7.1) | F:CAGAAGCACCAGAGGAGGA |
| R:CCAGAACGACGATGAAAAT | ||
| ISPH | 4-hydroxy-3-methylbut-2-en-1-yl diphosphate reductase (EC: 1.17.7.4) | F:AAATCATTCACAACCCGAC |
| R:CCAGACCTTAGACACCCAA | ||
| FPS/GPS | Geranyl pyrophosphate synthetase/Farnesyl pyrophosphate synthetase (EC: 2.5.1.1 2.5.1.10) | F:GGAAAAAATATACGAGGCA |
| R:AGACAGGGCAAGTAGAGCA | ||
| ACAT | acetyl-CoA C-acetyltransferase (EC: 2.3.1.9) | F:CCTGGTGGAAGAGGGAAAC |
| R:GAGCCGCATCAGCAAATC | ||
| HMGCS | hydroxymethylglutaryl-CoA synthase (EC: 2.3.3.10) | F:TGGACATCTACTTCCCTCG |
| R:CACAGTTTCGCTTCCTACT | ||
| HMGCR | hydroxymethylglutaryl-CoA reductase (EC: 1.1.1.34) | F:CGACGAGAACAAAGAGAAG |
| R:GTAAAACGCAACGGAAAAG | ||
| MVK | mevalonate kinase (EC: 2.7.1.36) | F:ACACGAAAGTTGGGAGAA |
| R:TATGGAGGAATGGCTGAC | ||
| MVAK | phosphomevalonate kinase (EC: 2.7.4.2) | F:TCAGAAGTAGCAGAGCCAT |
| R:GTATCAAGAACACGAGCAT | ||
| MVD | diphosphomevalonate decarboxylase (EC:4.1.1.33) | F:TCGTAGTCTTTTTGGTGGA |
| R:TGTATTATGCGTTTCGGTA | ||
| IDI | isopentenyl-diphosphate Delta-isomerase (EC: 5.3.3.2) | F:CAGCCATCCTCTATTCCGC |
| R:ATCAAGTTCATGCTCCCCC | ||
| SQS | squalene synthase (EC: 2.5.1.21) | F:TGACACCAGCATACCTAC |
| R:AATCGCCTCCTGATAACC | ||
| SQE | squalene epoxidase (EC: 1.14.14.17) | F:GGTTCGGTGTCTGGTTGA |
| R:GCTTCTATTTGGCATTGTTT | ||
| CYP450 | cytochromeP450 (1.14.x.x) | F:GGTCAGGTAGACGGGTTT |
| R:TCTCATCCGACGGTAGCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Zhang, Y.; Mu, D.; Lu, Y.; Chen, W.; Zhang, Y.; Zhang, R.; Qin, Y.; Yuan, J.; Pan, L.; et al. Selection of Reference Genes in Evodia rutaecarpa var. officinalis and Expression Patterns of Genes Involved in Its Limonin Biosynthesis. Plants 2023, 12, 3197. https://doi.org/10.3390/plants12183197
Zhou Y, Zhang Y, Mu D, Lu Y, Chen W, Zhang Y, Zhang R, Qin Y, Yuan J, Pan L, et al. Selection of Reference Genes in Evodia rutaecarpa var. officinalis and Expression Patterns of Genes Involved in Its Limonin Biosynthesis. Plants. 2023; 12(18):3197. https://doi.org/10.3390/plants12183197
Chicago/Turabian StyleZhou, Yu, Yuxiang Zhang, Detian Mu, Ying Lu, Wenqiang Chen, Yao Zhang, Ruiying Zhang, Ya Qin, Jianhua Yuan, Limei Pan, and et al. 2023. "Selection of Reference Genes in Evodia rutaecarpa var. officinalis and Expression Patterns of Genes Involved in Its Limonin Biosynthesis" Plants 12, no. 18: 3197. https://doi.org/10.3390/plants12183197