
Antifungal Activity of Glucosinolate-Derived Nitriles and Their Synergistic Activity with Glucosinolate-Derived Isothiocyanates Distinguishes Various Taxa of Brassicaceae Endophytes and Soil Fungi

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Isolation and Taxonomy of Fungi Based on Their ITS Barcode
2.2. Antifungal Potency of Glucosinolate-Derived Nitriles
2.3. Synergism between ITCs and Nitriles
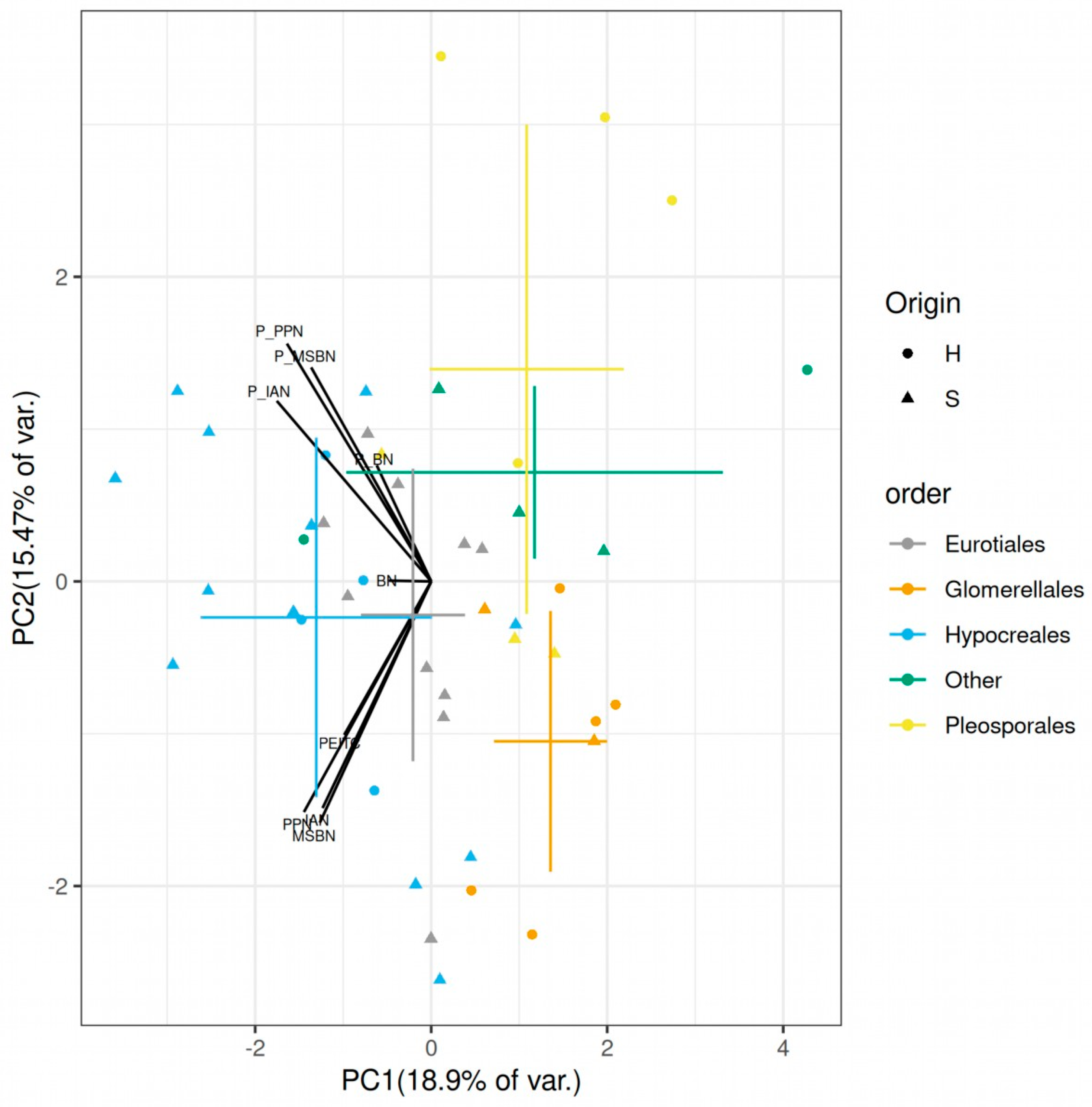
2.4. Differences among Endophytes and Soil Fungi, Fungal Taxa and Possible Microbiome Implications
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Fungi
4.3. Antimicrobial Assay
4.4. Minimum Inhibitory Concentration Measurement
4.5. Synergism Experiments
4.6. Statistical Tests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate Structural Diversity, Identification, Chemical Synthesis and Metabolism in Plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef] [PubMed]
- Abuyusuf, M.; Rubel, M.H.; Kim, H.-T.; Jung, H.-J.; Nou, I.-S.; Park, J.-I. Glucosinolates and Biotic Stress Tolerance in Brassicaceae with Emphasis on Cabbage: A Review. Biochem. Genet. 2023, 61, 451–470. [Google Scholar] [CrossRef] [PubMed]
- Hanschen, F.S.; Lamy, E.; Schreiner, M.; Rohn, S. Reactivity and Stability of Glucosinolates and Their Breakdown Products in Foods. Angew. Chem. Int. Ed. 2014, 53, 11430–11450. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Massih, R.M.; Debs, E.; Othman, L.; Attieh, J.; Cabrerizo, F.M. Glucosinolates, a Natural Chemical Arsenal: More to Tell than the Myrosinase Story. Front. Microbiol. 2023, 14, 1130208. [Google Scholar] [CrossRef] [PubMed]
- Plaszkó, T.; Szűcs, Z.; Vasas, G.; Gonda, S. Interactions of Fungi with Non-Isothiocyanate Products of the Plant Glucosinolate Pathway: A Review on Product Formation, Antifungal Activity, Mode of Action and Biotransformation. Phytochemistry 2022, 200, 113245. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Eugui, D.; Velasco, P. Natural Control of Plant Pathogens through Glucosinolates: An Effective Strategy against Fungi and Oomycetes. Phytochem. Rev. 2020, 19, 1045–1059. [Google Scholar] [CrossRef]
- Plaszkó, T.; Szűcs, Z.; Vasas, G.; Gonda, S. Effects of Glucosinolate-Derived Isothiocyanates on Fungi: A Comprehensive Review on Direct Effects, Mechanisms, Structure-Activity Relationship Data and Possible Agricultural Applications. J. Fungi 2021, 7, 539. [Google Scholar] [CrossRef]
- Eugui, D.; Escobar, C.; Velasco, P.; Poveda, J. Glucosinolates as an Effective Tool in Plant-Parasitic Nematodes Control: Exploiting Natural Plant Defenses. Appl. Soil Ecol. 2022, 176, 104497. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Ahiahonu, P.W.K. Phytotoxin Production and Phytoalexin Elicitation by the Phytopathogenic Fungus Sclerotinia sclerotiorum. J. Chem. Ecol. 2004, 30, 2163–2179. [Google Scholar] [CrossRef]
- Hiruma, K.; Fukunaga, S.; Bednarek, P.; Piślewska-Bednarek, M.; Watanabe, S.; Narusaka, Y.; Shirasu, K.; Takano, Y. Glutathione and Tryptophan Metabolism Are Required for Arabidopsis Immunity during the Hypersensitive Response to Hemibiotrophs. Proc. Natl. Acad. Sci. USA 2013, 110, 9589–9594. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Zheng, Q.-A. Metabolic Responses of Thellungiella Halophila/Salsuginea to Biotic and Abiotic Stresses: Metabolite Profiles and Quantitative Analyses. Phytochemistry 2010, 71, 581–589. [Google Scholar] [CrossRef]
- Frerigmann, H.; Piślewska-Bednarek, M.; Sánchez-Vallet, A.; Molina, A.; Glawischnig, E.; Gigolashvili, T.; Bednarek, P. Regulation of Pathogen-Triggered Tryptophan Metabolism in Arabidopsis Thaliana by MYB Transcription Factors and Indole Glucosinolate Conversion Products. Mol. Plant 2016, 9, 682–695. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Xu, J.; Wang, X.; He, X.; Wang, Y.; Zhou, J.; Zhang, S.; Meng, X. The Arabidopsis Pleiotropic Drug Resistance Transporters PEN3 and PDR12 Mediate Camalexin Secretion for Resistance to Botrytis cinerea. Plant Cell 2019, 31, 2206–2222. [Google Scholar] [CrossRef]
- Kuhn, H.; Lorek, J.; Kwaaitaal, M.; Consonni, C.; Becker, K.; Micali, C.; Ver Loren van Themaat, E.; Bednarek, P.; Raaymakers, T.M.; Appiano, M.; et al. Key Components of Different Plant Defense Pathways are Dispensable for Powdery Mildew Resistance of the Arabidopsis Mlo2 Mlo6 Mlo12 Triple Mutant. Front. Plant Sci. 2017, 8, 1006. [Google Scholar] [CrossRef]
- Yun, H.S.; Kang, B.G.; Kwon, C. Arabidopsis Immune Secretory Pathways to Powdery Mildew Fungi. Plant Signal. Behav. 2016, 11, e1226456. [Google Scholar] [CrossRef] [Green Version]
- Wittstock, U.; Kurzbach, E.; Herfurth, A.-M.; Stauber, E.J. Glucosinolate Breakdown. Adv. Bot. Res. 2016, 80, 125–169. [Google Scholar] [CrossRef]
- Vik, D.; Mitarai, N.; Wulff, N.; Halkier, B.A.; Burow, M. Dynamic Modeling of Indole Glucosinolate Hydrolysis and Its Impact on Auxin Signaling. Front. Plant Sci. 2018, 9, 550. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, R.; Li, R.; Kuwahara, A.; Nakabayashi, R.; Sotta, N.; Mori, T.; Ito, T.; Ohkama-Ohtsu, N.; Fujiwara, T.; Saito, K.; et al. Retrograde Sulfur Flow from Glucosinolates to Cysteine in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2021, 118, e2017890118. [Google Scholar] [CrossRef]
- Ting, H.-M.; Cheah, B.H.; Chen, Y.-C.; Yeh, P.-M.; Cheng, C.-P.; Yeo, F.K.S.; Vie, A.K.; Rohloff, J.; Winge, P.; Bones, A.M.; et al. The Role of a Glucosinolate-Derived Nitrile in Plant Immune Responses. Front. Plant Sci. 2020, 11, 257. [Google Scholar] [CrossRef]
- Witzel, K.; Hanschen, F.S.; Klopsch, R.; Ruppel, S.; Schreiner, M.; Grosch, R. Verticillium Longisporum Infection Induces Organ-Specific Glucosinolate Degradation in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 508. [Google Scholar] [CrossRef] [Green Version]
- Hanschen, F.S.; Pfitzmann, M.; Witzel, K.; Stützel, H.; Schreiner, M.; Zrenner, R. Differences in the Enzymatic Hydrolysis of Glucosinolates Increase the Defense Metabolite Diversity in 19 Arabidopsis thaliana Accessions. Plant Physiol. Biochem. 2018, 124, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, H.; Zhao, Z.; Sheng, X.; Shen, Y.; Gu, H. Natural Variation of Glucosinolates and Their Breakdown Products in Broccoli (Brassica Oleracea Var. Italica) Seeds. J. Agric. Food Chem. 2019, 67, 12528–12537. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium; Fungal Barcoding Consortium Author List; Bolchacova, E.; et al. Nuclear Ribosomal Internal Transcribed Spacer (ITS) Region as a Universal DNA Barcode Marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2017, 45, D37–D42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federhen, S. Type Material in the NCBI Taxonomy Database. Nucleic Acids Res. 2015, 43, D1086–D1098. [Google Scholar] [CrossRef] [Green Version]
- Xu, J. Fungal DNA Barcoding. Genome 2016, 59, 913–932. [Google Scholar] [CrossRef] [Green Version]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-Scale Generation and Analysis of Filamentous Fungal DNA Barcodes Boosts Coverage for Kingdom Fungi and Reveals Thresholds for Fungal Species and Higher Taxon Delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Plaszkó, T.; Szűcs, Z.; Cziáky, Z.; Ács-Szabó, L.; Csoma, H.; Géczi, L.; Vasas, G.; Gonda, S. Correlations Between the Metabolome and the Endophytic Fungal Metagenome Suggests Importance of Various Metabolite Classes in Community Assembly in Horseradish (Armoracia rusticana, Brassicaceae) Roots. Front. Plant Sci. 2022, 13, 921008. [Google Scholar] [CrossRef]
- Toju, H.; Kurokawa, H.; Kenta, T. Factors Influencing Leaf- and Root-Associated Communities of Bacteria and Fungi across 33 Plant Orders in a Grassland. Front. Microbiol. 2019, 10, 241. [Google Scholar] [CrossRef] [Green Version]
- Shofran, B.G.; Purrington, S.T.; Breidt, F.; Fleming, H.P. Antimicrobial Properties of Sinigrin and Its Hydrolysis Products. J. Food Sci. 1998, 63, 621–624. [Google Scholar] [CrossRef]
- Szűcs, Z.; Plaszkó, T.; Cziáky, Z.; Kiss-Szikszai, A.; Emri, T.; Bertóti, R.; Sinka, L.T.; Vasas, G.; Gonda, S. Endophytic Fungi from the Roots of Horseradish (Armoracia rusticana) and Their Interactions with the Defensive Metabolites of the Glucosinolate—Myrosinase—Isothiocyanate System. BMC Plant Biol. 2018, 18, 85. [Google Scholar] [CrossRef] [Green Version]
- Odds, F.C. Synergy, Antagonism, and What the Chequerboard Puts between Them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and Antagonism in Natural Product Extracts: When 1 + 1 Does Not Equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Yuan, K.; Zhou, Q.; Lu, C.; Du, L.; Liu, F. Mechanism of Antifungal Activity of Perilla Frutescens Essential Oil against Aspergillus Flavus by Transcriptomic Analysis. Food Control 2021, 123, 107703. [Google Scholar] [CrossRef]
- Wu, T.-L.; Zhang, B.-Q.; Luo, X.-F.; Li, A.-P.; Zhang, S.-Y.; An, J.-X.; Zhang, Z.-J.; Liu, Y.-Q. Antifungal Efficacy of Sixty Essential Oils and Mechanism of Oregano Essential Oil against Rhizoctonia Solani. Ind. Crops Prod. 2023, 191, 115975. [Google Scholar] [CrossRef]
- Clemente, I.; Aznar, M.; Nerín, C. Synergistic Properties of Mustard and Cinnamon Essential Oils for the Inactivation of Foodborne Moulds in Vitro and on Spanish Bread. Int. J. Food Microbiol. 2019, 298, 44–50. [Google Scholar] [CrossRef]
- Liu, Q.; Niu, H.; Zhang, W.; Mu, H.; Sun, C.; Duan, J. Synergy among Thymol, Eugenol, Berberine, Cinnamaldehyde and Streptomycin against Planktonic and Biofilm-Associated Food-Borne Pathogens. Lett. Appl. Microbiol. 2015, 60, 421–430. [Google Scholar] [CrossRef]
- Tunc, S.; Chollet, E.; Chalier, P.; Preziosi-Belloy, L.; Gontard, N. Combined Effect of Volatile Antimicrobial Agents on the Growth of Penicillium Notatum. Int. J. Food Microbiol. 2007, 113, 263–270. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, X.; Zhang, G.-A.; Zeng, S.-Y.; Lin, K.-C. Antifungal Vapour-Phase Activity of a Combination of Allyl Isothiocyanate and Ethyl Isothiocyanate Against Botrytis Cinerea and Penicillium Expansum Infection on Apples. J. Phytopathol. 2011, 159, 450–455. [Google Scholar] [CrossRef]
- Popović, M.; Maravić, A.; Čulić, V.Č.; Đulović, A.; Burčul, F.; Blažević, I. Biological Effects of Glucosinolate Degradation Products from Horseradish: A Horse That Wins the Race. Biomolecules 2020, 10, 343. [Google Scholar] [CrossRef] [Green Version]
- Andini, S.; Araya-Cloutier, C.; Waardenburg, L.; den Besten, H.M.W.; Vincken, J.-P. The Interplay between Antimicrobial Activity and Reactivity of Isothiocyanates. LWT 2020, 134, 109843. [Google Scholar] [CrossRef]
- Van Wambeke, E.; Ceustermans, A.; De Landtsheer, A.; Coosemans, J. Combinations of Soil Fumigants for Methyl-Bromide Replacement. Commun. Agric. Appl. Biol. Sci. 2009, 74, 75–84. [Google Scholar]
- Hiruma, K.; Onozawa-Komori, M.; Takahashi, F.; Asakura, M.; Bednarek, P.; Okuno, T.; Schulze-Lefert, P.; Takanoa, Y. Entry Mode-Dependent Function of an Indole Glucosinolate Pathway in Arabidopsis for Nonhost Resistance against Anthracnose Pathogens. Plant Cell 2010, 22, 2429–2443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipka, U.; Fuchs, R.; Kuhns, C.; Petutschnig, E.; Lipka, V. Live and Let Die—Arabidopsis Nonhost Resistance to Powdery Mildews. Eur. J. Cell Biol. 2010, 89, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, R.; Kopischke, M.; Klapprodt, C.; Hause, G.; Meyer, A.J.; Schwarzländer, M.; Fricker, M.D.; Lipka, V. Immobilized Subpopulations of Leaf Epidermal Mitochondria Mediate PENETRATION2-Dependent Pathogen Entry Control in Arabidopsis. Plant Cell 2016, 28, 130–145. [Google Scholar] [CrossRef] [Green Version]
- Lambrix, V.; Reichelt, M.; Mitchell-Olds, T.; Kliebenstein, D.J.; Gershenzon, J. The Arabidopsis Epithiospecifier Protein Promotes the Hydrolysis of Glucosinolates to Nitriles and Influences Trichoplusia Ni Herbivory. Plant Cell 2001, 13, 2793–2808. [Google Scholar] [CrossRef] [Green Version]
- Maciá-Vicente, J.G.; Nam, B.; Thines, M. Root Filtering, Rather than Host Identity or Age, Determines the Composition of Root-Associated Fungi and Oomycetes in Three Naturally Co-Occurring Brassicaceae. Soil Biol. Biochem. 2020, 146, 107806. [Google Scholar] [CrossRef]
- Ishimoto, H.; Fukushi, Y.; Yoshida, T.; Tahara, S. Rhizopus and Fusarium are Selected as Dominant Fungal Genera in Rhizospheres of Brassicaceae. J. Chem. Ecol. 2000, 26, 2387–2399. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, X. Effects of Green Manures on Rhizosphere Fungal Community Composition of Cucumber Seedlings. Curr. Microbiol. 2023, 80, 87. [Google Scholar] [CrossRef]
- Kudjordjie, E.N.; Hooshmand, K.; Sapkota, R.; Darbani, B.; Fomsgaard, I.S.; Nicolaisen, M. Fusarium Oxysporum Disrupts Microbiome-Metabolome Networks in Arabidopsis Thaliana Roots. Microbiol. Spectr. 2022, 10, e0122622. [Google Scholar] [CrossRef]
- Plaszkó, T.; Szűcs, Z.; Kállai, Z.; Csoma, H.; Vasas, G.; Gonda, S. Volatile Organic Compounds (VOCs) of Endophytic Fungi Growing on Extracts of the Host, Horseradish (Armoracia rusticana). Metabolites 2020, 10, 451. [Google Scholar] [CrossRef]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-Coverage ITS Primers for the DNA-Based Identification of Ascomycetes and Basidiomycetes in Environmental Samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [Green Version]
- Gómara, M.; Ramón-García, S. The FICI Paradigm: Correcting Flaws in Antimicrobial in Vitro Synergy Screens at Their Inception. Biochem. Pharmacol. 2019, 163, 299–307. [Google Scholar] [CrossRef]
- Houdkova, M.; Kokoska, L. Volatile Antimicrobial Agents and in Vitro Methods for Evaluating Their Activity in the Vapour Phase: A Review. Planta Medica 2020, 86, 822–857. [Google Scholar] [CrossRef]
- Inouye, S.; Takizawa, T.; Yamaguchi, H. Antibacterial Activity of Essential Oils and Their Major Constituents against Respiratory Tract Pathogens by Gaseous Contact. J. Antimicrob. Chemother. 2001, 47, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Nakahara, K.; Alzoreky, N.S.; Yoshihashi, T.; Nguyen, H.T.; Trakoontivakorn, G. Chemical Composition and Antifungal Activity of Essential Oil from Cymbopogon nardus (Citronella Grass). Jpn. Agric. Res. Q. JARQ 2013, 37, 249–252. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.-S.; Beuchat, L.R.; Kim, H.; Ryu, J.-H. Development of an Experimental Apparatus and Protocol for Determining Antimicrobial Activities of Gaseous Plant Essential Oils. Int. J. Food Microbiol. 2015, 215, 95–100. [Google Scholar] [CrossRef]
- Houdkova, M.; Rondevaldova, J.; Doskocil, I.; Kokoska, L. Evaluation of Antibacterial Potential and Toxicity of Plant Volatile Compounds Using New Broth Microdilution Volatilization Method and Modified MTT Assay. Fitoterapia 2017, 118, 56–62. [Google Scholar] [CrossRef]
- Ji, H.; Kim, H.; Beuchat, L.R.; Ryu, J.-H. Synergistic Antimicrobial Activities of Essential Oil Vapours against Penicillium corylophilum on a Laboratory Medium and Beef Jerky. Int. J. Food Microbiol. 2019, 291, 104–110. [Google Scholar] [CrossRef]
- Bonapace, C.R.; Bosso, J.A.; Friedrich, L.V.; White, R.L. Comparison of Methods of Interpretation of Checkerboard Synergy Testing. Diagn. Microbiol. Infect. Dis. 2002, 44, 363–366. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; ISBN 3-900051-07-0. [Google Scholar]



{kind=link}
{kind=link}
| ID | Genus | Order | Origin | MIC (mM) | FICI (PEITC + Nitrile) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PEITC | IAN | PPN | MSBN | BN | IAN | PPN | MSBN | BN | ||||
| F1 | Fusarium | Hy | H | 0.0536 | 2.56 | 6.11 | 27 | 79.6 | 0.56 | 0.75 | 0.56 | 0.63 |
| F2 | Fusarium | Hy | H | 0.0804 | 1.28 | 6.11 | 27 | 44.8 | 0.63 | 0.63 | 0.63 | 1.06 |
| F3 | Phoma | Pl | H | 0.0042 | 0.32 | 2.29 | 13.5 | 23.2 | 0.63 | 0.5 | 0.5 | 0.56 |
| F4 | Phoma | Pl | H | 0.0067 | 0.43 | 5.09 | 11.3 | 101.9 | 0.63 | 0.75 | 0.75 | 0.75 |
| F5 | Oidiodendron | - | H | 0.0050 | 0.27 | 1.27 | 15.8 | 49.7 | 0.38 | 0.5 | 0.25 | 0.5 |
| F6 | Fusarium | Hy | H | 0.0536 | 1.28 | 6.11 | 36 | 29.8 | 0.54 | 0.81 | 0.44 | 0.69 |
| F7 | Fusarium | Hy | H | 0.0804 | 1.28 | 6.11 | 40.5 | 101.9 | 0.56 | 0.5 | 0.5 | 0.52 |
| F8 | Paraphoma | Pl | H | 0.0134 | 1.28 | 4.07 | 27 | 59.7 | 0.44 | 0.56 | 0.63 | 0.63 |
| F9 | Plectosphaerella | Gl | H | 0.0402 | 1.28 | 4.58 | 27 | 79.6 | 0.44 | 0.38 | 0.38 | 0.5 |
| F10 | Plectosphaerella | Gl | H | 0.0268 | 1.71 | 5.09 | 20.3 | 46.4 | 0.38 | 0.44 | 0.38 | 0.5 |
| F11 | Pseudopyrenochaeta | Pl | H | 0.0067 | 0.43 | 2.54 | 13.5 | 41.4 | 0.5 | 0.56 | 0.75 | 0.69 |
| F12 | Plectosphaerella | Gl | H | 0.0268 | 1.92 | 8.14 | 27 | 53 | 0.38 | 0.38 | 0.44 | 0.31 |
| F13 | Phomopsis | - | H | 0.0201 | 1.28 | 9.16 | 27 | 79.6 | 0.69 | 0.63 | 0.63 | 0.69 |
| F14 | Plectosphaerella | Gl | H | 0.0179 | 1.49 | 4.07 | 27 | 59.7 | 0.46 | 0.46 | 0.46 | 0.67 |
| F15 | Plectosphaerella | Gl | H | 0.0313 | 1.71 | 7.12 | 36 | 79.6 | 0.5 | 0.31 | 0.46 | 0.54 |
| F16 | Stagonosporopsis | Pl | S | 0.0536 | 0.64 | 6.11 | 27 | 49.7 | 0.44 | 0.44 | 0.5 | 0.5 |
| F17 | Curvularia | Pl | S | 0.0201 | 1.71 | 6.11 | 27 | 56.4 | 0.38 | 0.56 | 1 | 0.69 |
| F18 | Penicillium | Eu | S | 0.0804 | 1.28 | 6.11 | 27 | 79.6 | 0.81 | 0.56 | 0.56 | 0.63 |
| F19 | Aspergillus | Eu | S | 0.0201 | 2.99 | 8.14 | 29.3 | 5 | 0.44 | 0.38 | 0.56 | 0.56 |
| F20 | Fusarium | Hy | S | 0.0536 | 1.28 | 6.11 | 36 | 66.3 | 0.63 | 0.56 | 0.81 | 0.63 |
| F21 | Fusarium | Hy | S | 0.1608 | 1.92 | 6.11 | 27 | 44.8 | 0.38 | 0.38 | 0.38 | 1.25 |
| F22 | Penicillium | Eu | S | 0.0201 | 1.92 | 4.58 | 40.5 | 44.8 | 0.56 | 0.44 | 0.56 | 0.56 |
| F23 | Fusarium | Hy | S | 0.0402 | 1.92 | 6.11 | 54 | 79.6 | 0.69 | 0.81 | 0.63 | 0.63 |
| F24 | Penicillium | Eu | S | 0.0402 | 1.28 | 6.11 | 27 | 19.9 | 0.69 | 0.69 | 0.63 | 0.63 |
| F25 | Penicillium | Eu | S | 0.0201 | 1.28 | 6.11 | 27 | 59.7 | 0.5 | 0.5 | 0.63 | 0.69 |
| F26 | Fusarium | Hy | S | 0.0357 | 1.28 | 9.16 | 40.5 | 82 | 0.44 | 0.38 | 0.44 | 0.47 |
| F27 | Penicillium | Eu | S | 0.0536 | 1.92 | 6.11 | 27 | 79.6 | 0.69 | 0.56 | 0.56 | 0.63 |
| F28 | Penicillium | Eu | S | 0.0447 | 1.28 | 6.11 | 36 | 26.5 | 0.58 | 0.56 | 0.5 | 0.56 |
| F29 | Fusarium | Hy | S | 0.1340 | 1.28 | 6.11 | 27 | 79.6 | 0.44 | 0.44 | 0.44 | 0.44 |
| F30 | Penicillium | Eu | S | 0.0134 | 1.28 | 5.09 | 36 | 44.8 | 0.63 | 0.63 | 0.63 | 0.63 |
| F31 | Cadophora | - | S | 0.0201 | 1.28 | 4.07 | 22.5 | 44.8 | 0.44 | 0.44 | 0.5 | 0.56 |
| F32 | Clonostachys | Hy | S | 0.0402 | 1.92 | 4.58 | 20.3 | 101.9 | 0.63 | 0.69 | 0.69 | 0.57 |
| F33 | Plectosphaerella | Gl | S | 0.0402 | 0.96 | 6.11 | 27 | 49.7 | 0.38 | 0.38 | 0.5 | 0.38 |
| F34 | Cladosporium | - | S | 0.0201 | 0.64 | 6.11 | 27 | 43.1 | 0.56 | 0.44 | 0.5 | 0.81 |
| F35 | Fusarium | Hy | S | 0.0804 | 1.92 | 6.11 | 27 | 79.6 | 0.81 | 0.81 | 0.75 | 0.69 |
| F36 | Fusarium | Hy | S | 0.0804 | 1.92 | 6.11 | 27 | 59.7 | 0.81 | 0.69 | 0.88 | 0.5 |
| F37 | Penicillium | Eu | S | 0.0536 | 1.28 | 4.58 | 27 | 29.8 | 0.63 | 0.44 | 0.63 | 0.5 |
| F38 | Penicillium | Eu | S | 0.0536 | 1.92 | 6.11 | 27 | 19.9 | 0.56 | 0.44 | 0.56 | 0.63 |
| F39 | Plectosphaerella | Gl | S | 0.0402 | 1.28 | 6.11 | 27 | 14.9 | 0.56 | 0.5 | 0.5 | 0.63 |
| F40 | Cladosporium | - | S | 0.0268 | 0.64 | 6.11 | 27 | 36.5 | 0.63 | 0.5 | 0.63 | 1 |
| F41 | Purpureocillium | Hy | S | 0.0313 | 1.28 | 6.11 | 27 | 14.9 | 0.5 | 0.44 | 0.5 | 0.75 |
| F42 | Clonostachys | Hy | S | 0.0626 | 2.56 | 6.11 | 27 | 59.7 | 0.56 | 0.63 | 0.81 | 0.5 |
| F43 | Aaosphaeria | Pl | S | 0.0201 | 1.28 | 6.11 | 27 | 79.6 | 0.38 | 0.56 | 0.56 | 0.38 |
| F44 | Fusarium | Hy | S | 0.0357 | 2.13 | 8.14 | 36 | 26.5 | 0.69 | 0.75 | 0.69 | 0.75 |
| F45 | Clonostachys | Hy | S | 0.0626 | 1.28 | 9.16 | 36 | 39.8 | 1 | 0.81 | 0.63 | 0.69 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szűcs, Z.; Plaszkó, T.; Bódor, E.; Csoma, H.; Ács-Szabó, L.; Kiss-Szikszai, A.; Vasas, G.; Gonda, S. Antifungal Activity of Glucosinolate-Derived Nitriles and Their Synergistic Activity with Glucosinolate-Derived Isothiocyanates Distinguishes Various Taxa of Brassicaceae Endophytes and Soil Fungi. Plants 2023, 12, 2741. https://doi.org/10.3390/plants12142741
Szűcs Z, Plaszkó T, Bódor E, Csoma H, Ács-Szabó L, Kiss-Szikszai A, Vasas G, Gonda S. Antifungal Activity of Glucosinolate-Derived Nitriles and Their Synergistic Activity with Glucosinolate-Derived Isothiocyanates Distinguishes Various Taxa of Brassicaceae Endophytes and Soil Fungi. Plants. 2023; 12(14):2741. https://doi.org/10.3390/plants12142741
Chicago/Turabian StyleSzűcs, Zsolt, Tamás Plaszkó, Eszter Bódor, Hajnalka Csoma, Lajos Ács-Szabó, Attila Kiss-Szikszai, Gábor Vasas, and Sándor Gonda. 2023. "Antifungal Activity of Glucosinolate-Derived Nitriles and Their Synergistic Activity with Glucosinolate-Derived Isothiocyanates Distinguishes Various Taxa of Brassicaceae Endophytes and Soil Fungi" Plants 12, no. 14: 2741. https://doi.org/10.3390/plants12142741