
Comparative Evaluation of Antioxidant Status and Mineral Composition of Diploschistes ocellatus, Calvatia candida (rostk.) Hollós, Battarrea phalloides and Artemisia lerchiana in Conditions of High Soil Salinity

 , , , and
, , , and
Abstract
:1. Introduction
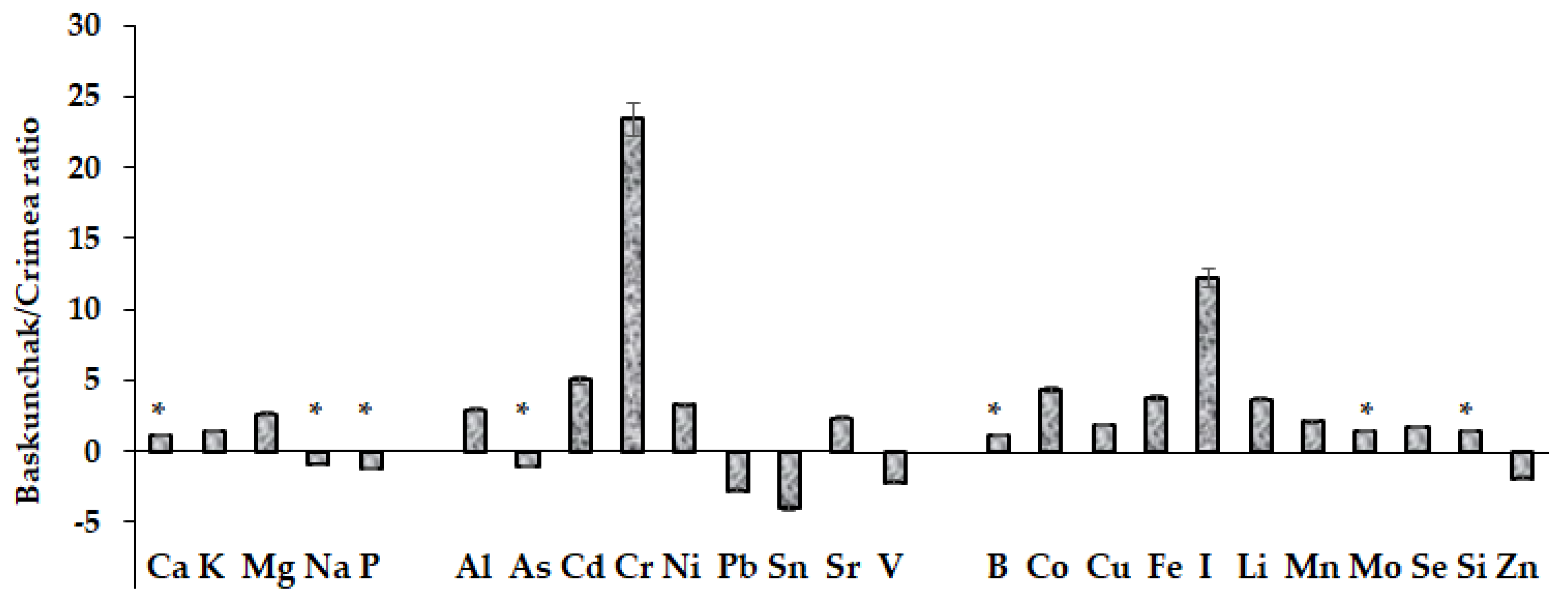
2. Results and Discussion
2.1. Mineral Composition
2.2. Antioxidants
2.3. Proline
2.4. Malonic dialdehyde (MDA)
2.5. Carbohydrates
2.6. Photosynthetic Pigments
3. Material and Methods
3.1. Mineral Composition
3.2. Total Polyphenols (TP)
3.3. Antioxidant Activity (AOA)
3.4. Proline
3.5. Malonic Dialdehyde (MDA)
3.6. Carbohydrates
3.7. Photosynthetic Pigments
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Volobaeva, O.V. Bogdinsko-Baskunchak Nature Reserve Flora. Ph.D. Thesis, Bashkir State University, Ufa, Russia, 2021. (In Russian). [Google Scholar]
- Chuicov, J.S. Bogdinsko-Baskunchak Nature Reserve Complex and Its Protection; OOO CNTEP: Astrakhan, Russia, 1998. (In Russian) [Google Scholar]
- Golubkina, N.; Logvinenko, L.; Molchanova, A.; Caruso, G. Genetic and Environmental Influence on Macro- and Micro-elements Accumulation in Plants of Artemisia Species. In Plant Micronutrients: Deficiency and Toxicity Management; Aftab, T., Ed.; Chapter 17; Springer: Berlin, Germany, 2020. [Google Scholar]
- Orlova, J.V. Ecophysiological Characteristics of Artemisia lerchiana Web in conditions of Low Volga region. Ph.D. Thesis, Moscow, Russia, 2009. (In Russian). [Google Scholar]
- Purvis, O.W.; Halls, S. A review of lichens in metal-enriched environments. Lichenologist 1996, 28, 571–601. [Google Scholar] [CrossRef]
- Evans, S. Conservation corner. Field Mycol. 2001, 2, 116–117. [Google Scholar] [CrossRef]
- Marren, P. Stinkhorns, earthstars and fungal flowers: The strange world gasteromycete fungi. Br. Wildl. 1995, 6, 366–376. [Google Scholar]
- Gargano, M.L.; Venturella, G.; Ferraro, V. Is Battarrea phalloides really an endangered species? Plant Biosyst.-Int. J. Plant Biol. (Off. J. Soc. Bot. Ital.) 2020, 155, 759–762. [Google Scholar]
- Hedawoo, G.B. Calvatia species: Wild edible puff balls from Armavati region (M.S.). Plantae Sci. 2020, 3, 30–34. [Google Scholar] [CrossRef]
- Fernández-Brime, S.; Llimona, X.; Lutzoni, F.; Gaya, E. Phylogenetic study of Diploschistes (lichen-forming Ascomycota: Ostropales: Graphidaceae), based on morphological, chemical, and molecular data. Taxon 2013, 62, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Mendili, M.; Bannour, M.; Araújo, M.E.M.; Aschi-Smit, S.; Seaward, M.R.D.; Khadhria, A. Secondary metabolites and antioxidant capacity of the Tunisian lichen Diploschistes ocellatus (Ascomycot a). Int.J. Med. Mushrooms 2019, 19, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Golubkina, N.A.; Kharchenko, V.A.; Caruso, G. Selenium: Prospects of functional food production with high antioxidant activity Reference Series in Phyto-chemistry. In Plant Antioxidants and Health; Ekiert, H., Ramawat, K.G., Arora, J., Eds.; Springer: Berlin, Germany, 2021. [Google Scholar] [CrossRef]
- Kalač, P. A review of chemical composition and nutritional value of wild-growing and cultivated mushrooms. J. Sci. Food Agric. 2013, 93, 209–218. [Google Scholar] [CrossRef]
- Delmail, D.; Grube, M.; Parrot, D.; Cook-Moreau, J.; Boustie, J.; Labrousse, P.; Tomasi, S. Halotolerance in lichens: Symbiotic coalition against salt stress. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Ed.; Springer Science + Business Media, LLC: New York, NY, USA, 2013; p. 115. [Google Scholar] [CrossRef]
- Piterance, A.V. Diploschistaceae family. Key of lichens of the USSR. Calcium–Gialectic 1975, 3, 45–53. (In Russian) [Google Scholar]
- Weber, W.A. Environmental modifications and the taxonomy of the crustose lichens. Svensk. Botanisk Tidskrift. 1962, 56, 293–333. [Google Scholar]
- Wadsten, T.; Moberg, R. Calcium oxalate hydrates on the surface of lichens. Lichenologist 1985, 17, 239–245. [Google Scholar] [CrossRef]
- Ručová, D.; Đorđević, T.; Baláž, M.; Weidinger, M.; Lang, I.; Gajdoš, A.; Goga, M.l. Investigation of calcium forms in lichens from Travertine sites. Plants 2022, 11, 620. [Google Scholar] [CrossRef] [PubMed]
- Cavigia, A.M.; Modenesi, P. Oxidative stress and ascorbic contents in Parmotrema reticulatum and Parmelia sulcata thalli. Lichenologist 1999, 31, 105–110. [Google Scholar] [CrossRef]
- Modenesi, P.; Piana, M.; Giordani, P.; Tafanelli, A.; Bartoli, A. Calcium oxalate and medullary architecture in Xanthomaculina convolute. Lichenologist 2000, 32, 505–512. [Google Scholar] [CrossRef]
- Modenesi, P.; Bombardi, V.; Giordani, P.; Brunialti, G.; Carallo, A. Dissolution of weddellite, calcium oxalate dehydrate, in Pyxine subcinerea. Lichenologist 2001, 33, 261–266. [Google Scholar] [CrossRef]
- Rosentsvet, O.A.; Nesterov, V.N.; Bogdanova, E.C.; Tabalenkova, G.N.; Zakhogy, I.G.; Popov, A.V. Effect of Ellton salinity soils on functional conditions of Artemisia representatives. Izv. RAS Biol. Ser. 2019, 3, 312–320. (In Russian) [Google Scholar]
- Golubkina, N.A.; Mironov, J.V. Element composition of mushrooms in contrasting anthropogenic loading. Geochem. Int. 2018, 56, 1263–1275. [Google Scholar]
- Falandysz, J. Selenium in edible mushrooms. J. Environ. Sci. Health Part C 2008, 26, 256–299. [Google Scholar] [CrossRef] [Green Version]
- Nöeske, O.; Läuchli, A.; Lange, O.L.; Vieweg, G.H.; Ziegler, H. Konzentration und Lokalisierung von Schwermetallen in Flechten der Erzschlackenhalden des Harzes. Vortr. Gesamtgeb. Ber. Dtsch. Bot. Ges. 1970, 4, 67–79. [Google Scholar]
- Lawrey, J.D.; Rudolph, E.D. Lichen accumulation of some heavy metals from acids surface substrates of coal mine ecosystems in southeastern Ohio. Ohio J. Sci. 1975, 75, 113–117. [Google Scholar]
- Purvis, O.W.; Elix, J.A.; Gaul, K.L. The occurrence of copper-norstictic acid in lichens from cupriferous substrata. Lichenologist 1990, 22, 345–354. [Google Scholar] [CrossRef]
- Mikkelsen, R.L.; Page, A.L.; Haghnia, G.H. Effect of salinity and its composition on the accumulation of selenium by alfalfa. Plant Soil 1988, 107, 63–67. [Google Scholar] [CrossRef]
- Alejandro, S.; Holler, S.; Meier, B.; Peiter, E. Manganese in plants: From acquisition to subcellular allocation. Front. Plant Sci. 2020, 11, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corapi, A.; Gallo, L.; Nicolardi, V.; Lucadamo, L.; Loppi, S. Temporal trends of element concentrations and ecophysiological parametrsin in the lichen Pseudevernia furfuracea transplanted in and around in industrial area S Italy. Environ. Monit. Assess. 2014, 186, 3149–3164. [Google Scholar] [CrossRef]
- Byazrov, L.G. Lichens in Ecological Monitoring; Nauchny Mir: Moscow, Russia, 2002; 336p. (In Russian) [Google Scholar]
- Bennett, J.P. Discrimination of lichen genera and species using element concentrations. Lichenologist 2008, 40, 135–151. [Google Scholar] [CrossRef]
- Nash, T.H., III. Nutrients, elemental accumulation, and mineral cycling. In Lichen Biology; Cambridge University Press: Cambridge, UK, 2010; pp. 236–253. [Google Scholar] [CrossRef]
- Insarova, I.D. Heavy metals effect on lichens. Problems of ecological monitoring and ecosystems modeling. Leningrad 1983, 6, 101–113. (In Russian) [Google Scholar]
- Benson, T.R.; Coble, M.A.; Rytuba, J.J.; Mahood, G.A. Lithium enrichment in intracontinental rhyolite magmas leads to Li deposits in caldera basins. Nat. Commun. 2017, 8, 270. [Google Scholar] [CrossRef] [PubMed]
- Winkel, L.H.E.; Vriens, B.; Jones, G.D.; Schneider, L.S.; Pilon-Smits, E.; Bañuelos, G.S. Selenium cycling across soil-plant-atmosphere interfaces: A critical review. Nutrients 2015, 7, 4199–4239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golubkina, N.; Lapchenko, V.; Lapchenko, H.; Zuev, A.; Caruso, G. Seasonal dynamics of groundwater mineral composition at the territory at the Crimean paleovolcano. Bull. Nikita Bot. Gard. 2022, 142, 129–135. [Google Scholar]
- Osyczka, P.; Latkowska, E.; Rola, K. Metabolic processes involved with sugar alcohol and secondary metabolite production in the hyperaccumulator lichen Diploschistes muscorum reveal its complex adaptation strategy against heavy-metal stress. Fungal Biol. 2021, 125, 999–1008. [Google Scholar] [CrossRef]
- Sarret, G.; Manceau, A.; Cuny, D.; van Haluwyn, C.; Déruelle, S.; Hazemann, J.-L.; Soldo, Y.; Eybert-Bŭrard, L.; Menthonnex, J.-J. Mechanisms of lichen resistance to metallic pollution. Environ. Sci. Technol. 1998, 32, 3325–3330. [Google Scholar] [CrossRef]
- Kosanić, M.; Ranković, B. Studies on antioxidant properties of lichen secondary metabolites. In Lichen Secondary Metabolites; Ranković, B., Ed.; Springer: New York, NY, USA, 2019. [Google Scholar] [CrossRef]
- Novaković, A.R.; Karaman, M.A.; Matavulj, M.N.; Pejin, B.M.; Belović, M.M.; Radusin, T.I.; Ilić, N.M. An insight into in vitro bioactivity of wild-growing puffball species Lycoperdon perlatum (Pers) 1796. Food Feed Res. 2015, 42, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Román, M.P.G.; Mantilla, N.B.; Flórez, S.A.C.; De Mandal, S.; Passari, A.K.; Ruiz-Villáfan, B.; Rodríguez-Sanoja, R.; Sánchez, S. Chapter Antimicrobial and antioxidant potential of wild edible mushrooms. In An Introduction to Mushroom; Intechopen: London, UK, 2020. [Google Scholar] [CrossRef] [Green Version]
- Golubkina, N.A.; Kekina, H.G.; Molchanova, A.V.; Antoshkina, M.S.; Nadezhkin, S.; Soldatenko, A.V. Plants Antioxidants and Methods of Their Determination; Infra-M: Moscow, Russia, 2020; (In Russian). [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S. An overview of selenium uptake, metabolism and toxicity in plants. Front. Plant Sci. 2017, 7, 2074. [Google Scholar] [CrossRef] [Green Version]
- Lanza, M.G.D.B.; dos Reis, A.R. Roles of selenium in mineral plant nutrition: ROS scavenging responses against abiotic stresses. Plant Physiol. Biochem. 2021, 164, 27–43. [Google Scholar] [CrossRef]
- Ouertani, R.N.; Abid, G.; Karmous, C.; Chikha, M.B.; Boudaya, O.; Mahmoudi, H.; Mejri, S.; Jansen, K.; Ghorbel, A. Evaluating the contribution of osmotic and oxidative stress components on barley growth under salt stress. AoB Plants 2021, 13, plab034. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. A review. Plant Signal Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, R.; Kunkiwska, A.B.; Schippers, J.H.M. Role of reactive oxygen species during cell expansion in leaves. Plant Physiol. 2016, 172, 2098–2106. [Google Scholar] [CrossRef] [Green Version]
- Shmakova, N.Y.; Markovskaya, E.F. Photosynthetic pigments of plants and lichens in Arctic tundra of west Spitsbergen. Rus. J. Plant Physiol. 2010, 57, 764–769. [Google Scholar] [CrossRef]
- Hinojosa-Vidal, E.; Marco, F.; Martínez_Alberola, F.; Escaray, F.J.; García-Breijo, F.J.; Reid-Armińana, J.; Carrasco, P.; Barreno, E. Characterization of the Responses to Saline Stress in the Symbiotic Green Microalga Treboixia sp. TR9. Planta 2018, 1, 14. [Google Scholar] [CrossRef]
- Green, T.G.A.; Nash, T.H., III; Lange, O.L. Physiological ecology of carbon dioxide exchange. In Lichen Biology; Chapter 9; Nash, T.H., III, Ed.; Cambridge University Press: Cambridge, UK, 2008; pp. 152–181. [Google Scholar] [CrossRef]
- Pintado, A.; Sancho, L.G.; Green, T.G.A.; Blanquer, J.M.; Lázaro, R. Function ecology of the biological soil crust in semiarid SE Spain: Sun and shade populations of Diploshistes diacapsis (Ach.) Lumbisch. Lichenologist 2005, 37, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Lange, O.L.; Green, T.G.A.; Reichenberger, H.; Chaidir, C. Do secondary substances in the thallus of a lichen promote CO2 diffusion and prevent depression of net photosynthesis at high water content? Oecologia 1997, 112, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Lange, O.L.; Green, T.G.A.; Reichenberger, H. The response of lichen photosynthesis to external CO2 concentration and its interaction with thallus water-status. J. Plant Physiol. 1999, 154, 157–166. [Google Scholar] [CrossRef]
- Golubkina, N.; Antoshkina, M.; Bondareva, L.; Sekara, A.; Campagna, E.; Caruso, G. Effect of foliar application of sodium selenate on mineral relationships in Brassicaceae crops. Horticulturae 2023, 9, 535. [Google Scholar] [CrossRef]
- Heath, R.L.; Parker, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Swamy, P.M. Laboratory Manual on Biotechnology; Rastogi: New Delhi, Rastogi Publications, New Delhi, India, 2008; pp. 101–104. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic bio-membranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Biological Effect | Biologically Active Compounds | |
|---|---|---|---|
| Calvatia species | Terrestrial saprophytes | Antibacterial, antiviral, antitumor and wound healing properties [9] | Calvatin and calvatic acid |
| Diploschistes ocellatus | Symbiotic organism of fungi and algae Trebouxia, calcephitis [10] | Photo-protective, anti-microbial to gram-positive and gram-negative bacteria, anti-fungal to Enterococcus faecalis [11] | Polyphenols, flavonoids |
| Artemisia lerchiana | Haloxerophitis with high tolerance to water deficiency and soil salinity | Allopathic, fungicidal, bactericidal and antioxidant [4] | Essential oil (camphor derivatives) |
| Element | Diplishistes ocellatus | Battarrea phalloides | Calvatia candida | Artemisia lerchiana | ||||
|---|---|---|---|---|---|---|---|---|
| Baskunchak | Crimea | Baskunchak | Crimea | Baskunchak | Crimea | Baskunchak | Crimea | |
| Ca | 114,617 ± 11,210 a | 100,656 ± 10,006 a | 10,014 ± 1000 c | 1020 ± 100 e | 3596 ± 317 d | 850 ± 82 e | 9728 ± 934 c | 13,600 ± 133 b |
| K | 1389 ± 129 e | 987 ± 96 f | 8044 ± 776 d | 34,406 ± 3101 a | 22,543 ± 2052 c | 26,754 ± 2588 bc | 22,306 ± 2180 c | 28,195 ± 2800 b |
| Mg | 934 ± 93 d | 352 ± 32 f | 3052 ± 298 a | 952 ± 93 d | 1514 ± 148 c | 672 ± 67 e | 1815 ± 180 bc | 1858 ± 184 b |
| Na | 873 ± 83 c | 831 ± 80 c | 328 ± 33 e | 502 ± 47 d | 10,851 ± 1001 a | 11,080 ± 1000 a | 3357 ± 328 b | 2939 ± 289 b |
| P | 302 ± 27 d | 373 ± 33 d | 4366 ± 431 b | 8167 ± 802 a | 4017 ± 389 b | 9655 ± 925 a | 1938 ± 190 c | 2175 ± 207 c |
| B | 7.90 ± 0.68 c | 7.24 ± 0.69 c | 23.65 ± 2.20 b | 6.59 ± 0.62 c | 4.10 ± 0.40 d | 2.00 ± 0.20 e | 22.23 ± 2.11 b | 46.99 ± 4.7 a |
| Co | 1.23 ± 0.12 a | 0.28 ± 0.03 c | 1.04 ± 0.11 a | 0.20 ± 0.02 d | 0.07 ± 0.01 e | 0.02 ± 0.01 f | 0.25 ± 0.02 c | 0.64 ± 0.06 b |
| Cu | 5.18 ± 0.49 d | 2.80 ± 0.30 e | 82.14 ± 8.01 a | 103.00 ± 10.10 a | 9.96 ± 0.90 c | 11.21 ± 1.10 c | 10.17 ± 1.01 c | 19.64 ± 1.88 b |
| Fe | 5072 ± 489 a | 1347 ± 127 b | 837 ± 82 c | 267 ± 25 e | 145 ± 14.2 f | 109 ± 10 g | 444 ± 44 d | 1542 ± 145 b |
| I | 15.05 ± 1.45 a | 1.23 ± 0.11 bc | 0.76 ± 0.07 d | 0.40 ± 0.04 e | 0.40 ± 0.04 e | 0.33 ± 0.03 e | 1.03 ± 0.09 c | 1.34 ± 0.12 b |
| Li | 3.50 ± 0.34 a | 0.96 ± 0.09 c | 0.65 ± 0.06 e | 0.49 ± 0.05 f | 0.08 ± 0.01 g | 0.07 ± 0.01 g | 0.89 ± 0.08 d | 2.36 ± 0.20 b |
| Mn | 29.20 ± 2.88 c | 14.00 ± 1.30 d | 62.46 ± 6.12 b | 14.50 ± 1.43 d | 9.09 ± 0.90 e | 6.59 ± 0.60 f | 57.48 ± 5.71 b | 117 ± 10 a |
| Mo | 0.31 ± 0.03 b | 0.23 ± 0.02 c | 0.93 ± 0.09 a | 0.15 ± 0.01 d | 0.15 ± 0.01 d | 0.12 ± 0.01 f | 0.31 ± 0.03 b | 0.98 ± 0.09 a |
| Se | 0.25 ± 0.02 e | 0.15 ± 0.01 f | 6.14 ± 0.59 b | 8.13 ± 0.80 a | 0.59 ± 0.06 d | 0.85 ± 0.08 c | 0.08 ± 0.01 g | 0.29 ± 0.03 e |
| Si | 22.26 ± 2.01 c | 15.06 ± 1.43 c | 5.93 ± 0.60 f | 54.90 ± 5.20 a | 5.68 ± 0.57 f | 38.67 ± 3.82 b | 10.06 ± 1.00 e | 13.42 ± 1.29 d |
| Zn | 16.08 ± 1.55 g | 30.24 ± 3.00 ef | 107.00 ± 10.00 a | 59.30 ± 5.35 b | 39.19 ± 3.85 cd | 28.12 ± 2.80 f | 33.19 ± 3.30 de | 44.91 ± 4.38 c |
| Al | 3139 ± 301 a | 1086 ± 100 b | 193 ± 18 e | 126 ± 12 f | 99 ± 9 g | 59 ± 6 h | 346 ± 32 d | 721 ± 72 c |
| As | 1.41 ± 0.11 a | 1.48 ± 0.15 a | 0.90 ± 0.09 b | 0.28 ± 0.02 d | 0.04 ± 0.01 d | 0.02 ± 0.01 f | 0.20 ± 0.02 e | 0.70 ± 0.07 c |
| Cd | 0.06 ± 0.01 f | 0.30 ± 0.03 d | 1.20 ± 0.10 a | 0.63 ± 0.06 c | 0.84 ± 0.08 b | 0.53 ± 0.05 c | 0.12 ± 0.01 e | 0.08 ± 0.01 f |
| Cr | 54.46 ± 5.10 a | 2.33 ± 0.21 c | 3.39 ± 0.34 b | 0.69 ± 0.06 e | 0.67 ± 0.06 e | 0.28 ± 0.03 f | 1.10 ± 0.10 d | 1.98 ± 0.20 c |
| Ni | 10.90 ± 1.92 a | 3.35 ± 0.33 bc | 3.77 ± 0.35 b | 0.51 ± 0.05 d | 0.30 ± 0.03 b | 0.12 ± 0.01 e | 3.03 ± 0.30 c | 3.04 ± 0.30 c |
| Pb | 5.54 ± 0.52 b | 15.48 ± 1.47 a | 1.09 ± 0.10 c | 0.70 ± 0.07 d | 0.10 ± 0.01 e | 0.07 ± 0.01 f | 0.62 ± 0.06 d | 1.09 ± 0.11 c |
| Sr | 114.00 ± 11.00 a | 49.09 ± 4.78 b | 56.00 ± 5.20 b | 4.67 ± 0.05 c | 2.79 ± 0.30 d | 1.06 ± 0.10 e | 48.14 ± 4.80 b | 107.00 ± 10.00 a |
| V | 6.78 ± 0.63 b | 14.68 ± 1.44 a | 2.89 ± 0.30 c | 0.51 ± 0.04 d | 0.31 ± 0.03 e | 0.29 ± 0.03 e | 2.69 ± 0.24 c | 6.59 ± 0.55 b |
| Object | Location | AOA (mg GAE g−1 d.w.) | TP (mg GAE g−1 d.w.) | Se (µg kg−1 d.w.) | Proline (mg g−1 d.w.) |
|---|---|---|---|---|---|
| Diploschistes ocellatus | Baskunchak | 19.0 ± 1.6 b | 11.0 ± 1.0 b | 250 ± 20 d | 1.03 ± 0.10 c |
| Crimea * | 18.2 ± 1.6 b | 10.1 ± 1.0 b | 261 ± 21 d | 1.02 ± 0.10 c | |
| Calvatia candida | Baskunchak | 21.1 ± 1.9 b | 12.1 ± 1.0 b | 590 ± 43 c | 1.31 ± 0.12 b |
| Crimea ** | 21.4 ± 2.0 b | 11.7 ± 1.0 b | 850 ± 78 b | 1.22 ± 0.11 b | |
| Battarrea phalloides | Baskunchak | 7.9 ± 0.7 c | 7.4 ± 0.7 c | 6140 ± 546 a | 1.12 ± 0.10 bc |
| Crimea ** | 7.2 ± 0.7 c | 6.9 ± 0.6 c | 8130 ± 750 a | 1.09 ± 0.10 bc | |
| Artemisia lerchiana | Baskunchak | 68.6 ± 5.1 a | 21.0 ± 1.8 a | 135 ± 11 e | 5.45 ± 0.44 a |
| Crimea ** | 58.5 ± 5.2 a | 24.1 ± 2.1 a | 116 ± 10.0 e | 5.29 ± 0.41 a |
| Parameter | Diploschistes ocellatus | Artemisia lerchiana |
|---|---|---|
| Chlorophyll a (mg 100 g−1 d.w.) | 6.18 ± 0.53 b | 41.70 ± 4.01 a |
| Chlorophyll b (mg 100 g−1 d.w.) | 13.80 ± 0.70 b | 91.00 ± 9.00 a |
| Total chlorophyll (mg 100 g−1 d.w.) | 19.98 ± 1.40 b | 132.70 ± 1.24 a |
| Chlorophyll b/Chlorophyll a ratio | 2.23 ± 0.20 a | 2.18 ± 0.20 a |
| Carotene (mg 100 g−1 d.w.) | 0.65 ± 0.05 b | 19.00 ± 1.6 a |
| Total chlorophyll/carotene ratio | 30.74 ± 3.00 a | 6.98 ± 0.70 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golubkina, N.; Tolpysheva, T.; Lapchenko, V.; Lapchenko, H.; Pirogov, N.; Zaitsev, V.; Sękara, A.; Tallarita, A.; Stoleru, V.; Murariu, O.C.; et al. Comparative Evaluation of Antioxidant Status and Mineral Composition of Diploschistes ocellatus, Calvatia candida (rostk.) Hollós, Battarrea phalloides and Artemisia lerchiana in Conditions of High Soil Salinity. Plants 2023, 12, 2530. https://doi.org/10.3390/plants12132530
Golubkina N, Tolpysheva T, Lapchenko V, Lapchenko H, Pirogov N, Zaitsev V, Sękara A, Tallarita A, Stoleru V, Murariu OC, et al. Comparative Evaluation of Antioxidant Status and Mineral Composition of Diploschistes ocellatus, Calvatia candida (rostk.) Hollós, Battarrea phalloides and Artemisia lerchiana in Conditions of High Soil Salinity. Plants. 2023; 12(13):2530. https://doi.org/10.3390/plants12132530
Chicago/Turabian StyleGolubkina, Nadezhda, Tatiana Tolpysheva, Vladimir Lapchenko, Helene Lapchenko, Nikolay Pirogov, Viacheslav Zaitsev, Agnieszka Sękara, Alessio Tallarita, Vasile Stoleru, Otilia Cristina Murariu, and et al. 2023. "Comparative Evaluation of Antioxidant Status and Mineral Composition of Diploschistes ocellatus, Calvatia candida (rostk.) Hollós, Battarrea phalloides and Artemisia lerchiana in Conditions of High Soil Salinity" Plants 12, no. 13: 2530. https://doi.org/10.3390/plants12132530