
iTRAQ-Based Proteomic and Physiological Analyses Reveal the Mechanisms of Dehydration and Cryopreservation Tolerance of Sophora tonkinensis Gagnep. Seeds
 , and
, and 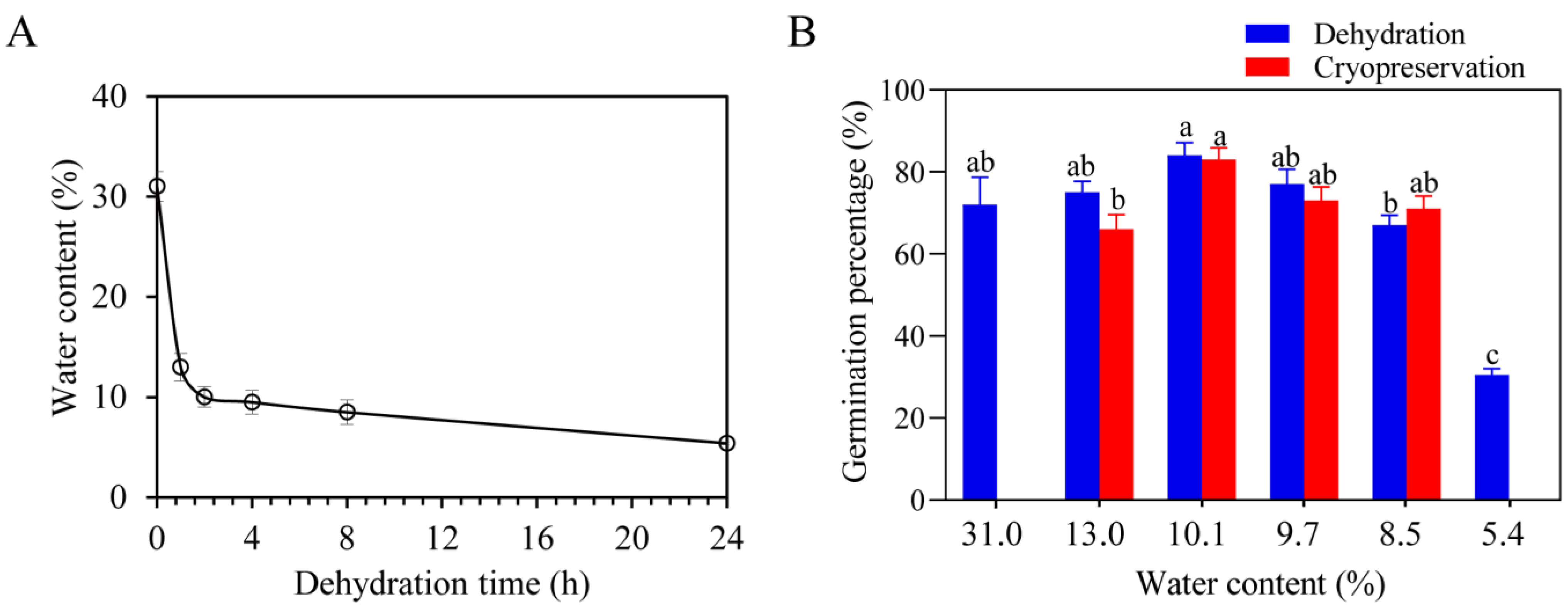{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Dehydration Tolerance of S. tonkinensis Seeds
2.2. Effects of Dehydration and Cryopreservation after Dehydration Treatment on the Cell Structure of S. tonkinensis Seed Plumular Axis
2.3. Response of Antioxidant Enzyme Activities in S. tonkinensis Seed Plumular Axis Treated with Dehydration and Dehydration Followed by Cryopreservation Treatment
2.4. Results of iTRAQ Analysis and Differential Protein Screening of S. tonkinensis Seeds Plumular Axis Treated with Dehydration and Dehydration Followed by Cryopreservation Treatment
2.5. KEGG and Go Functional Annotation of DEPs in S. tonkinensis Seeds Axis Treated with Dehydration and Cryopreservation
3. Discussion
3.1. Molecular Mechanism of Dehydration Tolerance in S. tonkinensis Seeds
3.2. Cryopreservation Treatment May Be Used as a Candidate Method for the Preservation of S. tonkinensis Seeds
4. Materials and Methods
4.1. Experimental Materials
4.2. Germination Experiment
4.3. Cell Structure and Antioxidant Enzyme Activity of Seedplumular Axis Treated with Cryopreservation after Dehydration and Dehydration Treatment
4.4. Protein Extraction and Enzyme Hydrolysis
4.5. iTRAQ Labeling and Grouping
4.6. Database Search and Proteomic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Smolikova, G.; Leonova, T.; Vashurina, N.; Frolov, A.; Medvedev, S. Desiccation Tolerance as the Basis of Long-Term Seed Viability. Int. J. Mol. Sci. 2021, 22, 101. [Google Scholar] [CrossRef]
- Song, S.-Q.; Liu, J.; Tang, C.-F.; Cheng, H.-Y.; Wang, W.-Q.; Zhang, Q.; Zhang, W.-H.; Gao, J.-D. Research Progress on the Physiology and Its Molecular Mechanism of Seed Desiccation Tolerance. Sci. Agric. Sin. 2022, 55, 1047–1063. [Google Scholar]
- Zhongqi, L.; Jiwai, H.; Haiqing, Z.; Aimin, L. Dehydration Tolerance of Plant Seeds: Current Research Situation and Prospects. Chin. Agric. Sci. Bull. 2020, 36, 36. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-4692-7. [Google Scholar]
- McKersie, B.D. Principles of Seed Science and Technology. Plant Sci. 2002, 162, 849. [Google Scholar] [CrossRef]
- Pammenter, N.W.; Berjak, P. Physiology of Desiccation-Sensitive (Recalcitrant) Seeds and the Implications for Cryopreservation. Int. J. Plant Sci. 2014, 175, 21–28. [Google Scholar] [CrossRef]
- Dekkers, B.J.W.; Costa, M.C.D.; Maia, J.; Bentsink, L.; Ligterink, W.; Hilhorst, H.W.M. Acquisition and Loss of Desiccation Tolerance in Seeds: From Experimental Model to Biological Relevance. Planta 2015, 241, 563–577. [Google Scholar] [CrossRef]
- Berjak, P.; Pammenter, N.W. From Avicennia to Zizania: Seed Recalcitrance in Perspective. Ann. Bot. 2007, 101, 213–228. [Google Scholar] [CrossRef]
- Berjak, P. Unifying Perspectives of Some Mechanisms Basic to Desiccation Tolerance across Life Forms. Seed Sci. Res. 2006, 16, 1–15. [Google Scholar] [CrossRef]
- Pammenter, N.W.; Berjak, P. A Review of Recalcitrant Seed Physiology in Relation to Desiccation-Tolerance Mechanisms. Seed Sci. Res. 1999, 9, 13–37. [Google Scholar] [CrossRef]
- Leprince, O.; Pellizzaro, A.; Berriri, S.; Buitink, J. Late Seed Maturation: Drying without Dying. EXBOTJ 2016, 68, 827–841. [Google Scholar] [CrossRef]
- Sano, N.; Rajjou, L.; North, H.M.; Debeaujon, I.; Marion-Poll, A.; Seo, M. Staying Alive: Molecular Aspects of Seed Longevity. Plant Cell Physiol. 2016, 57, 660–674. [Google Scholar] [CrossRef] [PubMed]
- Baud, S.; Dubreucq, B.; Miquel, M.; Rochat, C.; Lepiniec, L. Storage Reserve Accumulation in Arabidopsis: Metabolic and Developmental Control of Seed Filling. Arab. Book 2008, 6, e0113. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.; Buijs, G.; Ligterink, W.; Hilhorst, H. Evolutionary Ecophysiology of Seed Desiccation Sensitivity. Funct. Plant Biol. 2018, 45, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Lepiniec, L.; Devic, M.; Roscoe, T.J.; Bouyer, D.; Zhou, D.-X.; Boulard, C.; Baud, S.; Dubreucq, B. Molecular and Epigenetic Regulations and Functions of the LAFL Transcriptional Regulators That Control Seed Development. Plant Reprod. 2018, 31, 291–307. [Google Scholar] [CrossRef]
- Braybrook, S.A.; Stone, S.L.; Park, S.; Bui, A.Q.; Le, B.H.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. Genes Directly Regulated by LEAFY COTYLEDON2 Provide Insight into the Control of Embryo Maturation and Somatic Embryogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 3468–3473. [Google Scholar] [CrossRef] [PubMed]
- Song, S.-Q.; Tian, M.-H.; Kan, J.; Cheng, H.-Y. The Response Difference of Mitochondria in Recalcitrant Antiaris Toxicaria Axes and Orthodox Zea Mays Embryos to Dehydration Injury. J. Integr. Plant Biol. 2009, 51, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Møller, I.M.; Song, S.-Q. Proteomics of Desiccation Tolerance during Development and Germination of Maize Embryos. J. Proteom. 2012, 75, 1247–1262. [Google Scholar] [CrossRef]
- Carbonero, P.; Iglesias-Fernández, R.; Vicente-Carbajosa, J. The AFL Subfamily of B3 Transcription Factors: Evolution and Function in Angiosperm Seeds. J. Exp. Bot. 2017, 68, 871–880. [Google Scholar] [CrossRef]
- Fatihi, A.; Boulard, C.; Bouyer, D.; Baud, S.; Dubreucq, B.; Lepiniec, L. Deciphering and Modifying LAFL Transcriptional Regulatory Network in Seed for Improving Yield and Quality of Storage Compounds. Plant Sci. 2016, 250, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Sall, K.; Dekkers, B.J.W.; Nonogaki, M.; Katsuragawa, Y.; Koyari, R.; Hendrix, D.; Willems, L.A.J.; Bentsink, L.; Nonogaki, H. DELAY OF GERMINATION 1-LIKE 4 Acts as an Inducer of Seed Reserve Accumulation. Plant J. 2019, 100, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Barral, N.; del Rodríguez-Gacio, M.C.; Matilla, A.J. Delay of Germination-1 (DOG1): A Key to Understanding Seed Dormancy. Plants 2020, 9, 480. [Google Scholar] [CrossRef]
- Dekkers, B.J.W.; He, H.; Hanson, J.; Willems, L.A.J.; Jamar, D.C.L.; Cueff, G.; Rajjou, L.; Hilhorst, H.W.M.; Bentsink, L. The Arabidopsis DELAY OF GERMINATION 1 Gene Affects ABSCISIC ACID INSENSITIVE 5 (ABI5) Expression and Genetically Interacts with ABI3 during Arabidopsis Seed Development. Plant J. 2016, 85, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Barciszewska, M.Z.; Barciszewski, J.; Plitta, B.P.; Chmielarz, P. Global Changes in DNA Methylation in Seeds and Seedlings of Pyrus Communis after Seed Desiccation and Storage. PLoS ONE 2013, 8, e70693. [Google Scholar] [CrossRef]
- Bartels, A.; Han, Q.; Nair, P.; Stacey, L.; Gaynier, H.; Mosley, M.; Huang, Q.Q.; Pearson, J.K.; Hsieh, T.-F.; An, Y.-Q.C.; et al. Dynamic DNA Methylation in Plant Growth and Development. Int. J. Mol. Sci. 2018, 19, 2144. [Google Scholar] [CrossRef]
- Radwan, A.; Hara, M.; Kleinwächter, M.; Selmar, D. Dehydrin Expression in Seeds and Maturation Drying: A Paradigm Change. Plant Biol. J. 2014, 16, 853–855. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, H. Seed Germination and Dormancy: The Classic Story, New Puzzles, and Evolution. J. Integr. Plant Biol. 2019, 61, 541–563. [Google Scholar] [CrossRef]
- Chen, D.; Chen, B.; Fang, Y.; Zheng, C.; Ding, C.; Li, J.; Ma, Q. Flora of China; Science Press: Beijing, China, 2018; Volume 40, p. 76. [Google Scholar]
- Zhang, S.; Li, X.; Yang, W.; Zhou, Y. Sophorae Tonkinensis Radix et Rhizome-Induced Pulmonary Toxicity: A Study on the Toxic Mechanism and Material Basis Based on Integrated Omics and Bioinformatics Analyses. J. Chromatogr. B 2021, 1179, 122868. [Google Scholar] [CrossRef]
- Zhang, S.-N.; Li, X.-Z.; Tan, L.-Y.; Zhu, K.-Y. A Review of Pharmacological and Toxicological Effects of Sophora Tonkinensis with Bioinformatics Prediction. Am. J. Chin. Med. 2021, 49, 359–389. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; He, Y.; Li, X.; Yang, W.; Zhou, Y. Biolabel-Led Research Pattern Positions the Effects and Mechanisms of Sophorae Tonkinensis Radix et Rhizome on Lung Diseases: A Novel Strategy for Computer-Aided Herbal Medicine Research Based on Omics and Bioinformatics. Comput. Biol. Med. 2021, 136, 104769. [Google Scholar] [CrossRef]
- Song, L.; Jiang, N.; Tan, G.; Chen, Q.; Feng, S.; Zhang, Z. First Report of Round Leaf Spot on Sophora Tonkinensis Caused by Didymella Glomerata in China. Plant Dis. 2020, 105, 498. [Google Scholar] [CrossRef]
- Chen, D.-J.; Yuan, S.; Zhang, P.; An, Q.; Zou, J.-B.; Yuan, C.-M.; Zhao, L.-H.; Hu, Z.-X.; Hao, X.-J. Two New Isoflavones from the Roots of Sophora Tonkinensis. J. Asian Nat. Prod. Res. 2022, 25, 163–170. [Google Scholar] [CrossRef]
- Wei, K.H.; Xu, J.P.; Li, L.X.; Cai, J.Y.; Miao, J.H.; Li, M.H. In Vitro Induction and Generation of Tetraploid Plants of Sophora Tonkinensis Gapnep. Pharmacogn. Mag. 2018, 14, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Sheng-Li, Y.; Hao-Ke, H.E.; Meng-Yin, T.; Xin, L.I.; Su-Juan, W. Phenylpropanoid glycosides from Sophorae Tonkinensis Radix et Rhizoma. Zhongguo Zhong Yao Za Zhi 2020, 45, 5525–5529. [Google Scholar] [CrossRef]
- Yang, J.-H.; Kang, S.-Q.; Su, X.-H.; Lin, N.; Kong, X.-Y. Visual analysis of research progress of Sophorae Tonkinensis Radix et Rhizoma based on CiteSpace. Chin. Tradit. Herbal. Drugs 2021, 46, 1763–1768. [Google Scholar] [CrossRef]
- Qin, W.-L.; Lin, Z.-Z.; Wu, Q.-H.; Lan, Z.-Z.; Jiang, N. Study of the Cultivation of Wid Sophora tonkinsis Gapnep. Lishizhen Med. Mater. Med. Res. 2006, 17, 1668–1669. [Google Scholar]
- Chen, J.; Du, X.; Xu, X.; Zhang, S.; Yao, L.; He, X.; Wang, Y. Comparative Proteomic Analysis Provides New Insights into the Molecular Basis of Thermal-Induced Parthenogenesis in Silkworm (Bombyx Mori). Insects 2023, 14, 134. [Google Scholar] [CrossRef] [PubMed]
- Roberts, E.H. Predicting the Storage Life of Seeds. Seed Sci. Technol. 1973, 1, 499–514. [Google Scholar]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. An Intermediate Category of Seed Storage Behaviour? I. COFFEE. J. Exp. Bot. 1990, 41, 1167–1174. [Google Scholar] [CrossRef]
- Hong, T.D.; Linington, S.; Ellis, R.H.; Engels, J. Seed Storage Behaviour: A Compendium; International Plant Genetic Resources Institute (IPGRI): Rome, Italy, 1996; ISBN 978-92-9043-280-7. [Google Scholar]
- Liu, D.; Liu, J.-S. Research progress in dehydration tolerance of seed. J. Agric. Sci. Yanbian Univ. 2014, 36, 362–367. [Google Scholar] [CrossRef]
- Zong, M.; Cai, Y.-P.; Jin, X. Effective Factor s of Br owning and Relationship between Br owning and Desiccation Toler ance during the Development of Chestnut Seed. J. Anhui Agric. Sci. 2005, 8, 1405–1407. [Google Scholar] [CrossRef]
- Tao, Y.-L.; Zhu, C. Relationship among Desiccation-tolerance Proteinsand Soluble Sugars before and after Maturation of Castanea mollissima Seeds. Sci. Silv. Sin. 2004, 2, 45–50. [Google Scholar]
- Yang, Q.-H.; Ye, W.-H.; Song, S.-Q.; Yin, W.-S. Seed desiccation -tolerance and its relationship to seed types and developmental stages. ActaBot Boreali Occident. Sin. 2002, 06, 238–245. [Google Scholar]
- Xin, P.-P.; Hu, X.-R.; Zhang, Y.-L.; Hu, C.-L.; Guo, X.-R. Study on energy saving preservation of scallion and rape seed drying and ultra drying. Seed World 2001, 3, 21–22. [Google Scholar]
- Varghese, B.; Sershen; Berjak, P.; Varghese, D.; Pammenter, N.W. Differential Drying Rates of Recalcitrant Trichilia Dregeana Embryonic Axes: A Study of Survival and Oxidative Stress Metabolism. Physiol. Plant. 2011, 142, 326–338. [Google Scholar] [CrossRef]
- Gomes, M.P.; Garcia, Q.S. Reactive Oxygen Species and Seed Germination. Biologia 2013, 68, 351–357. [Google Scholar] [CrossRef]
- Araldi, C.G.; Coelho, C.M.M.; Gaziola, S.A.; Azevedo, R.A. Storage Elicits a Fast Antioxidant Enzyme Activity in Araucaria Angustifolia Embryos. Acta Physiol. Plant 2016, 38, 201. [Google Scholar] [CrossRef]
- Zhuang, Z.; Shi, F.-H.; Ding, Y.-F.; Shen, Y.-B. Study on dehydration tolerance of Citron Fruit seed. Sci. Agric. JiangSu 2015, 43, 229–231. [Google Scholar] [CrossRef]
- Pizarro, M.; Contreras, R.A.; Köhler, H.; Zúñiga, G.E. Desiccation Tolerance in the Antarctic Moss Sanionia Uncinata. Biol. Res. 2019, 52, 46. [Google Scholar] [CrossRef] [PubMed]
- Manghwar, H.; Li, J. Endoplasmic Reticulum Stress and Unfolded Protein Response Signaling in Plants. Int. J. Mol. Sci. 2022, 23, 828. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-X.; Howell, S.H. Endoplasmic Reticulum Protein Quality Control and Its Relationship to Environmental Stress Responses in Plants. Plant Cell 2010, 22, 2930–2942. [Google Scholar] [CrossRef]
- Al-Whaibi, M.H. Plant Heat-Shock Proteins: A Mini Review. J. King Saud. Univ. Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef]
- Cheng, X.; Wang, W.-J.; Cao, S.; Li, R.-X.; Zhang, B.-B.; Sun, A.-Q.; Zhang, C.-Q. Mechanism of DA-6 Treatment Regulating Wheat Seed Vigor After Anthesis. Sci. Agric. Sin. 2021, 54, 1821–1834. [Google Scholar]
- Demska, K.; Filip, E.; Skuza, L. Expression of Genes Encoding Protein Disulfide Isomerase (PDI) in Cultivars and Lines of Common Wheat with Different Baking Quality of Flour. BMC Plant Biol. 2018, 18, 294. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-Q.; Feng, L.-H.; Gu, M.-H.; Liu, Q.-Q. Progress on inheritance and gene cloning for rice grain quality in Jiangsu province. Hereditas 2021, 43, 425–441. [Google Scholar] [CrossRef]
- Urade, R. Oxidative Protein Folding in the Plant Endoplasmic Reticulum. Biosci. Biotechnol. Biochem. 2019, 83, 781–793. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Cur. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Arnér, E.S.; Holmgren, A. Physiological Functions of Thioredoxin and Thioredoxin Reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef]
- Obroucheva, N.; Sinkevich, I.; Lityagina, S. Physiological Aspects of Seed Recalcitrance: A Case Study on the Tree Aesculus hippocastanum. Tree Physiol. 2016, 36, 1127–1150. [Google Scholar] [CrossRef]
- Amara, I.; Zaidi, I.; Masmoudi, K.; Ludevid, M.D.; Pagès, M.; Goday, A.; Brini, F. Insights into Late Embryogenesis Abundant (LEA) Proteins in Plants: From Structure to the Functions. Am. J. Plant Sci. 2014, 5, 3440–3455. [Google Scholar] [CrossRef]
- Liu, H.; Xing, M.; Yang, W.; Mu, X.; Wang, X.; Lu, F.; Wang, Y.; Zhang, L. Genome-Wide Identification of and Functional Insights into the Late Embryogenesis Abundant (LEA) Gene Family in Bread Wheat (Triticum aestivum). Sci. Rep. 2019, 9, 13375. [Google Scholar] [CrossRef]
- Huang, R.; Xiao, D.; Wang, X.; Zhan, J.; Wang, A.; He, L. Genome-Wide Identification, Evolutionary and Expression Analyses of LEA Gene Family in Peanut (Arachis hypogaea L.). BMC Plant Biol. 2022, 22, 155. [Google Scholar] [CrossRef] [PubMed]
- Geng, W.; Wang, Y.; Zhang, J.; Liu, Z.; Chen, X.; Qin, L.; Yang, L.; Tang, H. Genome-Wide Identification and Expression Analyses of Late Embryogenesis Abundant (LEA) Gene Family in Tobacco (Nicotiana tabacum L.) Reveal Their Function in Abiotic Stress Responses. Gene 2022, 836, 146665. [Google Scholar] [CrossRef]
- Li, Z. Identification and Genetic Effect of LEA Genes During Seed Development in Linseed Flax. Master’Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2021. Available online: https://kns.cnki.net/kcms2/article/abstract?v=3uoqIhG8C475KOm_zrgu4lQARvep2SAkOTSE1G1uB0_um8HHdEYmZtFg9vXhdECNf9kdrWHWDI9g8wdpBt7Zg4FLKWL-UD4b&uniplatform=NZKPT (accessed on 15 February 2023).
- Cheng, J.-Y.; Zhang, D. Correlation Analysis of Gene Expression of LEA Protein and Moisture Content During Embryo Maturation in Nelumbo nucifera. J. Northwest For. Univ. 2020, 35, 75–81+120. [Google Scholar]
- Zhang, H.M. Proteomic Analysis of Drought Stress Responses and LEA proteins. Ph. D. Thesis, Northwest A & F University, Beijing, China, 2014. Available online: https://kns.cnki.net/kcms2/article/abstract?v=3uoqIhG8C447WN1SO36whLpCgh0R0Z-i4Lc0kcI_HPe7ZYqSOTP4QvUsX-vsGudGdptCq0hsEzYmYphSeKmAOmJonwPFphA1&uniplatform=NZKPT (accessed on 15 February 2023).
- Jin, X.; Cao, D.; Wang, Z.; Ma, L.; Tian, K.; Liu, Y.; Gong, Z.; Zhu, X.; Jiang, C.; Li, Y. Genome-Wide Identification and Expression Analyses of the LEA Protein Gene Family in Tea Plant Reveal Their Involvement in Seed Development and Abiotic Stress Responses. Sci. Rep. 2019, 9, 14123. [Google Scholar] [CrossRef]
- Delahaie, J.; Hundertmark, M.; Bove, J.; Leprince, O.; Rogniaux, H.; Buitink, J. LEA Polypeptide Profiling of Recalcitrant and Orthodox Legume Seeds Reveals ABI3-Regulated LEA Protein Abundance Linked to Desiccation Tolerance. J. Exp. Bot. 2013, 64, 4559–4573. [Google Scholar] [CrossRef]
- Yang, Q.-H.; Song, S.-Q.; Ye, W.-H.; Yin, S.-H. Relationship between dehydration tolerance and sugar in seeds. Bull. Bot. Res. 2003, 40, 204–210. [Google Scholar]
- Thormann, I.; Dulloo, M.; Engels, J. Techniques for Ex Situ Plant Conservation. In Plant Conservation Genetics; Haworth Press: Philadelphia, PA, USA, 2006; pp. 7–36. ISBN 978-1-56022-997-1. [Google Scholar]
- Walters, C.; Berjak, P.; Pammenter, N.; Kennedy, K.; Raven, P. Preservation of Recalcitrant Seeds. Science 1984, 339, 915–916. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Yoshida, S. Survival of Plant Tissue at Super-Low Temperature VI. Effects of Cooling and Rewarming Rates on Survival. Plant Physiol. 1967, 42, 1695–1701. [Google Scholar] [CrossRef]
- Zhao, D.-Q.; Song, J.-P.; Jia, H.-X.; Zhang, X.-H.; Yang, W.-L.; Wu, Y.-H.; Song, Z.; Wang, H.-P. Study on Cryopreservation Method of Allium Crop Germplasm. Plant Genet. Resour. 2022, 23, 1258–1268. [Google Scholar] [CrossRef]
- Hirano, T.; Godo, T.; Miyoshi, K.; Ishikawa, K.; Ishikawa, M.; Mii, M. Cryopreservation and Low-Temperature Storage of Seeds of Phaius tankervilleae. Plant Biotechnol. Rep. 2009, 3, 103–109. [Google Scholar] [CrossRef]
- de Oliveira, R.S.; Souza, F.V.D.; dos Santos, I.L.; de Souza, S.O.; Aona, L.Y.S.; de Souza, E.H. Cryopreservation and Low-Temperature Storage of Seeds of Tillandsia Species (Bromeliaceae) with Ornamental Potential. 3 Biotech. 2021, 11, 186. [Google Scholar] [CrossRef]
- Vishwakarma, P.K.; Vincent, L.; Vasugi, C.; Rajasekharan, P.E. Effect of Cryopreservation on Pollen Viability, Fertility and Morphology of Different Psidium Species. Cryobiology 2021, 98, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Rasl, T.; Schalk, M.; Temsch, E.; Kodym, A. Direct Cryopreservation of Winter-Acclimated Buds of Dracocephalum austriacum (Lamiaceae) from Field Material. Plant Cell Tissue Organ Culture 2020, 142, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Ninagawa, T.; Eguchi, A.; Kawamura, Y.; Konishi, T.; Narumi, A. A Study on Ice Crystal Formation Behavior at Intracellular Freezing of Plant Cells Using a High-Speed Camera. Cryobiology 2016, 73, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.-M.; Li, H.-L.; He, Y.; Hong, Q.-M.; Hu, W.-B.; Li, Q. Research Progress on Recalcitrant Seeds Storage of Tropical Crops. Chin. J. Trop. Agric. 2014, 34, 52–58. [Google Scholar]
- Kholina, A.B.; Voronkova, N.M. Seed Cryopreservation of Some Medicinal Legumes. J. Bot. 2012, 2012, 186891. [Google Scholar] [CrossRef]
- Hardstaff, L.K.; Sommerville, K.D.; Funnekotter, B.; Bunn, E.; Offord, C.A.; Mancera, R.L. Myrtaceae in Australia: Use of Cryobiotechnologies for the Conservation of a Significant Plant Family under Threat. Plants 2022, 11, 1017. [Google Scholar] [CrossRef]
- Dussert, S.; Chabrillange, N.; Rocquelin, G.; Engelmann, F.; Lopez, M.; Hamon, S. Tolerance of Coffee (Coffea Spp.) Seeds to Ultra-Low Temperature Exposure in Relation to Calorimetric Properties of Tissue Water, Lipid Composition, and Cooling Procedure. Physiol. Plant 2001, 112, 495–504. [Google Scholar] [CrossRef]
- Zeng, L.; He, M.-J.; Gu, Y.-K.; Wu, Y.; Zheng, X.-L. Study on Cryopreservation of Five Kinds of Medicinal Legume Seeds. Biot. Resour. 2017, 39, 42–47. [Google Scholar] [CrossRef]
- Xia, F.; Wang, X.; Li, M.; Mao, P. Mitochondrial Structural and Antioxidant System Responses to Aging in Oat (Avena sativa L.) Seeds with Different Moisture Contents. Plant Physiology and Biochemistry 2015, 94, 122–129. [Google Scholar] [CrossRef]
- Cheng, C.; Chen, L.; Chou, C.; Liang, J. Investigations of Riboflavin Photolysis via Coloured Light in the Nitro Blue Tetrazolium Assay for Superoxide Dismutase Activity. J. Photochem. 2015, 148, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Maehly, A.C.; Chance, B. The Assay of Catalases and Peroxidases. Methods Biochem. Anal. 1954, 1, 357–424. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Asada, K. Inactivation of Ascorbate Peroxidase in Spinach Chloroplasts on Dark Addition of Hydrogen Peroxide: Its Protection by Ascorbate. Plant Cell Physiol. 1984, 25, 1285. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Zhang, Y.; Jiang, Y.; Dai, Z.; Li, Q.; Mou, J.; Xu, L.; Deng, S.; Li, J.; Wang, R.; et al. iTRAQ-Based Proteomic and Physiological Analyses Reveal the Mechanisms of Dehydration and Cryopreservation Tolerance of Sophora tonkinensis Gagnep. Seeds. Plants 2023, 12, 1842. https://doi.org/10.3390/plants12091842
Luo Y, Zhang Y, Jiang Y, Dai Z, Li Q, Mou J, Xu L, Deng S, Li J, Wang R, et al. iTRAQ-Based Proteomic and Physiological Analyses Reveal the Mechanisms of Dehydration and Cryopreservation Tolerance of Sophora tonkinensis Gagnep. Seeds. Plants. 2023; 12(9):1842. https://doi.org/10.3390/plants12091842
Chicago/Turabian StyleLuo, Yongjian, Yixin Zhang, Yu Jiang, Zhangyan Dai, Qing Li, Jiaolin Mou, Li Xu, Shiming Deng, Jitao Li, Ru Wang, and et al. 2023. "iTRAQ-Based Proteomic and Physiological Analyses Reveal the Mechanisms of Dehydration and Cryopreservation Tolerance of Sophora tonkinensis Gagnep. Seeds" Plants 12, no. 9: 1842. https://doi.org/10.3390/plants12091842