
Low-Cost Hyperspectral Imaging to Detect Drought Stress in High-Throughput Phenotyping
 ,
,  , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
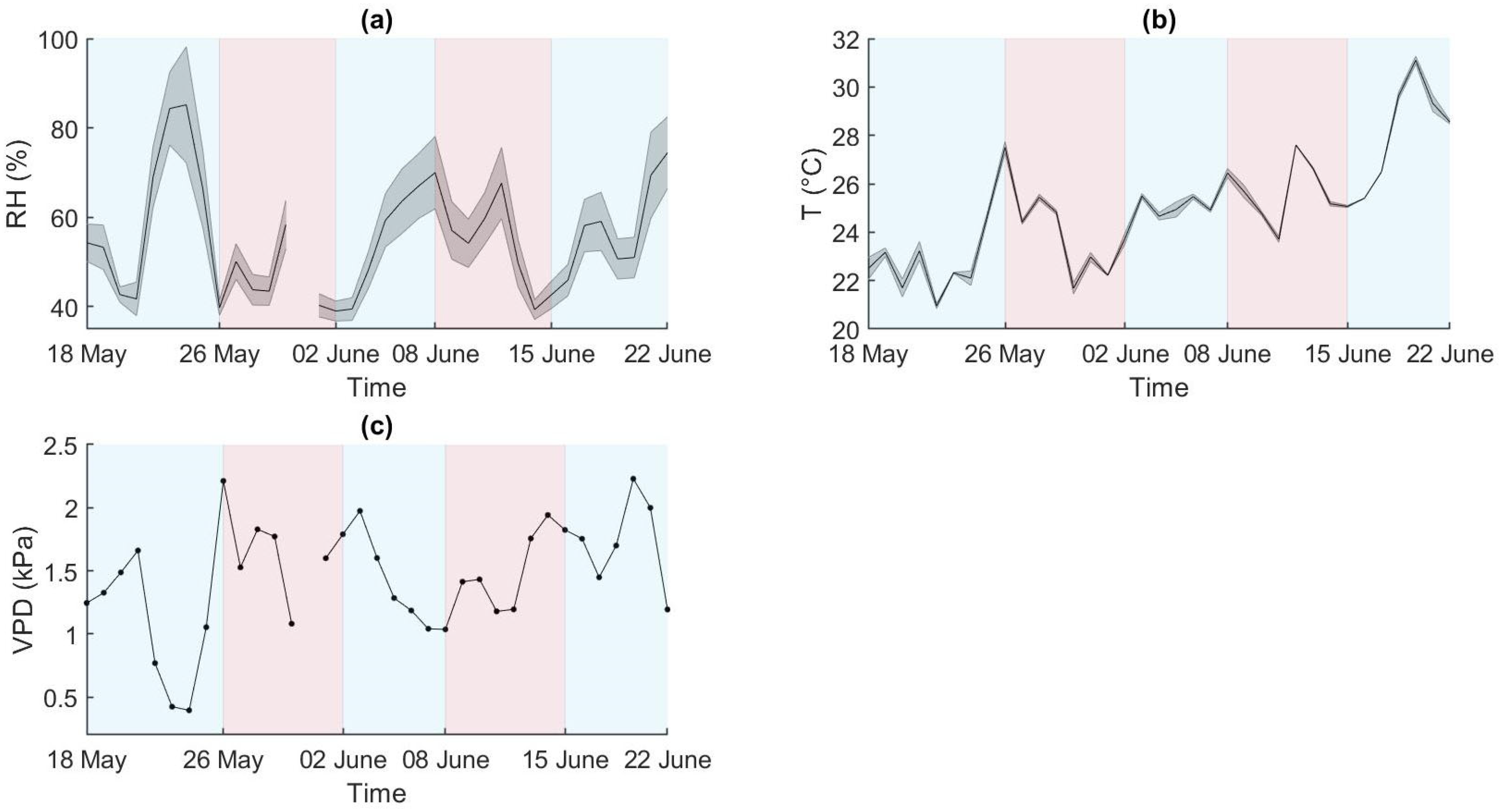
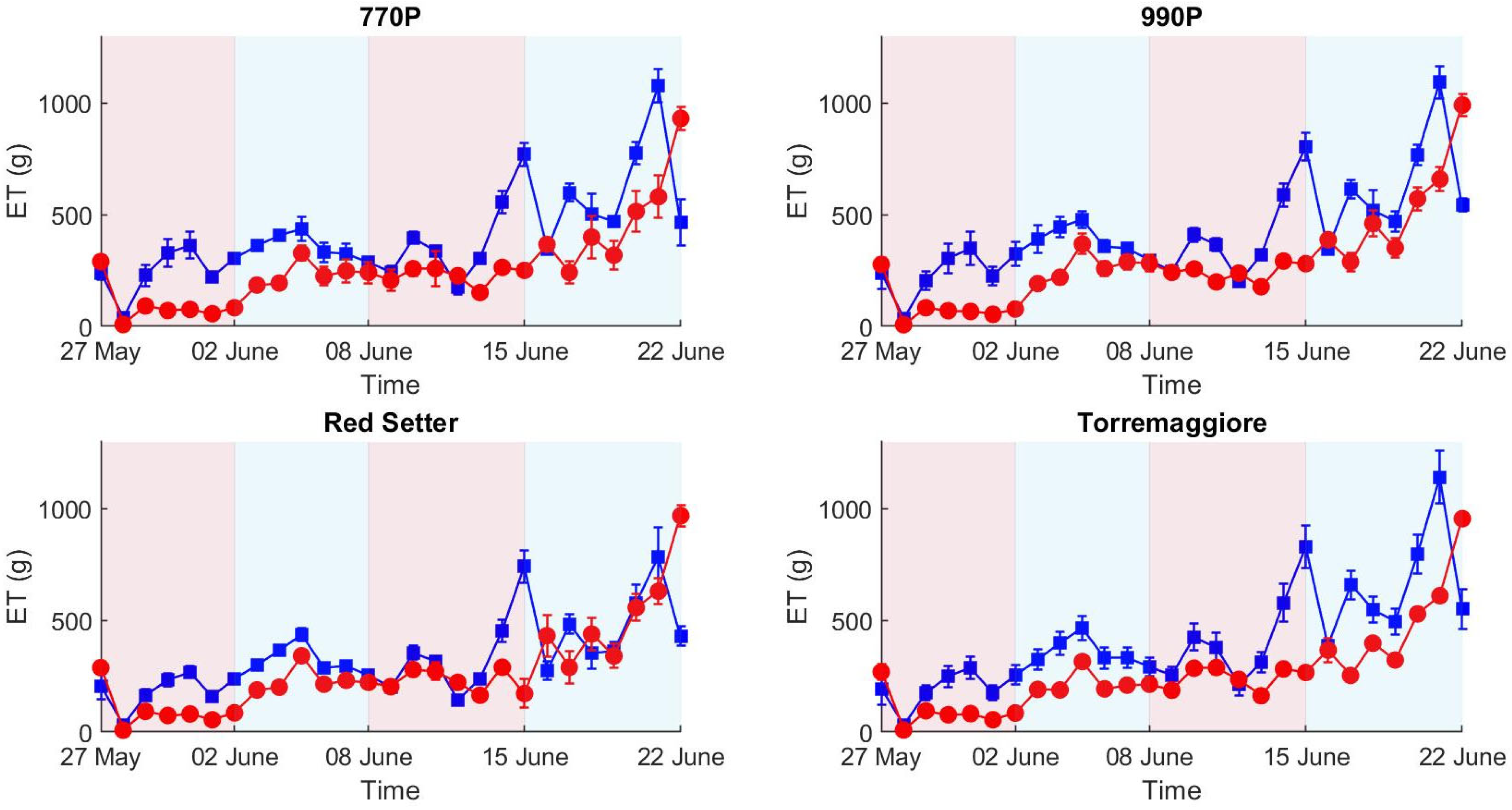
2.1. Environmental Variations and ET
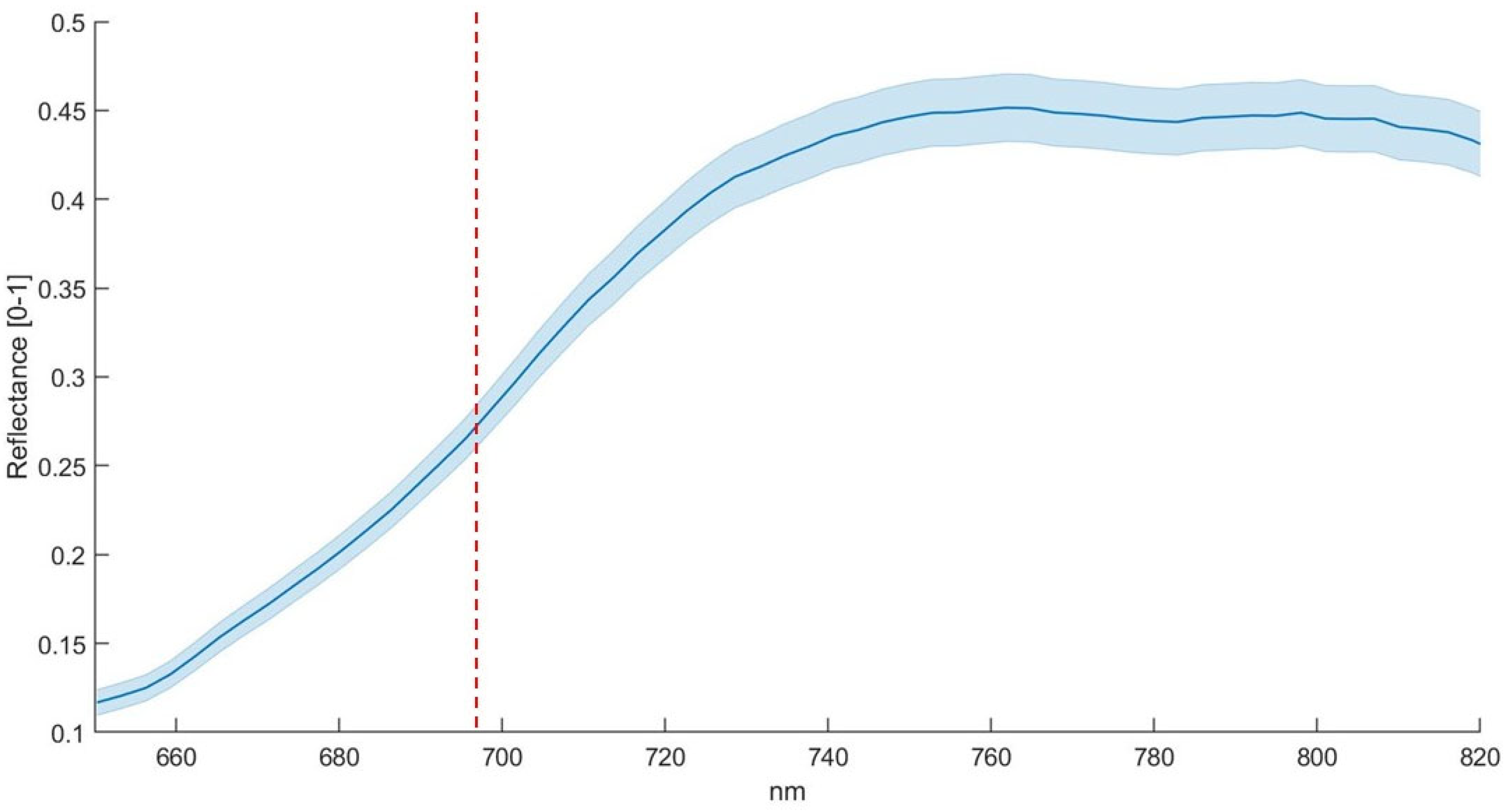
2.2. Hyperspectral Analysis
Segmentation Process
2.3. Hypespectral and RGB Cameras Derived Indices
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Phenotyping Platform and RGB Acquisition Process
4.3. Environmental Monitoring
4.4. Hyperspectral Data Acquisition and Processing
4.5. Stress Induction and Evapotranspiration
4.6. Automatic Segmentation for the Hyperspectral Images
4.7. Hyperspectral Data Analysis
4.8. RGB Images Segmentation and OIs
4.8.1. Projected Shot Area (PSA)
4.8.2. HUE Index (HUE)
4.8.3. Senescence Index (SI)
4.9. Statistical Analysis
5. Conclusions
- (i)
- The integration of the low-cost hyperspectral camera in the HTPP based on the LemnaTec Scanalzer 3D system permitted an increase in the water stress detection capability by the HTPP; indeed, a hyperspectral camera with an NIR spectral range appears more efficient in detecting the stress status during the first stress and recovery phases than the RGB technologies.
- (ii)
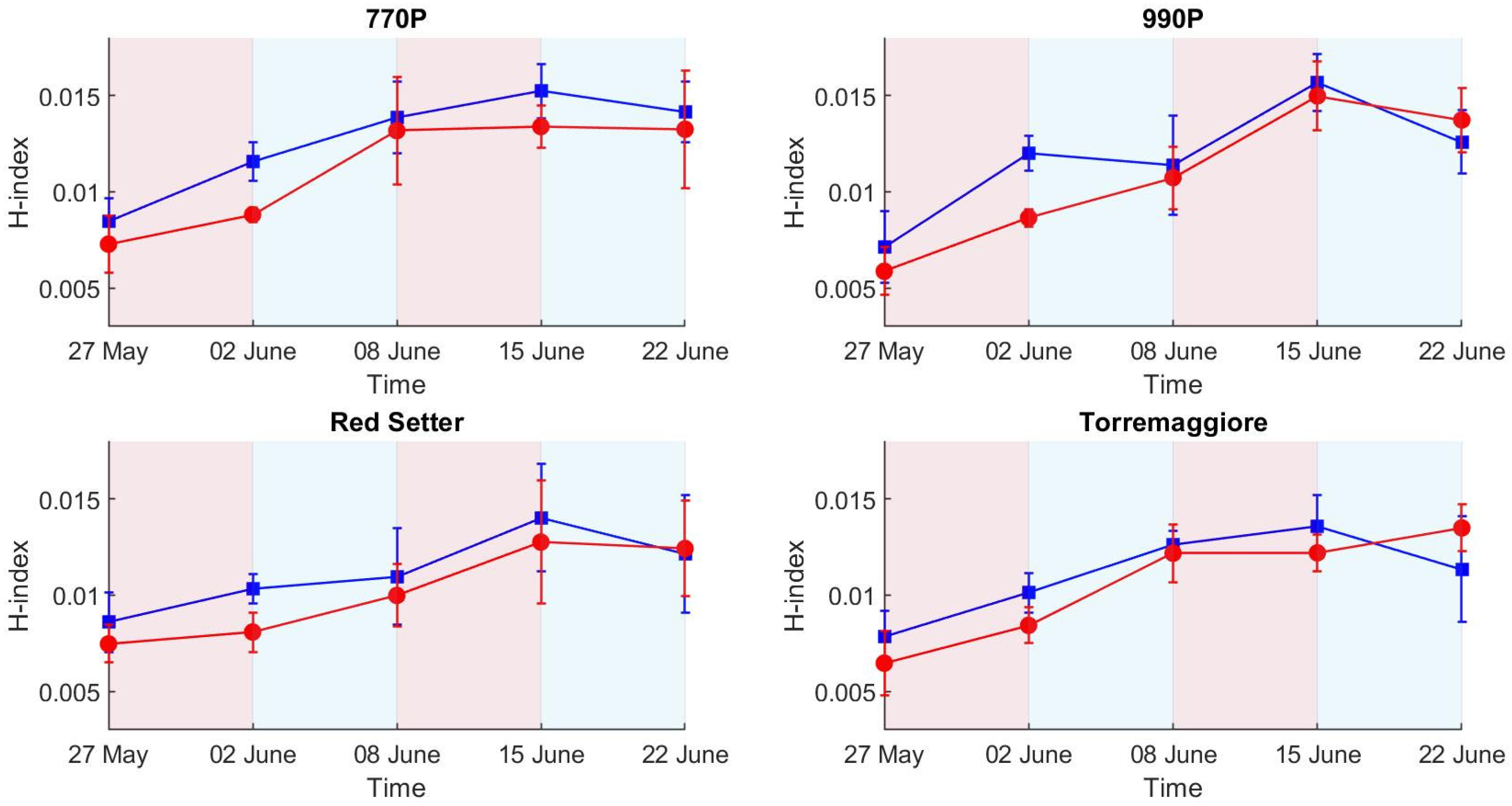
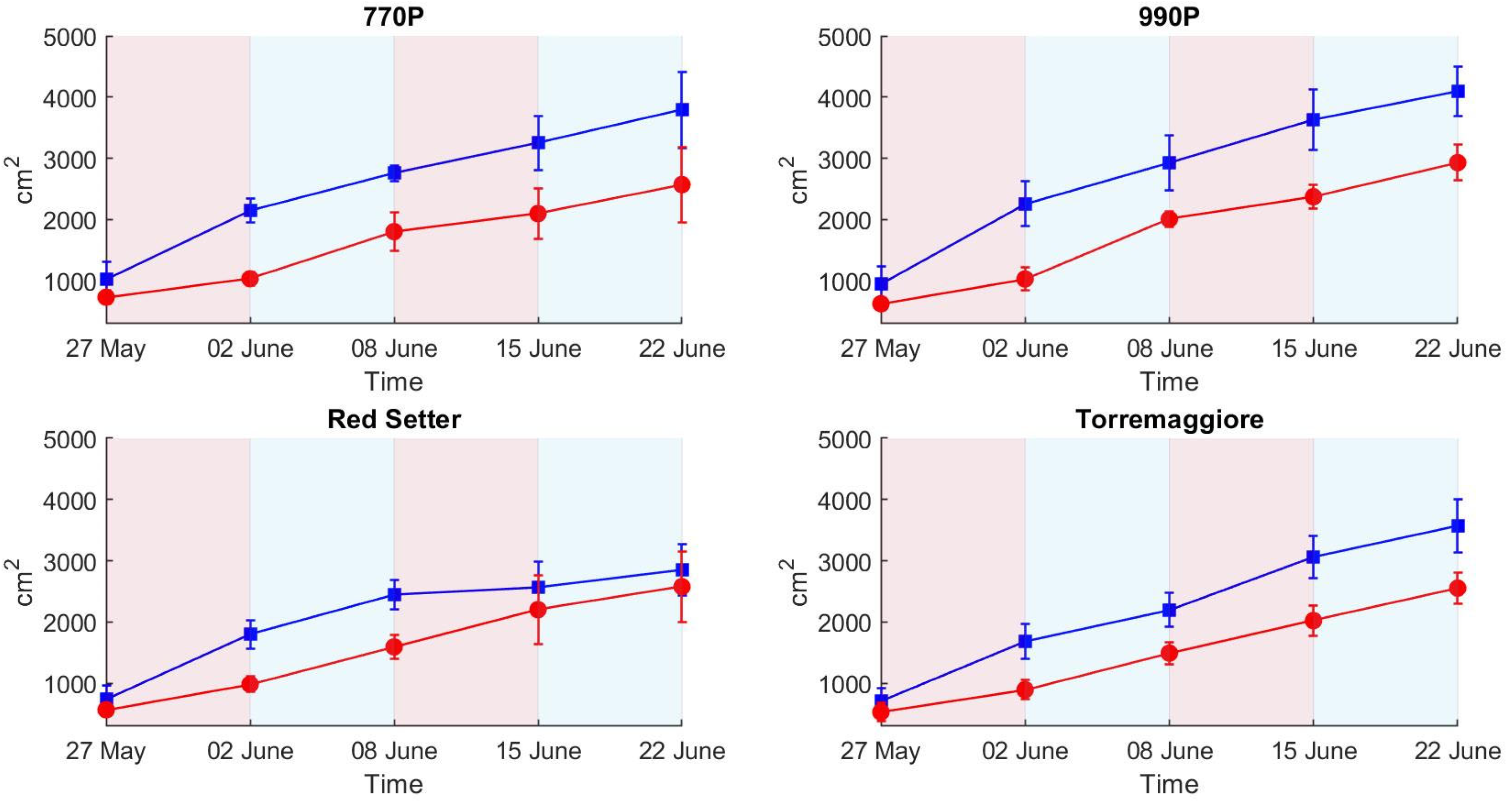
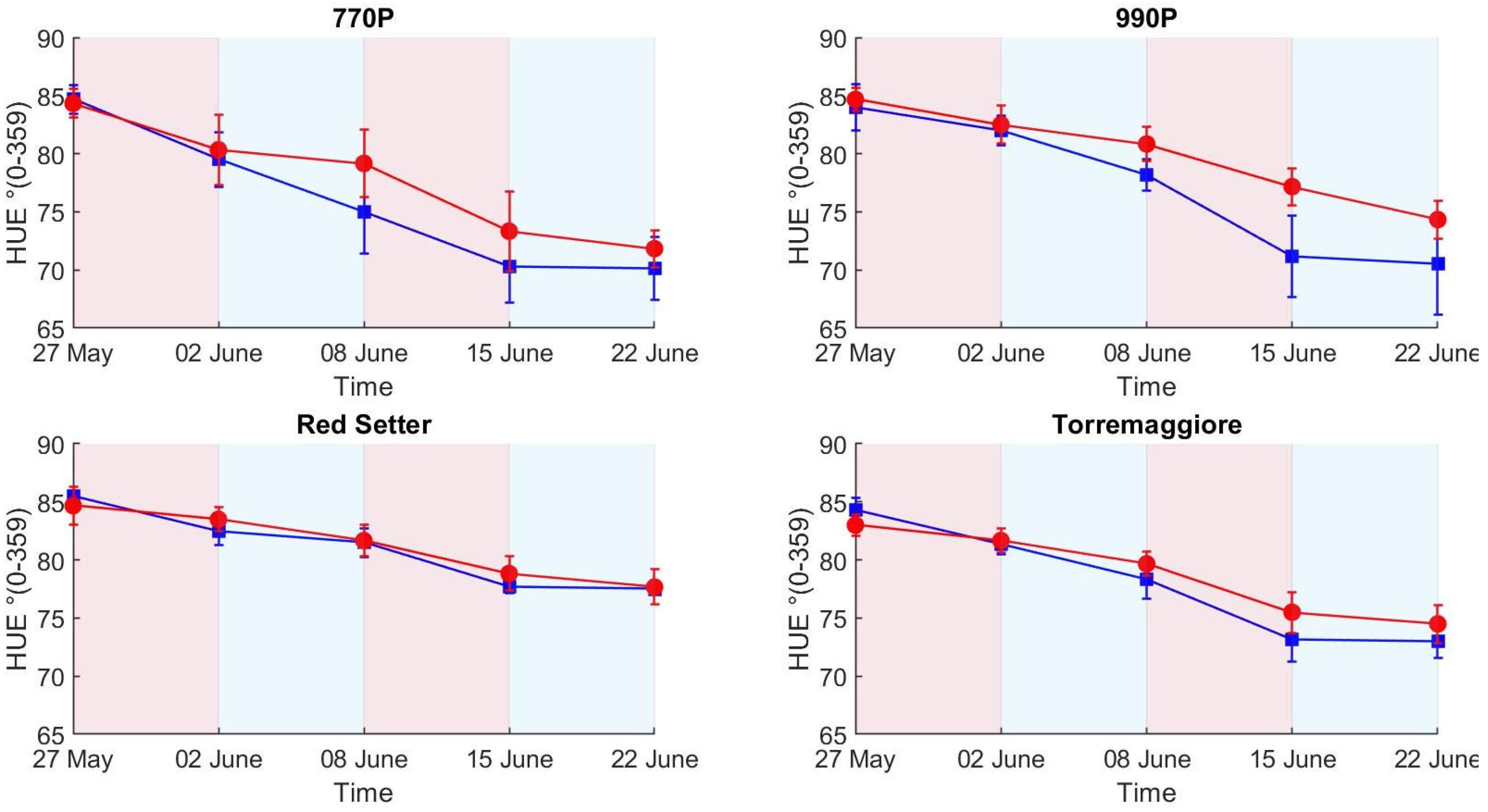
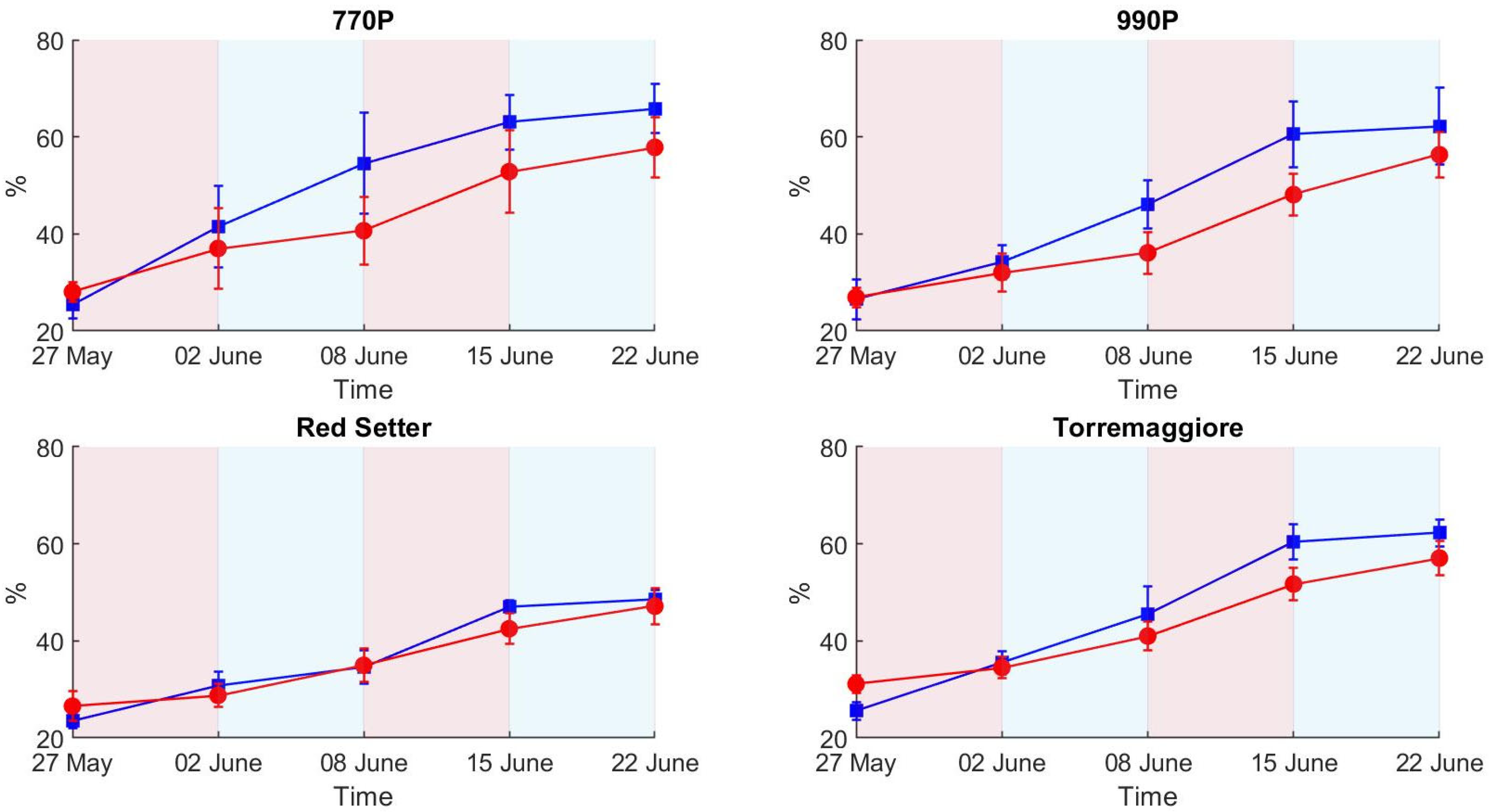
- The PSA more efficiently describes the structural effect of the differential irrigation regimes, particularly during the first stress phase, than HUE and SI, where SI and HUE do not show a clear picture of the irrigation trend compared to the ET results. The H-index more clearly describes the ecophysiological response during the first stress and recovery phases compared to PSA, HUE and SI, showing a higher sensitivity and better reflecting the ET trend. In addition, the H-index more clearly represents the second stress phase than PSA, HUE and SI, which globally appear less visible, probably due to an adaptation to the stress condition by the stressed theses. Based on these results, a low-cost hyperspectral camera with a VIS/NIR spectral range integrated with the RGB technologies commonly used in phenotyping activities appears to be the optimal combination for adoption in the phenotyping process. In addition, this integration could open new ways to develop innovative phenotyping techniques.
- (iii)
- The segmentation method based on the percentile technique in a standardized acquisition set efficiently reduced the hyperspectral dataset dimension, reporting a data reduction of 85.5%; simultaneously, the time needed to process the HI was reduced.
- (iv)
- The tolerance and susceptibility to drought stress of the four genotypes were successfully assessed by the combination of optical and hyperspectral indexes. Overall, the OIs and H-index results show that genotypes 770P and 990P were more susceptible to water stress than Red Setter and Torremaggiore genotypes.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lutz, W.; Sanderson, W.; Scherbov, S. The end of world population growth. Nature 2001, 412, 543–545. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, F.C.; Williams, J. Sustainable food production: Constraints, challenges and choices by 2050. Food Secur. 2015, 7, 221–233. [Google Scholar] [CrossRef]
- Cogato, A.; Meggio, F.; Migliorati, M.D.A.; Marinello, F. Extreme Weather Events in Agriculture: A Systematic Review. Sustainability 2019, 11, 2547. [Google Scholar] [CrossRef]
- Frame, D.J.; Rosier, S.M.; Noy, I.; Harrington, L.J.; Carey-Smith, T.; Sparrow, S.N.; Stone, D.A.; Dean, S.M. Climate change attribution and the economic costs of extreme weather events: A study on damages from extreme rainfall and drought. Clim. Chang. 2020, 162, 781–797. [Google Scholar] [CrossRef]
- Beillouin, D.; Schauberger, B.; Bastos, A.; Ciais, P.; Makowski, D.; Damien, B.; Bernhard, S.; Ana, B.; Phillipe, C.; David, M.; et al. Impact of extreme weather conditions on European crop production in 2018. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190510. [Google Scholar] [CrossRef]
- Dalhaus, T.; Schlenker, W.; Blanke, M.M.; Bravin, E.; Finger, R. The Effects of Extreme Weather on Apple Quality. Sci. Rep. 2020, 10, 7919. [Google Scholar] [CrossRef]
- Elahi, E.; Khalid, Z.; Tauni, M.Z.; Zhang, H.; Lirong, X. Extreme weather events risk to crop-production and the adaptation of innovative management strategies to mitigate the risk: A retrospective survey of rural Punjab, Pakistan. Technovation 2022, 117, 102255. [Google Scholar] [CrossRef]
- Lehmann, J.; Coumou, D.; Frieler, K. Increased record-breaking precipitation events under global warming. Clim. Chang. 2015, 132, 501–515. [Google Scholar] [CrossRef]
- Spinoni, J.; Barbosa, P.; De Jager, A.; McCormick, N.; Naumann, G.; Vogt, J.V.; Magni, D.; Masante, D.; Mazzeschi, M. A new global database of meteorological drought events from 1951 to 2016. J. Hydrol. Reg. Stud. 2019, 22, 100593. [Google Scholar] [CrossRef]
- Niu, S.; Luo, Y.; Li, D.; Cao, S.; Xia, J.; Li, J.; Smith, M.D. Plant growth and mortality under climatic extremes: An overview. Environ. Exp. Bot. 2014, 98, 13–19. [Google Scholar] [CrossRef]
- Asseng, S.; Ewert, F.; Martre, P.; Rotter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. Rising temperatures reduce global wheat production. Nat. Clim. Chang. 2015, 5, 143–147. [Google Scholar] [CrossRef]
- FAO. Damages and Losses; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Costa, C.; Wollenberg, E.; Benitez, M.; Newman, R.; Gardner, N.; Bellone, F. Roadmap for achieving net-zero emissions in global food systems by 2050. Sci. Rep. 2022, 12, 15064. [Google Scholar] [CrossRef] [PubMed]
- Godde, C.; Mason-D’croz, D.; Mayberry, D.; Thornton, P.; Herrero, M. Impacts of climate change on the livestock food supply chain; a review of the evidence. Glob. Food Secur. 2021, 28, 100488. [Google Scholar] [CrossRef] [PubMed]
- Molotoks, A.; Smith, P.; Dawson, T.P. Impacts of land use, population, and climate change on global food security. Food Energy Secur. 2021, 10, e261. [Google Scholar] [CrossRef]
- The 17 Goals|Sustainable Development. Available online: https://sdgs.un.org/goals (accessed on 20 November 2022).
- Food and Agriculture Organization of the United Nations (FAO). Global Update Report: Agriculture, Forestry and Fisheries in the Nationally Determined Contributions 2021 (Interim); FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Pérez-Méndez, N.; Garratt, M.P.D.; Gemmill-Herren, B.; Miguez, F.E.; Dicks, L.V. Policies for Ecological Intensification of Crop Production. Trends Ecol. Evol. 2019, 34, 282–286. [Google Scholar] [CrossRef]
- Khan, M.A.; Shirazi, M.U.; Shereen, A.; Ali, M.; Asma, A.; Jilani, N.S.; Mahboob, W. Agronomical and physiological perspectives for identification of wheat genotypes for high temperature tolerance. Pak. J. Bot. 2020, 52, 1973–1980. [Google Scholar] [CrossRef]
- Vogel, E.; Donat, M.G.; Alexander, L.V.; Meinshausen, M.; Ray, D.K.; Karoly, D.; Meinshausen, N.; Frieler, K. The effects of climate extremes on global agricultural yields. Environ. Res. Lett. 2019, 14, 054010. [Google Scholar] [CrossRef]
- Challinor, A.J.; Wheeler, T.R.; Craufurd, P.Q.; Ferro, C.A.T.; Stephenson, D.B. Adaptation of crops to climate change through genotypic responses to mean and extreme temperatures. Agric. Ecosyst. Environ. 2007, 119, 190–204. [Google Scholar] [CrossRef]
- Tariq, M.; Ahmed, M.; Iqbal, P.; Fatima, Z.; Ahmad, S. Crop Phenotyping. In Systems Modeling; Ahmed, M., Ed.; Springer: Singapore, 2020; pp. 45–60. [Google Scholar] [CrossRef]
- Xu, Y. Envirotyping for deciphering environmental impacts on crop plants. Theor. Appl. Genet. 2016, 129, 653–673. [Google Scholar] [CrossRef]
- Multi Environments and Genetic-Environmental Interaction (GxE) in Plant Breeding and its Challenges: A Review Article. Int. J. Res. Stud. Agric. Sci. 2021, 7, 11–18. [CrossRef]
- Pratap, A.; Gupta, S.; Nair, R.M.; Gupta, S.K.; Schafleitner, R.; Basu, P.S.; Singh, C.M.; Prajapati, U.; Gupta, A.K.; Nayyar, H.; et al. Using Plant Phenomics to Exploit the Gains of Genomics. Agronomy 2019, 9, 126. [Google Scholar] [CrossRef]
- Sarkar, S.; Cazenave, A.-B.; Oakes, J.; McCall, D.; Thomason, W.; Abbott, L.; Balota, M. Aerial high-throughput phenotyping of peanut leaf area index and lateral growth. Sci. Rep. 2021, 11, 21661. [Google Scholar] [CrossRef] [PubMed]
- Nomura, K.; Wada, E.; Saito, M.; Yamasaki, H.; Yasutake, D.; Iwao, T.; Tada, I.; Yamazaki, T.; Kitano, M. Estimation of the Leaf Area Index, Leaf Fresh Weight, and Leaf Length of Chinese Chive (Allium tuberosum) Using Nadir-looking Photography in Combination with Allometric Relationships. Hortscience 2022, 57, 777–784. [Google Scholar] [CrossRef]
- El Haddad, N.; Choukri, H.; Ghanem, M.E.; Smouni, A.; Mentag, R.; Rajendran, K.; Hejjaoui, K.; Maalouf, F.; Kumar, S. High-Temperature and Drought Stress Effects on Growth, Yield and Nutritional Quality with Transpiration Response to Vapor Pressure Deficit in Lentil. Plants 2021, 11, 95. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.X.; Wang, Y.M.; Chen, J.S.; Guo, R.; Shu, R.; Wang, J.Y. Status and application of advanced airborne hyperspectral imaging technology: A review. Infrared Phys. Technol. 2020, 104, 103115. [Google Scholar] [CrossRef]
- Wang, J.; Xu, Y.; Wu, G. The integration of species information and soil properties for hyperspectral estimation of leaf biochemical parameters in mangrove forest. Ecol. Indic. 2020, 115, 106467. [Google Scholar] [CrossRef]
- Pepe, M.; Pompilio, L.; Gioli, B.; Busetto, L.; Boschetti, M. Detection and Classification of Non-Photosynthetic Vegetation from PRISMA Hyperspectral Data in Croplands. Remote Sens. 2020, 12, 3903. [Google Scholar] [CrossRef]
- Tagliabue, G.; Boschetti, M.; Bramati, G.; Candiani, G.; Colombo, R.; Nutini, F.; Pompilio, L.; Rivera-Caicedo, J.P.; Rossi, M.; Rossini, M.; et al. Hybrid retrieval of crop traits from multi-temporal PRISMA hyperspectral imagery. ISPRS J. Photogramm. Remote Sens. 2022, 187, 362–377. [Google Scholar] [CrossRef]
- Damm, A.; Cogliati, S.; Colombo, R.; Fritsche, L.; Genangeli, A.; Genesio, L.; Hanus, J.; Peressotti, A.; Rademske, P.; Rascher, U.; et al. Response times of remote sensing measured sun-induced chlorophyll fluorescence, surface temperature and vegetation indices to evolving soil water limitation in a crop canopy. Remote Sens. Environ. 2022, 273, 112957. [Google Scholar] [CrossRef]
- Di Gennaro, S.F.; Toscano, P.; Gatti, M.; Poni, S.; Berton, A.; Matese, A. Spectral Comparison of UAV-Based Hyper and Multispectral Cameras for Precision Viticulture. Remote Sens. 2022, 14, 449. [Google Scholar] [CrossRef]
- Genangeli, A.; Allasia, G.; Bindi, M.; Cantini, C.; Cavaliere, A.; Genesio, L.; Giannotta, G.; Miglietta, F.; Gioli, B. A Novel Hyperspectral Method to Detect Moldy Core in Apple Fruits. Sensors 2022, 22, 4479. [Google Scholar] [CrossRef] [PubMed]
- Sarić, R.; Nguyen, V.D.; Burge, T.; Berkowitz, O.; Trtílek, M.; Whelan, J.; Lewsey, M.G.; Čustović, E. Applications of hyperspectral imaging in plant phenotyping. Trends Plant Sci. 2022, 27, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Koh, J.C.; Banerjee, B.P.; Spangenberg, G.; Kant, S. Automated hyperspectral vegetation index derivation using a hyperparameter optimisation framework for high-throughput plant phenotyping. New Phytol. 2022, 233, 2659–2670. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Behmann, J.; Steier, A.; Kraska, T.; Muller, O.; Rascher, U.; Mahlein, A.-K. Quantitative assessment of disease severity and rating of barley cultivars based on hyperspectral imaging in a non-invasive, automated phenotyping platform. Plant Methods 2018, 14, 45. [Google Scholar] [CrossRef]
- Stuart, M.B.; Davies, M.; Hobbs, M.J.; Pering, T.D.; McGonigle, A.J.S.; Willmott, J.R. High-Resolution Hyperspectral Imaging Using Low-Cost Components: Application within Environmental Monitoring Scenarios. Sensors 2022, 22, 4652. [Google Scholar] [CrossRef]
- Islam, R.; Ahmed, B.; Hossain, A. Feature reduction of hyperspectral image for classification. J. Spat. Sci. 2020, 67, 331–351. [Google Scholar] [CrossRef]
- Morisse, M.; Wells, D.M.; Millet, E.J.; Lillemo, M.; Fahrner, S.; Cellini, F.; Lootens, P.; Muller, O.; Herrera, J.M.; Bentley, A.R.; et al. A European perspective on opportunities and demands for field-based crop phenotyping. Field Crops Res. 2022, 276, 108371. [Google Scholar] [CrossRef]
- Xie, C.; Yang, C. A review on plant high-throughput phenotyping traits using UAV-based sensors. Comput. Electron. Agric. 2020, 178, 105731. [Google Scholar] [CrossRef]
- Golzarian, M.R.; Frick, R.A.; Rajendran, K.; Berger, B.; Roy, S.; Tester, M.; Lun, D.S. Accurate inference of shoot biomass from high-throughput images of cereal plants. Plant Methods 2011, 7, 2. [Google Scholar] [CrossRef]
- Rezzouk, F.Z.; Gracia-Romero, A.; Kefauver, S.C.; Gutiérrez, N.A.; Aranjuelo, I.; Serret, M.D.; Araus, J.L. Remote sensing techniques and stable isotopes as phenotyping tools to assess wheat yield performance: Effects of growing temperature and vernalization. Plant Sci. 2020, 295, 110281. [Google Scholar] [CrossRef]
- Sancho-Adamson, M.; Trillas, M.I.; Bort, J.; Fernandez-Gallego, J.A.; Romanyà, J. Use of RGB Vegetation Indexes in Assessing Early Effects of Verticillium Wilt of Olive in Asymptomatic Plants in High and Low Fertility Scenarios. Remote. Sens. 2019, 11, 607. [Google Scholar] [CrossRef]
- Noh, H.; Lee, J. The Effect of Vapor Pressure Deficit Regulation on the Growth of Tomato Plants Grown in Different Planting Environments. Appl. Sci. 2022, 12, 3667. [Google Scholar] [CrossRef]
- Sadok, W.; Lopez, J.R.; Smith, K.P. Transpiration increases under high-temperature stress: Potential mechanisms, trade-offs and prospects for crop resilience in a warming world. Plant, Cell Environ. 2021, 44, 2102–2116. [Google Scholar] [CrossRef] [PubMed]
- Jureková, Z.; Németh-Molnár, K.; Paganová, V. Physiological Responses of Six Tomato (Lycopersicon Esculentum Mill.) Cultivars to Water Stress. J. Hortic. For. 2011, 3, 294–300. Available online: https://academicjournals.org/journal/JHF/article-full-text-pdf/7B337231843 (accessed on 3 December 2022).
- Dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological Responses to Drought, Salinity, and Heat Stress in Plants: A Review. Stresses 2022, 2, 113–135. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Helyes, L. Physiological Responses of Selected Vegetable Crop Species to Water Stress. Agronomy 2019, 9, 447. [Google Scholar] [CrossRef]
- Patanè, C.; Tringali, S.; Sortino, O. Effects of deficit irrigation on biomass, yield, water productivity and fruit quality of processing tomato under semi-arid Mediterranean climate conditions. Sci. Hortic. 2011, 129, 590–596. [Google Scholar] [CrossRef]
- Quirino, B.; Noh, Y.-S.; Himelblau, E.; Amasino, R.M. Molecular aspects of leaf senescence. Trends Plant Sci. 2000, 5, 278–282. [Google Scholar] [CrossRef]
- Khan, S.H.; Khan, A.; Litaf, U.; Shah, A.S.; Khan, M.A.; Bilal, M.; Ali, M.U. Effect of Drought Stress on Tomato cv. Bombino. J. Food Process. Technol. 2015, 6, 465. [Google Scholar] [CrossRef]
- Janni, M.; Coppede, N.; Bettelli, M.; Briglia, N.; Petrozza, A.; Summerer, S.; Vurro, F.; Danzi, D.; Cellini, F.; Marmiroli, N.; et al. In Vivo Phenotyping for the Early Detection of Drought Stress in Tomato. Plant Phenomics 2019, 2019, 6168209. [Google Scholar] [CrossRef]
- Boochs, F.; Kupfer, G.; Dockter, K.; Kühbauch, W. Shape of the red edge as vitality indicator for plants. Int. J. Remote Sens. 1990, 11, 1741–1753. [Google Scholar] [CrossRef]
- Schlemmer, M.R.; Francis, D.D.; Shanahan, J.F.; Schepers, J.S. Remotely Measuring Chlorophyll Content in Corn Leaves with Differing Nitrogen Levels and Relative Water Content. Agron. J. 2005, 97, 106–112. [Google Scholar] [CrossRef]
- Tommaselli, A.M.G.; Santos, L.D.; Berveglieri, A.; Oliveira, R.A.; Honkavaara, E. A Study on the Variations of Inner Orientation Parameters of a Hyperspectral Frame Camera. ISPRS-Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2018, 42, 429–436. [Google Scholar] [CrossRef]
- RadhaKrishna, M.; Govindh, M.V.; Veni, P.K. A Review on Image Processing Sensor. J. Physics Conf. Ser. 2021, 1714, 012055. [Google Scholar] [CrossRef]
- Neto, A.J.S.; Lopes, D.C.; Pinto, F.A.C.; Zolnier, S. Vis/NIR spectroscopy and chemometrics for non-destructive estimation of water and chlorophyll status in sunflower leaves. Biosyst. Eng. 2017, 155, 124–133. [Google Scholar] [CrossRef]
- Ghiat, I.; Mackey, H.R.; Al-Ansari, T. A Review of Evapotranspiration Measurement Models, Techniques and Methods for Open and Closed Agricultural Field Applications. Water 2021, 13, 2523. [Google Scholar] [CrossRef]
- Basri, R.; Jacobs, D. Lambertian reflectance and linear subspaces. IEEE Trans. Pattern Anal. Mach. Intell. 2003, 25, 218–233. [Google Scholar] [CrossRef]
- Seneviratne, S.I.; Corti, T.; Davin, E.L.; Hirschi, M.; Jaeger, E.B.; Lehner, I.; Orlowsky, B.; Teuling, A.J. Investigating soil moisture—Climate interactions in a changing climate: A review. Earth-Sci. Rev. 2010, 99, 125–161. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 770P | 990P | Red Setter | Torremaggiore | ||
|---|---|---|---|---|---|
| H-index | ** | ** | ** | ** | |
| I Stress (2 June) | PSA | ** | ** | ** | ** |
| HUE | - | - | - | - | |
| SI | - | - | - | - | |
| H-index | - | - | - | - | |
| I Recovery (8 June) | PSA | ** | ** | ** | ** |
| HUE | - | ** | - | - | |
| SI | - | ** | - | - | |
| H-index | - | - | - | - | |
| II Stress (15 June) | PSA | ** | ** | - | ** |
| HUE | - | ** | - | - | |
| SI | - | ** | ** | ** | |
| H-index | - | - | - | - | |
| II Recovery (22 June) | PSA | ** | ** | - | ** |
| HUE | - | - | - | - | |
| SI | - | - | - | ** |
| Name | Provider | Origin | Plant Growth Habit | Fruit Type | Use |
|---|---|---|---|---|---|
| Red Setter | Portici Seed Collection | Commercial variety | Determinate | Elongated/Blocky | Processing |
| Torremaggiore | La Semiorto Sementi SRL | Southern Italy | Determinate | Round Cherry | Fresh Market |
| 770 P | Portici Seed Collection | Southern Italy | Determinate | Round Cherry with apex | Processing |
| 990 P | Portici Seed Collection | Southern Italy | Semi-Determinate | Round Cherry | Processing |
| Operation | Day |
|---|---|
| Transplant | 7 May 2021 |
| RGB and Hyperspectral Images acquisition | 27 May 2021 |
| Start 1st stress | 27 May 2021 |
| RGB and Hyperspectral Images acquisition | 2 June 2021 |
| Start Recovery | 2 June 2021 |
| RGB and Hyperspectral Images acquisition | 8 June 2021 |
| Start 2nd stress | 8 June 2021 |
| RGB and Hyperspectral Images acquisition | 15 June 2021 |
| Start Recovery | 15 June 2021 |
| RGB and Hyperspectral Images acquisition | 22 June 2021 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genangeli, A.; Avola, G.; Bindi, M.; Cantini, C.; Cellini, F.; Grillo, S.; Petrozza, A.; Riggi, E.; Ruggiero, A.; Summerer, S.; et al. Low-Cost Hyperspectral Imaging to Detect Drought Stress in High-Throughput Phenotyping. Plants 2023, 12, 1730. https://doi.org/10.3390/plants12081730
Genangeli A, Avola G, Bindi M, Cantini C, Cellini F, Grillo S, Petrozza A, Riggi E, Ruggiero A, Summerer S, et al. Low-Cost Hyperspectral Imaging to Detect Drought Stress in High-Throughput Phenotyping. Plants. 2023; 12(8):1730. https://doi.org/10.3390/plants12081730
Chicago/Turabian StyleGenangeli, Andrea, Giovanni Avola, Marco Bindi, Claudio Cantini, Francesco Cellini, Stefania Grillo, Angelo Petrozza, Ezio Riggi, Alessandra Ruggiero, Stephan Summerer, and et al. 2023. "Low-Cost Hyperspectral Imaging to Detect Drought Stress in High-Throughput Phenotyping" Plants 12, no. 8: 1730. https://doi.org/10.3390/plants12081730