
Effect of Seed Priming with Endophytic Bacillus subtilis on Some Physio-Biochemical Parameters of Two Wheat Varieties Exposed to Drought after Selective Herbicide Application

 ,
,
Abstract
:1. Introduction
2. Results
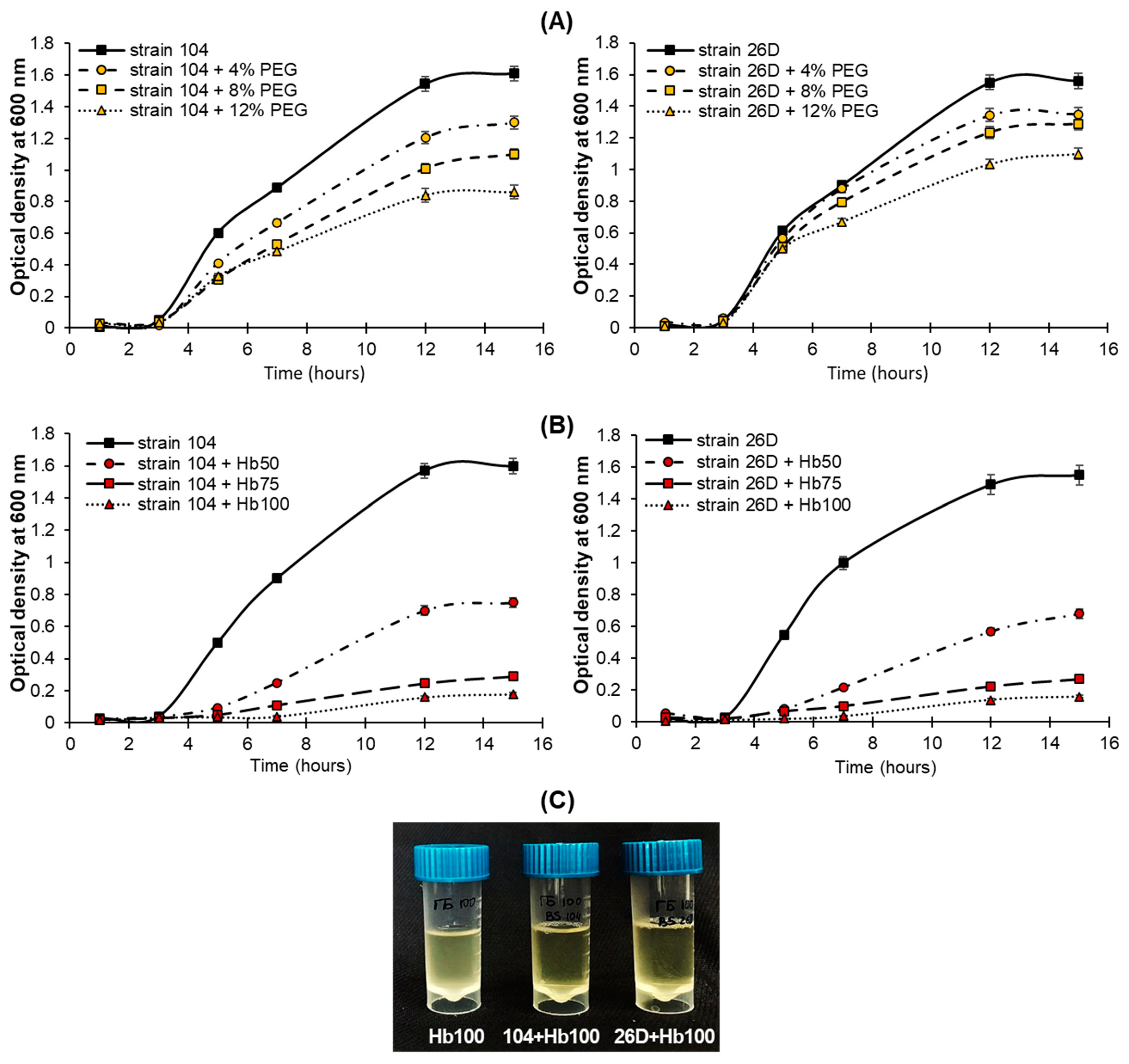
2.1. Herbicide and Drought Tolerance Test
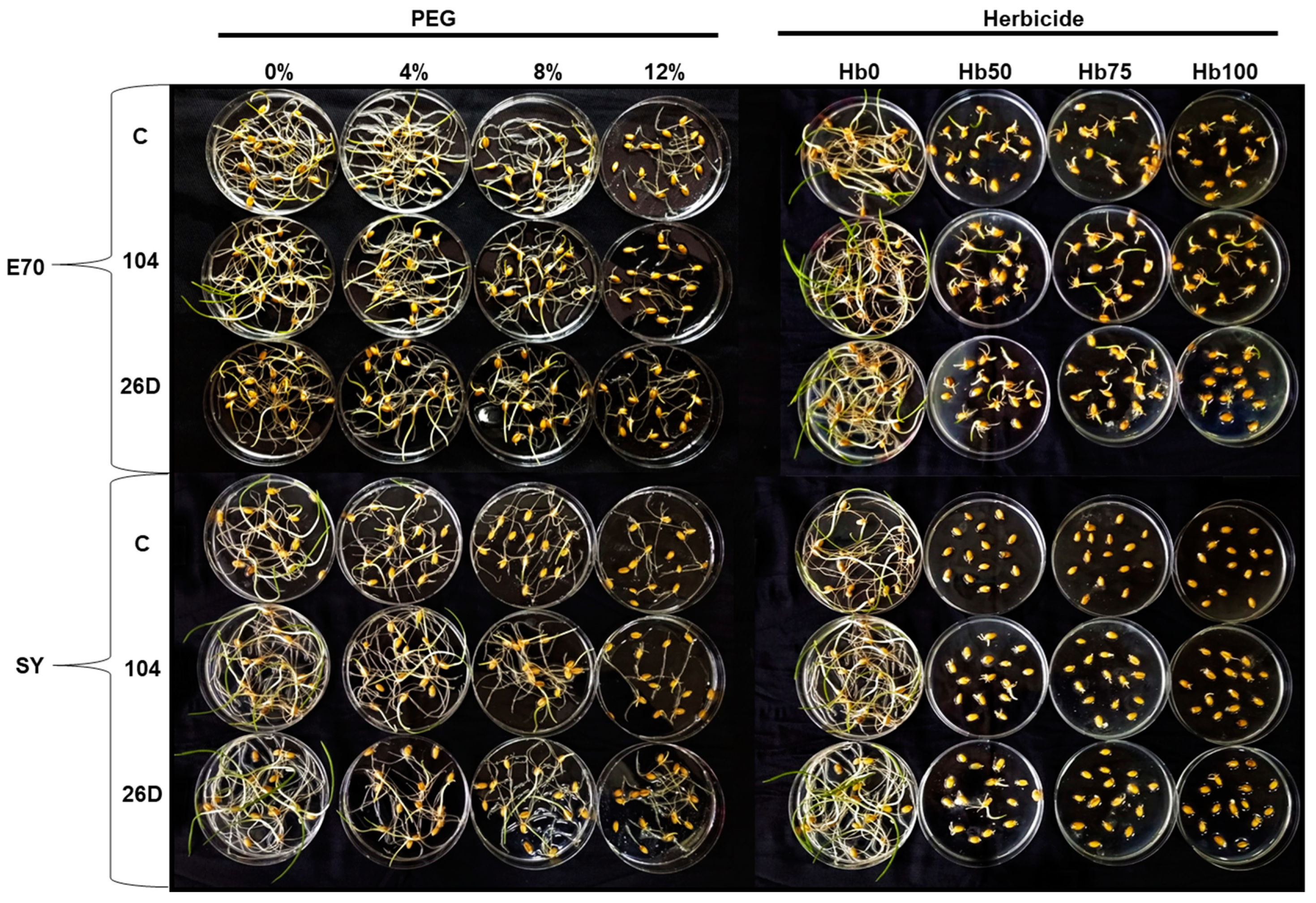
2.2. Effect of Herbicide Sekator® Turbo-Stress and PEG-Stress on Seed Germination and Early Hydroponic Seedling Growth
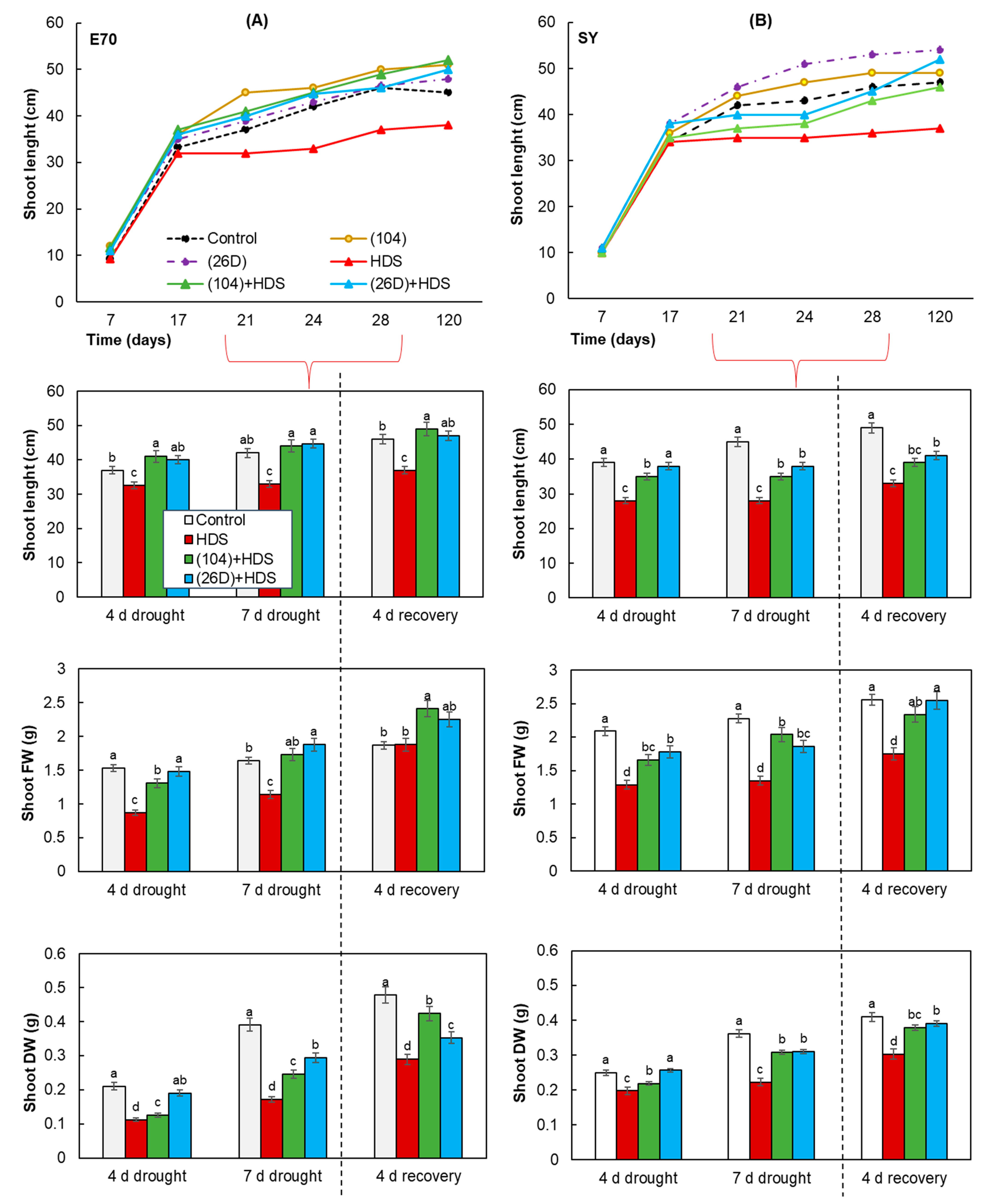
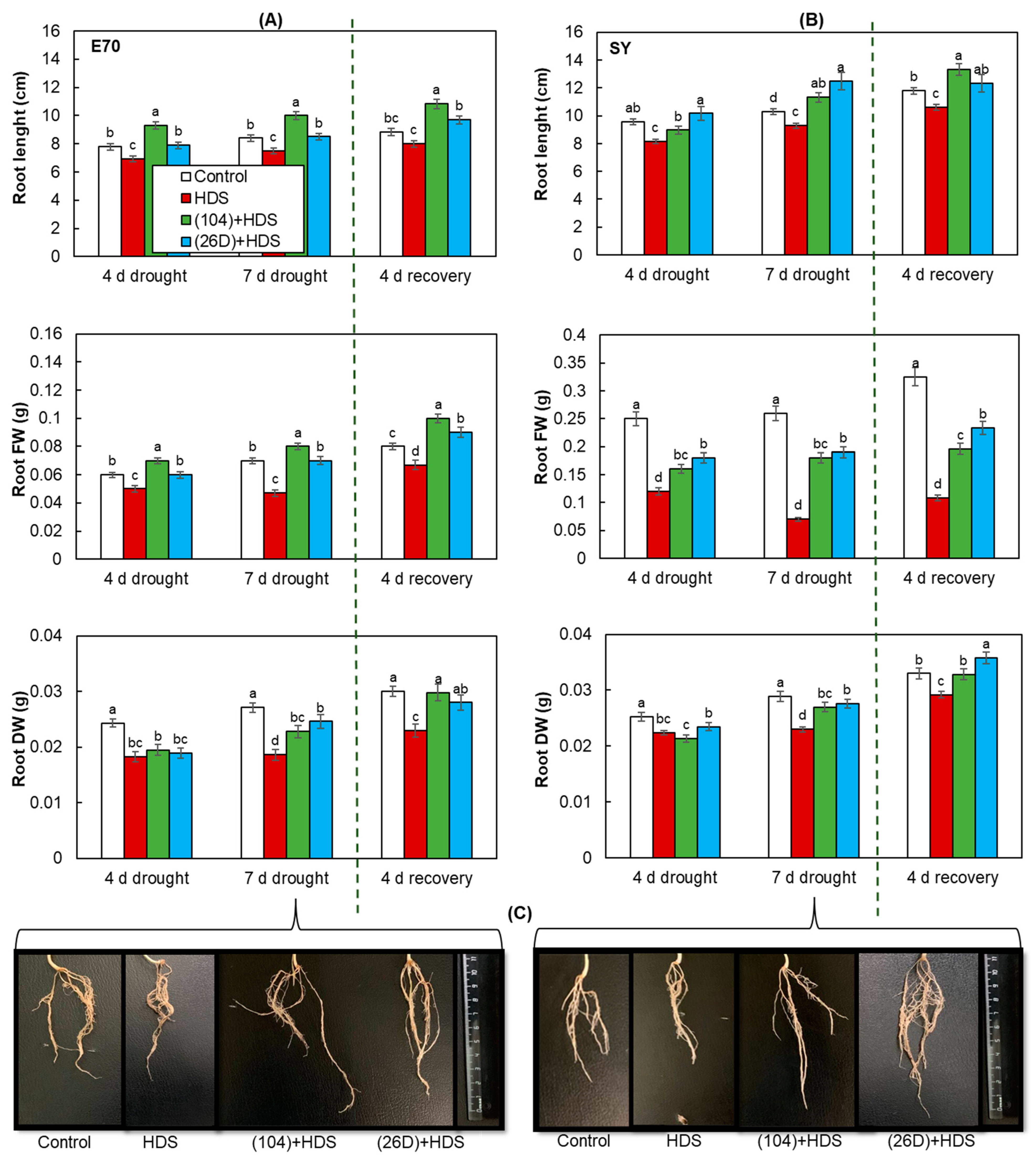
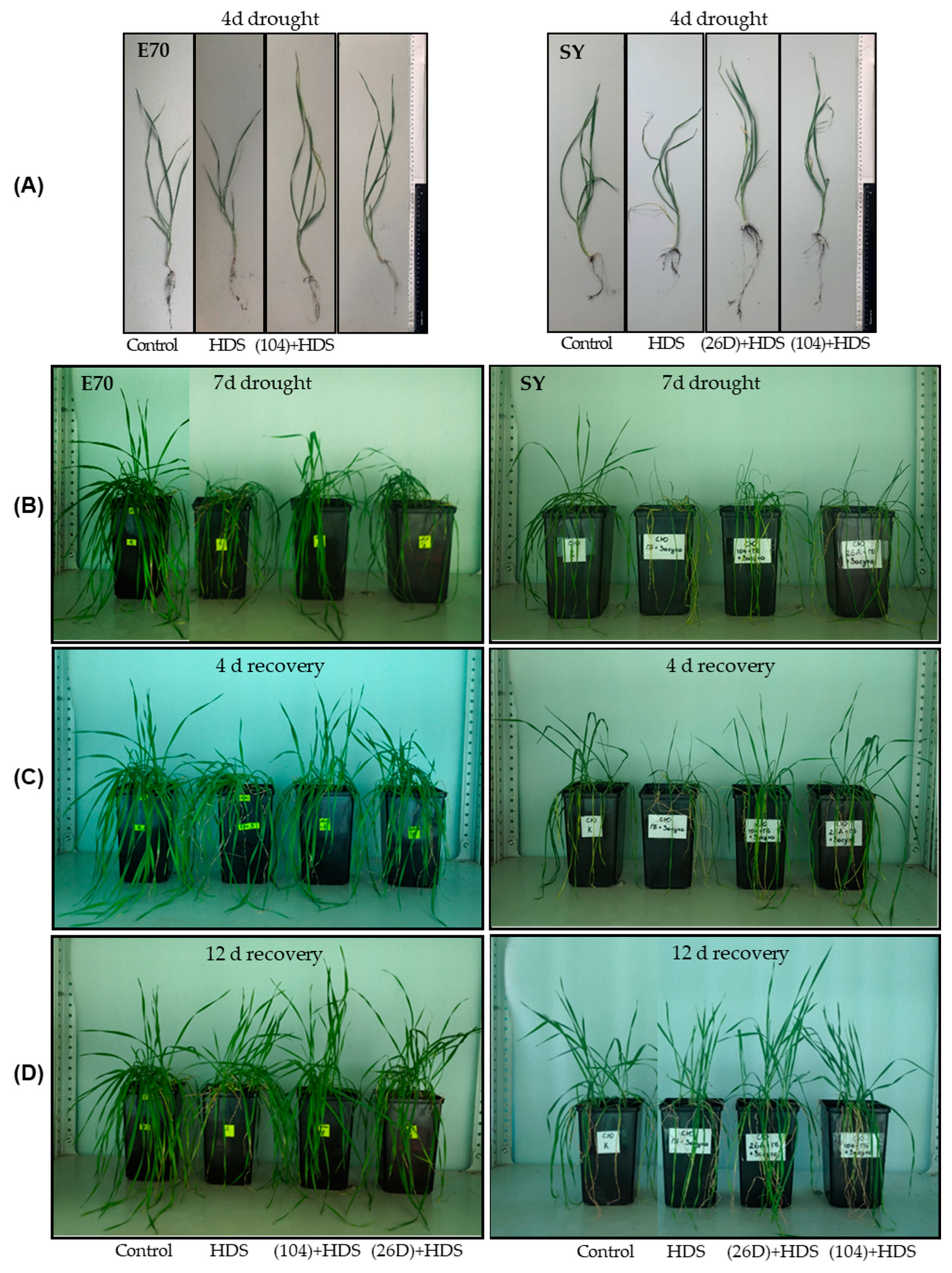
2.3. Growth of Wheat Plants Sprayed with Herbicide Sekator® Turbo and Exposed to Soil Drought in Pot Experiments
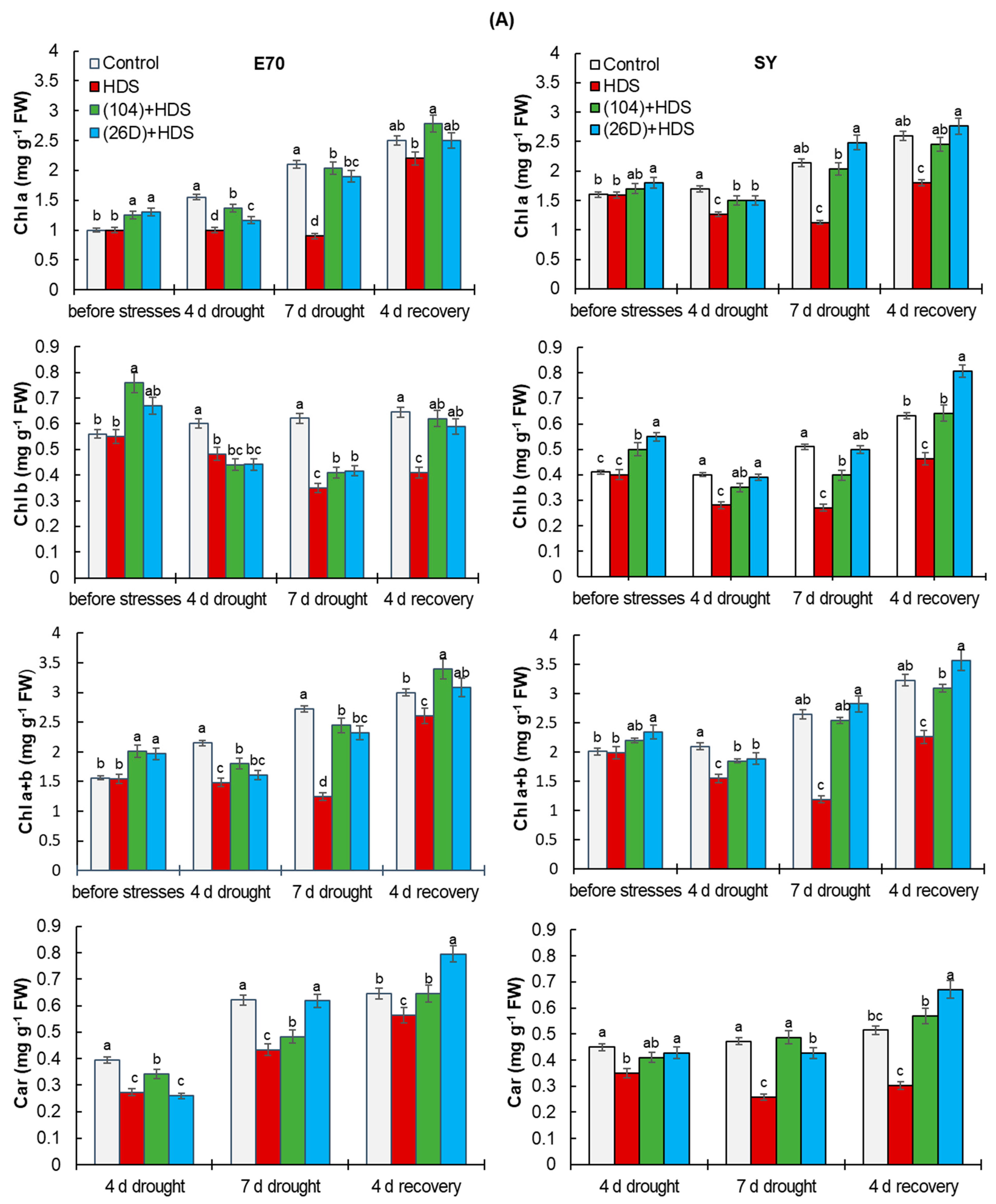
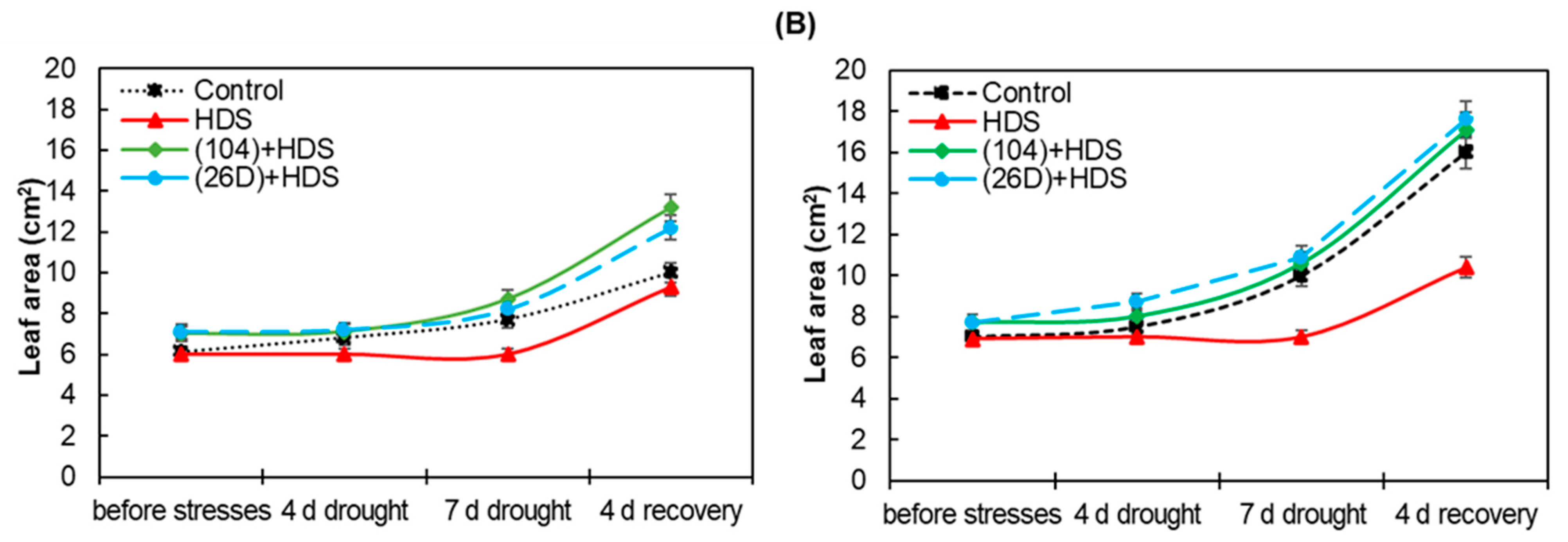
2.4. Chlorophyll Content and Leaf Area
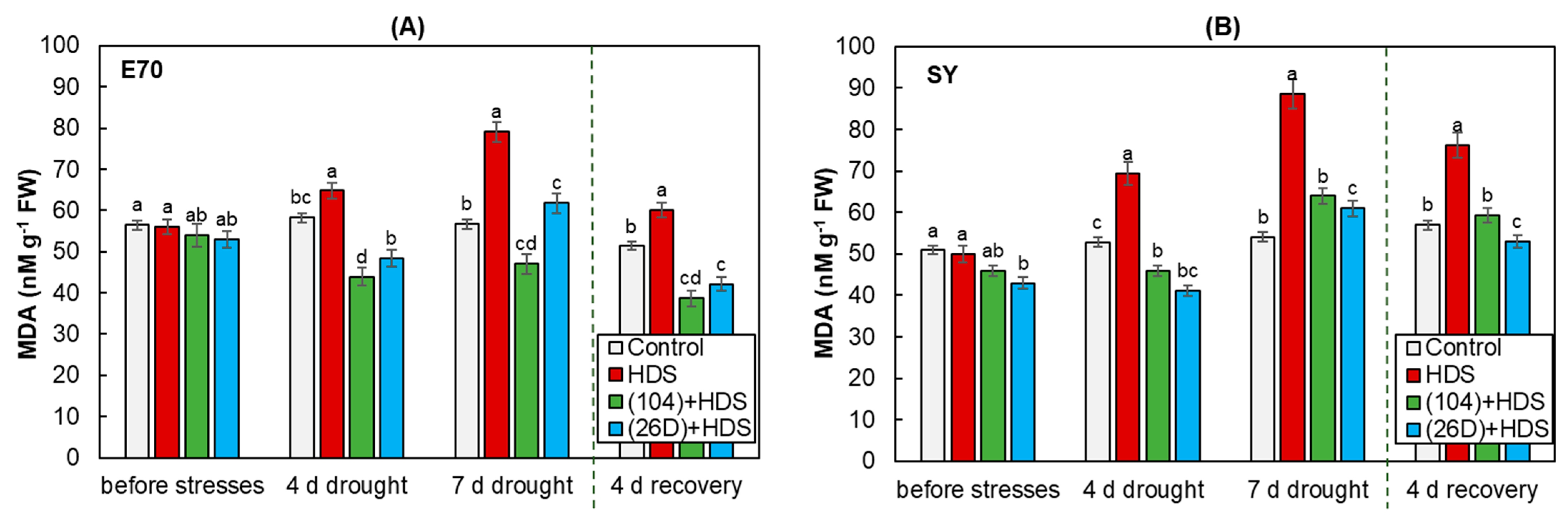
2.5. Lipid Peroxidation (Malondialdehyde Content)
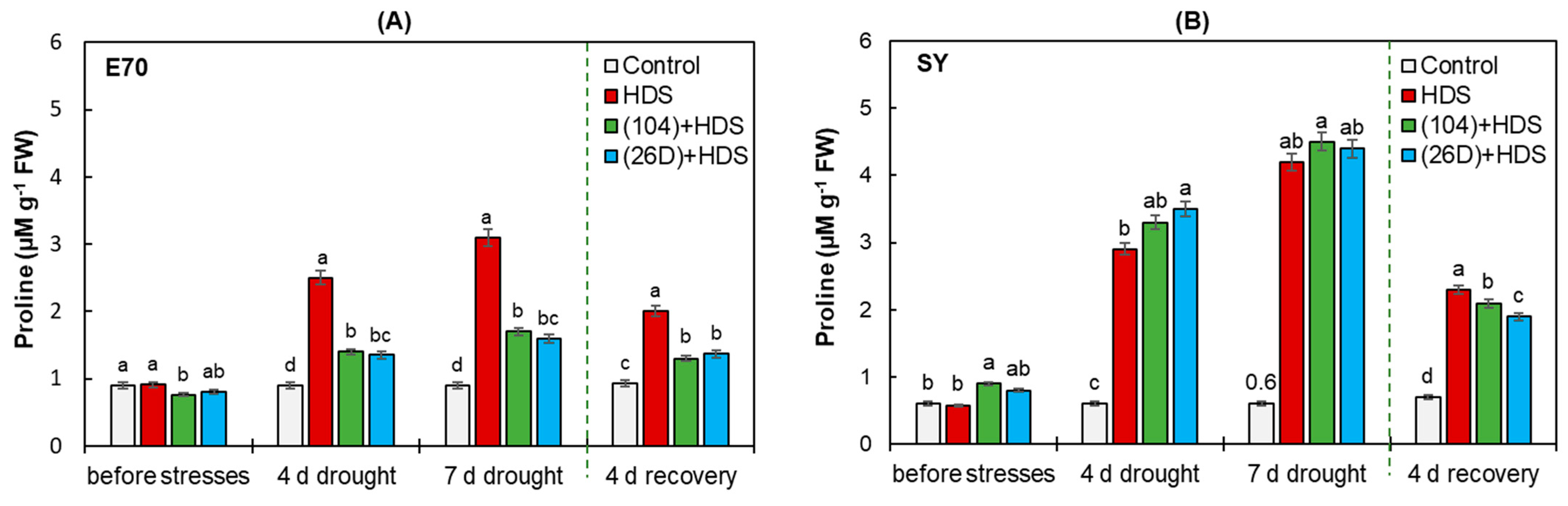
2.6. Proline Concentration
2.7. Grain Yield Parameters
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Bacterial Strains
4.2. Preparation of Bacterial Inoculum Suspensions and Seed Priming
4.3. Herbicide
4.4. Herbicide Stress Tolerance of Bacterial Strains
4.5. Drought Stress Tolerance of Bacterial Strains
4.6. In Vitro Seed Germination and Early Hydroponic Seedling Growth Assays
4.7. Pot Experiment Design, Treatments, and Growth Conditions
4.8. Determination of Photosynthetic Pigments and Leaf Area
4.9. Lipid Peroxidation (LPO) Assay
4.10. Determination of Proline Content
4.11. Evaluation of Grain Yield
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asseng, S.; Martre, P.; Maiorano, A.; Rötter, R.P.; O’Leary, G.J.; Fitzgerald, G.J.; Girousse, C.; Motzo, R.; Giunta, F.; Babar, M.A.; et al. Climate Change Impact and Adaptation for Wheat Protein. Glob. Chang. Biol. 2019, 25, 155–173. [Google Scholar] [CrossRef] [PubMed]
- Cramer, G.R.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of Abiotic Stress on Plants: A Systems Biology Perspective. BMC Plant Biol. 2011, 11, 163. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. In Agronomy for Sustainable Development; Springer: Berlin/Heidelberg, Germany, 2009; pp. 153–188. [Google Scholar]
- Ahmad, Z.; Waraich, E.A.; Akhtar, S.; Anjum, S.; Ahmad, T.; Mahboob, W.; Hafeez, O.B.A.; Tapera, T.; Labuschagne, M.; Rizwan, M. Physiological Responses of Wheat to Drought Stress and Its Mitigation Approaches. Acta Physiol. Plant. 2018, 40, 80. [Google Scholar] [CrossRef]
- Carvalho, M.; Gouvinhas, I.; Castro, I.; Matos, M.; Rosa, E.; Carnide, V.; Barros, A. Drought Stress Effect on Polyphenolic Content and Antioxidant Capacity of Cowpea Pods and Seeds. J. Agron. Crop Sci. 2021, 207, 197–207. [Google Scholar] [CrossRef]
- Verma, S.K. Integrated Weed Management in Wheat (Triticum aestivum L.): A Review. Curr. Res. Agric. Farming 2021, 2, 1–4. [Google Scholar] [CrossRef]
- Todorova, D.; Sergiev, I.; Brankova, L.; Katerova, Z.; Shopova, E.; Dimitrova, L. Assessment of the Biochemical Responses of Wheat Seedlings to Soil Drought after Application of Selective Herbicide. Plants 2021, 10, 733. [Google Scholar] [CrossRef] [PubMed]
- Todorova, D.; Aleksandrov, V.; Anev, S.; Sergiev, I. Photosynthesis Alterations in Wheat Plants Induced by Herbicide, Soil Drought or Flooding. Agronomy 2022, 12, 390. [Google Scholar] [CrossRef]
- Łozowicka, B.; Wołejko, E.; Kaczyński, P.; Konecki, R.; Iwaniuk, P.; Drągowski, W.; Jablońska-Trypuć, A. Effect of Microorganism on Behaviour of Two Commonly Used Herbicides in Wheat/Soil System. Appl. Soil Ecol. 2021, 162, 103879. [Google Scholar] [CrossRef]
- Varshney, S.; Hayat, S.; Alyemeni, M.N.; Ahmad, A. Effects of Herbicide Applications in Wheat Fields. Plant Signal. Behav. 2012, 7, 570–575. [Google Scholar] [CrossRef]
- Benevenuto, R.F.; Zanatta, C.B.; Guerra, M.P.; Nodari, R.O.; Agapito-Tenfen, S.Z. Proteomic Profile of Glyphosate-Resistant Soybean under Combined Herbicide and Drought Stress Conditions. Plants 2021, 10, 2381. [Google Scholar] [CrossRef]
- Vikrant; Jasmine, A.; Roobavathi, M.R. Impacts of Drought and Herbicide Stress on Seed Germination and Early Seedling Growth in a Legume Crop Mung Bean (Vigna Radiata L.). J. Plant Stress Physiol. 2021, 7, 32–41. [Google Scholar] [CrossRef]
- Radchenko, M.; Ponomareva, I.; Pozynych, I.; Morderer, Y. Stress and Use of Herbicides in Field Crops. Agric. Sci. Pract. 2021, 8, 50–70. [Google Scholar] [CrossRef]
- Pascual, L.S.; Segarra-Medina, C.; Gómez-Cadenas, A.; López-Climent, M.F.; Vives-Peris, V.; Zandalinas, S.I. Climate Change-Associated Multifactorial Stress Combination: A Present Challenge for Our Ecosystems. J. Plant Physiol. 2022, 276, 153764. [Google Scholar] [CrossRef] [PubMed]
- Lastochkina, O.; Aliniaeifard, S.; Kalhor, M.S.; Yuldashev, R.; Pusenkova, L.; Garipova, S. Plant Growth Promoting Bacteria Biotic Strategy to Cope with Abiotic Stresses in Wheat. In Wheat Production in Changing Environments: Management, Adaptation and Tolerance; Hasanuzzaman, M., Nahar, K., Hossain, A., Eds.; Springer: Singapore, 2019; pp. 579–614. [Google Scholar]
- Pathma, J.; Kennedy, R.K.; Bhushan, L.S.; Shankar, B.K.; Thakur, K. Microbial Biofertilizers and Biopesticides: Nature’s Assets Fostering Sustainable Agriculture. In Recent Developments in Microbial Technologies; Prasad, R., Kumar, V., Singh, J., Upadhyaya, C.P., Eds.; Springer: Singapore, 2021; pp. 39–69. [Google Scholar]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; AL-Harrasi, A. Plant Growth Promoting Bacteria as an Alternative Strategy for Salt Tolerance in Plants: A Review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant Survival under Drought Stress: Implications, Adaptive Responses, and Integrated Rhizosphere Management Strategy for Stress Mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef]
- Tataridas, A.; Kanatas, P.; Chatzigeorgiou, A.; Zannopoulos, S.; Travlos, I. Sustainable Crop and Weed Management in the Era of the EU Green Deal: A Survival Guide. Agronomy 2022, 12, 589. [Google Scholar] [CrossRef]
- Sood, G.; Kaushal, R.; Sharma, M. Significance of Inoculation with Bacillus subtilis to Alleviate Drought Stress in Wheat (Triticum aestivum L.). Vegetos 2020, 33, 782–792. [Google Scholar] [CrossRef]
- Lastochkina, O.; Seifikalhor, M.; Aliniaeifard, S.; Baymiev, A.; Pusenkova, L.; Garipova, S.; Kulabuhova, D.; Maksimov, I. Bacillus spp.: Efficient Biotic Strategy to Control Postharvest Diseases of Fruits and Vegetables. Plants 2019, 8, 97. [Google Scholar] [CrossRef]
- Fouda, A.; Hassan, S.E.D.; Eid, A.M.; El-Din Ewais, E. The Interaction Between Plants and Bacterial Endophytes under Salinity Stress. In Endophytes and Secondary Metabolites; Jha, S., Ed.; Reference Series in Phytochemistry; Springer: Cham, Switzerland, 2019; pp. 591–607. [Google Scholar]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef]
- Nanda, S.; Mohanty, B.; Joshi, R.K. Endophyte-Mediated Host Stress Tolerance as a Means for Crop Improvement. In Endophytes and Secondary Metabolites; Jha, S., Ed.; Reference Series in Phytochemistry; Springer: Cham, Switzerland, 2019; pp. 677–701. [Google Scholar] [CrossRef]
- Lastochkina, O.V. Adaptation and Tolerance of Wheat Plants to Drought Mediated by Natural Growth Regulators Bacillus spp.: Mechanisms and Practical Importance. Sel’skokhozyaistvennaya Biol. 2021, 56, 843–867. [Google Scholar] [CrossRef]
- Benedetto, N.A.; Corbo, M.R.; Campaniello, D.; Cataldi, M.P.; Bevilacqua, A.; Sinigaglia, M.; Flagella, Z. The Role of Plant Growth Promoting Bacteria in Improving Nitrogen Use Efficiency for Sustainable Crop Production: A Focus on Wheat. AIMS Microbiol. 2017, 3, 413. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Daim, I.A.; Bejai, S.; Fridborg, I.; Meijer, J. Identifying Potential Molecular Factors Involved in Bacillus Amyloliquefaciens 5113 Mediated Abiotic Stress Tolerance in Wheat. Plant Biol. 2018, 20, 271. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Daim, I.A.; Bejai, S.; Meijer, J. Bacillus Velezensis 5113 Induced Metabolic and Molecular Reprogramming During Abiotic Stress Tolerance in Wheat. Sci. Rep. 2019, 9, 16282. [Google Scholar] [CrossRef] [PubMed]
- Blake, C.; Christensen, M.N.; Kovács, Á. Molecular Aspects of Plant Growth Promotion and Protection by Bacillus subtilis. MPMI 2021, 34, 15. [Google Scholar] [CrossRef]
- Eichmann, R.; Richards, L.; Schäfer, P. Hormones as Go-Betweens in Plant Microbiome Assembly. Plant J. 2021, 105, 518. [Google Scholar] [CrossRef]
- Shobana, N.; Sugitha, T.; Sivakumar, U. Plant Growth-Promoting Bacillus sp. Cahoots Moisture Stress Alleviation in Rice Genotypes by Triggering Antioxidant Defense System. Microbiol. Res. 2020, 239, e126518. [Google Scholar] [CrossRef]
- Lastochkina, O.; Aliniaeifard, S.; Garshina, D.; Garipova, S.; Pusenkova, L.; Allagulova, C.; Fedorova, K.; Baymiev, A.; Koryakov, I.; Sobhani, M. Seed Priming with Endophytic Bacillus subtilis Strain-Specifically Improves Growth of Phaseolus vulgaris Plants under Normal and Salinity Conditions and Exerts Anti-Stress Effect through Induced Lignin Deposition in Roots and Decreased Oxidative and Osmotic Stresses. J. Plant Physiol. 2021, 263, 153462. [Google Scholar] [CrossRef]
- Lastochkina, O. Bacillus subtilis-Mediated Abiotic Stress Tolerance in Plants. In Bacilli and Agrobiotechnology: Phytostimulation and Biocontrol; Islam, M.T., Rahman, M.M., Pandey, P., Boehme, M.H., Haesaert, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; p. 97. [Google Scholar] [CrossRef]
- Barnawal, D.; Bharti, N.; Pandey, S.S.; Pandey, A.; Chanotiya, C.S.; Kalra, A. Plant Growth Promoting Rhizobacteria Enhances Wheat Salt and Drought Stress Tolerance by Altering Endogenous Phytohormone Levels and TaCTR1/TaDREB2 Expression. Physiol. Plant. 2017, 161, 502. [Google Scholar] [CrossRef]
- Lastochkina, O.; Pusenkova, L.; Yuldashev, R.; Babaev, M.; Garipova, S.; Blagova, D.; Khairullin, R.; Aliniaeifard, S. Effects of Bacillus subtilis On Some Physiological and Biochemical Parameters of Triticum aestivum L. (Wheat) under Salinity. Plant Physiol. Biochem. 2017, 121, 80. [Google Scholar] [CrossRef]
- Salem, G.; Stromberger, M.E.; Byrne, P.F.; Manter, D.K.; El-Feki, W.; Weir, T.L. Genotype-Specific Response of Winter Wheat (Triticum aestivum L.) to Irrigation and Inoculation with ACC Deaminase Bacteria. Rhizosphere 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Lastochkina, O.; Garshina, D.; Ivanov, S.; Yuldashev, R.; Khafizova, R.; Allagulova, C.; Fedorova, K.; Avalbaev, A.; Maslennikova, D.; Bosacchi, M. Seed Priming with Endophytic Bacillus subtilis Modulates Physiological Responses of Two Different Triticum aestivum L. Cultivars under Drought Stress. Plants 2020, 9, 1810. [Google Scholar] [CrossRef] [PubMed]
- Bourahla, M.; Djebbar, R.; Kaci, Y.; Abrous-Belbachir, O. Alleviation of Bleaching Herbicide Toxicity by PGPR Strain Isolated from Wheat Rhizosphere. An. Univ. Din Oradea Fasc. Biol. 2018, 25, 42–51. [Google Scholar]
- Shahid, M.; Khan, M.S. Glyphosate Induced Toxicity to Chickpea Plants and Stress Alleviation by HerbicideTolerant Phosphate Solubilizing Burkholderia cepacia PSBB1 Carrying Multifarious Plant Growth Promoting Activities. Biotech 2018, 8, 131. [Google Scholar] [CrossRef]
- Korobov, A.E.; Lelyak, V.A.; Ivleva, A.A. Spring Wheat Protection Against Herbicide Stress by Biological Preparation Phytop 8.67. In Proceedings of the International Scientific Conference Plant Protection in the Transition to Precision Farming, Minsk, Belarus, 27–29 July 2021; National Academy of Sciences: Washington, DC, USA, 2021; Volume 204. [Google Scholar]
- Mironenka, J.; Różalska, S.; Bernat, P. Potential of Trichoderma harzianum and Its Metabolites to Protect Wheat Seedlings against Fusarium culmorum and 2,4-D. Int. J. Mol. Sci. 2021, 22, 3058. [Google Scholar] [CrossRef] [PubMed]
- Bernat, P.; Nykiel-Szymańska, J.; Gajewska, E.; Różalska, S.; Stolarek, P.; Dackowa, J.; Słaba, M. Trichoderma harzianum Diminished Oxidative Stress Caused by 2,4-Dichlorophenoxyacetic Acid (2,4-D) in Wheat, with Insights from Lipidomics. J. Plant Physiol. 2018, 229, 158–163. [Google Scholar] [CrossRef]
- Chetverikov, S.P.; Timergalin, M.D.; Rameev, T.V.; Feoktistova, A.V.; Chetverikova, D.V.; Kendzhieva, A.A.; Sharipov, D.A.; Starikov, S.N.; Bakaeva, M.D.; Hudaygulov, G.G.; et al. A New «Agrobiolog» Biopreparation to Improve the Quality of Soft Spring Wheat Grain in Conditions of Drought and Herbicidal Stress. Nat. Tech. Sci. 2022, 6, 47–50. [Google Scholar] [CrossRef]
- Chennappa, G.; Sreenivasa, M.Y.; Nagaraja, H. Azotobacter Salinestris: A Novel Pesticide-Degrading and Prominent Biocontrol PGPR Bacteria. In Microorganisms for Green Revolution. Microorganisms for Sustainability; Panpatte, D., Jhala, Y., Shelat, H., Vyas, R., Eds.; Springer: Singapore, 2018; pp. 23–43. [Google Scholar] [CrossRef]
- Chetverikov, S.; Sharipov, D.; Bakaeva, M.; Chetverikova, D.; Timergalin, M.; Rameev, T. Bacterial Strain Pseudomonas Avellanae 6CH2 with Antifusarium Activity in Mitigation of Herbicidal Stress in Wheat Plants. Jordan J. Biol. Sci. 2021, 14, 805–812. [Google Scholar]
- Timergalin, M.D.; Feoktistova, A.V.; Rameev, T.V.; Kudoyarova, G.R.; Chetverikov, S.P. Role of Aauxin-Producing Bacteria in Overcoming the Stress of Wheat Plants Under Treatment with the Herbicide Chistalan. Agrochemistry 2020, 11, 35–40. [Google Scholar]
- Feoktistova, A.V.; Timergalin, M.D.; Rameev, T.V.; Chetverikov, S.P. Treatment of Wheat Plants with Bacteria Pseudomonas Protegens DA 1.2 Reduced the Negative Effect of the Herbicide Chistalan in Conditions of Water Deficit. Agrochemistry 2021, 10, 89–96. [Google Scholar] [CrossRef]
- Cherif, H.; Marasco, R.; Rolli, E.; Ferjani, R.; Fusi, M.; Soussi, A.; Mapelli, F.; Blilou, I.; Borin, S.; Boudabous, A.; et al. Oasis Desert Farming Selects Environment-Specific Date Palm Root Endophytic Communities and Cultivable Bacteria That Promote Resistance to Drought. Environ. Microbiol. Rep. 2015, 7, 668–678. [Google Scholar] [CrossRef]
- Chakraborty, U.; Chakraborty, B.N.; Chakraborty, A.P.; Dey, P.L. Water Stress Amelioration and Plant Growth Promotion in Wheat Plants by Osmotic Stress Tolerant Bacteria. World J. Microbiol. Biotechnol. 2013, 29, 789–803. [Google Scholar] [CrossRef]
- Arzanesh, M.H.; Alikhani, H.A.; Khavazi, K.; Rahimian, H.A.; Miransari, M. Wheat (Triticum aestivum L.) Growth Enhancement by Azospirillum sp. under Drought Stress. World J. Microbiol. Biotechnol. 2011, 27, 197–205. [Google Scholar] [CrossRef]
- Çakmakçı, R.; Turan, M.; Kıtır, N.; Güneş, A.; Nikerel, E.; Özdemir, B.S.; Yıldırım, E.; Olgun, M.; Topçuoğlu, B.; Tüfenkçi, Ş.; et al. The Role of Soil Beneficial Bacteria in Wheat Production: A Review. In Wheat Improvement, Management and Utilization; Wanyera, R., Owuoche, J., Eds.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Hussain, M.B.; Hussain, M.B.; Zahir, Z.A.; Asghar, H.N.; Asgher, M. Can Catalase and Exopolysaccharides Producing Rhizobia Ameliorate Drought Stress in Wheat? Int. J. Agric. Biol. 2014, 16, 3–13. [Google Scholar]
- Chen, C.; Xin, K.; Liu, H.; Cheng, J.; Shen, X.; Wang, Y.; Zhang, L. Pantoea alhagi, a Novel Endophytic Bacterium with Ability to Improve Growth and Drought Tolerance in Wheat. Sci. Rep. 2017, 7, 41564. [Google Scholar] [CrossRef]
- Chetverikov, S.P. Selection of Anti-Stress Bacterial Agents for the Protection of Agricultural Plants. Nat. Tech. Sci. 2019, 11, 81–84. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Rashid, U.; Yasmin, H.; Hassan, M.N.; Naz, R.; Nosheen, A.; Sajjad, M.; Ilyas, N.; Keyani, R.; Jabeen, Z.; Mumtaz, S.; et al. Drought-Tolerant Bacillus megaterium Isolated from Semi-Arid Conditions Induces Systemic Tolerance of Wheat under Drought Conditions. Plant Cell Rep. 2021, 41, 549–569. [Google Scholar] [CrossRef] [PubMed]
- Puthiyottil, P.; Akkara, Y. Pre-Treatment with Bacillus subtilis Mitigates Drought Induced Photo-Oxidative Damages in Okra by Modulating Antioxidant System and Photochemical Activity. Physiol. Mol. Biol. Plants 2021, 27, 945–957. [Google Scholar] [CrossRef]
- Batool, T.; Ali, S.; Seleiman, M.F.; Naveed, N.H.; Ali, A.; Ahmed, K. Plant Growth Promoting Rhizobacteria Alleviates Drought Stress in Potato in Response to Suppressive Oxidative Stress and Antioxidant Enzymes Activities. Sci. Rep. 2020, 10, 16975. [Google Scholar] [CrossRef]
- Rare, E. Stress Physiology: The Functional Significance of the Accumulation of Nitrogen-Containing Compounds. J. Horticult. Sci. 1990, 65, 231–243. [Google Scholar] [CrossRef]
- Mansour, M.M.F. Nitrogen Containing Compounds and Adaptation of Plants to Salinity Stress. Biol. Plant. 2000, 43, 491–500. [Google Scholar] [CrossRef]
- Kavi, K.P.B.; Hima, K.P.; Sunita, M.S.; Sreenivasulu, N. Role of Proline in Cell Wall Synthesis and Plant Development and its Implications in Plant Ontogeny. Front. Plant Sci. 2015, 66, e544. [Google Scholar] [CrossRef]
- Guan, C.; Cen, H.F.; Cui, X.; Tian, D.Y.; Tadesse, D.; Zhang, Y.W. Proline Improves Switchgrass Growth and Development by Reduced Lignin Biosynthesis. Sci. Rep. 2019, 9, e20117. [Google Scholar] [CrossRef]
- Vardharajula, S.; Zulfikar, A.S.; Grover, M.; Reddy, G.; Bandi, V. Drought-Tolerant Plant Growth Promoting Bacillus spp., Effect on Growth, Osmolytes, and Antioxidant Status of Maize under Drought Stress. J. Plant Interact. 2011, 6, 1–14. [Google Scholar] [CrossRef]
- Boleta, E.H.M.; Shintate Galindo, F.; Jalal, A.; Santini, J.M.K.; Rodrigues, W.L.; de Lima, B.H.; Arf, O.; Silva, M.R.; Buzetti, S.; Teixeira Filho, M.C.M. Inoculation with Growth-Promoting Bacteria Azospirillum brasilense and its Effects on Productivity and Nutritional Accumulation of Wheat Cultivars. Front. Sustain. Food Syst. 2020, 4, 1–10. [Google Scholar] [CrossRef]
- Alferov, A.A.; Chernova, L.S.; Zavalin, A.A.; Chebotar, V.K. The Efficiency of the Endophytic Biopreparations and Nitrogen Fertilizer Application. Bull. Russ. Agric. Sci. 2017, 5, 21–24. [Google Scholar]
- Chandra, D.; Srivastava, R.; Gupta, V.V.S.R.; Franco, C.M.M.; Paasricha, N.; Saifi, S.K.; Tuteja, N.; Sharma, A.K. Field Performance of Bacterial Inoculants to Alleviate Water Stress Effects in Wheat (Triticum aestivum L.). Plant Soil 2019, 441, 261–281. [Google Scholar] [CrossRef]
- Mokronosova, A.T. Small Workshop on Plant Physiology; Moscow State University: Moscow, Russia, 1994; p. 184. [Google Scholar]
- Jeffrey, S.; Humphrey, G. New Spectrophotometric Equations for Determining Chlorophylls A, B, C1 and C2 in Higher Plants, Algae and Natural Phytoplankton. Biochem. Physiol. Pfl. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Bioph. 1968, 125, 189. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldern, R.P.; Teare, D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205. [Google Scholar] [CrossRef]
- Vedrov, N.G.; Halipsky, A.N. Change in the Structure of Cell Yield and Economic Indicators of Biological-Long as a Result of the Change of Spring Wheat in Krasnodarsk Region. Bull. KSAU 2012, 4, 89–93. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration of PEG (%) | 3rd Day | 7th Day | Shoot Length (cm) | Root Length (cm) | ||||
|---|---|---|---|---|---|---|---|---|
| Germination (%) | Germination (%) | |||||||
| E70 | SY | E70 | SY | E70 | SY | E70 | SY | |
| 0 (control) | 99 ± 1 a | 80 ± 3 a | 99 ± 1 a | 82 ± 2 a | 6.8 ± 0.4 a | 5.3 ± 0.3 a | 5.8 ± 0.2 a | 5.4 ± 0.1 a |
| 4 | 84 ± 2 d | 73 ± 2 c | 84 ± 2 d | 73 ± 2 c | 5.7 ± 0.5 c | 4.9 ± 0.2 c | 5.2 ± 0.1 c | 4.8 ± 0.2 d |
| 4 + (strain 104) | 93 ± 2 b | 80 ± 1 a | 95 ± 1 b | 82 ± 1 ab | 6.2 ± 0.3 b | 5.0 ± 0.3 bc | 5.9 ± 0.3 a | 4.9 ± 0.3 c |
| 4 + (strain 26D) | 86 ± 3 c | 76 ± 2 b | 88 ± 3 c | 78 ± 5 b | 6.1 ± 0.2 b | 5.1 ± 0.2 b | 5.6 ± 0.2 b | 5.2 ± 0.4 b |
| 8 | 72 ± 1 g | 67 ± 4 e | 73 ± 2 e | 67 ± 2 e | 4.9 ± 0.1 e | 4.0 ± 0.5 f | 4.3 ± 0.3 e | 3.7 ± 0.2 f |
| 8 + (strain 104) | 87 ± 2 e | 70 ± 1 d | 88 ± 1 c | 70 ± 3 d | 5.3 ± 0.6 d | 4.5 ± 0.3 de | 5.1 ± 0.4 c | 4.2 ± 0.3 e |
| 8 + (strain 26D) | 84 ± 2 f | 69 ± 1 d | 84 ± 2 d | 69 ± 4 d | 5.0 ± 0.7 de | 4.6 ± 0.2 d | 4.9 ± 0.1 d | 4.8 ± 0.4 d |
| 12 | 68 ± 3 k | 60 ± 2 g | 68 ± 1 g | 60 ± 1 j | 3.8 ± 0.5 j | 3.1 ± 0.4 j | 3.8 ± 0.2 g | 2.9 ± 0.3 j |
| 12 + (strain 104) | 72 ± 1 g | 63 ± 3 f | 73 ± 3 e | 64 ± 2 f | 4.2 ± 0.6 f | 3.7 ± 0.1 g | 4.4 ± 0.5 e | 3.2 ± 0.2 g |
| 12 + (strain 26D) | 70 ± 2 j | 62 ± 1 f | 71 ± 2 f | 62 ± 3 g | 4.0 ± 0.3 fg | 3.9 ± 0.1 f | 4.1 ± 0.3 f | 3.7 ± 0.2 f |
| Concentration of Sekator® Turbo | 3rd day | 7th day | Shoot Length (cm) | Root Length (cm) | ||||
| Germination (%) | Germination (%) | |||||||
| E70 | SY | E70 | SY | E70 | SY | E70 | SY | |
| Hb0 (control) | 98 ± 2 a | 86 ± 4 a | 98 ± 1 a | 86 ± 3 a | 4.2 ± 0.2 a | 4.6 ± 0.1 a | 6.1 ± 0.2 a | 6.6 ± 0.3 a |
| Hb50 | 53 ± 4 d | 27 ± 5 f | 55 ± 2 f | 25 ± 5 f | 1.7 ± 0.1 d | 0.5 ± 0.3 d | 0.9 ± 0.1 e | 0.1 ± 0.05 f |
| Hb50 + (strain 104) | 76 ± 3 b | 67 ± 2 b | 78 ± 1 b | 67 ± 2 b | 2.2 ± 0.3 b | 0.9 ± 0.2 bc | 1.5 ± 0.2 b | 0.8 ± 0.1 c |
| Hb50 + (strain 26D) | 67 ± 5 c | 53 ± 3 c | 68 ± 3 d | 54 ± 2 c | 1.9 ± 0.2 c | 1.0 ± 0.1 b | 1.2 ± 0.2 c | 1.3 ± 0.1 b |
| Hb75 | 46 ± 1 f | 23 ± 4 g | 47 ± 4 g | 14 ± 1 k | 1.3 ± 0.1 f | 0.3 ± 0.1 e | 0.7 ± 02 f | 0.05 ± 0.01 g |
| Hb75 + (strain 104) | 73 ± 3 bc | 46 ± 2 d | 76 ± 2 c | 49 ± 1 d | 1.7 ± 0.1 d | 0.5 ± 0.1 d | 1.4 ± 0.1 bc | 0.3 ± 0.1 d |
| Hb75 + (strain 26D) | 60 ± 2 d | 33 ± 3 e | 63 ± 3 e | 37 ± 4 e | 1.5 ± 0.1 e | 0.4 ± 0.2 d | 1.1 ± 0.3 d | 0.2 ± 0.04 e |
| Hb100 | 37 ± 2 j | 14 ± 1 k | 41 ± 2 k | 15 ± 2 k | 0.5 ± 0.2 k | 0.07 ± 0.03 g | 0.4 ± 0.1 g | 0.03 ± 0.01 j |
| Hb100 + (strain 104) | 48 ± 4 e | 19 ± 3 j | 48 ± 1 g | 19 ± 3 j | 1.0 ± 0.3 g | 0.2 ± 0.1 f | 1.0 ± 0.1 de | 0.1 ± 0.02 e |
| Hb100 + (strain 26D) | 43 ± 3 g | 23 ± 4 g | 43 ± 1 j | 24 ± 2 fg | 0.7 ± 0.1 h | 0.3 ± 0.1 e | 0.8 ± 0.2 ef | 0.2 ± 0.1 e |
| Treatment | Plant Length (cm) | Spike Length (cm) | Grains Number per Spike | Mass of 1000 Grains (g) | Grain Yield (g per Plant) |
|---|---|---|---|---|---|
| E70 variety | |||||
| Control | 45 ± 1.1 d | 4.8 ± 0.7 b | 8.1 ± 0.3 d | 23.7 ± 2.7 c | 0.19 ± 0.03 e |
| 104 | 51 ± 0.3 a | 5.0 ± 0.9 a | 11.7 ± 0.5 a | 25.6 ± 1.3 a | 0.30 ± 0.02 a |
| 26D | 48 ± 0.6 bc | 4.9 ± 0.7 a | 10.8 ± 0.4 b | 24.4 ± 1.5 b | 0.26 ± 0.04 b |
| HDS | 38 ± 0.9 e | 4.1 * ± 0.4 d | 6.3 ± 0.3 e | 15.1 ± 2.6 e | 0.11 ± 0.06 f |
| 104 + HDS | 52 ± 0.7 a | 4.7 ± 0.3 bc | 10.2 ± 0.6 b | 23.5 ± 1.9 c | 0.24 ± 0.04 c |
| 26D + HDS | 50 ± 0.8 b | 4.5 ± 0.6 c | 9.9 ± 0.7 c | 22.3 ± 2.1 d | 0.22 ± 0.03 d |
| SY variety | |||||
| Control | 47 ± 0.8 d | 5.6 ± 1.3 c | 6.2 ± 1.7 d | 21.2 ± 1.6 c | 0.13 ± 0.02 c |
| 104 | 49 ± 0.9 c | 5.7 ± 0.9 b | 7.1 ± 1.1 b | 22.4 ± 0.6 b | 0.16 ± 0.01 b |
| 26D | 54 ± 0.4 a | 5.9 ± 0.6 a | 8.4 ± 1.9 a | 23.6 ± 1.3 a | 0.20 ± 0.02 a |
| HDS | 37 ± 0.7 e | 3.4 * ± 1.3 f | 3.5 ± 0.7 f | 14.2 ± 1.5 f | 0.05 ± 0.03 e |
| 104 + HDS | 46 ± 0.6 d | 4.5 ± 0.9 e | 5.6 ± 1.0 e | 17.2 ± 0.7 de | 0.10 ± 0.01 d |
| 26D + HDS | 52 ± 0.2 b | 5.4 ± 1.2 d | 6.7 ± 1.5 c | 18.3 ± 0.3 d | 0.13 ± 0.01 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lastochkina, O.; Yakupova, A.; Avtushenko, I.; Lastochkin, A.; Yuldashev, R. Effect of Seed Priming with Endophytic Bacillus subtilis on Some Physio-Biochemical Parameters of Two Wheat Varieties Exposed to Drought after Selective Herbicide Application. Plants 2023, 12, 1724. https://doi.org/10.3390/plants12081724
Lastochkina O, Yakupova A, Avtushenko I, Lastochkin A, Yuldashev R. Effect of Seed Priming with Endophytic Bacillus subtilis on Some Physio-Biochemical Parameters of Two Wheat Varieties Exposed to Drought after Selective Herbicide Application. Plants. 2023; 12(8):1724. https://doi.org/10.3390/plants12081724
Chicago/Turabian StyleLastochkina, Oksana, Albina Yakupova, Irina Avtushenko, Artem Lastochkin, and Ruslan Yuldashev. 2023. "Effect of Seed Priming with Endophytic Bacillus subtilis on Some Physio-Biochemical Parameters of Two Wheat Varieties Exposed to Drought after Selective Herbicide Application" Plants 12, no. 8: 1724. https://doi.org/10.3390/plants12081724